Embed Size (px)
Citation preview

Hameeda Naimi Barry Hinton: Department of Cell Biology 4 May 2015
Phenotyping Csk knockouts in the epididymis
Introduction: The goal for this semester was to continue analyzing the effects of Csk knockouts on development of the initial segment of the epididymis using data as a starting point for further investigation on this topic. The formal hypothesis investigated concerned whether Csk knockout mice displayed greater cell vascularization and/or cell proliferation through lack of inhibition of SRC family tyrosine-kinases, key initiators of the ERK pro-proliferative signaling pathway. Background: The Wolffian duct or epididymis is a key structure in the male reproductive system as it provides a luminal environment critical in the maturation of spermatozoa. Specifically, experiments blocking luminal factors from entering the epididymis, prevented development of epithelial cells in the initial segment, an area now suggested crucial to male fertility (1). Previous experiments have also shown that luminal fluid factors activate the pro-proliferative and pro-differentiating ERK pathway. A group of cytoplasmic non-receptor Src family tyrosine kinases (SFKs) serve as positive signal intermediaries between luminal fluid factors and the Erk pathway; Src family kinases activity is blocked by C-terminal Src Kinase (Csk), serving a critical tumor suppressor role under normal conditions. The previous semester’s Csk-knockout mouse immunofluorescence preliminary analyses suggested increased vasculogenesis, proposing potential evidence that reduced inhibition of Src family kinases permit greater differentiation through ERK pathway signaling components. This semester’s focus involved proper examination of factors related to the possibility of up-regulated vasculogenesis in the initial segment, an inquiry that might provide a more holistic understanding of epididymal development. Procedures:
• Determining proper analysis of vascularization: Various techniques were researched however three antibodies were ultimately chosen on the basis of their involvement in vascularization. Platelet derived growth factor (PDGF) has been noted in enhancing capillary growth in epididymal tissue (2). Platelet endothelial cell adhesion molecule (PECAM) has been used to evaluate the degree of tumor angiogenesis and finally, Paxillin, a focal adhesion scaffolding protein, has been connected with directing capillary endothelial cell motility (3). See the conclusions section below for further discussion regarding the applicability of the antibodies for future experiments.
• Immunofluorescence: Tissue samples from epididymides dissections, both control and knockout, were fixed in 4% PFA in PBS overnight at 4°C, embedded in paraffin, and sectioned. Slides were then deparaffinated and rehydrated. Antigen unmasking was completed through an antigen unmasking solution (Vector Laboratories) for 10 min on high in a 1,300-W microwave and cooled for 1hr at room temperature (RT). Slides were washed in TBS and then incubated in blocking solution comprised of 10% normal goat serum (Vector Laboratories), 0.5% cold-water fish skin gelatin (Sigma), and TBS for 70 min at RT. Slides were then incubated overnight at 4°C in blocking solution with their respective primary antibodies. The following day, slides were washed in TBS and incubated for 1.5hr at RT with blocking solution and secondary antibodies. Upon subsequent washing in TBS, slides were mounted with Prolong Anti-fade reagent (Molecular Probes) with DAPI for nuclear staining and viewed under a Zeiss microscope. Images can be seen in Figure 2 (A-H) and Figure 3 (A-F).
The following primary antibodies were purchased from Cell Signaling Technology: Phospho-p44/42 MAPK (Erk1/2) (Thr202/Tyr204) antibody (no.4370s, 1:200 working dilution), Phospho-c-Raf (Ser259) antibody (no.9421, 1:50 working dilution), PDGF Receptor β (28E1) antibody (no.3169, 1:100 working dilution), and Phospho-Paxillin (Tyr118) antibody (no.2541, 1:50 working dilution). The following primary antibodies were purchased from Santa Cruz Biotechnology: Phospho-p38 (D-8) antibody (no.sc-7973, 1:100 working dilution) and Pecam-1 (M-20) antibody (no.sc-1506, 1:50 working dilution). The following antibody was purchased from EMD Millipore: Anti-collagen Type IV antibody (no. AB769, 1:50 working dilution). The following secondary antibody was purchased from Life Technologies: Alexa Fluor (no.594, 1:200 working dilution).
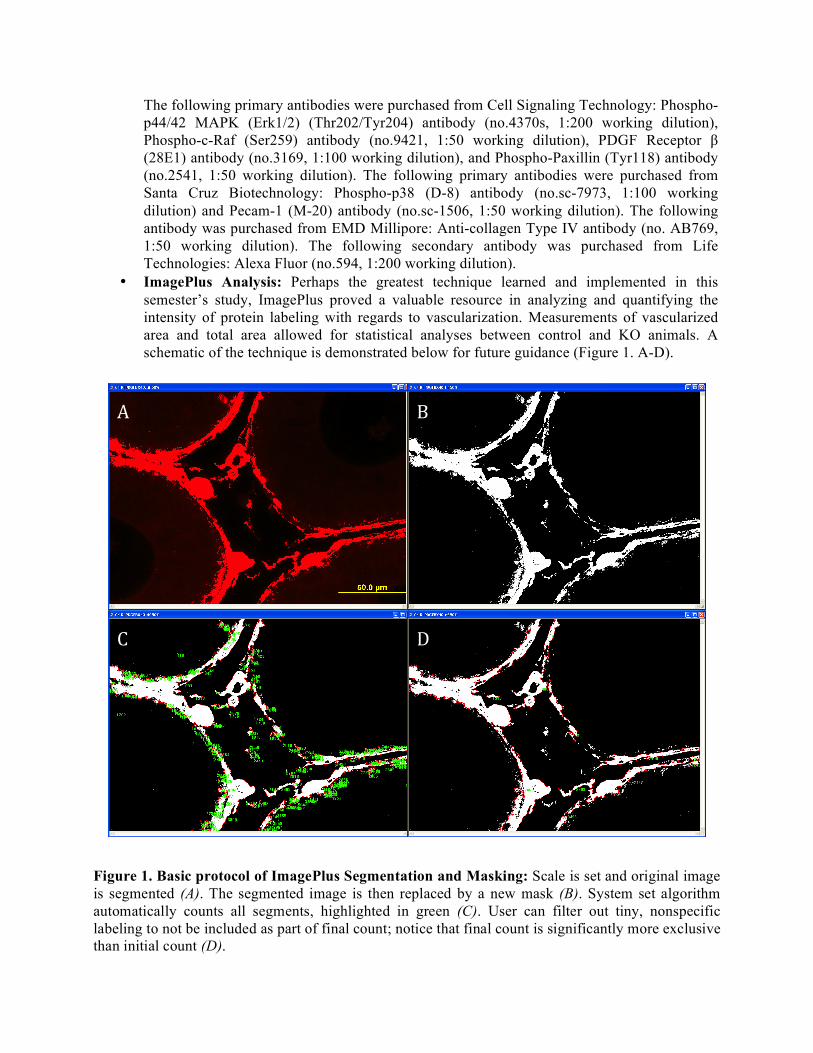
• ImagePlus Analysis: Perhaps the greatest technique learned and implemented in this semester’s study, ImagePlus proved a valuable resource in analyzing and quantifying the intensity of protein labeling with regards to vascularization. Measurements of vascularized area and total area allowed for statistical analyses between control and KO animals. A schematic of the technique is demonstrated below for future guidance (Figure 1. A-D).
Figure 1. Basic protocol of ImagePlus Segmentation and Masking: Scale is set and original image is segmented (A). The segmented image is then replaced by a new mask (B). System set algorithm automatically counts all segments, highlighted in green (C). User can filter out tiny, nonspecific labeling to not be included as part of final count; notice that final count is significantly more exclusive than initial count (D).
A B
C D
Results:
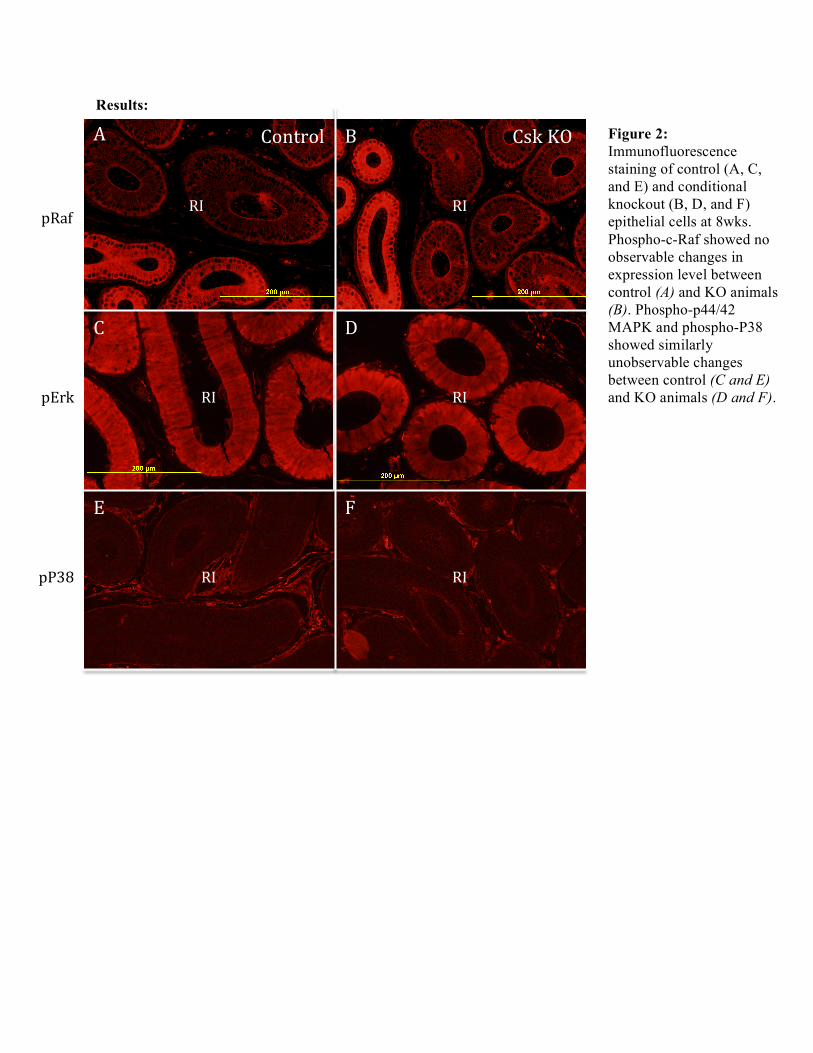
Figure 2: Immunofluorescence staining of control (A, C, and E) and conditional knockout (B, D, and F) epithelial cells at 8wks. Phospho-c-Raf showed no observable changes in expression level between control (A) and KO animals (B). Phospho-p44/42 MAPK and phospho-P38 showed similarly unobservable changes between control (C and E) and KO animals (D and F).
A
Control Csk KO
C
D
B
D
pRaf RI
pErk
RI
RI RI
pP38 RI RI
E F
A Control Csk KO B
C D
E F
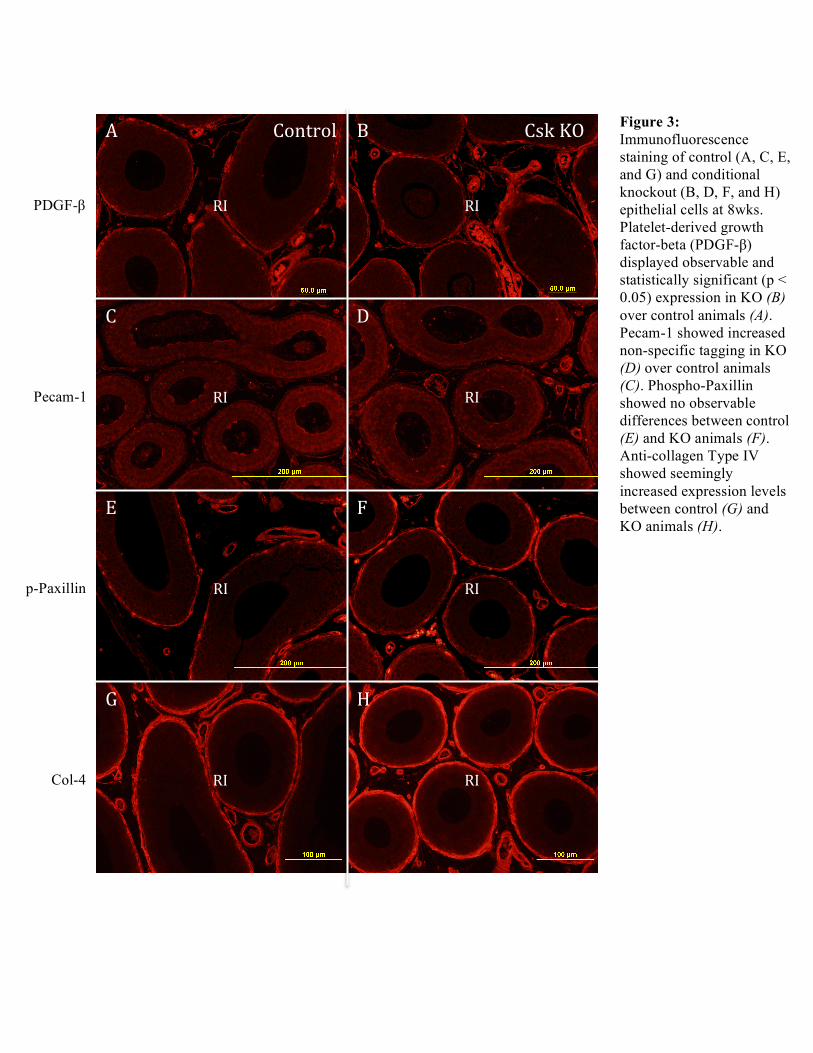
Figure 3: Immunofluorescence staining of control (A, C, E, and G) and conditional knockout (B, D, F, and H) epithelial cells at 8wks. Platelet-derived growth factor-beta (PDGF-β) displayed observable and statistically significant (p < 0.05) expression in KO (B) over control animals (A). Pecam-1 showed increased non-specific tagging in KO (D) over control animals (C). Phospho-Paxillin showed no observable differences between control (E) and KO animals (F). Anti-collagen Type IV showed seemingly increased expression levels between control (G) and KO animals (H).
G H
PDGF-β
Pecam-1
p-Paxillin
Col-4
RI RI
RI RI
RI RI
RI RI

Conclusions/Discussion: Contrary to what was expected, preliminary immunofluorescence analyses did not demonstrate any observable differences in phospho-p44/42 MAPK expression levels between control and KO animals. Similarly unobservable differences were indicated in expression levels for control and knockout animals in both phospho-c-Raf and phospho-P38 analyses. ImagePlus analysis was focused to those antibodies specific to angiogenesis: PDGF-β, Pecam-1, phospho-Paxillin, and Col-4. Scaled measurements of the vasculature area, total area, and a fraction of the two areas (vasculature area/total area) was taken only for PDGF-β at 4 and 8 weeks. Pecam-1 staining did not demonstrate efficient labeling and therefore data collected was temporarily excluded from the current investigation. Furthermore, phospho-Paxillin could not be statistically included as more replicates were necessary for calculations. PDGF-β, while it displayed no significant changes (p > 0.05) for control and KO animals at 4 weeks did exhibit significant changes (p < 0.05) among control and KO animals at 8 weeks. This preliminary investigation suggests a time-sensitive control on angiogenesis although further analyses will have to be well considered before such a conclusion can be made with regards to increased initial segment vascularization. Acknowledgements: A big thank you to Professor Hinton, Bingfang Xu, and Angela Washington for their patience and guidance throughout these past two semesters of research. J

References 1. Xu, Bingfang et al. “Testicular Lumicrine Factors Regulate ERK, STAT, and NFKB Pathways in
the Initial Segment of the Rat Epididymis to Prevent Apoptosis.” Biology of Reproduction 84.6 (2011): 1282–1291. Web.
2. Sato, N. et al. “Platelet-Derived Growth Factor Indirectly Stimulates Angiogenesis in Vitro.” The American Journal of Pathology 142.4 (1993): 1119–1130. Web.
3. Alexandra E. German, Tadanori Mammoto, Elisabeth Jiang, Donald E. Ingber, and Akiko Mammoto. Paxillin controls endothelial cell migration and tumor angiogenesis by altering neuropilin 2 expression. J Cell Sci 2014 127: 1672-1683; Advance Online Article February 12, 2014, doi: 10.1242/jcs.132316. Web.





























