Embed Size (px)
Citation preview

This article is protected by copyright. All rights reserved
Relationships between stomatal behavior, xylem vulnerability to cavitation
and leaf water relations in two cultivars of Vitis vinifera
Sergio Tombesia,*, Andrea Nardinib, Daniela Farinellia and Alberto Palliottia
aDepartment of Agricultural and Environmental Sciences, University of Perugia, Borgo XX giugno
74, 06121 Perugia, Italy bDepartment of Life Sciences, University of Trieste, Via L. Giorgieri 10, 34127 Trieste, Italy
*Corresponding author, e-mail: [email protected]
Current understanding of physiological mechanisms governing stomatal behavior under water stress
conditions is still incomplete and controversial. It has been proposed that coordination of stomatal
kinetics with xylem vulnerability to cavitation (VC) leads to different levels of isohydry/anisohydry in
different plant species/cultivars. In this study, this hypothesis is tested in Vitis vinifera cultivars
displaying contrasting stomatal behavior under drought stress. The cv Montepulciano (MP, near-
isohydric) and Sangiovese (SG, anisohydric) were compared in terms of stomatal response to leaf and
stem water potential, as eventually correlated to different petiole hydraulic conductivity (kpetiole) and
VC, as well as to leaf water relations parameters. MP leaves showed almost complete stomatal closure
at higher leaf and stem water potentials than SG leaves. Moreover, MP petioles had higher maximum
kpetiole and were more vulnerable to cavitation than SG. Water potential at the turgor loss point was
higher in MP than in SG. In SG, the percentage reduction of stomatal conductance (PLgs) under water
stress was almost linearly correlated with corresponding percentage loss of kpetiole (PLC), while in MP
PLgs was less influenced by PLC. Our results suggest that V. vinifera near-isohydric and anisohydric
genotypes differ in terms of xylem vulnerability to cavitation as well as in terms of kpetiole, and that the
coordination of these traits leads to their different stomatal responses under water stress conditions.
Abbreviations – gs, stomatal conductance; Kmax, maximum hydraulic conductance; kpetiole, petiole
hydraulic conductivity; MP, Montepulciano; PLC, percentage loss of hydraulic conductance; PLgs,
percentage reduction of stomatal conductance; RWC, relative water content; RWCtlp, relative water
content at turgor loss point; SG, Sangiovese; VC, vulnerability curves; ε, leaf modulus of elasticity;
πo, osmotic potential at full turgor; πtlp, turgor loss point; Ψleaf, leaf water potential.
This article has been accepted for publication and undergone full peer review but has not been through the copyediting, typesetting, pagination and proofreading process, which may lead to differences between this version and the Version of Record. Please cite this article as doi: 10.1111/ppl.12180

This article is protected by copyright. All rights reserved
Introduction
Plant-level gas exchange rates, photosynthesis, productivity and survival, ultimately depend on leaf-
level stomatal regulation of carbon gain versus water loss (Jones 1998). Under water stress conditions,
the onset of partial or total stomatal closure prevents excessive tissue dehydration and protects the
plant hydraulic system from runaway cavitation and catastrophic hydraulic failure that might occur
because of uncontrolled decline of leaf and stem water potential (Tyree and Sperry 1988). The ability
of different species to maintain a positive carbon balance under drought stress conditions is largely a
consequence of their different stomatal response to declining leaf water potentials (Chaves et al.
2010). On the basis of stomatal kinetics under water stress conditions, plants are generally classified
in two categories: isohydric species, which prevent water potential drop by means of early stomatal
closure, and anisohydric species, which maximize photosynthetic gain by keeping stomata open
despite larger water potential drops (Tardieu and Simonneau 1998). As an example, Vitis vinifera
comprises cultivars classified as near-isohydric, while other cultivars are known to be largely
anisohydric (Schultz 2003, Medrano et al. 2003, Poni et al. 2007, Palliotti et al. 2009, Pou et al. 2012),
although the classification into one or the other category is somehow difficult and uncertain because
of different experimental conditions and techniques adopted in different studies (Chavez et al. 2010).
The mechanism(s) responsible for stomatal closure under water stress are still controversial and
debated (Comstock 2002). Some studies have suggested that chemical signals are the main factor
driving stomatal closure, and further hypothesized that differential species-specific sensitivities to
[ABA] changes are at the basis of isohydric vs anisohydric behavior (Davies and Zang 1991, Tardieau
and Simonneau 1998, Lovisolo et al. 2002). On the other hand, stomatal closure has been reported to
be correlated to changes in turgor pressure of both epidermal and mesophyll cells (Cowan 1977,
Syvertsen 1982, Ache et al. 2010). Turgor loss is a critical outcome of water stress, and
osmoregulation leading to lower turgor loss point (Ψtlp) is generally regarded as an important strategy
adopted by plants to improve drought tolerance (Bartlett et al. 2012). In fact, plant species that
maintain positive turgor pressure at very low leaf water potential are also known to keep high stomatal
conductance, gas exchange and growth rates at relatively lower soil water potential than species
characterized by less negative Ψtlp values (Abrams and Kubiske 1990, Sack et al. 2003, Baltzer et al.
2008, Mitchell et al. 2008, Blackman et al. 2010).
Stomatal conductance to water vapor has been reported to be coordinated with liquid phase
hydraulic conductance of the soil-to-leaf pathway (Saliendra et al. 1995, Comstock 2002, Meinzer
2002), suggesting a possible feedback link between stomatal regulation and hydraulic signals (Nardini
and Salleo 2000, Franks 2004). Stomatal control of transpiration is known to be involved in xylem
cavitation avoidance mechanisms (Jones and Sutherland 1991, Tyree and Sperry 1989). Salleo et al.
(2000) have reported that the onset of partial stomatal closure in transpiring Laurel plants occurs when
stem water potential approaches the cavitation threshold, as revealed by concurrent production of

This article is protected by copyright. All rights reserved
ultrasound acoustic emissions at the stem and leaf level (Tyree and Dixon 1983).
Xylem hydraulic efficiency and vulnerability to cavitation-induced embolism are correlated to
several anatomical traits, including xylem conduit dimensions and arrangement, as well as to pit
membrane features (Sperry and Saliendra 1994, Lo Gullo et al. 1995, Choat et al. 2008, Lens et al.
2011). Previous studies have reported differences in terms of stem hydraulic conductance among
different genotypes of V. vinifera. In particular, Schultz (2003) has suggested that higher stem specific
conductivity was at the basis of the isohydric behavior of the cultivar Grenache when compared to the
anisohydric Syrah. Much less is known about the eventual relationships between xylem vulnerability
to cavitation and the relative level of isohydry of different grape cultivars, although it has been
suggested that contrasting hydraulic vulnerability to cavitation of different genotypes might also be an
important determinant of their different stomatal responses to declining water potential (Schultz
2003). Alsina et al. (2007) have reported large variability of vulnerability to cavitation across eight
different grape cultivars, with values of leaf water potential inducing 50% loss of hydraulic
conductance (Ψ50) ranging from –1 MPa in cv Parellada to about –3 MPa in cv Sauvignon Blanc.
However, xylem vulnerability was not correlated to drought tolerance mechanisms at the leaf level as
estimated on the basis of water relation parameters derived on water potential isotherms, in contrast
with data gathered from inter-specific comparisons (Vilagrosa et al. 2010). Overall, the above studies
did not provide clear-cut evidence about eventual correlations between vulnerability to cavitation and
stomatal regulation in grape.
In the present study, two grapevine cultivars were investigated for their stomatal and hydraulic
responses to drought stress. In particular, we selected two genotypes known for their contrasting
stomatal behavior i.e. the cv Sangiovese which is anisohydric, and the cv Montepulciano which
displays a near-isohydric behavior (Palliotti et al. 2009). The two cultivars are known to differ from
each other also in terms of xylem anatomy and other functional properties linked to photosynthetic
efficiency and energy dissipation potential like chlorophyll and carotenoids contents (Palliotti et al.
2008, 2011). Our aim was to identify the eventual relationships and mechanistic link between stomatal
behavior and xylem vulnerability to cavitation, as well as eventual coordination of the relative level of
isohydry/anisohydry to water relation parameters correlated to drought resistance.
Materials and methods
Plant material
This study was conducted during 2012 on 7-year-old potted V. vinifera vines of cultivar Sangiovese
(clone VCR30) and cultivar Montepulciano (clone R7), both grafted onto 1103 Paulsen rootstock and
grown in an outdoor area close to the Faculty of Agriculture of the University of Perugia (Region of
Umbria, central Italy, 42°58’N, 12°24’E, elevation 405 m a.s.l.). Pots (60 liters volume) were filled
with loam soil having field capacity and permanent wilting point water contents of 30.2 and 16.7%
(v/v), respectively. At the end of February, each vine was pruned to retain four spurs with two buds

This article is protected by copyright. All rights reserved
each. All shoots were oriented upright using suitable stakes. Ten vines per cultivar were used and
initially maintained at ~90% of maximum soil water availability. Daily water supply per pot was
determined by monitoring the soil water content with a Diviner 2000 capacitance probe (Sentek
Environ Tech., Sentek Environment Technologies, Stepney, South Australia), using access tubes
located in three pots per cultivar. Measurements were performed at 100 mm, 170 mm and 250 mm of
depth from the soil surface in the pot. Water was added in the required amounts every day at 8 pm.
Additional experiments were performed in 2013 on the same vines used in 2012. Vines were kept
irrigated to field capacity until 7 July, when drought was imposed by suspending irrigation and
closing pots into plastic bags. The drought treatment was continued until complete leaf abscission.
Stomatal conductance vs water potential
In 2012, stomatal conductance (gs) measurements were carried out on adult, non senescent, primary
leaves inserted between the 5th and the 10th node from the cane base. Measurements were performed
under saturating light conditions (PPFD > 1200 µmol photons m–2 s–1) using an open gas exchange
system (ADC-System, LCA-3, Hoddesdon, UK) equipped with a Parkinson leaf chamber.
Measurements were repeated during three consecutive sunny days (17, 18 and 19 July 2012) between
08.00 and 10.00 a.m. Two canes per vine, sampled from five different vines per cultivar, were
harvested at 08.00 by cutting the stem under water. Canes were transported to the laboratory and
allowed to dehydrate in the light. At different time intervals, leaves were measured for gs and
immediately detached to record their water potential using a pressure chamber (Soilmoisture Corp,
Santa Barbara, CA, USA).
In 2013, stomatal conductance (gs) measurements were carried out on adult, non senescent,
primary leaves grown between the 5th and the 10th node from the cane base in order to assess if
measurements of gs carried out on detached shoots were eventually biased by shoot cutting procedure.
Measurements were carried out between 12.00 a.m. and 1.00 p.m. from 8 July up to 16 July on 5 vines
per cultivar using the same equipment described above. Leaves were measured for gs and immediately
detached and wrapped in plastic film to record their water potential (Ψleaf) using a pressure chamber
(Soilmoisture Corp, Santa Barbara, CA, USA). Stem xylem water potential (Ψstem) was measured on
mature leaves that had been wrapped in plastic film and aluminum foil prior to the measurements
(McCutchen and Shackel 1992).
Xylem anatomy
In August 2012, three petioles per vine were sampled from five vines per cultivar. Petioles were
sampled from leaves grown between the 5th and the 10th node from the cane base. The basal portion
(~20–30 mm) of petioles was then sampled and immediately placed in ice and refrigerated until
sectioned. Samples were embedded in an agar solution (0.2%) and cross sections (four per sample) 20
µm thick were obtained using a microtome (2700 Frigocut, Reichert-Jung, Nossloch, Germany).

This article is protected by copyright. All rights reserved
Sections were stained with Iodine green to increase the contrast. Photographs of the cross-sections
were taken with a Leica ICCA camera (Leica Microsystems Wetzlar GmbH, Wetzlar, Germany)
coupled to a Leica DMLB light microscope (Leica Microsystems Wetzlar GmbH). Images were
acquired with Leica IM1000 software (Leica Microsystems Digital Imaging, Cambridge, UK). Two
photographs were taken from each cross-section at 200× magnification. Measurements of vessel
diameters were taken using SIGMASCAN PRO 5.0 software (Systat Software Inc., San Jose, CA, USA)
(Palliotti et al. 2011). Mean diameter distributions of vessels were then reported in frequency classes
of 5 µm expressed as percentage of the total number of vessels.
The theoretical hydraulic conductance (kh) (kg m MPa–1 s–1) was calculated according to the
modified Hagen-Poiseuille law as reported by Tombesi et al. (2010):
128
where d is the radius of the vessel (m), obtained from the mean of the largest and smallest radii, ρ is
the fluid density (assumed to be 1000 kg×m3 or equal to that of water at 20°C) and η is the viscosity
(assumed to be 1×10–9 MPa s–1, or equal to that of water at 20°C).
Vulnerability curves
Vulnerability curves were constructed using the double-ended pressure sleeve method described by
Cochard et al. (1992) and Ennajeh et al. (2011). A custom built apparatus was used, consisting of a T
steel fitting (50 mm long) that was used as sleeve, and two compression fittings at the two ends of the
sleeve. Rubber corks were used to seal the sleeve. Internal distance between corks when the sleeve
was sealed was ~ 4 mm. Measurements were performed in August 2012 on samples collected between
8.00 and 13.00. Minimum length of petioles used in this study was 11 cm i.e. substantially longer than
the longest xylem vessel in petioles of V. vinifera (Chatelet et al. 2011). Hydraulic conductance of
petioles was measured by connecting one sample end to a vertical plastic tubing filled with a filtered
(0.2 µm) 20 mM KCl solution. The pressure head was maintained at 6 kPa. Water flow through the
petioles was measured continuously while the air pressure inside the chamber was gradually
increased. Flow was measured gravimetrically by collecting the fluid from the distal end in pre-
weighted sponge pieces fitted in plastic tubes. Flow readings were taken over 1 min time intervals.
At the beginning of each experiment, the petioles were flushed at P = 0.2 MPa for 30 min, to
remove any eventual native embolism. After flushing samples, maximum hydraulic conductance
(Kmax) was measured. Then, the pressure in the chamber was progressively increased to 0.3, 0.5, 0.8,
1.0, 1.3, 1.5, 1.8, 2.0, 2.3 and 2.5 MPa. Sample hydraulic conductance (K) was measured at steady
state (10 min after the desired pressure value was reached) at each pressure step. Percentage loss of
hydraulic conductance (PLC) at each pressure value was calculated as:

This article is protected by copyright. All rights reserved
PLC = 100 × (Kmax–K)/Kmax
The leaf specific petiole hydraulic conductivity (kpetiole) was finally estimated according to Zufferey et
al. (2011) as:
kpetiole = (Kmax×L)/Aleaf
where L is petiole length and Aleaf is leaf blade surface area.
Relationship between percent loss of gs (PLgs) and PLC was obtained by interpolating gs value
from the regressions between gs and Ψstem for Ψstem values at which PLC values were measured. PLgs
representing as percentage of the maximum gs of the curve.
Pressure-volume curves
Leaf water potential isotherms, commonly known as pressure-volume (P-V) curves, were measured in
August 2012 on ten mature leaves per cultivar (two leaves from each of five different vines). Leaves
were harvested at 06.00 a.m. and petioles were kept immersed in deionized water for 2 h to allow full
rehydration. Leaves with Ψleaf < –0.05 MPa after rehydration were immediately discarded. Each leaf
was weighted using an analytic balance (OHAUS PA64C, OHAUS Corp., USA) and water potential
was immediately measured by a pressure chamber. Leaves were progressively dehydrated on the
bench and successive measurements of leaf weight and water potential were taken. Turgor loss point
(πtlp) and relative water content (RWC) at turgor loss point (RWCtlp) of each leaf was calculated as the
intersection point of the two phase linear equation representing the relationship between leaf RWC
and Ψleaf (Brodribb and Holbrook 2003). Leaf modulus of elasticity (ε) and the osmotic potential at
full rehydration (πo) were respectively calculated as the slope of the pressure potential curve between
100–RWC=0 and πtlp and the y-intercept of the solute potential curve (see Bartlett et al. 2012).
Differences between genotypes were assessed using the Student’s t-test (P<0.05). The
significance of regressions was tested using Pearson Product Moment Correlation. Test of
homogeneity of regression slope was used to assess if the slope of correlations was similar, an
ANCOVA was performed in order to test whether the null hypothesis was satisfied.
Results
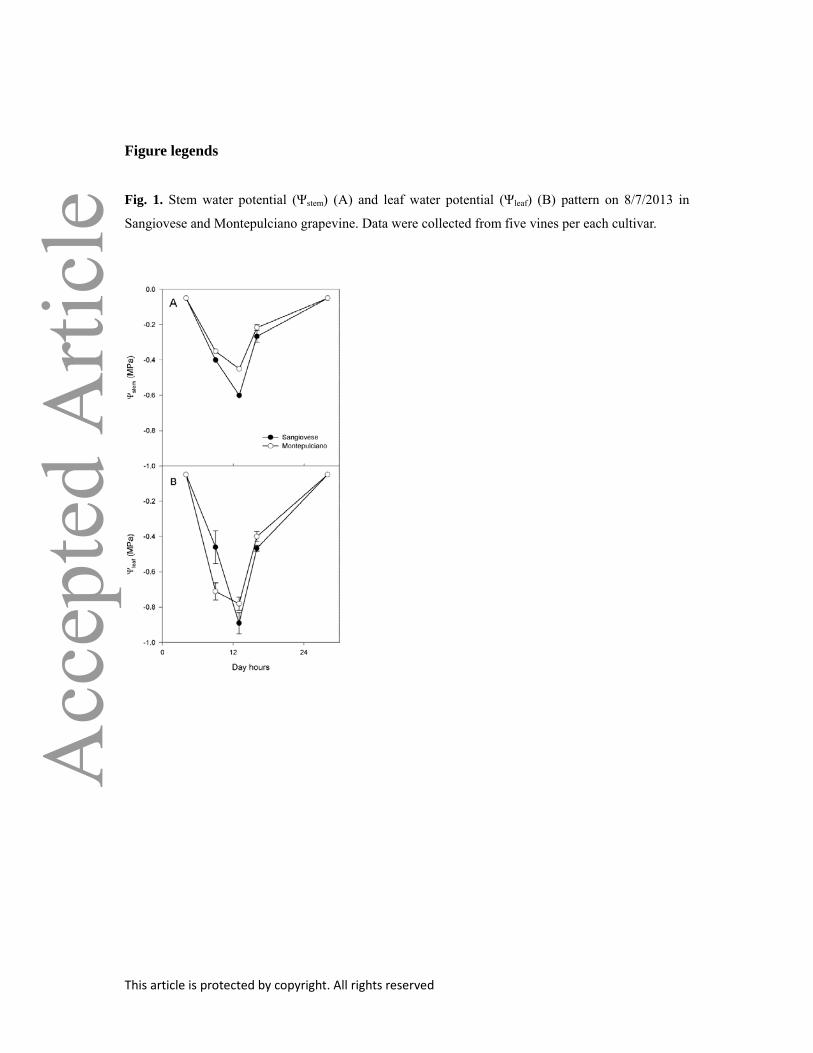
While predawn water potential was similar in Sangiovese and Montepulciano, stem water potential at
midday was more negative in Sangiovese than in Montepulciano (Fig 1A). A similar pattern was
observed for leaf water potential (Ψleaf), which in both cultivars reached more negative values than
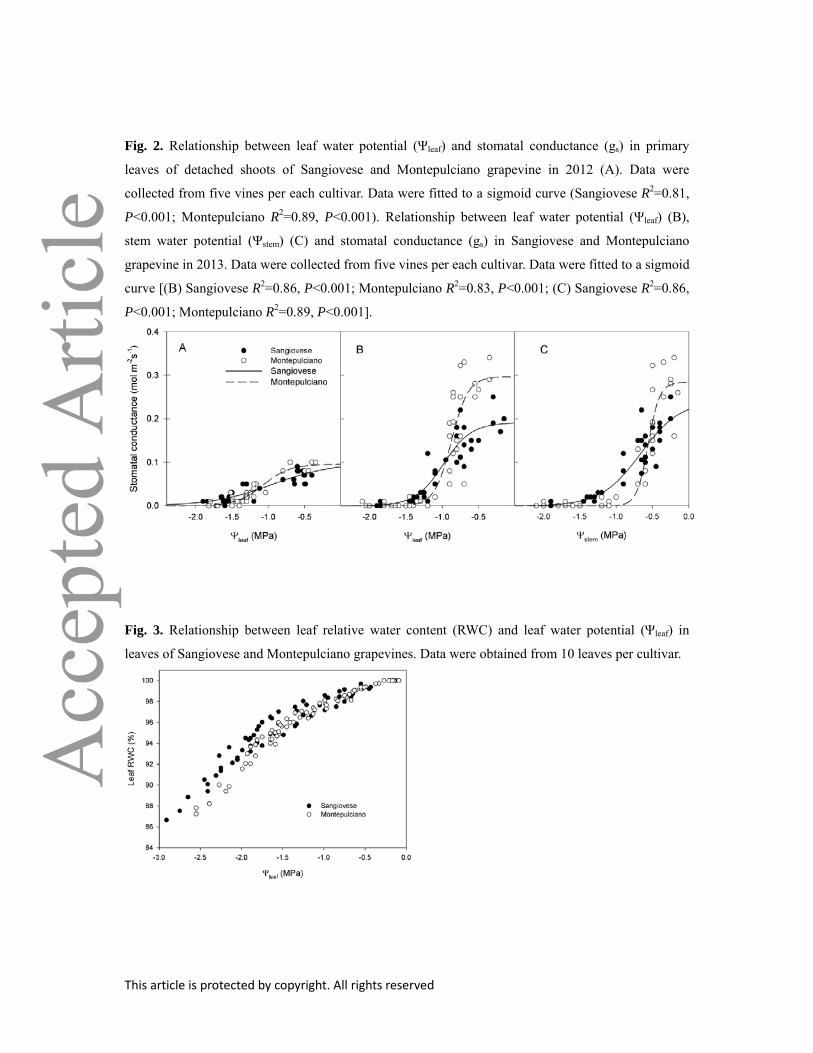
Ψstem (Fig 1B). In both genotypes, gs decreased as a function of declining Ψleaf and Ψstem, both when
measured on desiccating detached shoots or on whole drought-stressed plants (Fig. 2). The
relationship was significantly fitted by a sigmoid regression in all cases and the water potential

This article is protected by copyright. All rights reserved
inducing stomatal closure was independent on the method used. The two fitted curves were
significantly different depending on the genotype (test of homogeneity of regression slope P=0.0025,
P=0.005 and P=0.036 for relationships in fig 2A, 2B and 2C, respectively). However, stomatal
conductance measured on leaves from intact plants was larger than that measured on detached shoots.
Sangiovese had lower maximum gs than Montepulciano. Sangiovese also showed a very progressive
(almost linear) gs response to decreasing water potential and underwent complete stomatal closure at
Ψ = –2.0 MPa (Fig 2A, B and C). Montepulciano leaves displayed a sharp decrease of gs at Ψleaf < –
0.8 MPa and Ψstem < –0.5, and reached full stomatal closure at Ψleaf = –1.4 MPa and Ψstem = –1.0 MPa,
already.
Both πtlp and πo were significantly lower in Sangiovese than in Montepulciano (Table 1). On the
other hand, RWCtlp and ε were not significantly different in the two genotypes. Overall, the
relationship between RWC and Ψleaf was similar for the two cultivars in the water potential range
between 0 and –1.5 MPa. Below this threshold value, however, Sangiovese leaves retained higher
RWC than Montepulciano (Fig. 3) as Ψleaf continued to decline.
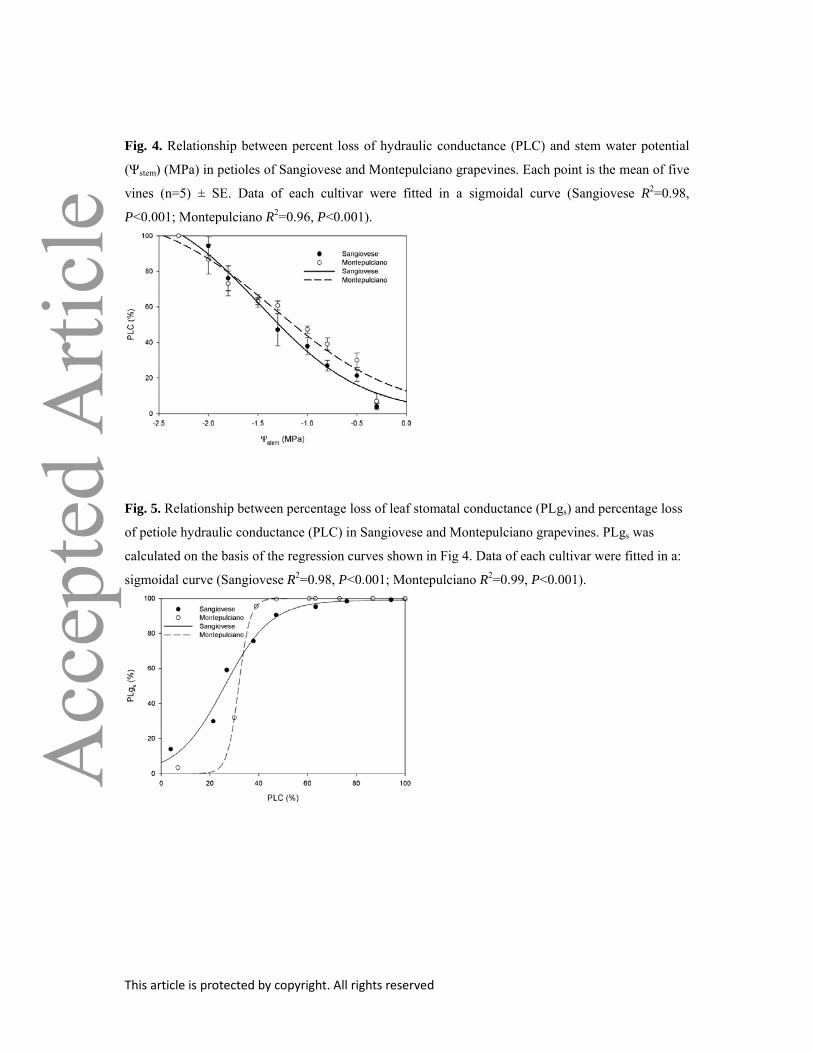
The two cultivars also displayed differential vulnerability to xylem embolism. Petiole PLC was
significant larger in Montepulciano in the P range from –0.5 to –1.3 MPa, with respect to Sangiovese
(Fig 4). Water potential values inducing 20% (P20) and 50% (P50) loss of hydraulic conductivity were
significantly lower in cv Sangiovese than in cv Montepulciano, while both cultivars reached 80% loss
of hydraulic conductivity at similar xylem pressures (Table 2). Leaf specific petiole hydraulic
conductivity (kpetiole) was approximately 40% higher in Montepulciano than in Sangiovese, averaging
about 7.1 and 5.0 mmol m–1 s–1 MPa–1, respectively (Table 2).
In both cultivars, PLgs was significantly correlated to PLC in a sigmoid fashion (Fig. 5).
However, cv Sangiovese had a higher gs sensitivity to PLC, as the PLgs to PLC ratio was close to 1
over the PLC range between 10 and 60%. On the other hand, the cv Montepulciano showed almost no
stomatal adjustment down to PLC of about 20%. After this PLC level, gs started to decline sharply and
PLgs approached 100% at PLC = 40%.
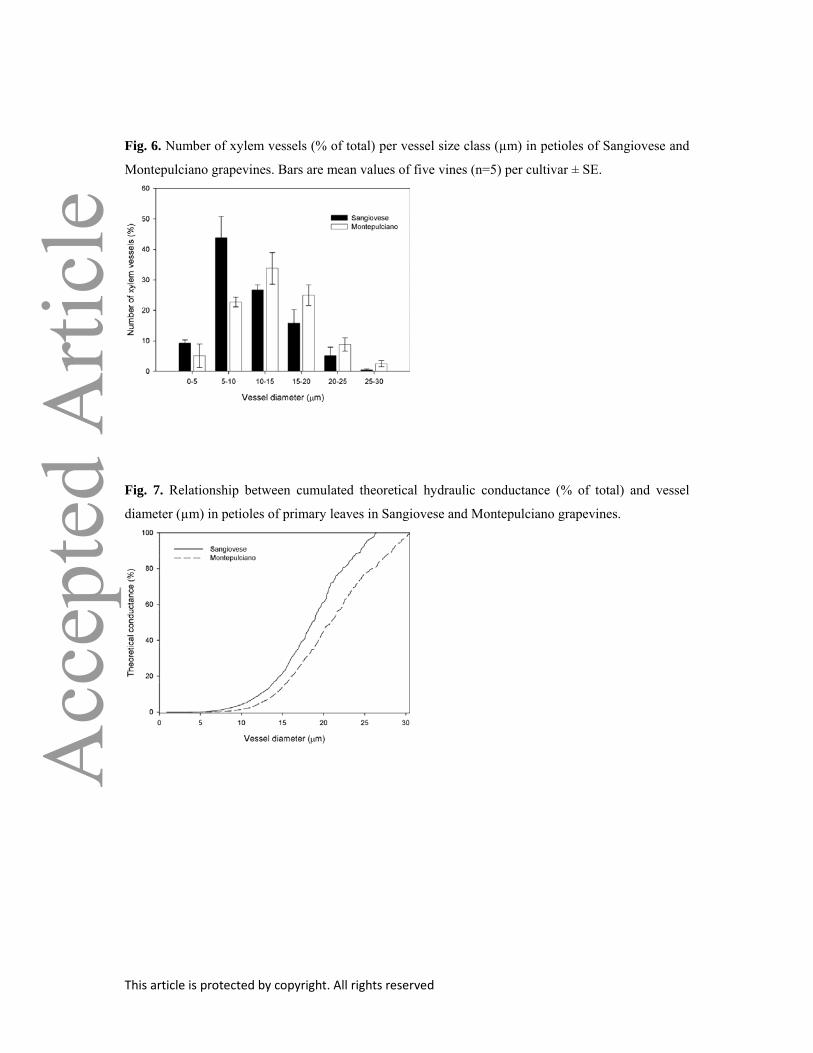
In Sangiovese petioles, xylem tissue had significantly more vessels in the 5–10 µm diameter
class when compared to Montepulciano, whereas this latter cultivar had significantly more vessels in
10–15 µm, 15–20 µm and 25–30 µm diameter classes than the former (Fig. 6). In Sangiovese, a larger
fraction of hydraulic conductance was apparently supported by smaller xylem conduits when
compared to Montepulciano (Fig. 7 and Table 2). About 50% of the theoretical xylem hydraulic
conductance was supported by xylem conduits smaller than 18 and 21 µm in Sangiovese and
Montepulciano, respectively (Table 2).
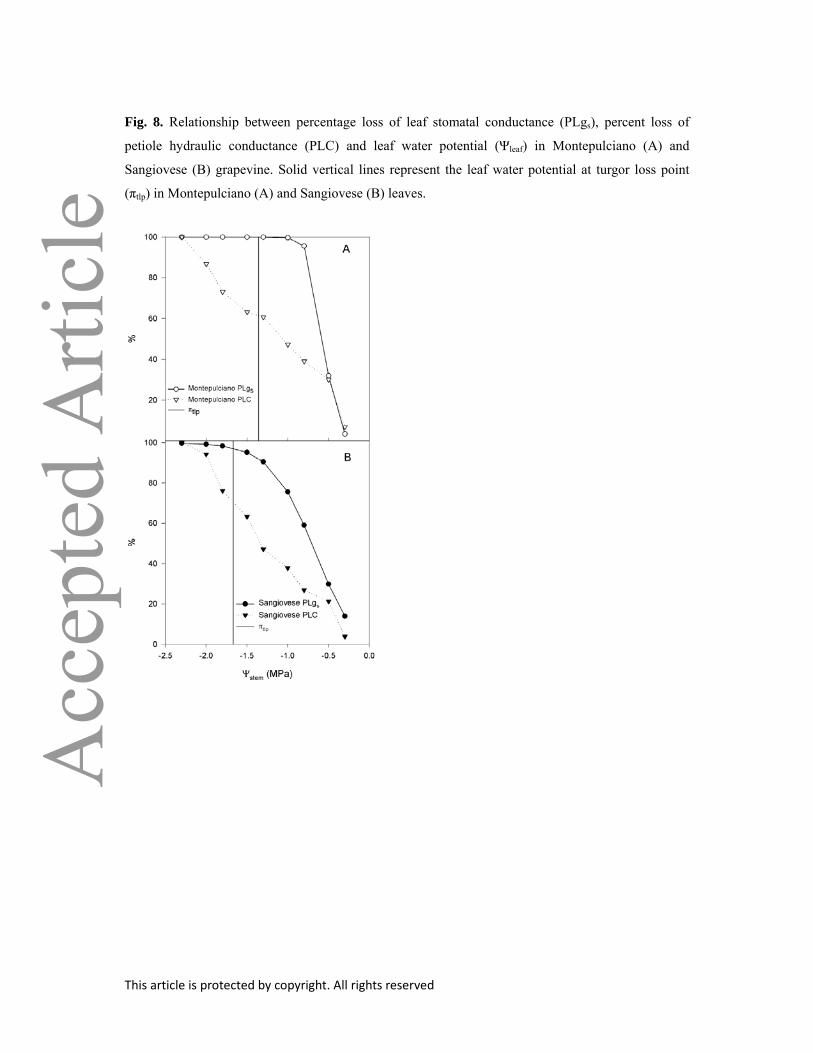
In both cultivars, PLgs was larger than PLC, but had reached similar values at Ψ = –2.3 MPa and
–2.0 in Montepulciano and Sangiovese, respectively (Fig. 8).
Discussion

This article is protected by copyright. All rights reserved
Montepulciano and Sangiovese vines displayed different levels of anisohydry. In both cultivars, Ψstem
declined during the central hours of the day but the decline was more marked in Sangiovese. This
different behavior can be attributed to different stomatal kinetics that resembled those previously
reported by Schultz (2003) for Grenache and Syrah cultivars, and were interpreted as evidence for
different levels of iso/anisohydry. Our results show that under water stress, Sangiovese reduces
stomatal conductance more gradually than Montepulciano and that, at relatively low water potentials
(< –1.5 MPa), it maintains higher gas exchange rates than Montepulciano. These results are in
accordance with previous data obtained on the same genotypes under field conditions (Palliotti et al.
2009). Furthermore, these results contribute to explain the different behavior observed in these same
cultivars under irrigation and after water stress acclimation (Palliotti et al. 2014). In our study, gs was
correlated both with Ψleaf and Ψstem. Stomatal closure was observed approximately at the same Ψleaf
and Ψstem values in both lab and field experiments, although the two curves differed in the central
region. In the field experiment carried out in 2013, gs values were higher than those recorded in the
experiment carried out in 2012 on excised sample, and this difference was probably due to the
severing of canes. However, the consistency of water potential values triggering stomatal closure in
the two experiments suggest that bench shoot dehydration could be a useful method for fast screening
of stomatal behavior across different grape genotypes.
Differently from other studies carried out on grapevine cultivars with different levels of
anisohydry (Tramontini et al. 2013, Hochberg et al. 2013), in our experiments the near-isohydric
cultivar Montepulciano had larger gs than Sangiovese. This is in accordance with the larger hydraulic
conductance of Montepulciano canes (Palliotti et al. 2011) and petioles in comparison with those of
Sangiovese. These results also differ from those reported by Beis and Patakas (2010) which did not
found any significant difference in plant hydraulic conductance in two grapevine cultivars with
different stomatal sensitivity to water stress. According to Schultz (2003), contrasting stomatal
behaviors among grapevine cultivars could be explained in terms of their different hydraulic
architecture. Our additional hypothesis was that differential vulnerability to cavitation might be at the
basis of contrasting stomatal kinetics under water stress conditions. In fact, xylem cavitation is a
hydraulic signal triggering stomatal closure (Salleo et al. 2000, Nardini et al. 2001). In turn, stomatal
regulation of transpiration limits stem and leaf water potential drop (Domec and Johnson 2012). The
regulation of water potential prevents further cavitation and embolism in the leaf and in the stem
(Tyree and Sperry 1988). Indeed, vulnerability curves of petioles, that could be considered as 'fuses' in
the hydraulic circuit (Zufferey et al. 2011), revealed that the near-isohydric cultivar Montepulciano
was more vulnerable to cavitation than the anisohydric one (Sangiovese), in agreement with Zufferey
et al. (2011) and Domec and Johnson (2012). Hence, our results apparently support the hypothesis of
coordination of stomatal closure with xylem vulnerability to cavitation, although only in Sangiovese
PLC was almost linearly correlated with PLgs, while the same relationship was weaker in
Montepulciano.

This article is protected by copyright. All rights reserved
Different stomatal behaviors as related to water potential drop and consequent triggering of
cavitation events can be also interpreted as a functional consequence of contrasting hydraulic anatomy
in different cultivars (Chouzouri and Schultz 2005). In fact, xylem vulnerability to cavitation and
embolism is dependent on xylem anatomy. As an example, mean cavitation pressure was correlated
with inter-vessel pit structure in different Acer species (Lens et al. 2011) and Cai and Tyree (2010)
demonstrated that in aspen, xylem vessels with large diameter suffered 50% PLC at xylem pressures
less negative than those necessary to embolize smaller conduits. As previously reported by some of us
(Palliotti et al. 2011), Sangiovese petioles had smaller xylem conduits than Montepulciano, in
accordance with relatively higher resistance to xylem embolism of the former cultivar with respect to
the latter. Moreover, in Sangiovese a larger fraction of total xylem hydraulic conductance was
apparently supported by smaller vessels.
Alternatively or in addition to cavitation-associated hydraulic signals, bulk leaf water status can
influence stomatal closure, which has been reported to be correlated or coordinated to the turgor loss
point (Cowan 1977). In the present study, πtlp was more negative in Sangiovese (anisohydric) than in
Montepulciano (near-isohydric) genotype, thus suggesting a possible coordination of this parameter
with stomatal behavior in response to bulk leaf water potential changes. In contrast with Beis and
Patakas (2010), in the present study we observed differences also in terms of osmotic potential (πo)
between the two cultivars, but no difference in terms of ε and RWCtlp. These results support the
hypothesis that also in Vitis vinifera genotypes, eventual differences in terms of πtlp are mainly driven
by osmotic adjustment and not by modifications of mechanical properties of cell walls, in agreement
with previous reports on several different species (Lenz et al. 2006, Baltzer et al. 2008, Mitchel et al.
2008, Bartlett et al. 2012). In both Sangiovese and Montepulciano, πtlp was reached when almost 90%
of stomatal conductance reduction or complete stomatal closure had occurred, already. Such results
are consistent with previous studies showing that initiation of stomatal closure is largely uncoupled
from bulk leaf water status (Mott and Franks 2001, Brodribb et al. 2003). On the other hand, in both
cultivars near-complete stomatal closure occurred at Ψleaf close to πtlp, similarly to observations
reported by Brodribb et al. (2003) in several different species and with data reported for walnut and
loblolly pine by Cochard et al. (2002) and Domec et al. (2009). Our data suggest that although πtlp is
not the main factor triggering partial stomatal closure, it is indeed correlated to the occurrence of
complete halt of leaf-level gas exchange.
Stomatal behavior in Montepulciano was apparently addressed at keeping high gas exchange
rates at moderately negative water potentials (Ψstem > –0.5), commonly reached under well-watered
conditions (fig 1). Moreover, Montepulciano petioles were more vulnerable to cavitation than
Sangiovese. If the stomatal kinetics were exclusively dominated by hydraulic signals, genotypes with
more vulnerable hydraulic apparatus would be expected to suffer gas exchange reduction even for
relatively small drops of leaf/stem water potential, as commonly experienced under non-stressful
conditions during the middle part of the day when irradiance and temperature are at their peaks.

This article is protected by copyright. All rights reserved
However, Montepulciano vines under well-watered conditions were able to transpire at faster rates
than Sangiovese (Palliotti et al. 2011). Under these conditions Ψleaf normally drops down to –0.8 MPa
(Zufferey et al. 2011) and Ψstem drops to –0.6 MPa, and, according to our data, PLC of Montepulciano
would be expected to be significantly higher than that of Sangiovese. According to the hypothesis of
complete coordination of stomatal behavior with xylem vulnerability to cavitation, stomatal
conductance should be lower in Montepulciano than in Sangiovese under these conditions. Our data
show an opposite behavior, suggesting that stomatal kinetics are not coordinated with xylem
vulnerability in the same way in all cultivars and at any level of water stress. This might arise as a
consequence of the fact that stomatal conductance is more closely related to actual values of kpetiole
rather than to relative loss of hydraulic conductivity. In fact, in accordance with previously reported
data on vine canes (Palliotti et al. 2011), Montepulciano had higher kpetiole than Sangiovese. As a
result, even at higher PLC in the former than in the latter cultivar, the actual water transport efficiency
per unit leaf surface area in the two cultivars might be similar. As an example, at Ψstem = –0.8 MPa, gs
was similar in the two cultivars, despite the fact that PLC of Montepulciano was higher (39%) than
that of Sangiovese (26%). However, taking into account the maximum kpetiole measured for the two
cultivars, at this water potential level it can be calculated that the actual kpetiole would still be slightly
higher in Montepulciano than in Sangiovese (4.6 vs 3.6 mmol m–1 s–1 MPa–1, respectively). This
would translate in similar water transport rates per unit leaf surface area and, hence, similar leaf
hydration and stomatal opening. However, in both cultivars stomatal closure occurred at Ψstem close to
those at which 50% of PLC occurs (P50). These results support the coordination of stomatal closure
with xylem vulnerability and the validity of P50 as an indicator of xylem hydraulic safety (Meinzer et
al. 2009, Choat et al. 2012).
In conclusion, different stomatal behaviors of Montepulciano and Sangiovese grapevines,
respectively near-isohydric and anisohydric, properly correlate to different levels of xylem
vulnerability to water stress-induced cavitation, provided actual values of stem/petiole hydraulic
conductivity are taken into account. Previous studies have analyzed many grapevine cultivars for their
contrasting stomatal behavior, and in several cases the same cultivar was classified as isohydric or
anisohydric by different studies (Chaves et al. 2010, Lovisolo et al. 2010). Experimental conditions
can be claimed as possible causes for such contradictory results. Our results point out that, depending
on the water potential range at which the experiment is carried out, the same cultivar can behave in
relatively opposite ways. Previous studies have proposed that cultivar-specific differences in stomatal
control in response to water stress arise as a result of differences in the ABA signaling and perception
machinery (Soar et al. 2006, Perrone et al. 2012a) and/or as a consequence of different patterns of
aquaporins expression and/or activation (Vandeleur et al.2009, Perrone et al.2012b, Pou et al 2013).
Although our data point to a mechanistic linkage between stomatal behavior and vulnerability to
cavitation, the possibility that other phenomena integrated hydraulic signals and triggered or mediated
stomatal closure cannot be ruled out.

This article is protected by copyright. All rights reserved
Author contributions – S.T., A.N. and A.P. conceived and planned the study. S.T. carried out the
experiment, analyzed the data and wrote the first draft of the manuscript. A.P. planted and managed
the vines that were used in the study, and reviewed the manuscript. A.N. revised and edited the
manuscript. A.P. and D.F. helped in the analysis of the data, revised and edited the manuscript, and
obtained funds to support the project.
References
Abrams MD, Kubiske ME (1990) Photosynthesis and water relations during drought in Acer rubrum
L genotypes from contrasting sites in central Pennsylvania. Funct Ecol 4: 727–733
Ache P, Bauer H, Kollist H, Al-Rasheid KA, Lautner S, Hartung W, Hedrich R (2010) Stomatal action
directly feeds back on leaf turgor: new insights into the regulation of the plant water status from
non-invasive pressure probe measurements. Plant J 62: 1072–1082
Alsina MM, de Herralde F, Aranda X, Save R, Biel C (2007) Water relations and vulnerability to
embolism are not related: experiments with eight grapevine cultivars. Vitis 46: 1–6
Baltzer JL, Davies SJ, Bunyavejchewin S, Noor NSM (2008) The role of desiccation tolerance in
determining tree species distributions along the Malay-Thai Peninsula. Funct Ecol 22: 221–231
Bartlett MK, Scoffoni C, Sack L (2012) The determinants of leaf turgor loss point and prediction of
drought tolerance of species and biomes: a global meta-analysis. Ecol Lett 15: 393–405
Beis A, Patakas A (2010) Differences in stomatal responses and root to shoot signalling between two
grapevine varieties subjected to drought. Funct Plant Biol 37: 139–146
Blackman CJ, Brodribb TJ, Jordan GJ (2010) Leaf hydraulic vulnerability is related to conduit
dimensions and drought resistance across a diverse range of woody angiosperms. New Phytol
188: 1113–1123
Brodribb TJ, Holbrook NM (2003) Stomatal closure during leaf dehydration, correlation with other
leaf physiological traits. Plant Physiol 132: 2166–2173
Brodribb TJ, Holbrook NM, Edwards EJ, Gutierrez MV (2003) Relations between stomatal closure,
leaf turgor and xylem vulnerability in eight tropical dry forest trees. Plant Cell Environ 26: 443–
450
Cai J, Tyree MT (2010) The impact of vessel size on vulnerability curves: data and models for within-
species variability in saplings of aspen, Populus tremuloides Michx. Plant Cell Environ 33:
1059–1069
Chatelet DS, Wistrom CM, Purcell AH, Rost TL, Matthews MA (2011) Xylem structure of four grape
varieties and 12 alternative hosts to the xylem-limited bacterium Xylella fastidiosa. Ann Bot 108:
73–85
Chaves MM, Zarrouk O, Francisco R, Costa JM, Santos T, Regalado AP, Rodrigues ML, Lopes CM
(2010) Grapevine under deficit irrigation: hints from physiological and molecular data. Ann Bot

This article is protected by copyright. All rights reserved
105: 661–676
Choat B, Cobb AR, Jansen S (2008) Structure and function of bordered pits: new discoveries and
impacts on whole-plant hydraulic function. New Phytol 177: 608–626
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Field TS, Gleason SM,
Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M,
Mitchell PJ, Nardini A, Pitterman J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE
(2012) Global convergence in the vulnerability of forests to drought. Nature 491: 752–755
Chouzouri A, Schultz HR (2005) Hydraulic anatomy, cavitation susceptibility and gas-exchange of
several grapevine cultivars of different geographical origin. Acta Hortic 689: 325–331
Cochard H, Coll L, Le Roux X, Ameglio T (2002) Unraveling the effects of plant hydraulics on
stomatal closure during water stress in walnut. Plant Physiol 128: 282–290
Cochard H, Cruiziat P, Tyree MT (1992) Use of positive pressures to establish vulnerability curves.
Further support for the air-seeding hypothesis and implications for pressure-volume analysis.
Plant Physiol 100: 205–209
Comstock J (2002) Hydraulic and chemical signalling in the control of stomatal conductance and
transpiration. J Exp Bot 53: 195–200
Cowan IR (1977) Stomatal behaviour and environment. Adv Bot Res 4: 114–228
Davies WJ, Zhang J (1991) Root signals and the regulation of growth and development of plants in
drying soil. Ann Rev Plant Physiol Plant Mol Biol 42: 55–76
Domec JC, Johnson DM (2012) Does homeostasis or disturbance of homeostasis in minimum leaf
water potential explain the isohydric versus anisohydric behaviour of Vitis vinifera L. cultivars?
Tree Physiol 32: 245–248
Domec JC, Noormets A, King JS, Sun G, Mcnulty SG, Gavazzi MJ, Boggs JL, Treasure EA (2009)
Decoupling the influence of leaf and root hydraulic conductances on stomatal conductance and
its sensitivity to vapour pressure deficit as soil dries in a drained loblolly pine plantation. Plant
Cell Environ 32: 980–991
Ennajeh M, Simões F, Khemira H, Cochard H (2011) How reliable is the double-ended pressure
sleeve technique for assessing xylem vulnerability to cavitation in woody Angiosperms? Physiol
Plant 142: 205–210
Franks PJ (2004) Stomatal control and hydraulic conductance, with special reference to tall trees. Tree
Physiol 24: 865–878
Hochberg U, Degu A, Fait A, Rachmilevitch S (2013) Near isohydric grapevine cultivar displays
higher photosynthetic efficiency and photorespiration rates under drought stress as compared
with near anisohydric grapevine cultivar. Physiol Plant 147: 443–452
Jones HG (1998) Stomatal control of photosynthesis and transpiration. J Exp Bot 49: 387–398
Jones HG, Sutherland RA (1991) Stomatal control of xylem embolism. Plant Cell Environ 11: 111–
121

This article is protected by copyright. All rights reserved
Lens F, Sperry JS, Christman MA, Choat B, Rabaey D, Jansen S (2011) Testing hypotheses that link
wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer. New Phytol
190: 709–723
Lenz TI, Wright IJ, Westoby M (2006) Interrelations among pressure-volume curve traits across
species and water availability gradients. Physiol Plant 127: 423–433
Lo Gullo MA, Salleo S, Piaceri EC, Rosso R (1995) Relations between vulnerability to xylem
embolism and xylem conduit dimensions in young trees of Quercus cerris. Plant Cell Environ
18: 661–669
Lovisolo C, Hartung W, Schubert A (2002) Whole-plant hydraulic conductance and root-to-shoot flow
of abscisic acid are independently affected by water stress in grapevines. Funct Plant Biol 29: 1–
8
Lovisolo C, Perrone I, Carra A, Ferrandino A, Flexas J, Medrano H, Schubert A (2010) Drought
induced changes in development and function of grapevine (Vitis spp.) organs and in their
hydraulic and non-hydraulic interactions at the whole plant level: a physiological and molecular
update. Funct Plant Biol 37: 98–116
McCutchan H, Shackel KA (1992) Stem-water potential as a sensitive indicator of water stress in
prune trees (Prunus domestica L. cv. French). J Am Soc Hortic Sci 117: 607–611
Medrano H, Escalona JM, Cifre J, Bota J, Flexas J (2003) A ten year study on the physiology of two
Spanish grapevine cultivars under field conditions: effects of water availability from leaf
photosynthesis to grape yield and quality. Funct Plant Biol 30: 607–619
Meinzer FC (2002) Co-ordination of vapour and liquid phase water transport properties in plants.
Plant Cell Env 25: 265–274
Meinzer FC, Johnson DM, Lachenbruch B, McCulloh KA, Woodruff DR (2009) Xylem hydraulic
safety margins in woody plants: coordination of stomatal control of xylem tension with hydraulic
capacitance. Funct Ecol 23: 922–930
Mitchell PJ, Veneklaas EJ, Lambers H, Burgess SSO (2008) Leaf water relations during summer
water deficit: differential responses in turgor maintenance and variation in leaf structure among
different plant communities in southwestern Australia. Plant Cell Environ 31: 1791–1802
Mott KA, Franks PJ (2001) The role of epidermal turgor in stomatal interactions following a local
perturbation in humidity. Plant Cell Environ 24: 657–662
Nardini A, Salleo S (2000) Limitation of stomatal conductance by hydraulic traits: sensing or
preventing xylem cavitation? Trees 15: 14–24
Nardini A, Tyree MT, Salleo S (2001) Xylem cavitation in the leaf of Prunus laurocerasus and its
impact on leaf hydraulics. Plant Physiol 125: 1700–1709
Palliotti A, Silvestroni O, Petoumenou D, Vignaroli S, Berrios JG (2008) Evaluation of low-energy
demand adaptive mechanisms in Sangiovese grapevine during drought. J Intern Sci de Vigne Vin
42: 41–47

This article is protected by copyright. All rights reserved
Palliotti A, Silvestroni O, Petoumenou D (2009) Photosynthetic and photoinhibition behaviour of two
field-grown grapevine cultivars under multiple summer stresses. Am J Enol Vitic 60: 189–198
Palliotti A, Poni S, Silvestroni O, Tombesi S, Bernizzoni F (2011) Morpho-structural and
physiological performance of Sangiovese and Montepulciano cvv. (Vitis vinifera) under non-
limiting water supply conditions. Funct Plant Biol 38: 888–898
Palliotti A, Tombesi S, Frioni T, Famiani F, Silvestroni O, Zamboni M, Poni S (2014) Morpho-
structural and physiological response of container-grown Sangiovese and Montepulciano cvv.
(Vitis vinifera) to re-watering after a pre-veraison limiting water deficit. Funct Plant Biol, DOI:
10.1071/FP13271
Perrone I, Pagliarani C, Lovisolo C, Chitarra W, Roman F, Schubert A (2012a) Recovery from water
stress affects grape leaf petiole transcriptome. Planta 235: 1383–1396
Perrone I, Gambino G, Chitarra W, Vitali M, Pagliarani C, Riccomagno N, Balestrini R, Kaldenhoff
R, Uehlein N, Gribaudo I, Schubert A, Lovisolo C (2012b) The grapevine root-specific aquaporin
VvPIP2;4N controls root hydraulic conductance and leaf gas exchange under well watered
conditions but not under water stress. Plant Physiol 160: 965–977
Poni S, Bernizzoni F, Civardi S (2007) Response of Sangiovese grapevines to partial root-zone
drying: gas-exchange, growth and grape composition. Sci Hortic 114: 96–103
Pou A, Medrano H, Tomas M, Martorell S, Ribas-Carbò M, Flexas J (2012) Anisohydric behavior in
grapevines results in better performance under moderate water stress and recovery than isohydric
behavior. Plant Soil 359: 335–349
Pou A, Medrano H, Flexas J, Tyerman SD (2013) A putative role for TIP and PIP aquaporins in
dynamics of leaf hydraulic and stomatal conductances in grapevine under water stress and
rewatering. Plant Cell Environ 36: 828–843
Sack L, Cowan PD, Jaikumar N, Holbrook NM (2003) The hydrology of leaves: co-ordination of
structure and function in temperate woody species. Plant Cell Environ 26: 1343–1356
Saliendra NZ, Sperry JS, Comstock J (1995) Influence of leaf water status on stomatal response to
humidity, hydraulic conductance, and soil drought in Betula occidentalis. Planta 196: 357–366
Salleo S, Nardini A, Pitt F, Lo Gullo MA (2000) Xylem cavitation and hydraulic control of stomatal
conductance in laurel (Laurus nobilis L.). Plant Cell Environ 23: 71–79
Schultz HR (2003) Differences in hydraulic architecture account for near-isohydric and anisohydric
behavior of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ 25:
1393–1405
Soar CJ, Dry PR and Loveys BR (2006) Scion photosynthesis and leaf gas exchange in Vitis vinifera
L. cv. Shiraz: Mediation of rootstock effects via xylem sap ABA. Aust J Grape Wine Res 12: 82–
96
Sperry JS, Saliendra NZ (1994) Intra- and inter-plant variation in xylem cavitation in Betula
occidentalis. Plant Cell Environ 17: 1233–1241

This article is protected by copyright. All rights reserved
Syvertsen JP (1982) Minimum leaf water potential and stomatal closure in citrus leaves of different
ages. Ann Bot 49: 827–834
Tardieu F, Simonneau T (1998) Variability among species of stomatal control under fluctuating soil
water status and evaporative demand: modelling isohydric and anisohydric behaviours. J Exp Bot
49: 419–432
Tombesi S, Johnson RS, Day KR, DeJong TM (2010) Interactions between rootstock, inter-stem and
scion xylem vessel characteristics of peach trees growing on rootstocks with contrasting size-
controlling characteristics. AoB Plants 2010: plq013
Tramontini S, Vitali M, Centioni L, Schubert A, Lovisolo C. (2013) Rootstock control of scion
response to water stress in grapevine. Env Exp Bot 93: 20–26
Tyree MT, Dixon MA (1983) Cavitation events in Thuja occidentalis L. Ultrasonic acoustic emissions
from the sapwood can be measured. Plant Physiol 72: 1094–1099
Tyree MT, Sperry JS (1988) Do woody plants operate near the point of catastrophic xylem
dysfunction caused by dynamic water stress? Plant Physiol 88: 574–580
Tyree MT, Sperry JS (1989) Vulnerability of xylem to cavitation and embolism. Ann Rev Plant
Physiol Plant Mol Biol 40: 19–38
Vandeleur RK, Mayo G, Shelden MC, Gilliham M, Kaiser BN, Tyerman SD (2009) The role of
plasma membrane intrinsic protein aquaporins in water transport through roots: diurnal and
drought stress responses reveal different strategies between isohydric and anisohydric cultivars of
grapevine. Plant Phys 149: 445–460
Vilagrosa A, Morales F, Abadía A, Bellot J, Cochard H, Gil-Pelegrin E (2010) Are symplast tolerance
to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated
analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-
resistant species. Environ Exp Bot 69: 233–242
Zufferey V, Cochard H, Ameglio T, Spring Jl, Viret O (2011) Diurnal cycles of embolism formation
and repair in petioles of grapevine (Vitis vinifera cv. Chasselas). J Exp Bot 62: 3885–3894
This article is protected by copyright. All rights reserved
Figure legends
Fig. 1. Stem water potential (Ψstem) (A) and leaf water potential (Ψleaf) (B) pattern on 8/7/2013 in
Sangiovese and Montepulciano grapevine. Data were collected from five vines per each cultivar.
This article is protected by copyright. All rights reserved
Fig. 2. Relationship between leaf water potential (Ψleaf) and stomatal conductance (gs) in primary
leaves of detached shoots of Sangiovese and Montepulciano grapevine in 2012 (A). Data were
collected from five vines per each cultivar. Data were fitted to a sigmoid curve (Sangiovese R2=0.81,
P<0.001; Montepulciano R2=0.89, P<0.001). Relationship between leaf water potential (Ψleaf) (B),
stem water potential (Ψstem) (C) and stomatal conductance (gs) in Sangiovese and Montepulciano
grapevine in 2013. Data were collected from five vines per each cultivar. Data were fitted to a sigmoid
curve [(B) Sangiovese R2=0.86, P<0.001; Montepulciano R2=0.83, P<0.001; (C) Sangiovese R2=0.86,
P<0.001; Montepulciano R2=0.89, P<0.001].
Fig. 3. Relationship between leaf relative water content (RWC) and leaf water potential (Ψleaf) in
leaves of Sangiovese and Montepulciano grapevines. Data were obtained from 10 leaves per cultivar.
This article is protected by copyright. All rights reserved
Fig. 4. Relationship between percent loss of hydraulic conductance (PLC) and stem water potential
(Ψstem) (MPa) in petioles of Sangiovese and Montepulciano grapevines. Each point is the mean of five
vines (n=5) ± SE. Data of each cultivar were fitted in a sigmoidal curve (Sangiovese R2=0.98,
P<0.001; Montepulciano R2=0.96, P<0.001).
Fig. 5. Relationship between percentage loss of leaf stomatal conductance (PLgs) and percentage loss
of petiole hydraulic conductance (PLC) in Sangiovese and Montepulciano grapevines. PLgs was
calculated on the basis of the regression curves shown in Fig 4. Data of each cultivar were fitted in a:
sigmoidal curve (Sangiovese R2=0.98, P<0.001; Montepulciano R2=0.99, P<0.001).
This article is protected by copyright. All rights reserved
Fig. 6. Number of xylem vessels (% of total) per vessel size class (µm) in petioles of Sangiovese and
Montepulciano grapevines. Bars are mean values of five vines (n=5) per cultivar ± SE.
Fig. 7. Relationship between cumulated theoretical hydraulic conductance (% of total) and vessel
diameter (µm) in petioles of primary leaves in Sangiovese and Montepulciano grapevines.
This article is protected by copyright. All rights reserved
Fig. 8. Relationship between percentage loss of leaf stomatal conductance (PLgs), percent loss of
petiole hydraulic conductance (PLC) and leaf water potential (Ψleaf) in Montepulciano (A) and
Sangiovese (B) grapevine. Solid vertical lines represent the leaf water potential at turgor loss point
(πtlp) in Montepulciano (A) and Sangiovese (B) leaves.

This article is protected by copyright. All rights reserved
Table 1. Leaf water potential at turgor loss point (πtlp) (MPa), modulus of elasticity (ε) (MPa), relative
water content at turgor loss point (RWCtlp) (%), and osmotic potential at full turgor (πo) (MPa) of
leaves of Sangiovese and Montepulciano grapevines. Values are means of leaves sampled from five
vines (n=5) ± SE. For each parameter significant differences are indicated with * (P<0.05).
Sangiovese Montepulciano t-test
πtlp –1.67 ± 0.09 –1.36 ± 0.08 *
πo –1.38 ± 0.04 –1.16 ± 0.08 *
RWCtlp 95.50 ± 0.81 96.55 ± 0.39 ns
ε 7.82 ± 0.75 7.96 ± 0.61 ns

This article is protected by copyright. All rights reserved
Table 2. Maximum diameter of xylem vessels (µm) accounting for 20, 50 and 80% (D20, D50, D80
respectively) of total theoretical hydraulic conductance (as shown in Fig. 7); xylem pressures (MPa)
inducing percentage loss of conductance (PLC) of 20, 50 and 80% (P20, P50, P80) in petioles and leaf
specific petiole hydraulic conductivity (kpetiole) in Sangiovese and Montepulciano grapevines. Values
are means of leaves from five vines (n=5) ± SE. For each parameter significant differences are
indicated with * (P<0.05).
Sangiovese Montepulciano t-test
D20 (µm) 14.63 ± 0.23 16.27 ± 0.19 *
D50 (µm) 18.59 ± 0.31 20.93 ± 0.42 *
D80 (µm) 22.33 ± 0.33 26.23 ± 0.53 *
P20 (MPa) –0.67 ± 0.09 –0.49 ± 0.05 *
P50 (MPa) –1.25 ± 0.07 –1.08 ± 0.05 *
P80 (MPa) –1.78 ± 0.08 –1.79 ± 0.05 ns
kpetiole (mmol m–1 s–1 MPa–1) 5.04 ± 0.46 7.13 ± 1.19 *





























