Embed Size (px)
Citation preview

Some observations on the behaviour of the Recent brachiopod MegerZina pisum under laboratory conditions N O R M A N M. S A V A G E
Savage, N. M.: Some observations on the behaviour of the Recent brachiopod Megerlina pisum under laboratory conditions. Lethaia, Vol. 5 , pp. 61-67. Oslo, January 15th, 1972.
Tank experiments with the terebratuloid Megerlina pisum indicate that food is obtained by filtration of sea-water drawn into the shell at the sides and exhaled anteriorly. Under laboratory conditions feeding periods of up to 12 hours were interspersed with resting periods of about equal duration. During the feeding periods the valves were maintained at full gape except for brief interruptions by complete or partial closures. Complete closures lasted several minutes whereas partial closures occurred about half as frequently and lasted only 1 or 2 seconds. Most of these closures did not involve the expulsion of faecal pellets. During the resting periods the valves remained closed for several hours at a time and this condition occasionally continued for 10 hours or more. Apparently a very low respiratory rate operates during these resting periods.
Norman M. Savage, Department of Geology, University of Oregon, Eugene, Oregon 97.103, U.S.A.
Ideally any study of brachiopod activity would take place in the sea and laboratory observations must be considered a poor substitute. However, so little is known about the feeding periodicity and related valve movements of extant brachiopods that even a tank experiment appears to offer instructive results.
The terebratuloids studied belong to the kraussinid species Megerlina pisum (Lamarck) and were collected from a submerged rock at the coast near Tongaat, a small town 35 km northeast of Durban, South Africa. The speci- mens were found in a rocky pool accessible only at low spring tide. Of several dozen boulders examined only one had brachiopods attached. This boulder weighed about 25 km and lay on a sandy rock platform with all the brachiopods and associated organisms on the undersurface. A piece of rock with several specimens attached was removed and the boulder was then carefully replaced. When the locality was revisited 2 weeks later the boulder had been moved several metres by wave action and most of the brachiopods had been stripped off. Several other boulders which had previ- ously been accessible were now buried under beach sand. This brachiopod habitat is clearly a physically perilous one. Even when the rocks are undis- turbed by storms, numerous barnacles, sponges, encrusting algae, alcyonariansr and tunicates compete for living space and there is presumably some dange,
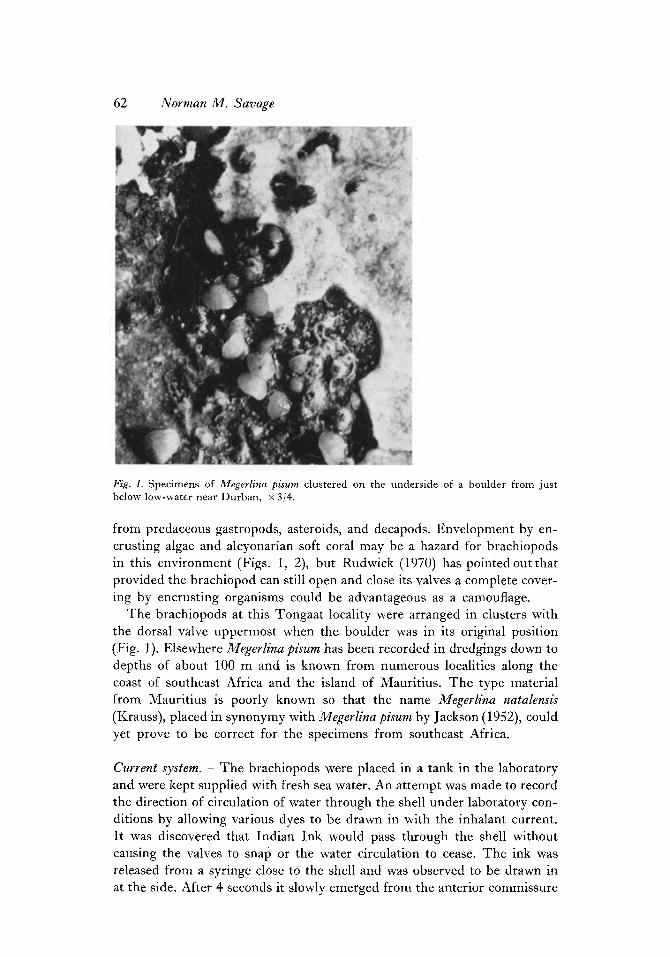
62 Norman M . Savage
Fig. 1. Specimens of Megerlina pisurn clustered on the underside of a boulder from just below low-water near Durban, x 3/4.
from predaceous gastropods, asteroids, and decapods. Envelopment by en- crusting algae and alcyonarian soft coral may be a hazard for brachiopods in this environment (Figs. 1, 2), but Rudwick (1970) has pointedoutthat provided the brachiopod can still open and close its valves a complete cover- ing by encrusting organisms could be advantageous as a camouflage.
The brachiopods at this Tongaat locality were arranged in clusters with the dorsal valve uppermost when the boulder was in its original position (Fig. 1). Elsewhere MegerEina pisum has been recorded in dredgings down to depths of about 100 m and is known from numerous localities along the coast of southeast Africa and the island of Mauritius. The type material from Mauritius is poorly known so that the name Megerlina natalensis (Krauss), placed in synonymy with Megerlina pisum by Jackson (1952), could yet prove to be correct for the specimens from southeast Africa,
Current system, - The brachiopods were placed in a tank in the laboratory and were kept supplied with fresh sea water. An attempt was made to record the direction of circulation of water through the shell under laboratory con- ditions by allowing various dyes to be drawn in with the inhalant current. I t was discovered that Indian Ink would pass through the shell without causing the valves to snap or the water circulation to cease. The ink was released from a syringe close to the shell and was observed to be drawn in at the side. After 4 seconds it slowly emerged from the anterior commissure

Behaeiour of brachio 1
Fig. 2. A cluster of Megerlina pisum partly covered by encroaching algae, l'/?
and was soon dispersed in the water (Fig. 3A-D). The brachiopod continued to feed and the experiment was repeated several times with similar results. It should not be assumed that circulation is normal when particles of ink are being inhaled and exhaled from the system, but it is noteworthy that the ink would not have been drawn in at the sides initially unless an inhalant current existed at that point.
Feeding peri0dicit.y. - Observations of the feeding periodicity and valve snapping of living brachiopods have been made previously, notably by Rud- wick (1961, 1962, 1970), but these were manually recorded and no automati- cally recorded observations over a period of several hours or days are known.
A specimen which had been detached from the rock several weeks earlier and had since been living at the bottom of the tank was transferred to a large 5,000 ml beaker and attached by the ventral valve to a small support using an epoxy resin. A small spot of the adhesive was used to fix a fine chread to the dorsal valve and the thread was connected to a balanced lever which rested against a smoked-drum recorder. The drums supporting the smoked paper were geared to give a record for 50 hours. They could then be moved up or down to give a further record of equal duration. In this way a continuous record was kept for 6 days. Living brachiopods are light sen- sitive and snap closed if illumination is changed. This reaction can occur if the observer passes between the tank and a distant window or quietly
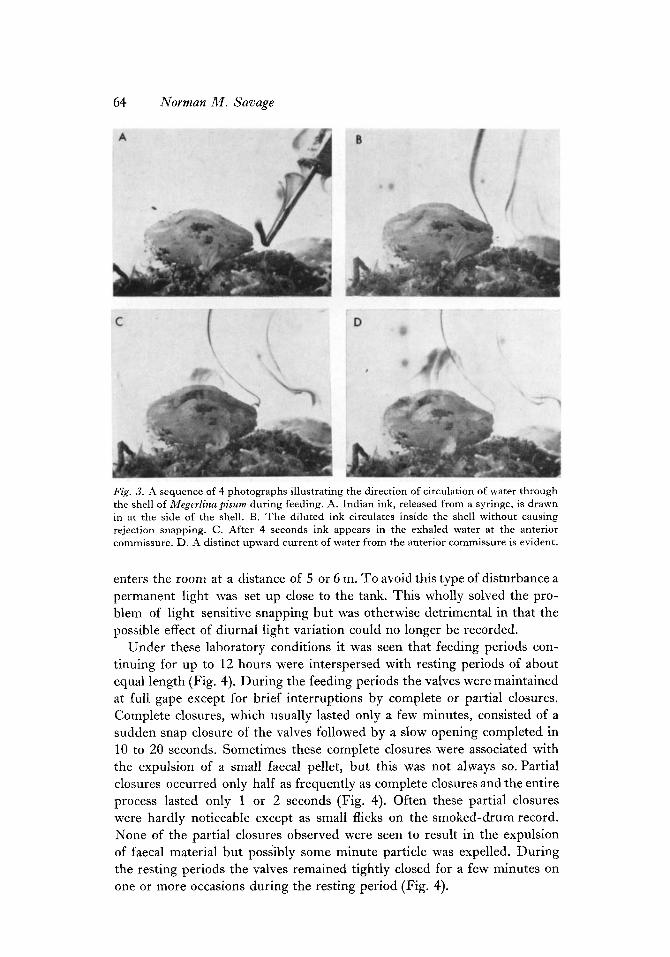
64 Norman M . Savage
Fig. 3. A sequence of 4 photographs illustrating the direction of circulation of water through the shell of Megevlina pisum during feeding. A. Indian ink, released from a syringe, is drawn in at the side of the shell. B. The diluted ink circulates inside the shell without causing rejection snapping. C . After 4 seconds ink appears in the exhaled water at the anterior commissure. D. A distinct upward current of water from the anterior commissure is evident.
enters the room at a distance of 5 or 6 m. To avoid this type of disturbance a permanent light was set up close to the tank. This wholly solved the pro- blem of light sensitive snapping but was otherwise detrimental in that the possible effect of diurnal light variation could no longer be recorded.
Under these laboratory conditions it was seen that feeding periods con- tinuing for up to 12 hours were interspersed with resting periods of about equal length (Fig. 4). During the feeding periods the valves were maintained at full gape except for brief interruptions by complete or partial closures. Complete closures, which usually lasted only a few minutes, consisted of a sudden snap closure of the valves followed by a slow opening completed in 10 to 20 seconds. Sometimes these complete closures were associated with the expulsion of a small faecal pellet, but this was not always so. Partial closures occurred only half as frequently as complete closures and the entire process lasted only 1 or 2 seconds (Fig. 4). Often these partial closures were hardly noticeable except as small flicks on the smoked-drum record. None of the partial closures observed were seen to result in the expulsion of faecal material but possibly some minute particle was expelled. During the resting periods the valves remained tightly closed for a few minutes on one or more occasions during the resting period (Fig. 4).

Behaviour of brarhiopods 65
Fig. .1. A photograph of one of the three smoked-drum records of the feeding periodicity of Megerlina pisum. Feeding periods of up to 12 hours were interspersed with resting periods of about egual length. During feeding periods complete closures lasting several minutes and partial closures lasting 1 or 2 seconds were recorded. During resting periods the valves remai- ned closed for several hours at a time and could remain in this condition for as long as 10 hours.
5 - Lethaia 5 : 1

66 Normun M. Savage
During the resting periods the valves remained tightly closed for several hours at a time. This state could continue for 10 hours or more without interruption, but usually a new feeding period began after 3 or 4 hours or else the valves opened for a few minutes on one or more occasions during the resting period (Fig. 4).
Towards the end of the &day recorded period the water became depleted of micro-organisms and the feeding pattern of the brachiopod gradually changed. The feeding periods became shorter'and the resting periods longer. Eventually the brachiopod remained closed almost continuously and it sur- vived in this way a further 2 weeks, after which the water was changed and the earlier feeding pattern was resumed.
Defaeration. - At the end of the &day recorded experiment the faecal pellets were counted as carefully as was possible on the glass floor of the beaker. There seemed to be only about one-third the number to be expected had each closure been a defaecation action.
Respiration. - The long periods of complete closure show that respiration is not dependent on a constant flow of water through the valves and that a very low respiratory rate operates during the resting periods. Possibly slow gas exchange occurs at the mantle edges, or around the pedicle foramen, but it is more likely that brachiopods are able to survive without oxygen for long periods. In this matter it is instructive to consider the respiratory requirements of certain bivalves. Ricketts & Calvin (1948) have shown that M y a arenaria can survive for 8 days without oxygen and Moore (1931) found that Abra alba survived for 3 days in deoxygenated water. Unlike bi- valves, brachiopods only very rarely occur upwards on the shore into the intertidal zone and it is unlikely that they have developed the same faculty for prolonged anaeobiosis. The example of the bivalves shows that there is nothing extraordinary in the ability of brachiopods to survive for over 12 hours without gaseous exchange.
Genercrl comment. - Epicontinental seas of the Palaeozoic era provided great areas in which the water was rarely more than 200 or 300 ni deep. In these seas brachiopods found quiet water, light, and abundant supplies of micro- scopic food. It is hardly surprising that they became numerous and varied. Today such seas are rare. Most living brachiopods are found along the margins of great oceans where the surf beats endlessly and where competi- tion for space is intense. Despite this changed environment it seems likely that the general behaviour pattern of extant forms is much the same as that of the multitudinous Pa1aeozoi.c forms. Dependence on photic micro- organisms for much of the food intake restricts the great majority of species to the sublittoral zone along with most other benthonic animals. The loph- ophores are wholly food collecting organs and respiration is an incidental,

Behauioiir o j brachiopods 67
non-critical, function which presumably occurs through the general body surface. When feeding, the valves open fully and any intermediate state rarely lasts more than a minute or two. Brachiopods are highly sensitive to slight variations in light intensity and this probably serves as a defence mechanism. Feeding and respiration can cease for many hours at a time and this will occur even when food and fresh water is available. Periodic snapping of the valves is not always a defaecation action and usually results in no faecal pellet. Possibly, minor irritant organisms are expelled in this way.
Acknowledgements. - I t is a pleasure to acknowledge the assistance of Miss Georgina Lambert and Mr. Michael Cooper of the Department of Geology, Durban, who helped locate and collect the specimens, and Dr. Benjamin Raudon of the Department of Animal Biology, Durban, who arranged the loan of the smoked-drum equipment and helped to make it work. Dr. Martin Rudwick and Dr. Anders Martinsson read the manuscript and made valuable suggestions.
R E F E R E N C E S
Jackson, J. W. 1952: A revision of some South African Brachiopoda; with descriptions of new species. Ann. S. Afv. Mus. 31, 1-40.
Moore, H. B. 1931: T h e muds of the Clyde Sea Area. 111. Chemical and physical condi- tions; rate and nature of sedimentation; and fauna. J . M a r . Biol. Ass. U.K. 17, 325-358.
Ricketts, E. F. & Calvin, J. 1948: Between Pacific Tides. 365 pp. Stanford Univ. Press, Stanford.
Rudwick, M. J. S. 1961 : ‘Quick’ and ‘catch’ adductor muscles in brachiopods. Nature 191, 1021.
Rudwick, M. J. S. 1962: Filter-feeding mechanisms in some brachiopods from New Zea- land. J . Linn. SOC. (Zool.) 43, 592-615.
Rudwick, M. J. S. 1970: Litling and Fossil Bvachiopods. 199 pp. Hutchinson & Co., London.





























