Embed Size (px)
Citation preview

Brain Research Bdletin, Vol. 21, pp. 105-I 16. 0 Pergamon Press plc, 1988. Printed in the U.S.A. 0361-9230/88 $3.00 + .OO
Somatostatin Immunoreactive Structures in the Developing Rat Spinal Cord
RAYMOND H. HO’
Department of Anatomy and Neuroscience Research Laboratory The Ohio State University, Columbus, OH 43210
Received 25 September 1987
HO, R. H. Somatostatin immunoreactive structures in the developing rat spinal cord. BRAIN RES BULL 21( 1) 105-l 16, 1988.-Somatostatin immunoreactive (SOM-I) perikarya were first observed in the ventral horn at E12, in the presumptive intermediate gray area at E14, and in the alar plate of the rostral spinal cord at E14. In general, after their initial appearance, their density increased and then decreased during development. A moderate density of SOM-I varicosities became obvious in the superficial laminae of the E20 dorsal horn. By El2 a few SOM-I perikarya, interpreted to be dorsal root ganglia, were observed lateral to the spinal cord, and by E13, SOM immunoreactivity was visualized within the central and peripheral processes of dorsal root ganglion axons. In the marginal zone, SOM-I fibers were first demonstrable in the ventral funiculus at E14, and in the lateral funiculus at E15. After their initial appearance, their density increased and then decreased with age, with the exception of the dorsal part of the lateral funiculus where it increased at the early stages of development to an apparently stable level. The early detection of SOM immunoreactivity in specific spinal regions corresponds well with the birth dates of cells in those regions. This indicates that the SOM-I cells are capable of synthesizing the substance at least as early as they have entered their final cell division.
Rat Spinal cord Somatostatin Development Immunohistochemistry Growth hormone release inhibiting factor Dorsal root ganglia Primary aRerents
SOMATOSTATIN immunoreactive (SOM-I) structures have been described in the spinal cord of a number of species ([13, 25-27, 30, 36, 39, SO] and see citations in [24]) including the human [41]. In the rat, SOM immunoreactivity was first demonstrated in primary afferent neurons implicated in nociception [20]. Subsequently, SOM-I structures were demonstrated in many regions of the spinal cord (see above and [3, 12, 18, 42, 511). SOM-I varicosities are densest in lamina (L) II and X of the spinal gray matter. Although dif- ferent investigators have reported SOM-I cells in different regions of the rat’s spinal cord, taken together, they have been observed in most areas of the gray matter [8, 16,22,24, 39, 421. The distribution of SOM-I structures in the spinal cord allows one to speculate that they may be involved in somatic and autonomic functions as well as somatosensory processing [3, 12, 20, 21, 491.
With respect to sensory processing, experiments using electrophysiological techniques [38], intrathecal administra- tion of SOM combined with behavioral observations [44], intrathecal SOM combined with physiological techniques [52,53], and peptide release in response to specific cutane- ous stimuli [28], have provided accumulating evidence for SOM’s involvement in nociception. Some of the cited studies [28,52, 53]have provided evidence that at least some
SOM-I primary afferents may transmit nociceptive impulses. If this is true, the reported inhibitory [38] role of SOM in the dorsal horn of the spinal cord would likely be mediated by nonprimary afferents which would include SOM-I structures intrinsic to the spinal cord or circuits involving SOM and an inhibitory interneuron.
SOM has been used clinically in humans to alleviate pain. Although the sites and mechanisms of action are unknown, intravenously administered SOM has been shown to relieve the painful sensations associated with cluster headaches in humans [46]. Further, administration of SOM to patients with surgically related pain and terminal cancer produced rapid and profound analgesia [5-7, 311.
Despite the above physiological and clinical studies, the neural circuits responsible for the functions implicated for SOM are unknown. One may study these circuits in the adult animal Alternatively, one may examine them during devel- opment when the connections are less complicated and possibly easier to demonstrate by immunohistochemistry. The latter approach would necessitate an initial devel- opmental study of SOM-I structures as described in the present communication, followed by future developmental studies of their connections.
Although the development of SOM-I structures in the
‘Requests for reprints should be addressed to Dr. Raymond H. Ho, Department of Anatomy, The Ohio State University, 333 West Tenth Avenue, Columbus, OH 43210.
105
HO
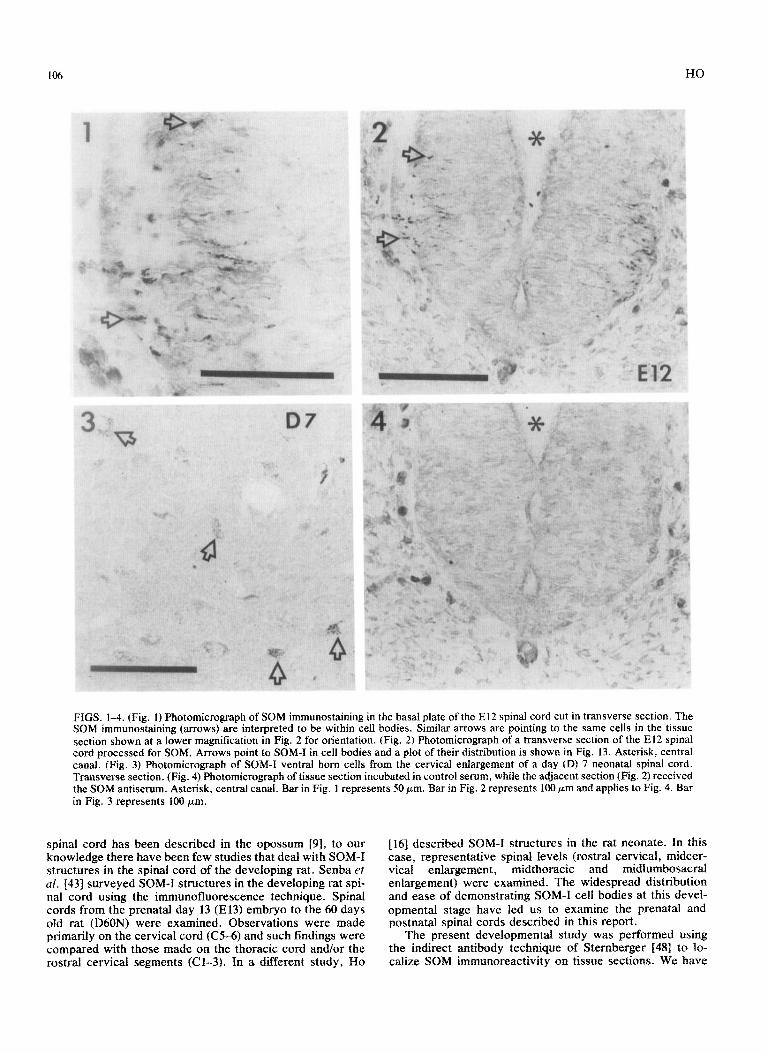
FIGS. 1-4. (Fig. 1) Photomicrograph of SOM immunostaining in the basal plate of the El2 spinal cord cut in transverse section. The SOM immunostaining (arrows) are interpreted to be within cell bodies. Similar arrows are pointing to the same cells in the tissue section shown at a lower magnification in Fig. 2 for orientation. (Fig. 2) Photomicrograph of a transverse section of the El2 spinai cord processed for SOM. Arrows point to SOM-I in cell bodies and a plot of their distribution is shown in Fig. 13. Asterisk, central canal. (Fig. 3) Photomicro~aph of SOM-I ventral horn cells from the cervical enlargement of a day (D) 7 neonatal spinal cord. Transverse section. (Fig. 4) Photomicrograph of tissue section incubated in control serum, while the adjacent section (Fig. 2) received the SOM antiserum. Asterisk, central canal. Bar in Fig. 1 represents 50 pm. Bar in Fig. 2 represents 100 ,xm and applies to Fig. 4. Bar in Fig. 3 represents 100 pm.
spinal cord has been described in the opossum [9], to our knowledge there have been few studies that deal with SOM-I structures in the spinal cord of the developing rat. Senba et ul. [43] surveyed SOM-I structures in the developing rat spi- nal cord using the immunofluorescence technique. Spinal cords from the prenatal day 13 (E13) embryo to the 60 days old rat (D60N) were examined. Observations were made primarily on the cervical cord (CS-6) and such findings were compared with those made on the thoracic cord and/or the rostra1 cervical segments (Cl-3). In a different study, Ho
1161 described SOM-I structures in the rat neonate. In this case, representative spinal levels frostral cervicai, midcer- vital enlargement, midthoracic and mi~um~sacral enlargement) were examined. The widespread distribution and ease of demonstrating SOM-I cell bodies at this devel- opmental stage have led us to examine the prenatal and postnatal spinal cords described in this report.
The present developmental study was performed using the indirect antibody technique of Sternberger [48] to lo- calize SOM immunoreactivity on tissue sections. We have
ONTOGENY OF SOMATOSTATIN SPINAL SYSTEMS 107
b
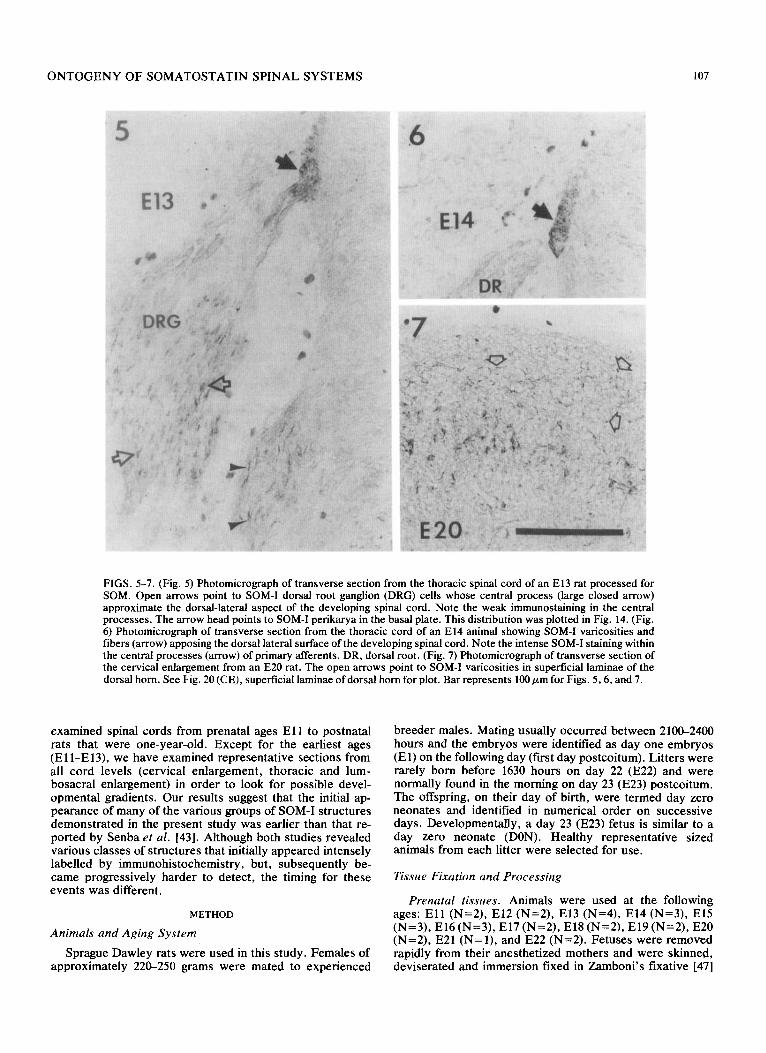
FIGS. 5-7. (Fig. 5) Photomicrograph of transverse section from the thoracic spinal cord of an El3 rat processed for SOM. Open arrows point to SOM-I dorsal root ganglion (DRG) cells whose central process (large closed arrow) approximate the dorsal-lateral aspect of the developing spinal cord. Note the weak immunostaining in the central processes. The arrow head points to SOM-I perikarya in the basal plate. This distribution was plotted in Fig. 14. (Fig. 6) Photomicrograph of transverse section from the thoracic cord of an El4 animal showing SOM-I varicosities and fibers (arrow) apposing the dorsal lateral surface of the developing spinal cord. Note the intense SOM-I staining within the central processes (arrow) of primary afferents. DR, dorsal root. (Fig. 7) Photomicrograph of transverse section of the cervical enlargement from an E20 rat. The open arrows point to SOM-I varicosities in superficial laminae of the dorsal horn. See Fig. 20 (CE), superficial laminae of dorsal horn for plot. Bar represents 100 pm for Figs. 5,6, and 7.
examined spinal cords from prenatal ages El1 to postnatal rats that were one-year-old. Except for the earliest ages (Eli-E13), we have examined representative sections from all cord levels (cervical enlargement, thoracic and lum- bosacral enlargement) in order to look for possible devel- opmental gradients. Our results suggest that the initial ap- pearance of many of the various groups of SOM-I structures demonstrated in the present study was earlier than that re- ported by Senba et al. [43]. Although both studies revealed various classes of structures that initially appeared intensely labelled by immunohistochemistry, but, subsequently be- came progressively harder to detect, the timing for these events was different.
METHOD
Animals and Aging System
Sprague Dawley rats were used in this study. Females of approximately 220-250 grams were mated to experienced
breeder males. Mating usually occurred between 210&2400 hours and the embryos were identified as day one embryos (El) on the following day (first day postcoitum). Litters were rarely born before 1630 hours on day 22 (E22) and were normally found in the morning on day 23 (E23) postcoitum. The offspring, on their day of birth, were termed day zero neonates and identified in numerical order on successive days. Developmentally, a day 23 (E23) fetus is similar to a day zero neonate (DON). Healthy representative sized animals from each litter were selected for use.
Tissue Fixation and Processing
Prenatal tissues. Animals were used at the following ages: El1 (N=2), El2 (N=2), El3 (N=4), El4 (N=3), El5 (N=3), E16 (N=3), El7 (N=2), El8 (N=2), El9 (N=2), E20 (N=2), E21 (N=l), and E22 (N=2). Fetuses were removed rapidly from their anesthetized mothers and were skinned, deviserated and immersion fixed in Zamboni’s fixative [47]
8
HO
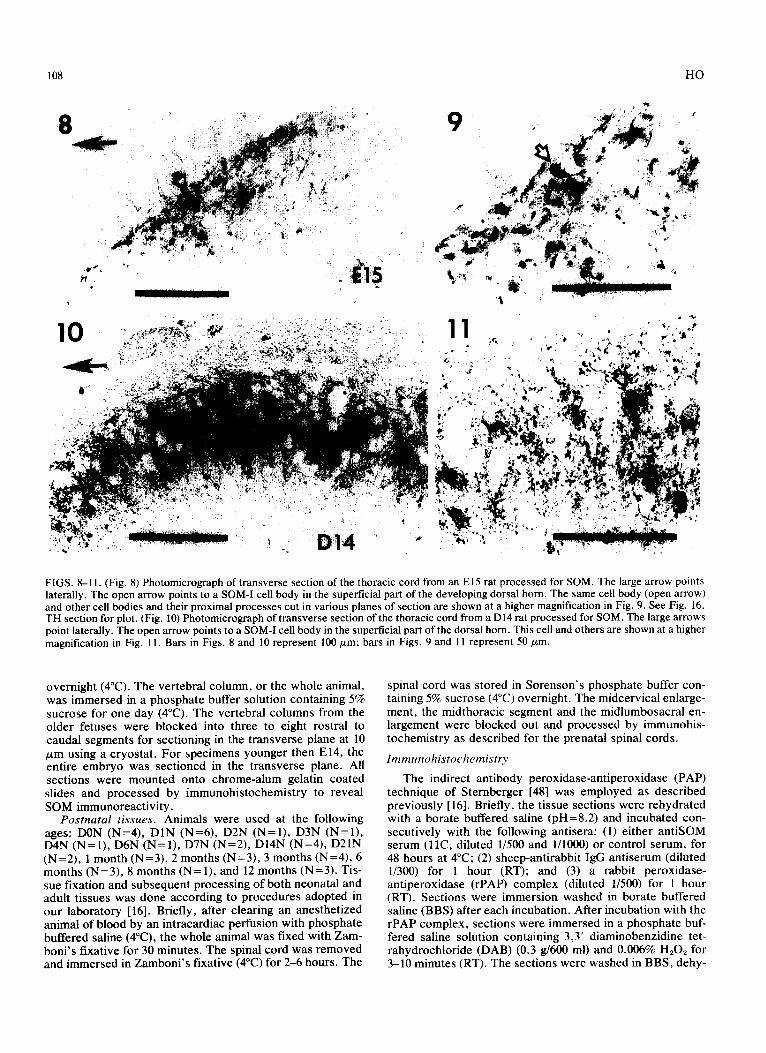
FIGS. 8-I 1. (Fig. 8) Photomicrograph of transverse section of the thoracic cord from an El5 rat processed for SOM. The large arrow points laterally. The open arrow points to a SOM-I cell body in the superficial part of the developing dorsal horn. The same cell body (open arrow) and other cell bbdies and their proximal processes cut in various planes of section are shown at a higher magnification in Fig. 9. See Fig. 16, TH section for plot. (Fig. 10) Photomicrograph of transverse section of the thoracic cord from a D14 rat processed for SOM. The large arrows point laterally. The ape, arrow points to a SOM-I cell body in the superficial part of the dorsal horn. This cell and others are shown at a higher magnification in Fig. 11. Bars in Figs. 8 and 10 represent 100 pm; bars in Figs. 9 and 11 represent 50 pm.
overnight (4°C). The vertebral column, or the whole animal, was immersed in a phosphate buffer solution containing 5% sucrose for one day (4°C). The vertebral columns from the older fetuses were blocked into three to eight rostra1 to caudal segments for sectioning in the transverse plane at 10 pm using a cryostat. For specimens younger then E14, the entire embryo was sectioned in the transverse plane. All sections were mounted onto chrome-alum gelatin coated slides and processed by immunohistochemistry to reveal SOM immunoreactivity.
Postnatal tissues. Animals were used at the following ages: DON (N=4), DlN (N=6), D2N (N=l), D3N (N=l), D4N (N= l), D6N (N=l), D7N (N=2), D14N (N=4), D21N (N=2), 1 month (N=3), 2 months (N=3), 3 months (N=4), 6 months (N=3), 8 months (N=l), and 12 months (N=3). Tis- sue fixation and subsequent processing of both neonatal and adult tissues was done according to procedures adopted in our laboratory [16]. Briefly, after clearing an anesthetized animal of blood by an intracardiac perfusion with phosphate buffered saline (4”C), the whole animal was fixed with Zam- boni’s fixative for 30 minutes. The spinal cord was removed and immersed in Zamboni’s fixative (4°C) for 2-6 hours. The
spinal cord was stored in Sorenson’s phosphate buffer con- taining 5% sucrose (4°C) overnight. The midcervical enlarge- ment, the midthoracic segment and the midlumbosacral en- largement were blocked out and processed by immunohis- tochemistry as described for the prenatal spinal cords.
Immunohistochemistry
The indirect antibody peroxidase-antiperoxidase (PAP) technique of Stemberger [48] was employed as described previously [16]. Briefly, the tissue sections were rehydrated with a borate buffered saline (pH=8.2) and incubated con- secutively with the following antisera: (1) either antiSOM serum (llC, diluted l/500 and l/1000) or control serum, for 48 hours at 4°C; (2) sheep-antirabbit IgG antiserum (diluted l/300) for 1 hour (RT); and (3) a rabbit peroxidase- antiperoxidase (rPAP) complex (diluted l/500) for 1 hour (RT). Sections were immersion washed in borate buffered saline (BBS) after each incubation. After incubation with the rPAP complex, sections were immersed in a phosphate buf- fered saline solution containing 3,3’ diaminobenzidine tet- rahydrochloride (DAB) (0.3 g/600 ml) and 0.006% H,Oz for 3-10 minutes (RT). The sections were washed in BBS, dehy-
ONTOGENY OF SOMATOSTATIN SPINAL SYSTEMS 109
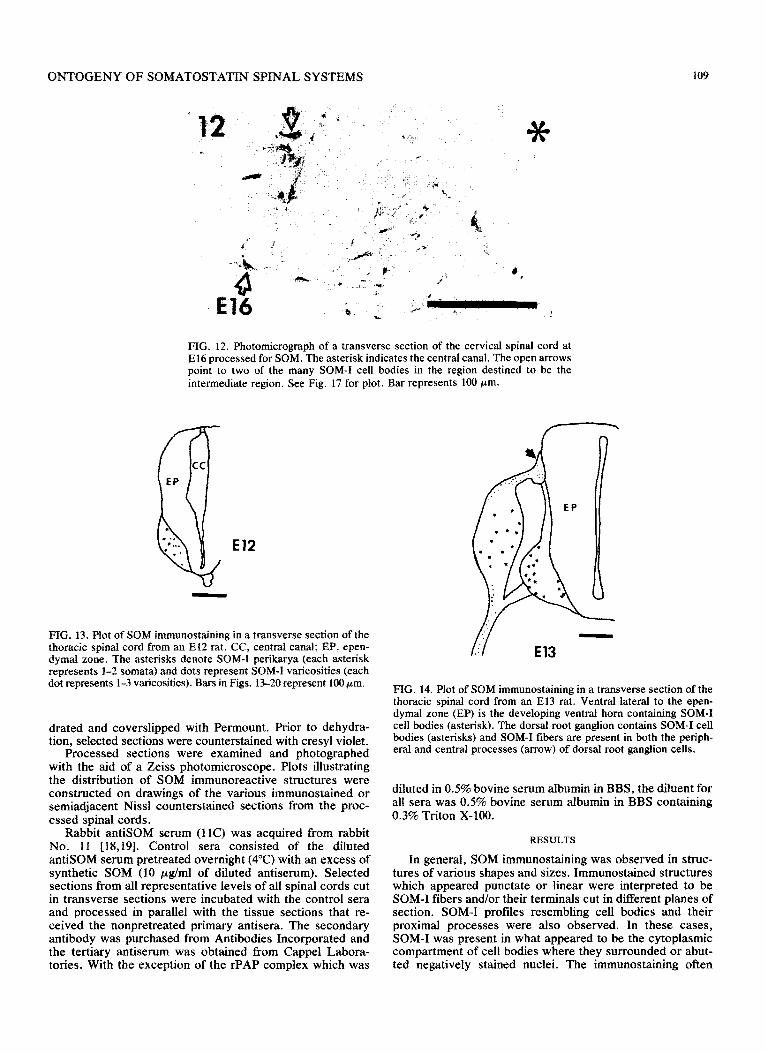
FIG. 12. Photomicrograph of a transverse section of the cervical spinal cord at El6 processed for SOM. The asterisk indicates the central canal. The open arrows point to two of the many SOM-I cell bodies in the region destined to be the intermediate region. See Fig. 17 for plot. Bar represents 100 pm.
El2
FIG. 13. Plot of SOM immunost~ning in a transverse section of the thoracic spinal cord from an El2 rat. CC, central canal; EP, epen- dymal zone. The asterisks denote SOM-I perikarya (each asterisk represents l-2 somata) and dots represent SOM-I varicosities (each dot represents l-3 varicosities). Bars in Figs. 13-20 represent 100 pm.
drated and coverslipped with Permount. Prior to dehydra- tion, selected sections were counterstained with cresyl violet.
Processed sections were examined and photographed with the aid of a Zeiss photomicroscope. Plots illustrating the distribution of SOM immunoreactive strnctures were constructed on drawings of the various immunostained or semiadjacent Nissl counterstained sections from the proc- essed spinal cords.
Rabbit antiSOM serum (IlC) was acquired from rabbit No. 11 [18,19]. Control sera consisted of the diluted antiSOM serum pretreated overnight (4°C) with an excess of synthetic SOM (10 &ml of diluted antiserum). Selected sections from all representative levels of all spinal cords cut in transverse sections were incubated with the control sera and processed in parallel with the tissue sections that re- ceived the nonpretreated primary antisera. The secondary antibody was purchased from Antibodies Incorporated and the tertiary antiserum was obtained from Cappel Labora- tories. With the exception of the rPAP complex which was
FIG. 14. Plot of SOM immunostaining in a transverse section of the thoracic spinal cord from an El3 rat. Ventral lateral to the epen- dymal zone (EP) is the developing ventral horn containing SOM-I cell bodies (asterisk). The dorsal root ganglion contains SOM-I cell bodies (asterisks) and SOM-I fibers are present in both the periph- eral and central processes (arrow) of dorsal root ganglion cells.
diluted in 0.5% bovine serum albumin in BBS, the diluent for all sera was 0.5% bovine serum albumin in BBS containing 0.3% Triton X-100.
RESULTS
In general, SOM immunostaining was observed in struc- tures of various shapes and sizes. Immunostained structures which appeared punctate or linear were interpreted to be SOM-I fibers and/or their terminals cut in different planes of section. SON-I profiles resembling cell bodies and their proximal processes were also observed. In these cases, SOM-I was present in what appeared to be the cytoplasmic compartment of cell bodies where they surrounded or abut- ted negatively stained nuclei. The immunostaining often
HO
CE
TH
LS
El4
-
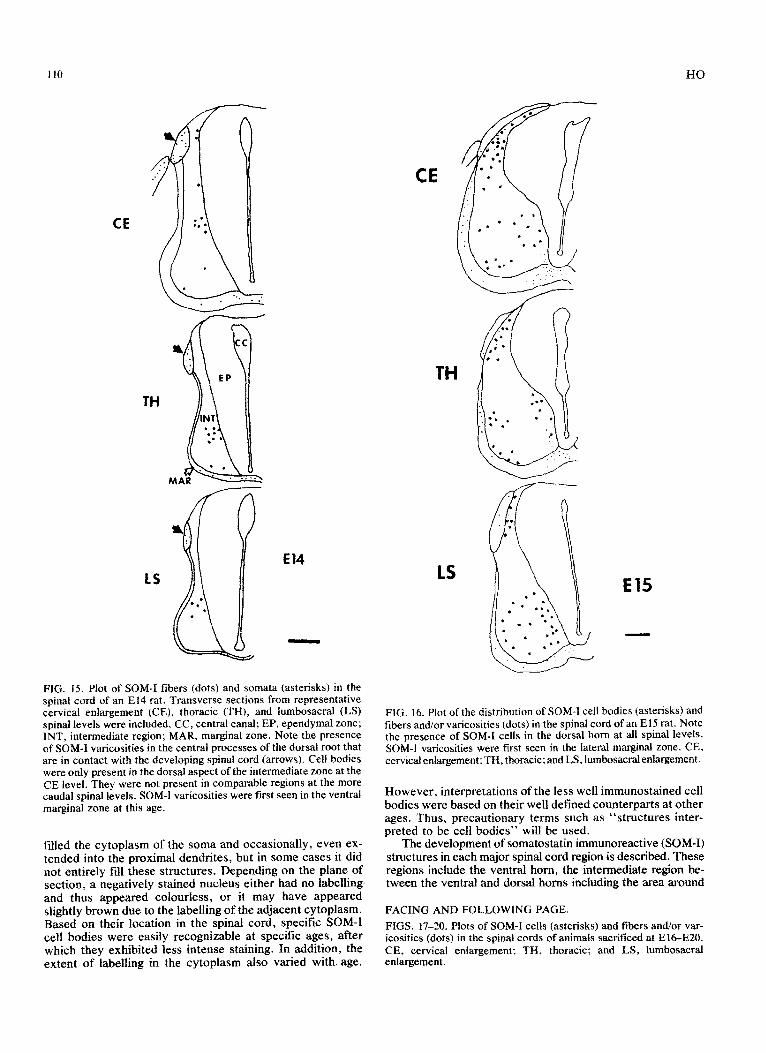
FIG. IS. Plot of SOM-I fibers (dots) and somata (asterisks) in the spinal cord of an El4 rat. Transverse sections from representative cervical enlargement (CE), thoracic (TH), and lumbosacral (LS) spinal levels were included. CC, central canal; EP, ependymal zone; INT, intermediate region; MAR, marginal zone. Note the presence of SOM-I varicosities in the central processes of the dorsal root that are in contact with the developing spinal cord (arrows). Cell bodies were only present in the dorsal aspect of the intermediate zone at the CE level. They were not present in comparable regions at the more caudal spinal levels. SOM-I varicosities were first seen in the ventral marginal zone at this age.
filled the cytoplasm of the soma and occasionally, even ex- tended into the proximal dendrites, but in some cases it did not entirely fill these structures. Depending on the plane of section, a negatively stained nucleus either had no labelling. and thus appeared colourless, or it may have appeared slightly brown due to the labelling of the adjacent cytoplasm. Based on their location in the spinal cord, specific SOM-I cell bodies were easily recognizable at specific ages, after which they exhibited less intense staining. In addition, the extent of labelling in the cytoplasm also varied with age.
El5
FIG. 16. Plot of the distribution of SOM-I cell bodies (asterisks) and fibers and/or varicosities (dots) in the spinal cord of an El5 rat. Note the presence of SOM-I cells in the dorsal horn at ail spinal levels. SOM-I varicosities were first seen in the lateral marginal zone. CE, ce~ic~enlargement: TH, thoracic; and LS, ium~sac~enla~ement.
However, interpretations of the less well immunostained cell bodies were based on their well defined counterparts at other ages. Thus, precautiona~ terms such as “structures inter- preted to be cell bodies” will be used.
The development of somatostatin immunoreactive (SOM-I) structures in each major spinal cord region is described. These regions include the ventral horn, the intermediate region be- tween the ventral and dorsal horns including the area around
FACING AND FOLLOWING PAGE,
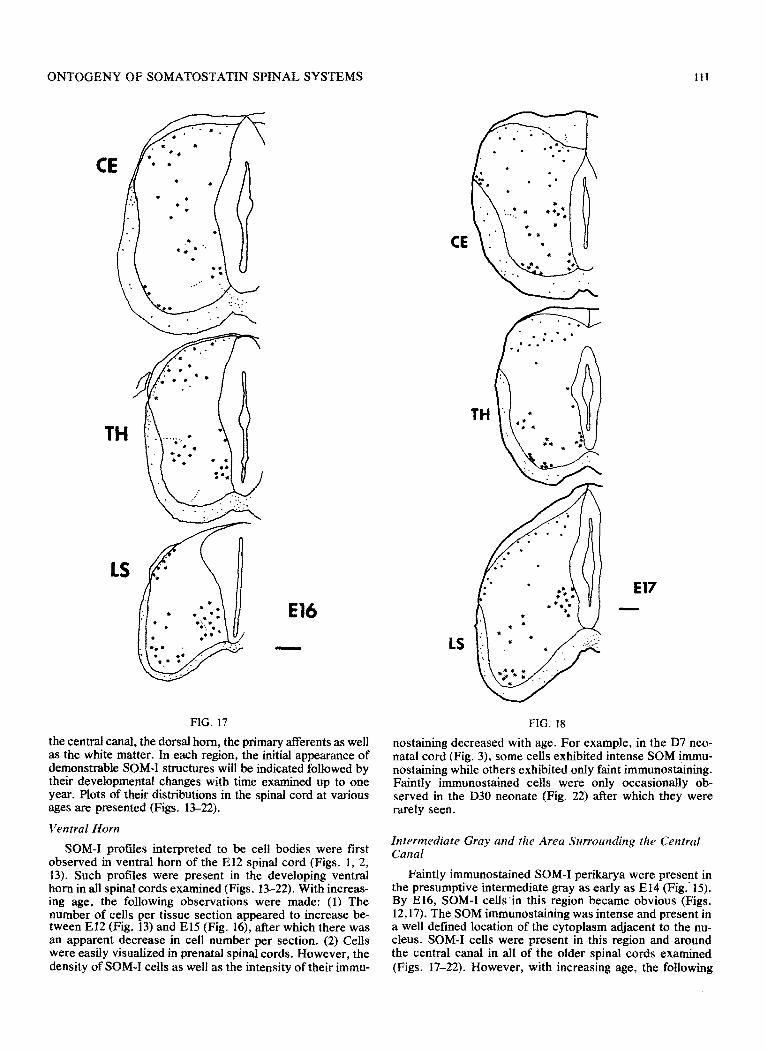
FIGS. 17-20. Plots of SOM-I cells (asterisks) and fibers and/or var- icosities (dots) in the spinal cords of animals sacrificed at E&E20. CE, cervical enlargement; TH, thoracic; and LS, lumbosacral enlargement.
ONTOGENY OF SOMATOSTATIN SPINAL SYSTEMS Ill
FIG. 17
El6
the central canal, the dorsal horn, the primary afferents as well as the white matter. In each region, the initial appearance of demonstrable SOM-I structures will be indicated followed by their developmental changes with time examined up to one year. Plots of their distributions in the spinal cord at various ages are presented (Figs. 13-22).
Ventral Horn
SOM-I profiles interpreted to be cell bodies were first observed in ventral horn of the El2 spinal cord (Figs. 1, 2, 13). Such profiles were present in the developing ventral horn in all spinal cords examined (Figs. 13-22). With increas- ing age, the folIowing observations were made: (1) The number of cells per tissue section appeared to increase be- tween El2 (Fig. 13) and El5 (Fig. 16), after which there was an apparent decrease in cell number per section. (2) Cells were easily visualized in prenatal spinal cords. However, the density of SOM-I ceils as well as the intensity of their immu-
FIG. 18
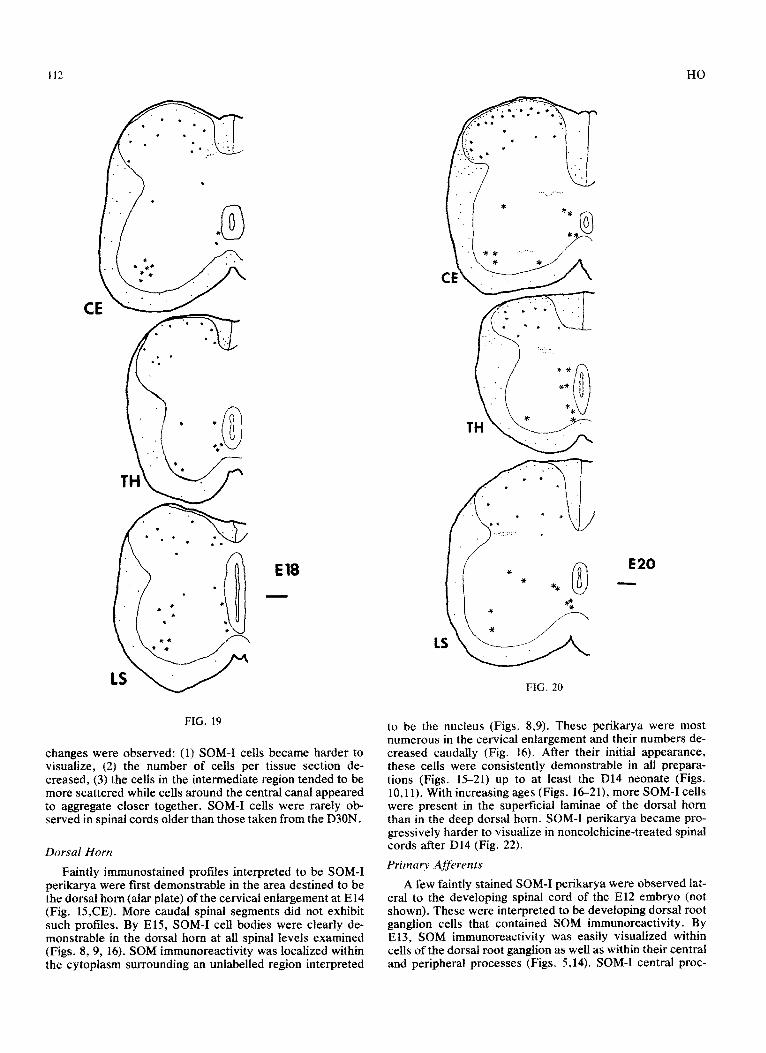
El7 -
nostaining decreased with age. For example, in the D7 neo- natal cord (Fig. 3), some cells exhibited intense SOM immu- nostaining while others exhibited only faint immunostaining. Faintly immunostained ceils were only occasionally ob- served in the D30 neonate (Fig. 22) after which they were rarely seen.
~~t~rmedi~te Gray and the Area ~~rr~und~~~ the Central Canal
Faintly immunostained SOM-I perikarya were present in the presumptive intermediate gray as early as El4 (Fig.. 15). By E16, SOM-I cells in this region became obvious (Figs. 12,17). The SGM immunostaining was intense and present in a well defined location of the cytoplasm adjacent to the nu- cleus. SOM-I cells were present in this region and around the central canal in all of the older spinal cords examined (Figs. 17-22). However, with increasing age, the following
HO
FIG. 19
changes were observed: (1) SOM-I cells became harder to visualize, (2) the number of cells per tissue section de- creased, (3) the cells in the intermediate region tended to be more scattered while cells around the central canal appeared to aggregate closer together. SOM-I cells were rarely ob- served in spinal cords older than those taken from the D30N.
Dorsal Horn
Faintly immunostained profiles interpreted to be SOM-I perikarya were first demonstrable in the area destined to be the dorsal horn (alar plate) of the cervical enlargement at El4 (Fig. 15,CE). More caudal spinal segments did not exhibit such profiles. By E15, SOM-I cell bodies were clearly de- monstrable in the dorsal horn at all spinal levels examined (Figs. 89, 16). SOM immunoreactivity was localized within the cytoplasm surrounding an unlabelled region interpreted
-
FIG. 20
E20
to be the nucleus (Figs. 8,9). These perikarya were most numerous in the cervical enlargement and their numbers de- creased caudally (Fig. 16). After their initial appearance, these cells were consistently demonstrable in all prepara- tions (Figs. 15-21) up to at least the D14 neonate (Figs. 10,ll). With increasing ages (Figs. 1621), more SOM-I cells were present in the superficial laminae of the dorsal horn than in the deep dorsal horn. SOM-I perikarva became uro- gressively harder to visualize in noncolchicink-treated sp’inal cords after D14 (Fig. 22).
Primnry Afferents
A few faintly stained SOM-I perikarya were observed lat- eral to the developing spinal cord of the El2 embryo (not shown). These were interpreted to be developing dorsal root ganglion cells that contained SOM immunoreactivity. By E13, SOM immunoreactivity was easily visualized within cells of the dorsal root ganglion as well as within their central and peripheral processes (Figs. 5,14). SOM-I central proc-
ONTOGENY OF SOMATOSTATIN SPINAL SYSTEMS 113
07
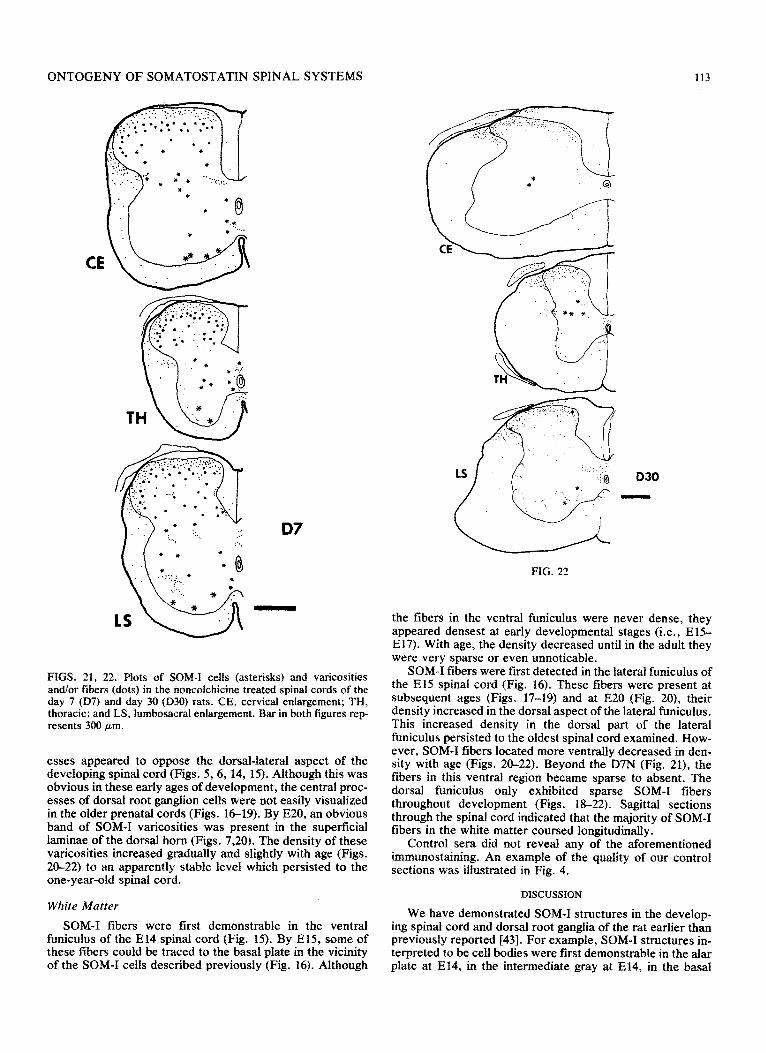
FIGS. 21, 22. Plots of SOM-I cells (asterisks) and varicosities and/or fibers (dots) in the noncolchicine treated spinal cords of the day 7 (D7) and day 30 (D30) rats. CE, cervical enlargement; TH, thoracic; and LS, lumbosacral enlargement. Bar in both figures rep- resents 300 pm.
esses appeared to oppose the dorsal-lateral aspect of the developing spinal cord (Figs. 5,6, 14, 15). Although this was obvious in these early ages of development, the central proc- esses of dorsal root ganglion cells were not easily visualized in the older prenatal cords (Figs. 1619). By E20, an obvious band of SOM-I varicosities was present in the superficial laminae of the dorsal horn (Figs. 7,20). The density of these varicosities increased gradually and slightly with age (Figs. 20-22) to an apparently stable level which persisted to the one-year-old spinal cord.
White Matter
SOM-I fibers were first demonstrable in the ventral funiculus of the El4 spinal cord (Fig. 15). By E15, some of these fibers could be traced to the basal plate in the vicinity of the SOM-I cells described previously (Fig. 16). Although
030
-
FIG. 22
the fibers in the ventral funiculus were never dense, they appeared densest at early developmental stages (i.e., El% E17). With age, the density decreased until in the adult they were very sparse or even unnoticable.
SOM-I fibers were first detected in the lateral fnniculus of the El5 spinal cord (Fig. 16). These fibers were present at subsequent ages (Figs. 17-19) and at EC20 (Fig. 20), their density increased in the dorsal aspect of the lateral funiculus. This increased density in the dorsal part of the lateral funiculus persisted to the oldest spinal cord examined. How- ever, SOM-I fibers located more ventrally decreased in den- sity with age (Figs. 20-22). Beyond the D7N (Fig. 21), the fibers in this ventral region became sparse to absent. The dorsal funiculus only exhibited sparse SOM-I fibers throughout development (Figs. ll3-22). Sagittal sections through the spinal cord indicated that the majority of SOM-I fibers in the white matter coursed longitudinally.
Control sera did not reveal any of the aforementioned immunostaining. An example of the quality of our control sections was illustrated in Fig. 4.
DISCUSSION
We have demonstrated SOM-I structures in the develop- ing spinal cord and dorsal root ganglia of the rat earlier than previously reported [43]. For example, SOM-I structures in- terpreted to be cell bodies were first demonstrable in the alar plate at E14, in the intermediate gray at E14, in the basal

114 HO
plate at E12, and in the dorsal root ganglia and their associ- ated central and peripheral processes at E13. The ability to demonstrate these structures at such early ages allowed us to make potential new correlations with important early devel- opmental events in the rat spinal cord. The initial timing for the demonstration of SOM-I cells in various regions of the developing spinal cord corresponded well with birth dates of cells in those regions [ 1, 29, 34, 351. Thus, we may examine these cells by using immunohistochemistry at least as early as they have entered their final cell division.
Similar to other reports using young animals [9,37] col- chicine treatment was not needed to demonstrate SOM-I cell bodies. It should be emphasized that this study was based on animals that were not treated with colchicine. Thus, the densities and distributions of SOM-I cell bodies in our spinal cords may be different from those seen in colchicine-treated animals of corresponding ages. We hypothesize that differ- ences in SOM immunostaining of cell bodies with and with- out colchicine are most obvious in older animals and that they decrease progressively with decreasing age. Although this hypothesis remains to be tested, the results of the pres- ent study will serve as an important baseline for the evalua- tion of colchicine-treated cords as well as for experimental studies in which normal untreated spinal cord preparations are needed. Further, since colchicine inhibits axonal trans- port, the SOM-I varicosities interpreted to be fibers and/or their terminals demonstrated in this study may be more numerous than after colchicine treatment.
SOM immunoreactivity was also present very early in the development of the spinal cord and/or dorsal root ganglia of other species. SOM immunoreactivity was first demon- strated in the dorsal root ganglion of the quail embryo at day 13 [ll]. SOM immunoreactivity was first detected in the opossum at postnatal day 5 [9]. SOM-I fibers were relatively numerous within the marginal zone and present within presumptive laminae I-III as well as in more ventral areas of the gray matter. It should be noted, however, that in the rat, SOM immunoreactivity was demonstrable earlier than other peptides such as substance-P and enkephalin [37,43]. neurotensin [43], cholecystokinin-8 and vasoactive intestinal peptide [lo]. In the opossum, the earliest demonstration of SOM immunoreactivity lagged behind that of substance-P and enkephalin structures [9]. More studies are needed, how- ever, before one can speculate on the significance, if any, of these developmental sequences. It should be emphasized, however, that SOM and other peptides could be synthesized by spinal neurons very early in development. Whether the ability of spinal neurons to synthesize these peptides paral- lels their ability to release them to carry out the functions which characterize them in the adult animal remains to be determined.
It should be emphasized that the neuronal elements dem- onstrating SOM immunoreactivity may have been present before sufftcient concentrations of SOM immunoreactivity were available to be demonstrable. We interpret that SOM immunoreactivity was present within the structures de- scribed at least as early as it could be detected by im- munohistochemistry, but it was possible that it was present in very small quantities even earlier. Further, it should be emphasized that at the light microscopic level cell or nuclear membranes were not visualized. Profiles that resembled SOM-I cell bodies were interpreted to be SOM-I perikarya, but it was possible that some of them were SOM-I var- icosities or terminals closely apposing SOM negative somata. Support for our assumption was obtained from other studies in the adult and neonatal rat in which similar profiles in corresponding spinal regions have been interpreted to be cell bodies [8, 16, 22, 24, 39, 421. It was also possible that potential SOM-I varicosities or terminals apposing a SOM-I cell body may escape detection. Electron microscopic studies will be helpful to clarify these issues.
The function(s) of SOM in the adult rat and other species remain(s) speculative. but there is sufficient evidence to suggest that SOM is involved in nociceptive mechanisms. Thus, it is likely that at least one of the cell types described may be involved in nociceptive circuitry. Physiological studies [2. 4. 32, 33, 54, 551 have implicated LI, outer LII, LV and LX in nociception and the SOM-I structures de- scribed in this report within these regions may be involved. However. cells in other regions of the spinal cord may send processes to these ‘nociceptive implicated’ regions. In addi- tion, descending SOM-I systems exist [23,40,45] which may play a role in nociception. Thus, the spinal circuitry involv- ing SOM structures is complicated. Whatever the function(s) of SOM systems in the adult animal, it is premature to as- sume that they are the same in the developing animal.
Although the exact mechanism is unknown, synthetic SOM has been shown to promote outgrowth of axotomized neurites isolated from the pond snail [14]. Possible mech- anisms by which SOM may act in this capacity have been discussed [14]. Thus, in the spinal cord of the developing rat, the possibility that SOM may play a role in the initial events of axonal production and/or elongation cannot be excluded.
ACKNOWLEDGEMENTS
Because we could label SOM-I structures at very young ages, it should be possible to carry out more detailed studies of their development. For example, electron microscopic studies of synaptogenesis of SOM-I and related structures may be undertaken. Based on these studies of the normal relationship(s) between SOM-I and other structures and their development, experimental studies of developmental plas- ticity may follow.
This work was supported by NIH grants NS17080, NS10165 and NS231.59. I thank Dr. George F. Martin for his helpful sugges- tions in the preparation of this manuscript. The author is grateful for the technical assistance of Ranee Wolf and Marquita Allen and the photographic assistance of Mr. Karl Rubin. I thank Mr. Steve R. Zimmerman for his generous help with the word processor system. The author thanks Mary Braidich and Susan Fairman for the tvpina of the manuscript. Preliminary observations from this study werebre- sented at the Neuroscience Annual Meeting in 1982 [ 151 and 1983 [ 17.
REFERENCES
1. Altman, J.: Bayer, S. A. The development of the rat spinal cord. In: Beck, F.; Hild, W.: van Limborgh, J.; Ortmann, R.; Pauly, J. E.; Schiebler, T. H., eds. Advances in anatomy, embryology and cell biology. vol. 85. Berlin: Springer-Verlag: 1984.
2. Basbaum, A. I.; Fields, H. L. Endogenous pain control sys- terns: Brainstem spinal pathways and endorphin cicuitry. Annu. Rev. Neurosci. 7:30%338; 1984.

ONTOGENY OF SOMATOSTATIN SPINAL SYSTEMS 115
3.
4.
5.
6.
7.
8.
9.
10.
Il.
12.
Buck, S. H.; Walsh, J. H.; Yamamura, H. 1.; Burks, T. F. Mini review: neuropeptides in sensory neurons. Life Sci. 30:1857- 1866; 1982. Cervero, F.: Iggo, A. The substantia gelatinosa of the spinal cord. A critical review. Brain 103:717-772; 1980. Chrubasik, J.: Meynadier, J.; Blond, S.; Scherpereel, P.; Ack- erman, E.; Weinstock, M.; Bonath, K. ; Cramer, H.; Wunsch, E. Somatostatin, a potent analgesic. Lancet 2:1208-1209; 1984. Chrubasik, J.; Meynadier, J.; Scherpereel, P. Somatostatin ver- sus morphine in epidura1 treatment after major a~omin~ op- erations. Anesthesiology 63:A237; 1985. Chrubasik, 3.; Meynadier, J.; Scherpereel, P.: Wunsch, E. The effect of epidural somatostatin on postoperative pain. Anesth. Analg. 64:1085-1088; 1985. Dalsgaard, C.-J.; Hokfelt, T.; Johansson, 0.; Elde, R. Somato- statin immunoreactive cell bodies in the dorsal horn and the parasympathetic intermediolateral nucleus of the rat spinal cord. Neurosci. Lett. 27:335-339: 1981. DiTirro, F. J.; Martin, G. F.: Ho, R. H. A developmental study of substance-P, somatostatin, enkephalin, and serotonin immuno- reactive elements in the spinal cord of the North American op- nossum. J. Comu. Neurol. 213:241-261: 1983. ‘Fuji, K.; Senba,‘E.; Fujii, S.; Nomura, I.; Wu, J.-Y.; Ueda, Y.; Tohyama, M. Distribution, ontogeny and projections of cholecystokinin-8, vasoactive intestinal polypeptide and ~-~inobutyrate-containing neuron systems in the rat spinal cord: an immunohistochemical analysis. Neuroscience 14:881- 894; 1985. Garcia-Arraras, J. E.: Chanconie, M.; Fontaine-Perus, J. In vivo and in vitro development of somatostatin-like-immuno- reactivity in the peripheral nervous system of quail embryos. J. Neurosci. 4:154%1558; 1984. Gibson, S. J.; Polak, J. M.; Bloom, S. R.; Wall, P. D. The distribution of nine peptides in rat spinal cord with special em- phasis on the substantia gelatinosa and on the area around the central canal (lamina X). J. Camp, Neurol. 201:65-79; 1981.
13. Giesler, G. J., Jr.; Elde, R. P. Immunocytochemical studies of the peptidergic content of fibers and terminals within the lateral spinal and lateral cervical nuclei. J. Neurosci. 5:1833-1841: 1985.
14. Grimm-Jorgensen, Y. Somatostatin and calcitonin stimulate neurite regeneration of molluscan neurons in vitro. Brain Res. 403: 121-126: 1987.
15. Ho, R. H. Prenatal ontogeny of somatostatin (SOM) im- munoreactive elements in the rat spinal cord. Sot. Neurosci. Abstr. 8:807: 1982.
16. Ho, R. H. Widespread distribution of substance P- and somatostatin-immunoreactive elements in the spinal cord of the neonatal rat. Cell Tissue Res. 232:471-486: 1983.
17. Ho, R. H. Ontogeny of somatostatin (SdM) immunoreactive elements in the rat spinal cord. Sot. Neurosci. Abstr. 9455; 1983.
18. Ho, R. H.; Berelowitz, M. Somatostatin 28 (1-14) immunoreac- tivity in primary afferent neurons of the rat spinal cord. Neurosci. Lett. 46: 161-166; 1984.
19. Ho, R. H.; DePalatis, L. R. Substance P immunoreactivity in the median eminence of the North American opossum and domestic fowl. Brain Res. 189:565-569; 1980.
20. Hokfelt, T.; Elde, R.; Johansson, 0.; Luft, R.; Arimura, A. Immunohistochemical evidence for the presence of somatosta- tin, a powerful inhibitory peptide, in some primary sensory neurons. Neurosci. Lett. 1:231-235; 1975.
21. Holets, V.; Elde, R. The differential distribution and relation- ship of serotoninergic and peptidergic fibers to sympathoadrenal neurons in the intermediolateral cell column of the rat: a com- bined retrograde axonal transport and immunofluorescence study. Neuroscience 7:1155-1174; 1982.
22. Hunt, S. P.; Kelly, J. S.; Emson, P. C.; Kimmel, J. R.; Miller, R. J.; Wu, J-Y. An immunohistochemical study of neuronal Population containing neuropeptides or y-aminobutyrate within the superficial layers of the rat dorsal horn. Neuroscience 6: 1883-1898; 1981.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
3.5.
36.
37.
Inagaki, S.; Kawai, Y.; Matsuzaki, T.; Shiosaka, S.; Tohyama, M. Precise terminal fields of the descending somatostatinergic neuron system from the amygdaloid complex of the rat. J. Himforsch. 24:345-356; 1983. Johansson, 0.; Hokfelt, T.; Elde, R. P. Immunohistochemical distribution of somatostatin-like immunoreactivity in the central nervous system of the adult rat. Neuroscience 13:265-339; 1984. Krukoff, T. L.; Ciriello, J.; Calaresu, F. R. Segmental distribu- tion of peptide-like immunoreactivity in cell bodies of the tho~columb~ sympathetic nuclei of the cat. J. Comp. Neurol. 24090-102; 1985. Krukoff, T. L.; Cirieffo, J.; Calaresu, F. R. Segmental distribu- tion of peptide- and SHT-like immunoreactivity in nerve termi- nals and fibers of the thoracolumbar sympathetic nuclei of the cat. J. Comp. Neurol. 240~103-116; 1985. Krukoff, T. L.; Ciriello, J.; Calaresu, F. R. Somatostatin-like immunoreactivity in neurons, nerve terminals, and fibers of the cat spinal cord. J. Comp. Neurol. 243:13-22: 1986. Kuraishi, Y.; Hirota, N.; Sato, Y.; Hino, Y.; Satoh, M.: Takagi, H. Evidence that substance P and somatostatin transmit sepa- rate information related to pain in the spinal dorsal horn. Brain Res. 325:294-298; 1985. Lawson, S. N.; Caddy, K. W. T.; Biscoe, T. J. Development of rat dorsal root ganglion neurones: studies of cell birthdays and changes in mean cell diameter. Cell Tissue Res. 153:399-413; 1974. Massari, V. J.; Tizabi, Y.; Park, C. H.; Moody, T. W.; Helke, C. J.; O’Donohue, T. L. Distribution and origin of bombesin, substance P and somatostatin in cat spinal cord. Peptides 4:673-681; 1983. Meynadier, J.; Chrubasik, J.; Dubar, M.; Wunsch, E. Intrathecal somatostatin in terminally ill patients: a report of two cases. Pain 239-12; 1985. Nahin, R. L.; Madsen, A. M.; Giesler, G. J., Jr. Anatomical and physiological studies of the gray matter surrounding the spinal cord central canal. J. Comp. Neural. 220:321-335; 1983. Nahin, R. L.; Madsen, A. M.; Micevych, P. E.; Haist, F.; Ter- man, G. W. Giesler, G. J., Jr. Studies of the area around the central canal: efferents and afferents. Sot. Neurosci. Abstr. 10:487; 1984. Nomes, H. 0.; Das, G. D. Temporal pattern of neurogenesis in spinal cord: cytoarchitecture and directed growth of axons. Proc. Natl. Acad. Sci. USA 69:t962-1966; 1972. Nornes, H. 0.: Das. G. D. Temporal pattern of neurogenesis in spinal cord of rat. I. An autoradiographic study-time and sites of origin and migration and settling patterns of neuroblasts. Brain Res. 73:121-138; 1974. Papadopoulos, G. C.; Karamanlidis, A. N.; Dinopoulos, A.; Antonopoulos, J. Somatostatinlike immunoreactive neurons in the hedgehog (Erinaceus europaeus) and the sheep (Ovis aries) central nervous system. J. Comp. Neurol. 244: 174-192; 1986. Pickel, V. M.; Sumal, K. K.; Miller, R. J. Early prenatal devel- opment of substance P and enkephalin-containing neurons in the rat. J. Comp. Neural. 210:411-422; 1982. ^ *..
38. Rand& M.; Miletic, V. Depressant actions ot methionme- enkephalin and somatostatin in cat dorsal horn neurones ac- tivated by noxious stimuli. Brain Res. 152:19&202; 1978.
39. Sasek, C. A.; Seybold, V. S.; Elde, R. P. The immunohis- tochemical localization of nine peptides in the sacral parasym- pathetic nucleus and the dorsal gray commissure in rat spinal cord. Neuroscience 12:855-873; 1984.
40. Sawchenko, P. E.; Swanson, L. W. Immunohistochemical identification of neurons in the paraventricular nucleus of the hypothalamus that project to the medulla or to the spinal cord in the rat. J. Comu. Neurol. 205:260-272; 1982.
41. Schoenen, J.; Lotstra, F.; Vierendeels, G.; Reznik, M.; Van- derhaeghen, J. J. Substance P, enkephalins, somatostatin, cholecystokinin, oxytocin, and vasopressin in human spinal cord. Neurology 35:881-890; 1985.
42. Schroder, H. D. Somatostatin in the caudal spinal cord: an im- munohistochemic~ study of the spinal centers involved in the innervation of pelvic organs. J. Comp. Neurol. 223:40%414; 1984.

116 HO
43. Senba. E.: Shiosaka. S.: Hara, Y.; inagaki, S.: Sakanaka. M.: Takatsuki, K.; Kawai. Y.; Tohyama. M. Ontogeny of the pep- tidergic system in the rat spinal cord: immunohistochemical analysis. J. Comp. Neurol. 208:54-66; 1982.
44. Seybold, V. S.: Hylden. J. L. K.: Wilcox, G. L. lntrathecal substance P and somatostatin in rats: Behaviors indicative of sensation. Peptides 3:49-543 1982.
45. Shimada, S.; Shiosaka, S.; Takami, K.: Yamano, M.; Tohyama, M. Somatostatinergic neurons in the insular cortex project to the spinal cord: combined retrograde axonal transport and im- munohistochemical study. Brain Res. 326: 197-200; 1985.
46. Sicuteri, F.; Geppetti, F.; Marabini, S.; Lembeck. F. Pain relief by somatostatin in attacks of cluster headache. Pain 18:359-365: 1984.
47. Stefanini, M.: DeMartino, C.; Zamboni, L. Fixation of ejacu- lated spermatozoa for electron microscopy. Nature 216: 173- 174; 1967.
48. Sternberger, L. A. Immunocytochemistry. 2nd ed. New York: John Wiley and Sons; 1979.
49. Tonoue, T.; Hata, I-I.: Ohnishi, T.; Kido. N.: Nakamura. T. Somatostatin and GABA correlate with cervical autonomic nerve activity. Regul. Pept. 10:299307: 1985.
50. Tuchscherer, M. M.: Seybold, V. S. Immunohistochemical studies of substance P. cholecystokinin-octapeptide and somatostatin in dorsal root ganglia of the rat. Neuroscience 14:593-605: 1985.
51. Vincent. S. R.: McIntosh. C. H. S.: Buchan, A. M. J.: Brown. J. C. Central somatostatin systems revealed with monoclonal antibodies. J. Comp. Neurol. 238:169186: 1985.
52. Wiesenfeld-Hallin, Z. Intrathecal somatostatin modulates spinal sensory and reflex mechanisms: behavioral and elec- trophysiological studies in the rat. Neurosci. Lett. 6269-74; 1985.
53. Wiesenfeld-Hallin, Z. Substance P and somatostatin modulate spinal cord excitability via physiologically different sensory pathways. Brain Res. 372: t72-175: 1986.
54. Willis, W. D., Jr. The pain system. The neural basis of nocicep- tive transmission in the mammalian nervous system. In: Gilden- berg, P. L.. ed. Pain and headache. vol. 8. New York: Karger: 1985.
55. Willis, W. D.: Coggeshall, R. E. Sensory mechanisms of the spinal cord. New York: Plenum Press: 1978.





























