Embed Size (px)
Citation preview

R
GD
Ra
oAb
Sdc
C
AtitbtsHmdatTsnptgmicvtpbAsimegdP
*5EAoRiD
V
Neuroscience 130 (2005) 567–580
0d
EVIEW
LUTAMATE AND GABA RECEPTOR SIGNALLING IN THE
EVELOPING BRAINKG
AR
Trbchnttsosmmelb
hnc
. LUJÁN,a* R. SHIGEMOTOb AND G. LÓPEZ-BENDITOc
Facultad de Medicina and Centro Regional de Investigaciones Bi-médicas, Universidad de Castilla-La Mancha, Campus Biosanitario,vda. de Almansa s/n, 02006 Albacete, Spain
Division of Cerebral Structure, National Institute for Physiologicalciences, CREST Japan Science and Technology Corporation, Myo-aiji, Okazaki 444-8585, Japan
Instituto de Neurociencias, Universidad Miguel Hernández-CSIC,ampus de San Juan, 03550 San Juan de Alicante, Spain
bstract—Our understanding of the role played by neuro-ransmitter receptors in the developing brain has advancedn recent years. The major excitatory and inhibitory neuro-ransmitters in the brain, glutamate and GABA, activateoth ionotropic (ligand-gated ion channels) and metabo-ropic (G protein-coupled) receptors, and are generally as-ociated with neuronal communication in the mature brain.owever, before the emergence of their role in neurotrans-ission in adulthood, they also act to influence earlierevelopmental events, some of which occur prior to syn-pse formation: such as proliferation, migration, differen-iation or survival processes during neural development.o fulfill these actions in the constructing of the nervousystem, different types of glutamate and GABA receptorseed to be expressed both at the right time and at the rightlace. The identification by molecular cloning of 16 iono-ropic glutamate receptor subunits, eight metabotropiclutamate receptor subtypes, 21 ionotropic and twoetabotropic GABA receptor subunits, some of which ex-
st in alternatively splice variants, has enriched our appre-iation of how molecular diversity leads to functional di-ersity in the brain. It now appears that many differentypes of glutamate and GABA receptor subunits haverominent expression in the embryonic and/or postnatalrain, whereas others are mainly present in the adult brain.lthough the significance of this differential expression ofubunits is not fully understood, it appears that the changen subunit composition is essential for normal develop-
ent in particular brain regions. This review focuses onmerging information relating to the expression and role oflutamatergic and GABAergic neurotransmitter receptorsuring prenatal and postnatal development. © 2005 IBRO.ublished by Elsevier Ltd. All rights reserved.
Corresponding author. Tel: �34-967-599200x2933; fax: �34-967-99327.-mail address: [email protected] (R. Luján).bbreviations: AMPA, �-amino-3-hydroxy-5-methyl-4-isoxazole propi-nic acid; CNS, central nervous system; CP, cortical plate; CR, Cajal-etzius; GABA, �-aminobutyric acid; iGlu, ionotropic glutamate; IZ,
ntermediate zone; mGlu, metabotropic glutamate; NMDA, N-methyl-
T-aspartate; RMS, rostral migratory stream; SVZ, subventricular zone;Z, ventricular zone.
306-4522/05$30.00�0.00 © 2005 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2004.09.042
567
ey words: neurotransmitter receptors, AMPA, NMDA, kainateABAB receptors, mGlu receptors, development.
ContentsDevelopmental expression of neurotransmitter receptor
subunits 568AMPA receptor subunits 568NMDA receptor subunits 568Kainate receptor subunits 568mGlu receptor subtypes 569GABAA receptor subunits 570GABAB receptor subunits 570
Neurotransmitter receptor signaling in cell proliferation 570Involvement of neurotransmitter receptor in neuronal
migration 572Radial migration 572Tangential migration 572
Role of neurotransmitter receptors in early neuronaldifferentiation 573
Neurotransmitter receptor signaling during synaptogenesis574
Acquisition and localization of neurotransmitter receptors atsynapses 574Glutamatergic synapses 574GABAergic synapses 576
Concluding remarks 576cknowledgments 576eferences 576
he ability of our nervous system to learn, change andespond to the environment, reflects an underlying capa-ility of neurons to dynamically alter the strengths of theironnections. These connections, called synapses, areighly specialized sites of contact between presynapticerve terminals and postsynaptic neurons. Synapses con-ain a large variety of molecules at very high local densi-ies, including neurotransmitter receptors, associatedtructural proteins and signaling molecules, whose preciserganization gives rise to proper function. Among theseynaptic molecules, neurotransmitter receptors will ulti-ately define the functionality of a synapse. Furthermore,any of the observed changes in synaptic transmissionfficacy, that play a central role in processes such as
earning and memory or neurodegeneration, are mediatedy neurotransmitter receptors.
The present view of the central nervous system (CNS)as developed dramatically over the past few years andew principles regarding the role of neurotransmitter re-eptors in the developing CNS are beginning to emerge.
he development of the CNS results from a well charac-ved.

tntmsbt�iwe1mgrrapiScfflN(tstcpGsrov
stamcOraaaGG2
tsaciwGpf
Dr
Omsmvtbb
tipiaGpP(esm1obwai
upuapswsipoap1
rtAegahapr
R. Luján et al. / Neuroscience 130 (2005) 567–580568
erized temporo-spatial pattern of events that begins witheuronal proliferation, followed by migration, differentia-ion, and ending with synapse formation and circuit refine-ents. A growing body of evidence suggests that each
tep in that developmental sequence of the CNS involvesoth the appropriate expression and function of neuro-ransmitters and their receptors. Although glutamate and-aminobutyric acid (GABA) are the primary excitatory and
nhibitory neurotransmitters in adulthood, it is now fairlyell established that both are abundant and widespreadarly in embryonic life (Miranda-Contreras et al., 1998,999; Benı́tez-Diaz et al., 2003). Glutamate and GABAediate their actions by the activation of ionotropic (ligand-ated ion channels) and metabotropic (G protein-coupled)eceptors. Three subclasses of ionotropic glutamate (iGlu)eceptors are known and are named after their selectivegonists: i) �-amino-3-hydroxy-5-methyl-4-isoxazole pro-ionic acid (AMPA), ii) N-methyl-D-aspartate (NMDA) and
ii) kainate receptors (Hollmann and Heinemann, 1994).ixteen functional subunits may assemble in tetramericomplexes to form the following receptors: GluR1–GluR4or AMPA (that occur in two alternatively spliced versions,ip and flop); GluR5–GluR7 and KA1–KA2 for kainate; andR1, NR2A–NR2D and NR3A–B for NMDA receptors
Hollmann and Heinemann, 1994). The metabotropic glu-amate (mGlu) receptors consist of at least eight differentubtypes (mGlu1–mGlu8), that have been classified intohree groups based on their sequence homology, pharma-ological profile and coupling to intracellular transductionathways (Pin and Duvoisin, 1995; Conn and Pin, 1997).roup I mGlu receptors consist of mGlu1, mGlu5 and their
plice variants (mGlu1�, �, c, d and mGlu5a,b); group IIeceptors include mGlu2 and mGlu3; and group III consistsf mGlu4, mGlu6, mGlu7 and mGlu8, and some spliceariants.
Based on the presence of eight subunit families con-isting of 21 subunits (�1-6, �1-4, �1–4, �, �, �, �, 1-3),he ionotropic GABA receptors (GABAA receptors) displayn extraordinary structural heterogeneity. It is thought thatost functional GABAA receptors in vivo are formed upon
o-assembly of �-, �-, and �-subunits (Macdonald andlsen, 1994). The metabotropic GABA receptors (GABAB
eceptors) consist of two subunits: GABAB1, which exists inlternatively spliced forms designated 1a, b, c, d and e,nd GABAB2 (reviewed by Billinton et al., 2001; Bowery etl., 2002). Physiological responses following activation ofABAB receptors require the co-assembly of GABAB1 andABAB2 (reviewed by Couve et al., 2000; Bowery et al.,002).
In this review, we summarize the current knowledge onhe involvement of neurotransmitter receptors in neuronalignaling during development. We will focus on glutamatend GABA receptors, which are inextricably linked in theontrol of neuronal excitability, and discuss issues concern-ng their expression and role in the developing brain. Firstly,e will provide an overview of the diversity of glutamate andABA receptor subunits and their developmental expressionattern, and then discuss their potential functions in the brain
rom proliferation to synapse formation. h
evelopmental expression of neurotransmittereceptor subunits
ne indicator of the functional importance of neurotrans-itter receptor subunit diversity comes from examining the
ubunit mRNA or protein changes seen during develop-ent. Although the exact changes in subunit expression
ary with brain region, it now appears that many differentypes of neurotransmitter receptors are present in the em-ryonic brain, while others are dominant in the postnatalrain or in the adult brain.
AMPA receptor subunits. The GluR1 subunit is de-ected in the whole brain at embryonic day E15, and levelsncrease progressively during late embryonic and earlyostnatal days (Martin et al., 1998). Regionally, GluR1
ncreases in the cerebral cortex but decreases in the stri-tum with postnatal development. In the cerebellum,luR1 is expressed transiently at particular time pointsostnatally, by both granule and Purkinje cells, but from21 onwards these neurons have very low GluR1 levels
Martin et al., 1998; Fig. 1). The GluR2/3 subunits are alsoxpressed in embryonic development, whereas the GluR4ubunit is mainly expressed in the late postnatal develop-ent and adult (Hall and Bahr, 1994; Furuta and Martin,999; Metin et al., 2000; Fig. 1). Concerning the isoformsf AMPA receptors, flip variants expression dominatesefore birth and continues to be expressed into adulthood,hereas flop variants are in low abundance before P8 andre up-regulated to about the same level as the flip forms
n adulthood (Hollmann and Heinemann, 1994; Fig. 1).
NMDA receptor subunits. The functional NR1 sub-nit is ubiquitously present in the brain throughout pre- andostnatal development (Fig. 1), while the modulatory sub-nits (NR2A-D) are differentially expressed (Watanabe etl., 1993; Takai et al., 2003). The NR2A subunit is ex-ressed postnatally and widely in the brain while the NR2Bubunit is detected throughout the entire embryonic brain,ith a restricted expression to the forebrain at postnataltages (Fig. 1). The NR2C subunit appears postnatally and
s prominent in the cerebellum; the NR2D subunit is mainlyresent in the diencephalon and the brainstem at embry-nic and neonatal stages (Watanabe et al., 1993; Takai etl., 2003). The NR3 subunit is abundant within the laterenatal and early postnatal brain development (Sun et al.,998).
Kainate receptor subunits. The mRNA for all kainateeceptor subunits, except the KA-1 subunit, can be de-ected in the embryonic brain by E12 (Bahn et al., 1994).ll subunits undergo a peak in their expression in the latembryonic and early postnatal period (Fig. 1). At the re-ional level, the GluR5 subunit shows a peak of expressionround the period of birth in the sensory cortex, in CA1ippocampal interneurons (stratum oriens), the septum,nd in the thalamus, while the GluR6 subunit shows arenatal expression peak in the neocortical cingulate gy-us. The KA-1 subunit appears with the development of the
ippocampus and remains largely confined to discrete
astc
roTd1lu
aitedtddtam
FtoDetb pressed
R. Luján et al. / Neuroscience 130 (2005) 567–580 569
reas such as the CA3 region, the dentate gyrus, andubiculum, whereas the KA-2 subunit is found throughouthe CNS from early embryonic stages (Fig. 1), remainingonstant until adulthood (Bahn et al., 1994).
mGlu receptor subtypes. The expression of mGlueceptor subtypes is differentially regulated during devel-pment, showing distinct regional and temporal profiles.he two receptor subtype of group I, mGlu1 and mGlu5, areetected during embryonic development (Shigemoto et al.,992; López-Bendito et al., 2002a; Fig. 1), albeit at low
evels. However, while the level of mGlu expression grad-
ig. 1. Schematic representation of the expression of glutamate and Ghe gray scale shows the relative differences in the expression betweef mGlu receptors, and the distinct subunits of GABAB receptors, but nifferences in the onset of expression between substructures, e.g. cortxpressed relatively earlier than AMPA and kainate receptors. Regardi
he other seven subtypes, whereas the GABAB1 subunit of the GABAB
ut equalizes in the adult brain. Embryonic and postnatal ages are ex
1
ally increases during early postnatal days (Shigemoto et 1
l., 1992; López-Bendito et al., 2002a), mGlu5 expressionncreases perinatally, peaking around the second postna-al week and decreases thereafter to adult levels (Cataniat al., 1994; Romano et al., 1996, 2002; Fig. 1). Thisifferent expression pattern is also detected at the level ofheir isoforms. Thus, it seems that mGlu1� predominatesuring development, as mGlu1� and mGlu1c is mostly notetected until adulthood (Casabona et al., 1997). In con-
rast, the change in expression of mGlu5 is associated withdecline in the mGlu5a splice variant and an increase inGlu5b, which dominates in adulthood (Minakami et al.,
eptors throughout the developing rat cerebral cortex. The gradient ininct subunits for each subclass of iGlu receptors, the distinct subtypesn the different receptor subclasses or subfamilies; for details see text.s, have not been considered; for details see text. NMDA receptors areotropic receptors, mGlu1 seems to be expressed relatively earlier thanrs exceed that of the GABAB2 subunit during embryonic developmentin days.
ABA recn the distot betweeical layerng metab
recepto
995; Romano et al., 1996, 2002).

tlwda
tbtld1
emt2od1tpgu(ew1mwvoni(
iebdaitattGutvrtwdga2
mvrdcGit
Np
PmisfcctiN
dtstbcttsrc
iatb2ptd
ptTi1sdcm1cIhz
R. Luján et al. / Neuroscience 130 (2005) 567–580570
In addition, group II mGlu receptors are also differen-ially expressed. In the brain, mGlu2 mRNA expression isow at birth and increases during postnatal development,hereas mGlu3 is highly expressed at birth and decreasesuring maturation to adult levels of expression (Catania etl., 1994; Fig. 1).
Finally, with reference to group III mGlu receptors,he mRNA and protein expression for mGlu4 is low atirth and increases during postnatal development (Ca-ania et al., 1994; Elezgarai et al., 1999; Fig. 1). Simi-arly, mGlu7a levels are highest at P7 and P14, and thenecline thereafter in cortical regions (Bradley et al.,998).
GABAA receptor subunits. GABAA receptor subunitxpression is differentially regulated during brain develop-ent, with each subunit exhibiting a unique regional and
emporal developmental expression profile. Some of the1 GABAA subunits dominate expression during embry-nic development (e.g. �2, �3, and �5), whereas othersominate postnatally or in the adult brain (Laurie et al.,992; Fritschy et al., 1994). For example, the expression ofhe �1 subunit is low at birth, but increases during the firstostnatal week, whereas the �2 subunit decreases pro-ressively (Fritschy et al., 1994). Additionally, the �5 sub-nit is found throughout pre- and postnatal developmentKillisch et al., 1991). The �2/3 subunits are ubiquitouslyxpressed during development, indicating their associationith � subunits in distinct receptor subtypes (Fritschy et al.,994). The �1 and �3 subunits expression levels droparkedly during development, whereas �2 expression isidespread and remains mostly constant throughout de-elopment (Fritschy et al., 1994). Although the significancef the differential expression of GABAA receptor subunits isot completely understood, it seems that subunit switching
n certain brain regions is essential for normal developmentCuliat et al., 1994; Gunther et al., 1995).
GABAB receptor subunits. In situ hybridization andmmunohistochemical studies have defined a pattern ofarly and strong GABAB1 receptor expression in discreterain regions during embryonic development (López-Ben-ito et al., 2002b, 2003, 2004b; Kim et al., 2003; Martin etl., 2004; Panzanelli et al., 2004). GABAB1 receptor mRNA
s intensely expressed by E11, and at E12 is detected inhe hippocampal formation, cerebral cortex, intermediatend posterior neuroepithelium, and the pontine neuroepi-helium (Kim et al., 2003; Martin et al., 2004). Furthermore,he most widely studied isoforms of the GABAB1 subunit,ABAB1a and GABAB1b, seem to be developmentally reg-lated, with GABAB1b being the most abundant isoform inhe adult, while GABAB1a dominates during postnatal de-elopment (Fritschy et al., 1999; Fig. 1). However, GABAB2
eceptor mRNA and protein are not detected at the sameime period, as the expression of the GABAB1 subunit,hose isoforms greatly exceed that of the GABAB2 subunituring embryonic development but equalizes in most re-ions in the adult brain (Kim et al., 2003; López-Bendito etl., 2002b, 2004b; Martin et al., 2004; Panzanelli et al.,
004; Fig. 1). Thus it is likely that the GABAB1 subunit is pore important than the GABAB2 subunit in the early de-elopment of the CNS. Indeed, it appears that GABAB
eceptor subunits are not coordinately regulated duringevelopment. Despite the fact that functional GABAB re-eptor requires heterodimerization of GABAB1 andABAB2 subunits, the expression of each of them is under
ndependent control during embryonic development (Mar-in et al., 2004).
eurotransmitter receptor signaling in cellroliferation
roliferation of neuronal progenitors is one of the funda-ental developmental processes responsible for generat-
ng the correct number of cells of each type in the correctequence in the brain. Both cell-intrinsic and -extrinsicactors contribute to changes in cell production and affecterebral cortical growth. Among other extracellular mole-ules, neurotransmitter receptors have been implicated inhe extrinsic regulation of cell proliferation in the develop-ng telencephalon (see review by Cameron et al., 1998;guyen et al., 2001; Owens and Kriegstein, 2002).
Functional iGlu receptors emerge during terminal cellivision and early neuronal differentiation of rat neuroepi-helial cells (Maric et al., 2000). Several studies have alsohown that both NMDA and non-NMDA glutamate recep-ors are expressed in early postmitotic neurons (see reviewy Lauder, 1993; Bardoul et al., 1998), and that glutamatean inhibit DNA synthesis in cells proliferating at the cor-ical neuroepithelium when AMPA/kainate receptor are ac-ivated (LoTurco et al., 1995). However, proliferation oftriatal neuronal progenitors is promoted by an NMDAeceptor-dependent mechanism but not AMPA/kainate re-eptors at the ventral telencephalon (Luk et al., 2003).
AMPA responses are first observed in terminally divid-ng neuronal progenitors which begin to express Tuj1 (anntigen which is expressed by neuronal precursors) nearhe time of terminal division. Postmitotic neurons expressoth AMPA/kainate and NMDA receptors (Maric et al.,000). Taken together these results suggest that the ap-earance of functional iGlu receptors may regulate theransition from proliferation to postmitotic neuronalifferentiation.
The role of NMDA receptor activation controlling cellroliferation has also been shown to occur in brain regionshat retain a neurogenic population of cells through life.his is the case in the dentate gyrus of the hippocampus,
n rodents (Cameron et al., 1995), primates (Gould et al.,998) and human (Eriksson et al., 1998). Several in vivotudies have shown that blockage of NMDA receptorsuring either the first postnatal week or in adult rats, in-reases cell proliferation in the hippocampus; affectingainly granule cells (Gould et al., 1994; Cameron et al.,995). However, the mechanisms by which glutamate de-reases dentate gyrus cell proliferation remains unknown.n relation to the possible involvement of mGlu receptors, itas also been shown the presence of mGlu5 receptor inones of active neurogenesis both in the prenatal and
ostnatal brain (Di Giorgi Gerevini et al., 2004).
GsrsaVtc(soalitoG
gAfirTgapgbGs2
ucp
Fnbinstm nt corticar
R. Luján et al. / Neuroscience 130 (2005) 567–580 571
Regarding the GABAergic system, several types ofABAA receptor transcripts and subunits have been de-cribed as components of functional GABAA receptors inat neuroepithelial cells, neuroblasts and glioblasts, duringpinal and cortical neurogenesis (LoTurco et al., 1995; Mand Barker, 1995; Ma et al., 1998; Serafini et al., 1998;erkhratsky and Steinhauser, 2000). In precursor cells, in
he neocortical proliferative zone, activation of GABAA re-eptors has been shown to influence DNA synthesisLoTurco et al., 1995; Haydar et al., 2000). A more recenttudy has pointed to a more complicated scenario. Usingrganotypic mouse slice cultures in which the spatial sep-ration between the ventricular zone (VZ) and subventricu-
ar zone (SVZ) of the cerebral cortex has been maintained,nherent differences between VZ and SVZ progenitors inheir physiological responses to GABA or glutamate werebserved (Haydar et al., 2000). Exogenously applied
ig. 2. Neurotransmitter receptors are involved in the Prolif and migratieuronal progenitors while postmitotic neurons (green cells) express by GABA and Glu, respectively, has been shown to shorten the cell cy
s markedly decreased in response to GABA (black circled negativeeuronal migration of cortical neurons at early stages of development.timulates migration of neurons from the VZ and IZ, respectively, wharget destinations in the CP. The U-shape discontinuous line withigration. The gradient in the background color represents the differe
adial glia.
ABA and glutamate shortened the cell cycle of VZ pro- c
enitors; this effect suggesting a mediation by GABAA orMPA/kainate receptors. Surprisingly, in contrast to thesendings, proliferation in the SVZ markedly decreased inesponse to GABA, glutamate, and their agonists (Fig. 2).hus, depending on the cortical compartment, GABA andlutamate have differential effects on cortical cell prolifer-tion. Altogether, these results suggest that in culturedrogenitor cells of the developing neocortex, GABA andlutamate regulate cell proliferation by providing a feed-ack signal that controls division (Fig. 2). Additionally, aABA-dependent cell proliferation has also been demon-
trated in the hippocampus (Ben-Yaakov and Golan,003).
The exact mode of GABA or glutamate action is stillnresolved. Nevertheless, functional GABAergic-signalingomponents among differentiating neurons and their ca-acity to release GABA from both cell body and growth
tical neurons. AMPA responses are first observed in terminally dividing/kainate and NMDA receptors. Activation of GABA and Glu receptorsprogenitors (black circled positive symbol), while the Prolif in the SVZNeurotransmitter receptor activation has been reported to influenceBAC and GABAB-like receptor activation (red circled positive symbol)BAA receptor activation arrests migration as neurons approach theirpresents the direction followed by the cells types during Prolif andl layers. Glu, glutamate; MZ, marginal zone; Prolif, proliferation; RG,
on of coroth AMPAcle of VZsymbol).Thus, GAereas GAarrows re
one compartments emerge during neuronal lineage pro-

gbamsb(Omss
Im
AonnIntmMiaiPHcGe
prsoiLsrf
gtaov1tacicp
tse
ttcGgwn(vbl
mtgracG[lt
imaimvwbtBhm(eec3s
msiAtGartAstrrt
R. Luján et al. / Neuroscience 130 (2005) 567–580572
ression (Maric et al., 2001). Those postmitotic cells coulde the source of GABA or glutamate that leads to thectivation of GABA and glutamate receptors expressed byitotic cells in the proliferative zones. Regardless of the
ubcellular compartment, it seems that at least GABA cane released in a paracrine manner to active its receptorsDemarque et al., 2002), as synapses are still not present.ther interesting questions as how are GABA and gluta-ate concentrations regulated or what kind of intracellular
ignaling mechanisms are involved in their activation aretill unanswered.
nvolvement of neurotransmitter receptor in neuronaligration
fter division, most cortical neurons migrate from their sitef origin to their final destination in the cerebral cortex. Thiseuronal movement is essential for the establishment oformal brain organization (see for instance Hatten, 1999).n the developing cerebral cortex, glutamatergic projectingeurons are primarily generated in the VZ and then moveo the developing cortical plate (CP) by means of “radialigration” (see review by Marı́n and Rubenstein, 2001).ost GABAergic interneurons, however, originate mainly
n the medial ganglionic eminence of the ventral telenceph-lon and follow tangential migratory routes through the
ntermediate zone (IZ) to reach the cortex (see review byarnavelas, 2000; Marı́n and Rubenstein, 2001; Fig. 3).owever, the caudal ganglionic eminence has been re-ently demonstrated to give rise to a distinct population ofABAergic interneurons (Nery et al., 2002; López-Benditot al., 2004a).
Neurotransmitter receptor activation has been re-orted to influence neuronal migration of cortical neu-ons at early stages of development. The presence ofome of these neurotransmitter receptors in the embry-nic cerebral cortex has been shown by in situ hybrid-
zation and immunohistochemistry (Behar et al., 2001;ópez-Bendito et al., 2002b; Fig. 3). In the followingections, we will review the role of neurotransmittereceptors in the processes of neuronal migration in theorebrain.
Radial migration. Due to its early expression pattern,lutamate is a likely candidate to play a role as a chemoat-ractant signal in the developing mouse cerebral cortex. Anssay measuring the effects of glutamate on the migrationf acutely dissociated murine embryonic cortical cells, re-ealed stimulation of the migration of cortical neurons to a0-fold greater extent than GABA (Behar et al., 1999). Inhe cerebellum, if NMDA receptors are blocked by specificntagonists or by high Mg2� concentration, the granuleell migration from the external to the internal granule layers retarded (Rossi and Slater, 1993). Conversely, an in-rease of NMDA or glycine concentration increases thisrocess (Komuro and Rakic, 1993).
Some illuminating results on the role of neurotransmit-er receptors on cortical radial migration, came from aeries of pharmacological studies, which showed the influ-
nce of the activation of GABA receptors in neuronal mo- oility and migration in both dissociated cells and organo-ypic slice cultures (Behar et al., 1996, 2000, 2001). Inultured rat brain slices, it has been determined thatABAC and GABAB-like receptor activation stimulates mi-ration of neurons from the VZ and IZ, respectively,hereas GABAA receptor activation arrests migration aseurons approach their target destinations in the CPBehar et al., 1998). Altogether these data show that acti-ation of neurotransmitter receptors, independent of therain area or neuronal phenotype, plays a role in control-
ing the migration of neurons.
Tangential migration. The cellular and molecularechanisms controlling tangential migration of cortical in-
erneurons are still relatively unknown. It has been sug-ested that activation of AMPA receptors leads to neuriteetraction in tangentially migrating interneurons (Poluch etl., 2001). In addition, tangentially migrating cells in theerebral cortex posses functional NMDA, non-NMDA andABAA receptors whose activation leads to changes in
Ca2�]i (Soria and Valdeolmillos, 2002). However, the re-ationship between these changes in [Ca2�]i signaling andhe actual process of tangential migration is still unclear.
Several studies though have provided some illuminat-ng ideas on the role of GABA receptors in tangential
igration. Interestingly, one of these studies involved thectivation of neurotransmitter receptors and the end result,
n particular, reinforced the role of GABAB receptors inigration of neurons (López-Bendito et al., 2003). Using in
itro embryonic rat organotypic cultures, in combinationith immunohistochemical techniques, it was shown thatlockade of GABAB receptor modified the distribution of
angential migratory interneurons within the cortex (López-endito et al., 2003; Fig. 3). Additionally, other studiesave shown the functionality of some of these neurotrans-itter receptors, in the tangentially migratory interneurons
Metin et al., 2000; Soria and Valdeolmillos, 2002). Forxample, GABAergic calbindin-positive IZ cells seem toxpress inwardly rectifying calcium-permeable AMPA re-eptors, but not NMDA receptors (Metin et al., 2000; Fig.). Likewise, these cells displayed electrophysiological re-ponses to GABAA agonists.
Another population of neurons that adopt a tangentialode of migration to reach their final target is the precur-
ors of olfactory interneurons. These interneurons are bornn the embryonic subpallium (Altman, 1969; Lois andlvarez-Buylla, 1994) and their migration continues
hrough adulthood, providing a constant supply of newABAergic local circuit neurons to the olfactory bulb (Loisnd Alvarez-Buylla, 1994). Migration of olfactory interneu-ons in the adult occurs along a highly restricted routeermed the rostral migratory stream (RMS; Lois andlvarez-Buylla, 1994; Kornack and Rakic, 2001). A recenttudy pointed out the possibility that GABA may also affecthese neuronal progenitors via the activation of specificeceptors (Wang et al., 2003). Every neuronal progenitoresponded to GABA via picrotoxin-sensitive GABAA recep-or activation, demonstrating that the neuronal progenitors
f the SVZ/RMS contain and are depolarized by GABA.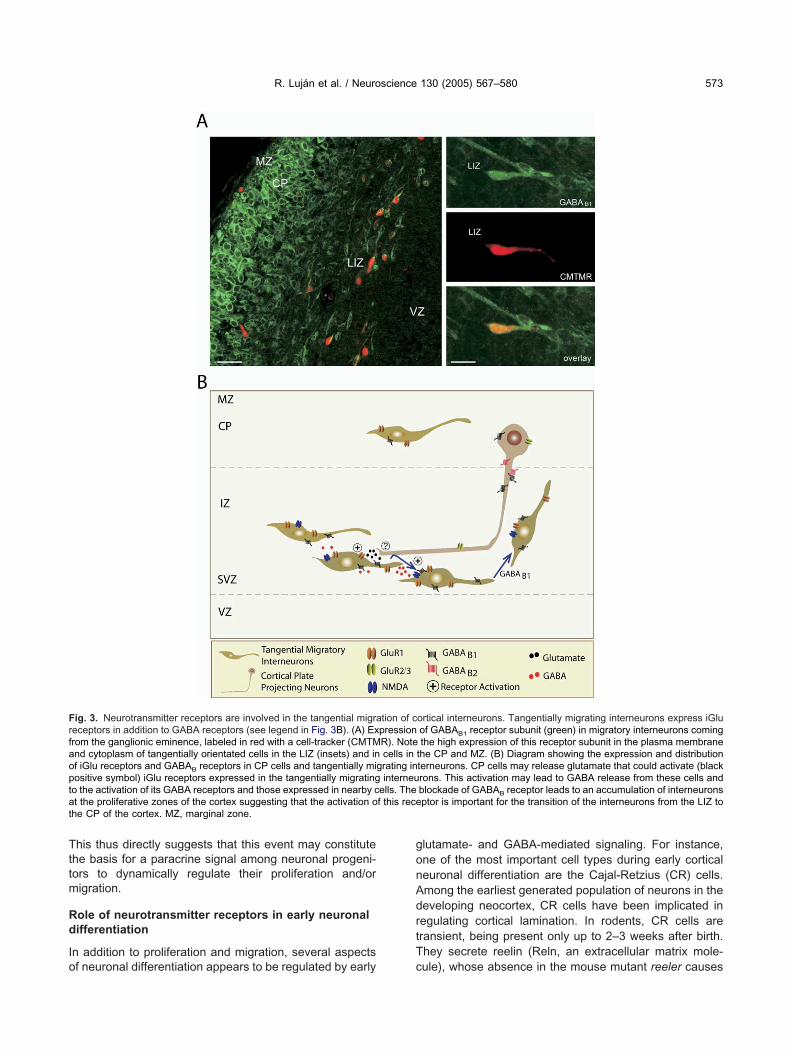
Tttm
Rd
Io
gonAdrtT
Frfaoptat
R. Luján et al. / Neuroscience 130 (2005) 567–580 573
his thus directly suggests that this event may constitutehe basis for a paracrine signal among neuronal progeni-ors to dynamically regulate their proliferation and/origration.
ole of neurotransmitter receptors in early neuronalifferentiation
n addition to proliferation and migration, several aspects
ig. 3. Neurotransmitter receptors are involved in the tangential migraeceptors in addition to GABA receptors (see legend in Fig. 3B). (A) Exrom the ganglionic eminence, labeled in red with a cell-tracker (CMTMnd cytoplasm of tangentially orientated cells in the LIZ (insets) and inf iGlu receptors and GABAB receptors in CP cells and tangentially miositive symbol) iGlu receptors expressed in the tangentially migratingo the activation of its GABA receptors and those expressed in nearby ct the proliferative zones of the cortex suggesting that the activation ohe CP of the cortex. MZ, marginal zone.
f neuronal differentiation appears to be regulated by early c
lutamate- and GABA-mediated signaling. For instance,ne of the most important cell types during early corticaleuronal differentiation are the Cajal-Retzius (CR) cells.mong the earliest generated population of neurons in theeveloping neocortex, CR cells have been implicated inegulating cortical lamination. In rodents, CR cells areransient, being present only up to 2–3 weeks after birth.hey secrete reelin (Reln, an extracellular matrix mole-
ortical interneurons. Tangentially migrating interneurons express iGluof GABAB1 receptor subunit (green) in migratory interneurons comingthe high expression of this receptor subunit in the plasma membrane
the CP and MZ. (B) Diagram showing the expression and distributionterneurons. CP cells may release glutamate that could activate (blackrons. This activation may lead to GABA release from these cells andblockade of GABAB receptor leads to an accumulation of interneuronsptor is important for the transition of the interneurons from the LIZ to
tion of cpressionR). Notecells in
grating ininterneu
ells. Thef this rece
ule), whose absence in the mouse mutant reeler causes

aFGsatettRiacliwvt1
toappo1pWta1tme
Ns
TifitcrfcmdvamcAtr2C
ikp2rsO2ocsaou
ddcdt
Ar
AGstcgrtomra
icEsNada4dap1ht2cvnil
R. Luján et al. / Neuroscience 130 (2005) 567–580574
severe cortical laminar disruption (see review byrotscher, 1998; Curran and D’Arcangelo, 1998; Tissir andoffinet, 2003). The molecular cascade acting down-tream of Reln is just beginning to be understood (Walshnd Goffinet, 2000). However, the functional mechanismshat regulate Reln synthesis and secretion have been littlexplored. CR cells express diverse types of neurotransmit-er receptors, including iGlu receptors and GABAA recep-ors (Schwartz et al., 1998; Mienville and Pesold, 1999).ecently, the expression of the mGlu receptor 1�, mGlu1�,
n CR cells (Martı́nez-Galán et al., 2001; López-Bendito etl., 2002a) and the existence of functional mGlu1 in layer Iells of the postnatal mouse cerebral cortex was estab-ished (Martı́nez-Galán et al., 2001). Postnatal CR cellsncur a dramatic increase in their NMDA receptor density,hich ultimately may trigger their death. The fact that inivo pharmacological blockade of NMDA receptors cur-ailed the disappearance of CR cells (Mienville and Pesold,999) strongly supports such a hypothesis.
Apart from the correct positioning and laminar acquisi-ion of cortical neurons, glutamate causes a graded seriesf changes in pyramidal neuron cytoarchitecture inducingselective inhibition in dendritic outgrowth and dendritic
runing at subtoxic levels. Some studies have also re-orted that NMDA receptor activation promotes neuriteutgrowth from cerebellar granule cells (Pearce et al.,987) and dendritic outgrowth and branching of hippocam-al cells (Mattson et al., 1988; Brewer and Cotman, 1989;ilson et al., 2000). Furthermore, GABAA receptor activa-
ion has also been shown to promote neurite outgrowthnd maturation of GABAergic interneurons (Barbin et al.,993; Marty et al., 1996). In addition it is seen to regulate
he morphological development of cortical neurons throughembrane depolarization and increases in [Ca2�]i (Marict al., 2001).
eurotransmitter receptor signaling duringynaptogenesis
he establishment of neural networks begins with grow-ng axons recognizing their postsynaptic targets, thusorming synaptic contacts. This process can be dividednto two separate phases: the first one (synapse forma-ion) comprises the establishment of functional synapticommunication, and the second phase (synapse matu-ation) comprises the functional and morphological dif-erentiation of synapses; both phases requiring tightommunication between pre- and postsynaptic ele-ents. How specific pre- and postsynaptic elements areifferentiated, how synaptic contacts are generated de-elopmentally and how these synapses are remodelednd maintained in mature brain seems to be in partediated by the action of neurotransmitter receptors. A
omplete description on the involvement of NMDA andMPA receptors in the formation, stabilization, matura-
ion and elimination of synapses has been extensivelyeviewed elsewhere (see for instance Bolton et al.,000; Lee and Sheng, 2000; Zhang and Poo, 2001;
ohen-Cory, 2002; Garner et al., 2002). Although the anvolvement of mGlu receptors in such processes is un-nown, it has been recently suggested that they may alsoarticipate in synapse-stabilizing responses (Miskevich et al.,002). Additionally, a description on the role of GABAA
eceptor subunits in synaptogenesis has also been exten-ively reviewed recently (see for instance Ben-Ari, 2002;wens and Kriegstein, 2002; Fritschy et al., 2003; Meier,003). Altogether, those reviews outline that the assemblyf CNS synapses likely involve a complex interplay ofell–cell adhesion, interneuronal signaling and site-pecific recruitment of protein complexes. Furthermore,ctivity-dependent mechanisms control postsynaptic levelf neurotransmitter receptors, with different mechanismssed for the synaptic targeting of distinct receptor types.
Much less is known, however, about the acquisition ofifferent glutamate and GABA receptor subunits occurringuring synapse formation, and how they accumulate andhange at synapses to reach their final distribution patternuring synapse maturation and adulthood. In the next sec-ion, we will concentrate on such processes.
cquisition and localization of neurotransmittereceptors at synapses
critical aspect in the development of glutamatergic andABAergic synapses is the progressive recruitment and
ubsequent accumulation of neurotransmitter receptors atheir functional site. Such processes are important, be-ause proper function of synaptic transmission at anyiven synapse, depends on adequate placement of neu-otransmitter receptors of the appropriate number andype, in the neuronal membrane. The differential regulationf the distinct subunits during pre- and postnatal develop-ent (see Developmental expression of neurotransmitter
eceptor subunits) may favor the correct acquisition andccumulation processes of receptor complexes.
Glutamatergic synapses. Electrophysiological andmmunohistochemical studies have shown that NMDA re-eptors are expressed firstly in glutamatergic synapses.arly in postnatal development, and during development,ynapses acquire AMPA receptors with little change inMDA receptor numbers (Golshani et al., 1998; Petralia etl., 1999). With regard to their localization, during postnatalevelopment and adulthood, AMPA and NMDA receptorsre concentrated at the postsynaptic specialization (Fig.). However, while AMPA receptors seem to be evenlyistributed along the postsynaptic specialization (Nusser etl., 1998b; Somogyi et al., 1998), NMDA receptors arereferentially distributed to the center of it (Somogyi et al.,998; Racca et al., 2000). Interestingly, NMDA receptorsave been found on membranes postsynaptic to GABAergic
erminals (Fig. 4) in the adult hippocampus (Gundersen et al.,004), though no information is yet available about the factorsontrolling their recruitment in inhibitory synapses during de-elopment. Finally, the acquisition and arrangement of kai-ate receptors at synapses remains to be fully established. It
s well known, however, that the KA-1 and KA-2 subunits areocated at synaptic sites, as well as at presynaptic sites in the
dulthood (Darstein et al., 2003; Fig. 4).
iiaofatnbcdace
tmdeccdaa2
slt
FatrGEgidr asing tera ciated wi
R. Luján et al. / Neuroscience 130 (2005) 567–580 575
The acquisition and distribution of mGlu receptors dur-ng development seems to be different to that described forGlu receptors. This may be because mGlu receptors arebsent from the postsynaptic specialization. Electrophysi-logical studies (Golshani et al., 1998) have shown thatunctional mGlu receptors are detected in cortical syn-pses from birth (approximately at the same time as func-ional NMDA receptors), although only after the first post-atal week could slow EPSPs mediated by mGlu receptorse evoked by high-frequency stimulation. Immunohisto-hemical studies have demonstrated that early in postnatalevelopment in the cerebral cortex, group I mGlu receptorsre excluded from the main body of the postsynaptic spe-ialization (Fig. 4). Its highest concentration occurs at the
ig. 4. Schematic diagrams showing the location of glutamate and GAnd adulthood. Panels A and B show the localization of glutamate rece
he localization of GABA receptors at excitatory and inhibitory synapseceptor subunits varies among brain regions and cell types. ThereABAergic synapses. NMDA, AMPA and kainate receptors are concenarly in postnatal development, and during development, synapses acroup I mGlu receptors are absent from the postsynaptic specialization
n the preterminal portion of axon terminal, whereas group III mGu reevelopment, mGlu receptors progressively concentrate at those subeceptor subunits are concentrated at synapses opposite to GABA relend postsynaptic sites (panels C and D). They are mainly found asso
dge of the postsynaptic specialization (termed perisynap- a
ic location), as well as along the extrasynaptic plasmaembrane (López-Bendito et al., 2002a; Fig. 4). A similaristribution for mGlu1 and mGlu5 is seen in the adult (Lujánt al., 1996, 1997; López-Bendito et al., 2002a). In theerebellum, however, mGlu1 is already present in Purkinjeell spines before the arrival of excitatory synapses, and asevelopment progresses, mGlu1 accumulates in perisyn-ptic positions, in association with the maturation of par-llel fiber-Purkinje cell synapses (López-Bendito et al.,001).
The acquisition of group II and III mGlu receptors atynapses is unexplored. We mainly know their final
ocalization in the adulthood. Here, group II mGlu recep-ors, mGlu and mGlu , can be found both at postsyn-
tors at synapses in the cerebral cortex during postnatal developmentxcitatory and inhibitory synapses, respectively. Panels C and D showctively. The precise localization of the different glutamate and GABAschematic diagrams are not representative of all glutamatergic or
the postsynaptic specialization, as well as extrasynaptically (panel A).PA receptors with little change in NMDA receptor numbers. However,ng peri- or extrasynaptically (panel A). Group II mGlu receptors occurare found in the presynaptic active zone (panel A). During postnatalcompartments to reach their final density in the adult brain. GABAA
minals (panel D). In contrast, GABAB receptors can be present at pre-th glutamatergic synapses (panel C).
BA recepptors at ees, respefore, thetrated inquire AM, occurriceptorscellular
2 3
ptic and presynaptic sites (Fig. 4), depending on the

bmjapamatzemtaDgSaagi
rhcn�mnnpSlratGha
oorEcptdtaa2p(e2ac
aBe
fat(La
C
Abtrrtetacmtfatranndlhbpcrcmdfiptnssaci
AMsd
A
R. Luján et al. / Neuroscience 130 (2005) 567–580576
rain region and cell type. At their postsynaptic location,Glu2 is preferentially located outside the synapse (Lu-
án et al., 1997). In contrast, mGlu3 is found to bessociated with glutamatergic synapses, including theostsynaptic specialization, and glial cells (Tamaru etl., 2001; Fig. 4). At presynaptic locations, mGlu2 andGlu3 are concentrated in the pre-terminal portion ofxons or along the extrasynaptic membrane of axon
erminals, and always outside the presynaptic activeone (Luján et al., 1997; Shigemoto et al., 1997; Tamarut al., 2001; Fig. 4). Regarding group III mGlu receptors,Glu4, mGlu7 and mGlu8 are mainly concentrated along
he presynaptic active zone of glutamatergic (Shigemoto etl., 1996, 1997; Kinoshita et al., 1998; Corti et al., 2002;alezios et al., 2002; Somogyi et al., 2003) and GABAer-ic synapses (Corti et al., 2002; Dalezios et al., 2002;omogyi et al., 2003; Fig. 4), where they may act as auto-nd heteroreceptors, respectively. No information is as yetvailable about the factors that control this differential tar-eting to different subcellular domains or their recruitment
n inhibitory synapses during development.
GABAergic synapses. The involvement of GABAA
eceptors in the construction of postsynaptic domains of in-ibitory synapses is not fully known. GABAA receptors areoncentrated at synapses opposite to GABA releasing termi-als (Fig. 4), but receptor complexes containing only �- and-subunits form channels that are inserted in the plasmaembrane (Gunther et al., 1995). While the � subunits areot necessary for receptor assembly and translocation to theeuronal surface, they seem to be required for clustering ofostsynaptic GABAA receptor subtypes (Essrich et al., 1998;chweizer et al., 2003). GABAA receptor subunits are also
ocated along the extrasynaptic plasma membrane of neu-ons (Fig. 4), generally at low densities (Nusser et al., 1995),lthough the � subunit is localized exclusively at extrasynap-ic sites (Nusser et al., 1998a; Fig. 4). Furthermore, someABAA receptor subunits (e.g. the �6, �2/3 and �2 subunits)ave also been found at glutamatergic synapses (Nusser etl., 1996, 1998a).
No information is as yet available about the acquisitionf GABAB receptors at synapses. They show, however,ne of the most intriguing locations of all neurotransmittereceptors during postnatal development and adulthood.lectrophysiological, pharmacological and immunohisto-hemical studies have identified GABAB receptors onostsynaptic sites and on presynaptic GABAergic and glu-amatergic axons (Misgeld et al., 1995). Early in postnatalevelopment, GABAB receptors are located at extrasynap-ic and perisynaptic sites, as well as at pre- and postsyn-ptic sites, similarly to the distribution observed in thedulthood (López-Bendito et al., 2002b; Panzanelli et al.,004; Fig. 4). They are abundant on dendritic spinesostsynaptic to glutamatergic axon terminals in the brainFritschy et al., 1999; Gonchar et al., 2001; López-Benditot al., 2002b, 2004b; Kulik et al., 2002, 2003; Luján et al.,004) and on the postsynaptic specialization (Billinton etl., 2001; Gonchar et al., 2001; Kulik et al., 2002). In
ontrast, GABAB receptors are rarely associated with syn-pses opposite to GABA releasing terminals (López-endito et al., 2002b, 2004b; Kulik et al., 2002, 2003; Lujánt al., 2004).
At their presynaptic location, GABAB receptors areound in the preterminal portion of axons, in the extrasyn-ptic membrane of axon terminals and in presynaptic ac-ive zones of glutamatergic and GABAergic synapsesLópez-Bendito et al., 2002b; Kulik et al., 2002, 2003;uján et al., 2004; Fig. 4), where they likely act as hetero-nd autoreceptors, respectively.
oncluding remarks
combination of electrophysiological, molecular, in situ hy-ridization and immunohistochemical techniques has begun
o shed light on the role and distribution of neurotransmittereceptors in the developing brain. This review illuminatesecent evidence showing how glutamate and GABA recep-ors exert different roles during primary nervous structurestablishment prior to the emergence of their role in neuro-
ransmission. In this way, the early activation of glutamatend GABA receptor subunits, expressed by several kinds ofells, appears to account for the regulation of proliferation,igration and differentiation during CNS development. Addi-
ionally, early expression of the receptor subunits accountsor their involvement in the establishment of synaptic contactnd the refinement of neuronal circuits. Moreover, changes in
he level of expression and distribution of neurotransmittereceptors are critical steps in normal synapse development,nd abnormalities in these processes may be responsible foreural diseases. Which specific changes are critical to bothormal development and disease processes remain to beefined. This may not be particularly easy because the mo-
ecular composition and functional properties of synapses areeterogeneous. Indeed, glutamate and GABA receptors cane targeted during synaptogenesis to any subcellular com-artment of the neuronal surface, in a receptor subunit- andell type-dependent manner. Presumably, the different neu-otransmitter receptor subunits are targeted to specific sub-ellular compartments because their intrinsic properties areost suited for the physiological functions at those sitesuring development and in the adulthood. Continued identi-cation of the molecular components of individual synapsesromises to offer new insights on the mechanisms that con-
rol synaptogenesis processes and the signaling betweeneurons in the developing brain. A more complete under-tanding of the roles of the many neurotransmitter receptorubunits described in this review, and of the functional inter-ction with other signaling proteins, should provide an in-reasing understanding of brain development and neurolog-
cal diseases.
cknowledgments— The authors are grateful to Diane Latawiec,Sc, for the English revision of the manuscript. This work was
upported by grants from the Consejerı́a de Sanidad of the Juntae Comunidades de Castilla-La Mancha.
REFERENCES
ltman J (1969) Autoradiographic and histological studies of postnatal
neurogenesis: IV. Cell proliferation and migration in the anterior
B
B
B
B
B
B
B
B
B
B
B
B
B
B
B
B
C
C
C
C
C
C
C
C
C
C
D
D
D
D
E
E
E
F
F
F
F
F
G
G
G
R. Luján et al. / Neuroscience 130 (2005) 567–580 577
forebrain, with special reference to persisting neurogenesis in theolfactory bulb. J Comp Neurol 137:433–457.
ahn S, Volk B, Wisden W (1994) Kainate receptor gene expression inthe developing rat brain. J Neurosci 14:5525–5547.
arbin G, Pollard H, Gaiarsa JL, Ben-Ari Y (1993) Involvement ofGABAA receptors in the outgrowth of cultured hippocampal neu-rons. Neurosci Lett 152:150–154.
ardoul M, Drain MJ, Konig N (1998) Modulation of intracellular cal-cium in early neural cells by non-NMDA ionotropic glutamate re-ceptors. Perspect Dev Neurobiol 5:353–371.
ehar TN, Li YX, Tran HT, Ma W, Dunlap V, Scott C, Barker JL (1996)GABA stimulates chemotaxis and chemokinesis of embryonic cor-tical neurons via calcium-dependent mechanisms. J Neurosci16:1808–1818.
ehar TN, Schaffner AE, Scott CA, O’Connell C, Barker JL (1998)Differential response of cortical plate and ventricular zone cells toGABA as a migration stimulus. J Neurosci 18:6378–6387.
ehar TN, Scott CA, Greene CL, Wen X, Smith SV, Maric D, Liu QY,Colton CA, Barker JL (1999) Glutamate acting at NMDA receptorsstimulates embryonic cortical neuronal migration. J Neurosci19:4449–4461.
ehar TN, Schaffner AE, Scott CA, Greene CL, Barker JL (2000)GABA receptor antagonists modulate postmitotic cell migration inslice cultures of embryonic rat cortex. Cereb Cortex 10:899–909.
ehar TN, Smith SV, Kennedy RT, McKenzie JM, Maric I, Barker JL(2001) GABA(B) receptors mediate motility signals for migratingembryonic cortical cells. Cereb Cortex 11:744–753.
en-Ari Y (2002) Excitatory actions of GABA during development: thenature of the nurture. Nat Rev Neurosci 3:728–739.
enı́tez-Diaz P, Miranda-Contreras L, Mendoza-Briceño RV, Peña-Contreras Z, Palacios-Prü E (2003) Prenatal and postnatal con-tents of amino acid neurotransmitters in mouse parietal cortex. DevNeurosci 25:366–374.
en-Yaakov G, Golan H (2003) Cell proliferation in response to GABAin postnatal hippocampal slice culture. Int J Dev Neurosci21:153–157.
rewer GJ, Cotman CW (1989) NMDA receptor regulation of neuronalmorphology in cultured hippocampal neurons. Neurosci Lett99:268–273.
illinton A, Ige AO, Bolam JP, White JH, Marshall FH, Emson PC(2001) Advances in the molecular understanding of GABA(B) re-ceptors. Trends Neurosci 24:277–282.
olton MM, Blanpied TA, Ehlers MD (2000) Localization and stabili-zation of ionotropic glutamate receptors at synapses. Cell Mol LifeSci 57:1517–1525.
owery NG, Bettler B, Frostl W, Gallagher JP, Marshall F, Raiteri M,Bonner TI, Enna SJ (2002) International Union of Pharmacology:XXXIII. Mammalian �-aminobutyric acidB receptors: structure andfunction. Pharmacol Rev 54:247–264.
radley SR, Rees HD, Yi H, Levey AI, Conn PJ (1998) Distribution anddevelopmental regulation of metabotropic glutamate receptor 7a inrat brain. J Neurochem 71:636–645.
ameron HA, McEwen BS, Gould E (1995) Regulation of adult neu-rogenesis by excitatory input and NMDA receptor activation in thedentate gyrus. J Neurosci 15:4687–4692.
ameron HA, Hazel TG, McKay RD (1998) Regulation of neurogen-esis by growth factors and neurotransmitters. J Neurobiol36:287–306.
asabona G, Knopfel T, Kuhn R, Gasparini F, Baumann P, Sortino MA,Copani A, Nicoletti F (1997) Expression and coupling to polyphos-phoinositide hydrolysis of group I metabotropic glutamate receptors inearly postnatal and adult rat brain. Eur J Neurosci 9:12–17.
atania MV, Landwehrmeyer GB, Testa CM, Standaert DG, Penney JBJr, Young AB (1994) Metabotropic glutamate receptors are differen-tially regulated during development. Neuroscience 61:481–495.
ohen-Cory S (2002) The developing synapse: construction and mod-
ulation of synaptic structures and circuits. Science 298:770–776.onn PJ, Pin JP (1997) Pharmacology and functions of metabotropicglutamate receptors. Annu Rev Pharmacol Toxicol 37:205–237.
orti C, Aldegheri L, Somogyi P, Ferraguti F (2002) Distribution andsynaptic localisation of the metabotropic glutamate receptor 4(mGluR4) in the rodent CNS. Neuroscience 110:403–420.
ouve A, Moss SJ, Pangalos MN (2000) GABAB receptors: a newparadigm in G protein signalling. Mol Cell Neurosci 16:296–312.
uliat CT, Stubbs LJ, Montgomery CS, Russell LB, Rinchik EM (1994)Phenotypic consequences of deletion of the gamma 3, alpha 5, orbeta 3 subunit of the type A gamma-aminobutyric acid receptor inmice. Proc Natl Acad Sci USA 91:2815–2818.
urran T, D’Arcangelo G (1998) Role of reelin in the control of braindevelopment. Brain Res Brain Res Rev 26:285–294.
alezios Y, Luján R, Shigemoto R, Roberts JDB, Somogyi P (2002)The presence of mGluR7a in the presynaptic active zones ofGABAergic and non-GABAergic terminals on interneurones in therat somatosensory cortex. Cereb Cortex 12:961–974.
arstein M, Petralia RS, Swanson GT, Wenthold RJ, Heinemann SF(2003) Distribution of kainate receptor subunits at hippocampalmossy fiber synapses. J Neurosci 23:8013–8019.
emarque M, Represa A, Becq H, Khalilov I, Ben-Ari Y, Aniksztejn L(2002) Paracrine intercellular communication by a Ca2�- andSNARE-independent release of GABA and glutamate prior to syn-apse formation. Neuron 36:1051–1061.
i Giorgi Gerevini VD, Caruso A, Cappuccio I, Ricci Vitiani L, RomeoS, Della Rocca C, Gradini R, Melchiorri D, Nicoletti F (2004) ThemGlu5 metabotropic glutamate receptor is expressed in zones ofactive neurogenesis of the embryonic and postnatal brain. BrainRes Dev Brain Res 150:17–22.
lezgarai I, Benitez R, Mateos JM, Lazaro E, Osorio A, Azkue JJ,Bilbao A, Lingenhoehl K, Van Der Putten H, Hampson DR, Kuhn R,Knopfel T, Grandes P (1999) Developmental expression of thegroup III metabotropic glutamate receptor mGluR4a in the medialnucleus of the trapezoid body of the rat. J Comp Neurol411:431–440.
riksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C,Peterson DA, Gage FH (1998) Neurogenesis in the adult humanhippocampus. Nat Med 4:1313–1317.
ssrich C, Lorez M, Benson JA, Fritschy JM, Luscher B (1998)Postsynaptic clustering of major GABAA receptor subtypes re-quires the gamma 2 subunit and gephyrin. Nat Neurosci 1:563–571.
ritschy JM, Paysan J, Enna A, Mohler H (1994) Switch in the expres-sion of rat GABAA-receptor subtypes during postnataldevelopment: an immunohistochemical study. J Neurosci 14:5302–5324.
ritschy JM, Meskenaite V, Weinmann O, Honer M, Benke D, MohlerH (1999) GABAB-receptor splice variants GB1a and GB1b in ratbrain: developmental regulation, cellular distribution and extrasyn-aptic localization. Eur J Neurosci 11:761–768.
ritschy JM, Schweizer C, Brunig I, Luscher B (2003) Pre- and post-synaptic mechanisms regulating the clustering of type A gamma-aminobutyric acid receptors (GABAA receptors). Biochem SocTrans 31:889–892.
rotscher M (1998) Cajal-Retzius cells, Reelin, and the formation oflayers. Curr Opin Neurobiol 8:570–575.
uruta A, Martin LJ (1999) Laminar segregation of the cortical plateduring corticogenesis is accompanied by changes in glutamatereceptor expression. J Neurobiol 39:67–80.
arner CC, Zhai RG, Gundelfinger ED, Ziv NE (2002) Molecular mech-anisms of CNS synaptogenesis. Trends Neurosci 25:243–251.
olshani P, Warren RA, Jones EG (1998) Progression of change inNMDA, non-NMDA, and metabotropic glutamate receptor func-tion at the developing corticothalamic synapse. J Neurophysiol80:143–154.
onchar Y, Pang L, Malitschek B, Bettler B, Burkhalter A (2001)Subcellular localization of GABA(B) receptor subunits in rat visual
cortex. J Comp Neurol 431:182–197.
G
G
G
G
H
H
H
H
K
K
K
K
K
K
K
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
M
M
M
M
M
M
M
M
M
M
M
R. Luján et al. / Neuroscience 130 (2005) 567–580578
ould E, Cameron HA, McEwen BS (1994) Blockade of NMDA recep-tors increases cell death and birth in the developing rat dentategyrus. J Comp Neurol 340:551–565.
ould E, Tanapat P, McEwen BS, Flugge G, Fuchs E (1998) Proliferationof granule cell precursors in the dentate gyrus of adult monkeys isdiminished by stress. Proc Natl Acad Sci USA 95:3168–3171.
undersen V, Holten AT, Storm-Mathisen J (2004) GABAergic syn-apses in hippocampus exocytose aspartate on to NMDA receptors:quantitative immunogold evidence for co-transmission. Mol CellNeurosci 26:156–165.
unther U, Benson J, Benke D, Fritschy JM, Reyes G, Knoflach F,Crestani F, Aguzzi A, Arigoni M, Lang Y (1995) Benzodiazepine-insensitive mice generated by targeted disruption of the gamma 2subunit gene of gamma-aminobutyric acid type A receptors. ProcNatl Acad Sci USA 92:7749–7753.
all RA, Bahr BA (1994) AMPA receptor development in rattelencephalon: [3H]AMPA binding and Western blot studies. J Neu-rochem 63:1658–1665.
atten ME (1999) Central nervous system neuronal migration. AnnuRev Neurosci 22:511–539.
aydar TF, Wang F, Schwartz ML, Rakic P (2000) Differential modu-lation of proliferation in the neocortical ventricular and subventricu-lar zones. J Neurosci 20:5764–5774.
ollmann M, Heinemann S (1994) Cloned glutamate receptors. AnnuRev Neurosci 17:31–108.
illisch I, Dotti CG, Laurie DJ, Luddens H, Seeburg PH (1991) Expres-sion patterns of GABAA receptor subtypes in developing hip-pocampal neurons. Neuron 7:927–936.
im MO, Li S, Park MS, Hornung JP (2003) Early fetal expression ofGABA(B1) and GABA(B2) receptor mRNAs on the development ofthe rat central nervous system. Brain Res Dev Brain Res 143:47–55.
inoshita A, Shigemoto R, Ohishi H, van der Putten H, Mizuno N(1998) Immunohistochemical localization of metabotropic gluta-mate receptors, mGluR7a and mGluR7b, in the central nervoussystem of the adult rat and mouse: a light and electron microscopicstudy. J Comp Neurol 393:332–352.
omuro H, Rakic P (1993) Modulation of neuronal migration by NMDAreceptors. Science 260:95–97.
ornack DR, Rakic P (2001) Cell proliferation without neurogenesis inadult primate neocortex. Science 294:2127–2130.
ulik A, Nakadate K, Nyiri G, Notomi T, Malitschek B, Bettler B,Shigemoto R (2002) Distinct localization of GABAB receptors rel-ative to synaptic sites in the rat cerebellum and ventrobasal thal-amus. Eur J Neurosci 15:291–307.
ulik A, Vida I, Luján R, Haas C, López-Bendito G, Shigemoto R,Frotscher M (2003) Subcellular localization of metabotropicGABAB receptor subunits GABAB1a/b and GABAB2 in the rat hip-pocampus. J Neurosci 23:11026–11035.
auder JM (1993) Neurotransmitters as growth regulatory signals: role ofreceptors and second messengers. Trends Neurosci 16:233–240.
aurie DJ, Wisden W, Seeburg PH (1992) The distribution of thirteenGABA A receptor subunit mRNAs in the rat brain: III. Embryonicand postnatal development. J Neurosci 12:4151–4172.
ee SH, Sheng M (2000) Development of neuron-neuron synapses.Curr Opin Neurobiol 10:125–131.
ois C, Alvarez-Buylla A (1994) Long-distance neuronal migration inthe adult mammalian brain. Science 264:1145–1148.
ópez-Bendito G, Shigemoto R, Luján R, Juiz JM (2001) Developmen-tal changes in the localisation of the mGluR1� subtype of metabo-tropic glutamate receptors in Purkinje cells. Neuroscience105:413–429.
ópez-Bendito G, Shigemoto R, Fairén A, Luján R (2002a) Differentialdistribution of group I metabotropic glutamate receptors during ratcortical development. Cereb Cortex 12:625–638.
ópez-Bendito G, Shigemoto R, Kulik A, Paulsen O, Fairén A, Luján R(2002b) Expression and distribution of metabotropic GABA recep-tor subtypes GABA R1 and GABA R2 during rat neocortical de-
B Bvelopment. Eur J Neurosci 15:1766–1778.
ópez-Bendito G, Luján R, Shigemoto R, Ganter P, Paulsen O, MolnárZ (2003): Blockade of GABAB receptors alters the tangential mi-gration of cortical neurons. Cereb Cortex 13:932–942.
ópez-Bendito G, Sturgess K, Erdelyi F, Szabo G, Molnar Z, PaulsenO (2004a) Preferential origin and layer destination of GAD65-GFPcortical interneurons. Cereb Cortex 14:1122–1133.
ópez-Bendito G, Shigemoto R, Kulik A, Vida I, Fairén A, Luján R(2004b) Distribution of metabotropic GABAB receptor subunitsGABAB1a/b and GABAB2 in the rat hippocampus during prenataland postnatal development. Hippocampus 14:836–848.
oTurco JJ, Owens DF, Heath MJ, Davis MB, Kriegstein AR (1995)GABA and glutamate depolarize cortical progenitor cells and inhibitDNA synthesis. Neuron 15:1287–1298.
uján R, Nusser Z, Roberts JDB, Shigemoto R, Somogyi P (1996)Perisynaptic location of metabotropic glutamate receptors mGluR1and mGluR5 on dendrites and dendritic spines in the rat hippocam-pus. Eur J Neurosci 8:1488–1500.
uján R, Roberts JDB, Shigemoto R, Ohishi H, Somogyi P (1997)Differential plasma membrane distribution of metabotropic gluta-mate receptors mGluR1�, mGluR2 and mGluR5, relative to neu-rotransmitter release sites. J Chem Neuroanat 13:219–241.
uján R, Shigemoto R, Kulik A, Juiz JM (2004) Localisation of theGABAB receptor 1a/b subunit relative to glutamatergic synapses inthe dorsal cochlear nucleus of the rat. J Comp Neurol 475:36–46.
uk KC, Kennedy TE, Sadikot AF (2003) Glutamate promotes prolif-eration of striatal neuronal progenitors by an NMDA receptor-mediated mechanism. J Neurosci 23:2239–2250.
a W, Barker JL (1995) Complementary expressions of transcriptsencoding GAD67 and GABAA receptor alpha 4, beta 1, andgamma 1 subunits in the proliferative zone of the embryonic ratcentral nervous system. J Neurosci 15:2547–2560.
a W, Liu QY, Maric D, Sathanoori R, Chang YH, Barker JL (1998) BasicFGF-responsive telencephalic precursor cells express functionalGABA(A) receptor/Cl-channels in vitro. J Neurobiol 35:277–286.
acdonald RL, Olsen RW (1994) GABAA receptor channels. AnnuRev Neurosci 17:569–602.
aric D, Liu QY, Grant GM, Andreadis JD, Hu Q, Chang YH, BarkerJL, Joseph J, Stenger DA, Ma W (2000) Functional ionotropicglutamate receptors emerge during terminal cell division and earlyneuronal differentiation of rat neuroepithelial cells. J Neurosci Res61:652–662.
aric D, Liu QY, Maric I, Chaudry S, Chang YH, Smith SV, Sieghart W,Fritschy JM, Barker JL (2001) GABA expression dominates neu-ronal lineage progression in the embryonic rat neocortex and fa-cilitates neurite outgrowth via GABA(A) autoreceptor/Cl chan-nels. J Neurosci 21:2343–2360.
arı́n O, Rubenstein JL (2001) A long, remarkable journey: tangentialmigration in the telencephalon. Nat Rev Neurosci 2:780–790.
artin LJ, Furuta A, Blackstone CD (1998) AMPA receptor protein indeveloping rat brain: glutamate receptor-1 expression and local-ization change at regional, cellular, and subcellular levels withmaturation. Neuroscience 83:917–928.
artin SC, Steiger JL, Gravielle MC, Lyons HR, Russek SJ, Farb DH(2004) Differential expression of gamma-aminobutyric acid type Breceptor subunit mRNAs in the developing nervous system andreceptor coupling to adenylyl cyclase in embryonic neurons.J Comp Neurol 473:16–29.
artı́nez-Galán JR, López-Bendito G, Lujan R, Shigemoto R, FairenA, Valdeolmillos M (2001) Cajal-Retzius cells in early postnatalmouse cortex selectively express functional metabotropic gluta-mate receptors. Eur J Neurosci 13:1147–1154.
arty S, Berninger B, Carroll P, Thoenen H (1996) GABAergic stim-ulation regulates the phenotype of hippocampal interneuronsthrough the regulation of brain-derived neurotrophic factor. Neuron16:565–570.
attson MP, Lee RE, Adams ME, Guthrie PB, Kater SB (1988) Inter-
actions between entorhinal axons and target hippocampal
M
M
M
M
M
M
M
M
N
N
N
N
N
N
O
P
P
P
P
P
P
R
R
R
R
S
S
S
S
S
S
S
S
S
S
T
T
T
V
W
R. Luján et al. / Neuroscience 130 (2005) 567–580 579
neurons: a role for glutamate in the development of hippocampalcircuitry. Neuron 1:865–876.
eier J (2003) The enigma of transmitter-selective receptor accumu-lation at developing inhibitory synapses. Cell Tissue Res311:271–276.
etin C, Denizot JP, Ropert N (2000) Intermediate zone cells expresscalcium-permeable AMPA receptors and establish close contactwith growing axons. J Neurosci 20:696–708.
ienville JM, Pesold C (1999) Low resting potential and postnatalupregulation of NMDA receptors may cause Cajal-Retzius celldeath. J Neurosci 19:1636–1646.
inakami R, Iida K, Hirakawa N, Sugiyama H (1995) The expressionof two splice variants of metabotropic glutamate receptor subtype5 in the rat brain and neuronal cells during development. J Neu-rochem 65:1536–1542.
iranda-Contreras L, Mendoza-Briceno RV, Palacios-Pru EL (1998)Levels of monoamine and amino acid neurotransmitters in thedeveloping male mouse hypothalamus and in histotypic hypotha-lamic cultures. Int J Dev Neurosci 16:403–412.
iranda-Contreras L, Benitez-Diaz PR, Mendoza-Briceno RV, Del-gado-Saez MC, Palacios-Pru EL (1999) Levels of amino acidneurotransmitters during mouse cerebellar neurogenesis and inhistiotypic hypothalamic cultures. Dev Neurosci 12:147–158.
isgeld U, Bijak M, Jarolimek W (1995) A physiological role forGABAB receptors and the effects of baclofen in the mammaliancentral nervous system. Prog Neurobiol 46:423–462.
iskevich F, Lu W, Lin SY, Constantine-Paton M (2002) Interactionbetween metabotropic and NMDA subtypes of glutamate receptorsin sprout suppression at young synapses. J Neurosci 22:226–238.
ery S, Fishell G, Corbin JG (2002) The caudal ganglionic eminenceis a source of distinct cortical and subcortical cell populations. NatNeurosci 5:1279–1287.
guyen L, Rigo JM, Rocher V, Belachew S, Malgrange B, Rogister B,Leprince P, Moonen G (2001) Neurotransmitters as early signalsfor central nervous system development. Cell Tissue Res305:187–202.
usser Z, Roberts JD, Baude A, Richards JG, Somogyi P (1995)Relative densities of synaptic and extrasynaptic GABAA receptorson cerebellar granule cells as determined by a quantitative immu-nogold method. J Neurosci 15:2948–2960.
usser Z, Sieghart W, Stephenson FA, Somogyi P (1996) The alpha 6subunit of the GABAA receptor is concentrated in both inhibitory andexcitatory synapses on cerebellar granule cells. J Neurosci16:103–114.
usser Z, Sieghart W, Somogyi P (1998a) Segregation of differentGABAA receptors to synaptic and extrasynaptic membranes ofcerebellar granule cells. J Neurosci 18:1693–1703.
usser Z, Lujan R, Laube G, Roberts JD, Molnar E, Somogyi P(1998b) Cell type and pathway dependence of synaptic AMPAreceptor number and variability in the hippocampus. Neuron21:545–559.
wens DF, Kriegstein AR (2002) Is there more to GABA than synapticinhibition? Nat Rev Neurosci 3:715–727.
anzanelli P, López-Bendito G, Luján R, Sassoe-Pogneto M (2004)Localization and developmental expression of GABAB receptors inthe rat olfactory bulb. J Neurocytol 33:87–99.
arnavelas JG (2000) The origin and migration of cortical neurones:new vistas. Trends Neurosci 23:126–131.
earce IA, Cambray-Deakin MA, Burgoyne RD (1987) Glutamateacting on NMDA receptors stimulates neurite outgrowth from cer-ebellar granule cells. FEBS Lett 223:143–147.
etralia RS, Esteban JA, Wang YX, Partridge JG, Zhao HM, WentholdRJ, Malinow R (1999) Selective acquisition of AMPA receptorsover postnatal development suggests a molecular basis for silentsynapses. Nat Neurosci 2:31–36.
in JP, Duvoisin R (1995) The metabotropic glutamate receptors:
structure and functions. Neuropharmacology 34:1–26.oluch S, Drian MJ, Durand M, Astier C, Benyamin Y, Konig N (2001)AMPA receptor activation leads to neurite retraction in tangentiallymigrating neurons in the intermediate zone of the embryonic ratneocortex. J Neurosci Res 63:35–44.
acca C, Stephenson FA, Streit P, Roberts JD, Somogyi P (2000)NMDA receptor content of synapses in stratum radiatum of thehippocampal CA1 area. J Neurosci 20:2512–2522.
omano C, van den Pol AN, O’Malley KL (1996) Enhanced earlydevelopmental expression of the metabotropic glutamate receptormGluR5 in rat brain: protein, mRNA splice variants, and regionaldistribution. J Comp Neurol 367:403–412.
omano C, Smout S, Miller JK, O’Malley KL (2002) Developmentalregulation of metabotropic glutamate receptor 5b protein in rodentbrain. Neuroscience 111:693–698.
ossi DJ, Slater NT (1993) The developmental onset of NMDA recep-tor-channel activity during neuronal migration. Neuropharmacol-ogy 32:1239–1248.
chwartz TH, Rabinowitz D, Unni V, Kumar VS, Smetters DK, TsiolaA, Yuste R (1998) Networks of coactive neurons in developinglayer 1. Neuron 20:541–552.
chweizer C, Balsiger S, Bluethmann H, Mansuy IM, Fritschy JM,Mohler H, Luscher B (2003) The gamma 2 subunit of GABA(A)receptors is required for maintenance of receptors at mature syn-apses. Mol Cell Neurosci 24:442–450.
erafini R, Maric D, Maric I, Ma W, Fritschy JM, Zhang L, Barker JL(1998) Dominant GABA(A) receptor/Cl channel kinetics correlatewith the relative expressions of alpha2, alpha3, alpha5 and beta3subunits in embryonic rat neurones. Eur J Neurosci 10:334–349.
higemoto R, Nakanishi S, Mizuno N (1992) Distribution of the mRNAfor a metabotropic glutamate receptor (mGluR1) in the centralnervous system: an in situ hybridization study in adult and devel-oping rat. J Comp Neurol 322:121–135.
higemoto R, Kulik A, Roberts JD, Ohishi H, Nusser Z, Kaneko T,Somogyi P (1996) Target-cell-specific concentration of a metabo-tropic glutamate receptor in the presynaptic active zone. Nature381:523–525.
higemoto R, Kinoshita A, Wada E, Nomura S, Ohishi H, Takada M,Flor PJ, Neki A, Abe T, Nakanishi S, Mizuno N (1997) Differentialpresynaptic localization of metabotropic glutamate receptor sub-types in the rat hippocampus. J Neurosci 17:7503–7522.
omogyi P, Tamas G, Lujan R, Buhl EH (1998) Salient features ofsynaptic organisation in the cerebral cortex. Brain Res Brain ResRev 26:113–135.
omogyi P, Dalezios Y, Luján R, Roberts JDB, Watanabe M,Shigemoto R (2003) High level of mGluR7 in the presynaptic activezones of select populations of GABAergic terminals innervating inter-neurons in the rat hippocampus. Eur J Neurosci 17:2503–2520.
oria JM, Valdeolmillos M (2002) Receptor-activated calcium signalsin tangentially migrating cortical cells. Cereb Cortex 12:831–839.
un L, Margolis FL, Shipley MT, Lidow MS (1998) Identification of along variant of mRNA encoding the NR3 subunit of the NMDAreceptor: its regional distribution and developmental expression inthe rat brain. FEBS Lett 441:392–396.
akai H, Katayama K, Uetsuka K, Nakayama H, Doi K (2003) Distri-bution of N-methyl-D-aspartate receptors (NMDARs) in the devel-oping rat brain. Exp Mol Pathol 75:89–94.
amaru Y, Nomura S, Mizuno N, Shigemoto R (2001) Distribution ofmetabotropic glutamate receptor mGluR3 in the mouse CNS: dif-ferential location relative to pre- and postsynaptic sites. Neuro-science 106:481–503.
issir F, Goffinet AM (2003) Reelin and brain development. Nat RevNeurosci 4:496–505.
erkhratsky A, Steinhauser C (2000) Ion channels in glial cells. BrainRes Brain Res Rev 32:380–412.
ang DD, Krueger DD, Bordey A (2003) GABA depolarizes neuronalprogenitors of the postnatal subventricular zone via GABAA recep-
tor activation. J Physiol 550:785–800.
W
W
W
Z
R. Luján et al. / Neuroscience 130 (2005) 567–580580
alsh CA, Goffinet AM (2000) Potential mechanisms of mutations thataffect neuronal migration in man and mouse. Curr Opin Genet Dev10:270–274.
atanabe M, Inoue Y, Sakimura K, Mishina M (1993) Distinct spatio-temporal distributions of the NMDA receptor channel subunit
mRNAs in the brain. Ann NY Acad Sci 707:463–466.ilson MT, Kisaalita WS, Keith CH (2000) Glutamate-inducedchanges in the pattern of hippocampal dendrite outgrowth: a rolefor calcium-dependent pathways and the microtubule cytoskele-ton. J Neurobiol 43:159–172.
hang LI, Poo MM (2001) Electrical activity and development of neural
circuits. Nat Neurosci 4:1207–1214.(Accepted 23 September 2004)(Available online 11 November 2004)

![Glutamate carboxypeptidase II gene knockout attenuates ... · metabotropic glutamate receptor (mGluR3) [- 7–9]. Acti vating mGluR3 by NAAG reduces the synaptic glutamate ... (Leica](https://img.dokumen.tips/doc/110x75/5c4d740293f3c34aee567cc7/glutamate-carboxypeptidase-ii-gene-knockout-attenuates-metabotropic-glutamate.jpg)



























