Embed Size (px)
Citation preview

lable at ScienceDirect
International Biodeterioration & Biodegradation 62 (2008) 483–486
Contents lists avai
International Biodeterioration & Biodegradation
journal homepage: www.elsevier .com/locate/ ib iod
Short communication
Pseudonocardia in white colonizations in two caves with Paleolithic paintings
F. Stomeo, M.C. Portillo, J.M. Gonzalez*, L. Laiz, C. Saiz-JimenezInstituto de Recursos Naturales y Agrobiologia, IRNAS-CSIC, Avda. Reina Mercedes 10, 41012 Sevilla, Spain
a r t i c l e i n f o
Article history:Received 4 September 2007Received in revised form 7 November 2007Accepted 11 December 2007Available online 24 June 2008
Keywords:PseudonocardiaActinobacteriaMicrobial colonizationPaleolithic paintingsSubterranean environments
* Corresponding author. Tel.: þ34 95 462 4711; faxE-mail address: [email protected] (J.M. Gonza
0964-8305/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.ibiod.2007.12.011
a b s t r a c t
Deterioration caused by microbial colonization is one of the factors affecting the conservation of Pa-leolithic paintings in caves. Assessing the composition of the microbial communities represents the firststage in understanding and controlling these colonizations. The microbial communities of the Cave ofDona Trinidad (Ardales, Malaga, Southern Spain) and Santimamine (Kortezubi, Biscay, Northern Spain)were analyzed by molecular techniques based on RNA and DNA. Microbial community fingerprintsobtained by denaturing gradient gel electrophoresis and 16S rRNA gene libraries and sequencing de-tected that the major constituents were Actinobacteria belonging to the genus Pseudonocardia. Unlikeprevious reports on different caves, this study confirms the highly significant role of Actinobacteria asmetabolically active microorganisms in the biodeterioration of subterranean environments.
� 2008 Elsevier Ltd. All rights reserved.
1. Introduction
Caves often contain unique artwork; some of it dated at over20,000 years old. This is the case of Paleolithic paintings and en-gravings. At present, most of the well known and visited caves thatcontain paintings are affected by progressive biodeterioration(Holden, 2002; Allemand and Bahn, 2005) and the control, treat-ment, and preservation of these environments are serious prob-lems garnering the interest of scientists, conservators, and thepublic.
Recent reports on molecular surveys of microbial communitiescolonizing cave environments suggest the participation of variousclasses of microorganisms includingddepending on thesitedphototrophic microorganisms such as cyanobacteria andmicroscopic algae (Albertano and Urzı, 1999; Albertano et al., 2000),a variety of bacteria (Schabereiter-Gurtner et al., 2002a,b, 2004;Zimmermann et al., 2005), low-temperature Crenarchaeota (Gon-zalez et al., 2006), and fungi (Dupont et al., 2007). Among thebacteria, the division Actinobacteria represents a group of micro-organisms generally present both in soils and in subterranean en-vironments (Goodfellow and Williams, 1983; Bull et al., 1992; Laizet al., 2003; Groth et al., 2001), and their role in biodeterioratingprocesses has been emphasized (Monte and Ferrari, 1993; Grothet al., 1999). However, different methods used for their detection(culturing vs. molecular techniques) have provided highly di-vergent information on their representation in the microbial
: þ34 95 462 4002.lez).
All rights reserved.
communities. While culturing procedures show the Actinobacteriaas the major component of microbial communities in caves andtombs (Monte and Ferrari, 1993; Groth et al., 1999, 2001), moleculartechniques suggest that this bacterial group constitutes a minorityof the bacterial community in caves with Paleolithic paintings(Schabereiter-Gurtner et al., 2002a,b, 2004; Laiz et al., 2003).
Recent developments in molecular techniques provide us withthe opportunity to perform microbial surveys of natural environ-ments based on both DNA and RNA. DNA analyses reveal informationon the presence of microorganisms (Ward et al.,1990). RNA analysesallow the detection of those microorganisms showing metabolicactivity in situ (Mills et al., 2004; Gonzalez et al., 2006) since RNAcontent in bacteria is related to growth rate and activity (Molin andGivskov, 1999). Consequently, RNA-based strategies result indetecting the microbes metabolically active in colonizing thestudied site.
In this study, we report on the major participation of Actino-bacteria in the active development of whitish microbial coloniza-tions in two caves containing valuable Paleolithic paintings andengravings.
2. Materials and methods
The microbial communities developing as white microbial colonizations (Fig. 1)in the Caves of Dona Trinidad (Ardales, Malaga, Southern Spain) and Santimamine(Kortezubi, Biscay, Northern Spain) were analyzed. The works of art decorating DonaTrinidad and Santimamine Caves have been previously described by Cantalejo et al.(2006) and Apellaniz (1971), respectively. These caves are located in different cli-matic and geographical areas. The Dona Trinidad and Santimamine Caves were inthe range of 15–18 �C (S. Sanchez-Moral, National Museum of Natural Sciences, CSIC,Madrid, personal communication) and 11–14 �C (M. Zaparain, CRN, Madrid, personal

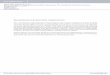
Fig. 1. White colonizations developing at Dona Trinidad (A) and Santimamine Caves (B). Bars indicate 5 cm.
F. Stomeo et al. / International Biodeterioration & Biodegradation 62 (2008) 483–486484
communication), respectively, and both caves maintained a relative humidity higherthan 90% throughout the year. At present, no permanent artificial lights are allowedin these caves. In Dona Trinidad Cave white colonizations can be observed along thestairway from the entrance up to some of the speleothems in the first hall, witha marked indication of progression through the whole cave. In Santimamine Cave,the development of white colonizations is found mainly on the walls of the entrancehall.
Three colonies from each location were collected under aseptic conditions andimmediately preserved in RNAlater solution (Ambion, Inc., Austin, TX, USA) and kepton ice until arrival in the laboratory, where they were maintained at �80 �C. Eachsample consisted in an entire single colony. RNA was extracted using the RNA-queous4PCR total RNA extraction kit (Ambion, Inc.), following the manufacturer’srecommendations. Thermoscript reverse transcriptase was used to synthesize cDNAfrom bacterial 16S rRNA genes using the primer 518R (Neefs et al., 1990). DNA ex-traction was carried out using the Nucleospin Food DNA extraction kit (Mackerel-Nagel, Duren, Germany). PCR amplifications were performed using ExTaq DNApolymerase (Takara, Tokyo, Japan) and the primer pairs 27F and 907R for DNA or 27Fand 518R for cDNA (Gonzalez et al., 2003). Amplification products were purifiedusing the JetQuick PCR purification kit (Genomed, Bad Oeynhausen, Germany) andDNA libraries were constructed with the TOPO-TA cloning kit (Invitrogen, Carlsbad,CA, USA). DNA libraries were screened as described by Gonzalez et al. (2003). Se-lected clones were sequenced. Sequence information was manually inspected usingthe software Chromas v.1.45 (Technelysium, Tewantin, Australia). Edited sequenceswere submitted for homology search using the algorithm Blastn (Altschul et al.,1990; http://www.ncbi.nlm.nih.gov/blast) and screened for the presence of chi-meras as described by Gonzalez et al. (2005).
Microbial community fingerprints were obtained by denaturing gradient gelelectrophoresis (DGGE) as previously described (Muyzer et al., 1993; Gonzalez andSaiz-Jimenez, 2004) using a GC-rich tailed primer (341F-GC) and the reverse primer518R.
Actinobacteria(Pseudonocardia)
Firm(28Other
Actinobacteria(3%)
Planctomycetes(9%)
Chloroflexi (3%)
A
86%
Fig. 2. Composition of the microbial communities detected by the use of molecular analya molecular survey of white colonizations at Dona Trinidad Cave.
The unique sequences obtained from Dona Trinidad and Santimamine Caves canbe retrieved under the accession numbers EF447064–EF447082 and EF447083–EF447092, respectively.
3. Results
A DNA-based molecular survey of the bacterial communitiesfrom white colonizations at Dona Trinidad Cave (Fig. 2) showed thepresence of a variety of microorganisms belonging to differentbacterial phyla. Based on DNA analysis, the phylum Actinobacteriawere the most frequently found in the analyzed 16S rRNA gene li-brary, reaching 44% of sequences out of a total of 25 clones. Withinthe Actinobacteria, most of the clones corresponded to microor-ganisms belonging to Streptomyces, the Rubrobacteridae, and a highproportion of them to the genus Pseudonocardia. Several un-cultured Pseudonocardia were detected, constituting up to 36% ofthe analyzed clones from a DNA-based clone library. Only 20% ofthese Pseudonocardia-related sequences corresponded to the met-abolically active dominant phylotype detected in Dona TrinidadCave (see below). Phylum Firmicutes constituted about 28% of theanalyzed clones from a DNA-based molecular survey and wasrepresented mainly by the genera Bacillus and Streptococcus. Otherbacterial phyla represented in these colonies were the Planctomy-cetes, Nitrospirae, Acidobacteria, and Proteobacteria. The Proteo-bacteria were constituted by members of the beta-, gamma-, and
OtherActinobacteria
(8%)
icutes%)
Beta-Proteobacteria (4%)
Gamma-Proteobacteria(8%)
Acidobacteria(4%)
Planctomycetes(4%)
Nitrospirae(4%)
Delta-Proteobacteria (4%)
B
36%
ses based on RNA (A) and DNA (B) as percentages of the processed sequences during
Planctomycetes (6%)
Alphaproteobacteria(9%)
Betaproteobacteria (3%)
Acidobacteria (3%)Nitrospirae (3%)
Actinobacteria(Pseudonocardia)
(66%)
Actinobacteria(others) (11%)
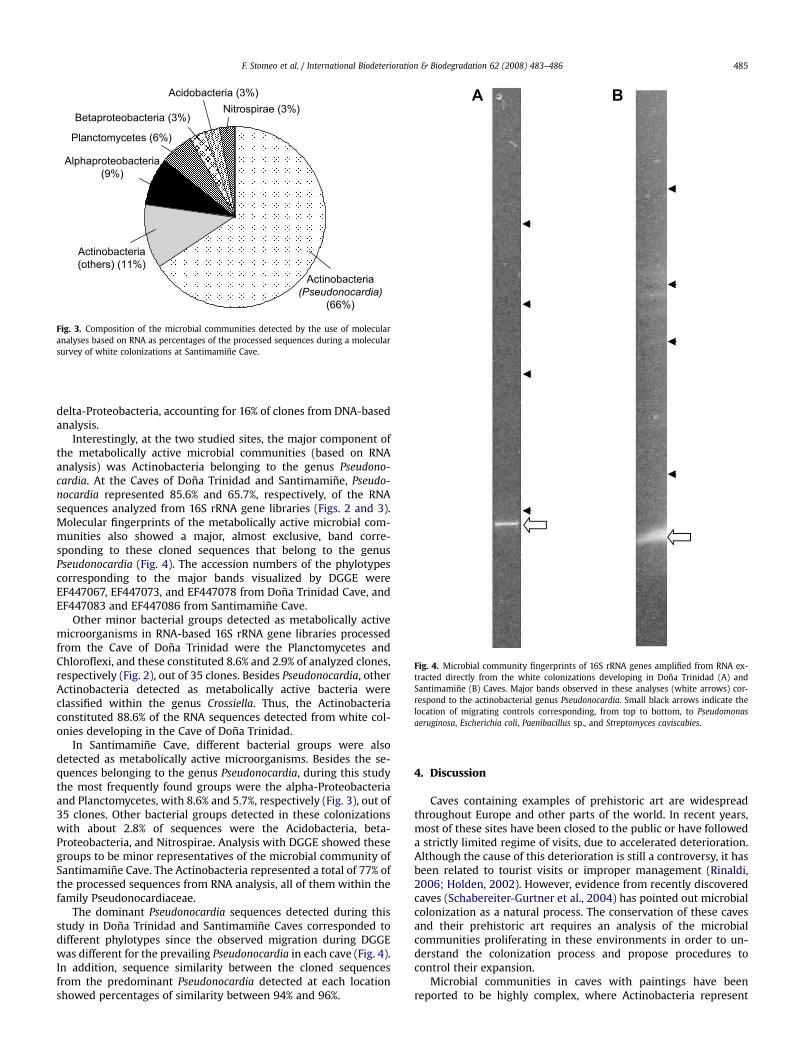
Fig. 3. Composition of the microbial communities detected by the use of molecularanalyses based on RNA as percentages of the processed sequences during a molecularsurvey of white colonizations at Santimamine Cave.
Fig. 4. Microbial community fingerprints of 16S rRNA genes amplified from RNA ex-tracted directly from the white colonizations developing in Dona Trinidad (A) andSantimamine (B) Caves. Major bands observed in these analyses (white arrows) cor-respond to the actinobacterial genus Pseudonocardia. Small black arrows indicate thelocation of migrating controls corresponding, from top to bottom, to Pseudomonasaeruginosa, Escherichia coli, Paenibacillus sp., and Streptomyces caviscabies.
F. Stomeo et al. / International Biodeterioration & Biodegradation 62 (2008) 483–486 485
delta-Proteobacteria, accounting for 16% of clones from DNA-basedanalysis.
Interestingly, at the two studied sites, the major component ofthe metabolically active microbial communities (based on RNAanalysis) was Actinobacteria belonging to the genus Pseudono-cardia. At the Caves of Dona Trinidad and Santimamine, Pseudo-nocardia represented 85.6% and 65.7%, respectively, of the RNAsequences analyzed from 16S rRNA gene libraries (Figs. 2 and 3).Molecular fingerprints of the metabolically active microbial com-munities also showed a major, almost exclusive, band corre-sponding to these cloned sequences that belong to the genusPseudonocardia (Fig. 4). The accession numbers of the phylotypescorresponding to the major bands visualized by DGGE wereEF447067, EF447073, and EF447078 from Dona Trinidad Cave, andEF447083 and EF447086 from Santimamine Cave.
Other minor bacterial groups detected as metabolically activemicroorganisms in RNA-based 16S rRNA gene libraries processedfrom the Cave of Dona Trinidad were the Planctomycetes andChloroflexi, and these constituted 8.6% and 2.9% of analyzed clones,respectively (Fig. 2), out of 35 clones. Besides Pseudonocardia, otherActinobacteria detected as metabolically active bacteria wereclassified within the genus Crossiella. Thus, the Actinobacteriaconstituted 88.6% of the RNA sequences detected from white col-onies developing in the Cave of Dona Trinidad.
In Santimamine Cave, different bacterial groups were alsodetected as metabolically active microorganisms. Besides the se-quences belonging to the genus Pseudonocardia, during this studythe most frequently found groups were the alpha-Proteobacteriaand Planctomycetes, with 8.6% and 5.7%, respectively (Fig. 3), out of35 clones. Other bacterial groups detected in these colonizationswith about 2.8% of sequences were the Acidobacteria, beta-Proteobacteria, and Nitrospirae. Analysis with DGGE showed thesegroups to be minor representatives of the microbial community ofSantimamine Cave. The Actinobacteria represented a total of 77% ofthe processed sequences from RNA analysis, all of them within thefamily Pseudonocardiaceae.
The dominant Pseudonocardia sequences detected during thisstudy in Dona Trinidad and Santimamine Caves corresponded todifferent phylotypes since the observed migration during DGGEwas different for the prevailing Pseudonocardia in each cave (Fig. 4).In addition, sequence similarity between the cloned sequencesfrom the predominant Pseudonocardia detected at each locationshowed percentages of similarity between 94% and 96%.
4. Discussion
Caves containing examples of prehistoric art are widespreadthroughout Europe and other parts of the world. In recent years,most of these sites have been closed to the public or have followeda strictly limited regime of visits, due to accelerated deterioration.Although the cause of this deterioration is still a controversy, it hasbeen related to tourist visits or improper management (Rinaldi,2006; Holden, 2002). However, evidence from recently discoveredcaves (Schabereiter-Gurtner et al., 2004) has pointed out microbialcolonization as a natural process. The conservation of these cavesand their prehistoric art requires an analysis of the microbialcommunities proliferating in these environments in order to un-derstand the colonization process and propose procedures tocontrol their expansion.
Microbial communities in caves with paintings have beenreported to be highly complex, where Actinobacteria represent

F. Stomeo et al. / International Biodeterioration & Biodegradation 62 (2008) 483–486486
a low percentage (Schabereiter-Gurtner et al., 2002a,b, 2004;Zimmermann et al., 2005; Gonzalez et al., 2006); however, thisstudy has found that Actinobacteria (i.e., Pseudonocardia) were thepredominant component of the white colonizations present in thecaves. The RNA-based molecular approach confirmed Actino-bacteria as the major metabolically active member of the microbialcommunity forming white colonizations. Two methods, clone li-braries and microbial fingerprints, supported this finding. Conse-quently, Actinobacteria constituted the major componentcontributing to the expansion of white colonizations in the twocaves under analysis.
The potential bias generated during PCR amplification againstthe amplification of sequences from the Actinobacteria (Heueret al., 1997; Stach et al., 2003) and the probable overestimation ofthe proportion of Actinobacteria from culturing bacteria in stan-dard culture media (Laiz et al., 2003) might be the two factorscontributing to the different estimates of Actinobacteria in naturalmicrobial communities. In this study, bacterial domain primershave been used to detect Actinobacteria. Thus, the results suggestthat PCR bias might be an unsupported cause for the low estimatesof Actinobacteria in soil and subterranean environments usingmolecular methods. Herein, culture-independent strategies basedon DNA and RNA were used. In contrast to recent reports ona number of caves with rock art paintings (Schabereiter-Gurtneret al., 2002a,b, 2004), this study corroborates the importance ofActinobacteria, specifically those belonging to the genus Pseudo-nocardia, in the colonization processes of cave environments. At-tempts to isolate the active Pseudonocardia strains are in progressand, if they are successful, it would be of great interest to un-derstand the biodeterioration processes and pursue adequateconservation strategies at caves with prehistoric paintings andengravings.
Acknowledgements
The authors acknowledge funding from the Junta de Andalucıa,Consejerıa de Cultura, through an agreement to study the Cave ofDona Trinidad in Ardales. Santimamine Cave was studied thanks toa research agreement with CRN, S.A., and the Country Council ofBiscay. FS acknowledges support from European Communityproject MEST-CT-2004-513915. JMG is grateful for the supportprovided by the project CGL2006-11561/BTE, and CSJ appreciatessupport from project GCL2006-07424/BOS.
References
Albertano, P., Urzı, C., 1999. Structural interactions among epilithic cyanobacteriaand heterotrophic microorganisms in Roman hypogea. Microbial Ecology 38,244–252.
Albertano, P., Bruno, L., Moscone, D., D’Ottavi, D., Palleschi, G., 2000. The effect ofphotosynthesis on pH variation in cyanobacterial biofilms from Roman cata-combs. Journal of Applied Phycology 12, 379–384.
Allemand, L., Bahn, P.G., 2005. Best way to protect rock art is to leave it alone.Nature 433, 800.
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J., 1990. Basic localalignment search tool. Journal of Molecular Biology 215, 403–410.
Apellaniz, J.M., 1971. La Caverna de Santimamine. Excma. Diputacion de Vizcaya,Bilbao, Spain.
Bull, A.T., Goodfellow, M., Slater, J., 1992. Biodiversity as a source of innovation inbiotechnology. Annual Review of Microbiology 46, 219–252.
Cantalejo, P., Maura, R., Espejo, M.M., Ramos, J.F., Medianero, J., Aranda, A.,Duran, J.J., 2006. La Cueva de Ardales: Arte Prehistorico y Ocupacion en elPaleolıtico Superior. Centro de Ediciones de la Diputacion de Malaga, 427pp.
Dupont, J., Jacquet, C., Dennetiere, B., Lacoste, B., Bousta, F., Orial, G., Cruaud, C.,Couloux, A., Roquebert, M.F., 2007. Invasion of the French paleolithic paintedcave of Lascaux by members of the Fusarium solani species complex. Mycologia99, 526–533.
Gonzalez, J.M., Ortiz-Martinez, A., Gonzalez-del Valle, M.A., Laiz, L., Saiz-Jimenez, C.,2003. An efficient strategy for screening large libraries of amplified 16S rDNAsequences from complex environmental communities. Journal of Microbiolog-ical Methods 55, 459–463.
Gonzalez, J.M., Saiz-Jimenez, C., 2004. Microbial diversity in biodeteriorated mon-uments as studied by denaturing gradient gel electrophoresis. Journal ofSeparation Science 27, 174–180.
Gonzalez, J.M., Zimmermann, J., Saiz-Jimenez, C., 2005. Evaluating putative chimericsequences from PCR amplified products and other cross-over events.Bioinformatics 21, 333–337.
Gonzalez, J.M., Portillo, M.C., Saiz-Jimenez, C., 2006. Metabolically active Cren-archaeota in Altamira Cave. Naturwissenschaften 93, 42–45.
Goodfellow, M., Williams, S.T., 1983. Ecology of actinomycetes. Annual Review ofMicrobiology 37, 189–216.
Groth, I., Vetermann, R., Schuetze, B., Schumann, P., Saiz-Jimenez, C., 1999. Acti-nomycetes in karstic caves of Northern Spain (Altamira and Tito Bustillo).Journal of Microbiological Methods 36, 115–122.
Groth, I., Schumann, P., Laiz, L., Sanchez–Moral, S., Canaveras, J.C., Saiz-Jimenez, C.,2001. Geomicrobiological study of the Grotta dei Cervi, Porto Badisco, Italy.Geomicrobiology Journal 18, 241–258.
Heuer, H., Krsek, M., Baker, P., Smalla, K., Wellington, E.M.H., 1997. Analysis ofactinomycete communities by specific amplification of genes encoding 16SrRNA and gel-electrophoretic separation in denaturing gradients. Applied andEnvironmental Microbiology 63, 3233–3241.
Holden, C., 2002. Cave paintings in jeopardy. Science 297, 47.Laiz, L., Gonzalez, J.M., Saiz-Jimenez, C., 2003. Microbial communities in caves:
ecology, physiology, and effects on Paleolithic paintings. In: Koestler, R.J.,Koestler, V.R., Charola, A.E., Nieto-Fernandez, F.E. (Eds.), Art, Biology, and Con-servation: Biodeterioration of Works of Art. The Metropolitan Museum of Art,New York, pp. 210–225.
Mills, H.J., Martinez, R.J., Story, S., Sobecky, P.A., 2004. Identification of members ofthe metabolically active microbial populations associated with Beggiatoa spe-cies mat communities from Gulf of Mexico cold-seep sediments. Applied andEnvironmental Microbiology 70, 5447–5458.
Molin, S., Givskov, M., 1999. Application of molecular tools for in situ monitoring ofbacterial growth activity. Environmental Microbiology 1, 383–391.
Monte, M., Ferrari, R., 1993. Biodeterioration in subterranean environments. Aero-biologia 9, 141–148.
Muyzer, G., de Waal, E.C., Uitterlinden, A.G., 1993. Profiling of complex microbialpopulations by denaturing gradient gel electrophoresis analysis of polymerasechain reaction-amplified genes coding for 16S rRNA. Applied and Environ-mental Microbiology 59, 695–700.
Neefs, J.M., Van de Peer, Y., Hendriks, L., De Wachter, R., 1990. Compilation ofsmall ribosomal subunit RNA sequences. Nucleic Acids Research 18, 2237–2317.
Rinaldi, A., 2006. Saving a fragile legacy. EMBO Reports 7, 1075–1079.Schabereiter-Gurtner, C., Saiz-Jimenez, C., Pinar, G., Lubitz, W., Rolleke, S., 2002a.
Altamira cave Paleolithic paintings harbor partly unknown bacterial commu-nities. FEMS Microbiology Letters 211, 7–11.
Schabereiter-Gurtner, C., Saiz-Jimenez, C., Pinar, G., Lubitz, W., Rolleke, S., 2002b.Phylogenetic 16S rRNA analysis reveals the presence of complex and partlyunknown bacterial communities in Tito Bustillo Cave, Spain, and on its Paleo-lithic paintings. Environmental Microbiology 4, 392–400.
Schabereiter-Gurtner, C., Saiz-Jimenez, C., Pinar, G., Lubitz, W., Rolleke, S., 2004.Phylogenetic diversity of bacteria associated with Paleolithic paintings andsurrounding rock walls in two Spanish caves (Llonın and La Garma). FEMSMicrobiology Ecology 47, 235–247.
Stach, J.E.M., Maldonado, L.A., Ward, A.C., Goodfellow, M., Bull, A.T., 2003. Newprimers for the class Actinobacteria: application to marine and terrestrialenvironments. Environmental Microbiology 5, 828–841.
Ward, D.M., Weller, R., Bateson, M.M., 1990. 16S rRNA sequences reveal numerousuncultured microorganisms in a natural community. Nature 345, 63–65.
Zimmermann, J., Gonzalez, J.M., Ludwig, W., Saiz-Jimenez, C., 2005. Detection andphylogenetic relationships of a highly diverse uncultured acidobacterialcommunity on Paleolithic paintings in Altamira Cave using 23S rRNA sequenceanalyses. Geomicrobiology Journal 22, 379–388.