Embed Size (px)
Citation preview

Principles of Biochemistry Fourth Edition
Chapter 9Lipids and Membranes
Copyright © 2006 Pearson Prentice Hall, Inc.
Horton • Moran • Scrimgeour • Perry • Rawn
陳 玉 婷

Lipids
Storage energy: triacylglycerol (fat, oil)Cell membrane: glycerophospholipids, sphingolipids, sterolsProtective surface: wax (in cell wall, exoskeleton, skin)Enzyme cofactor:Electron carrier: cytochrome P450Light-absorbing pigments: chlorophyllHydrphobic anchorsEmulsifying agentsHormones: testosterone, estradiolInteracellular messengers: sialic acid
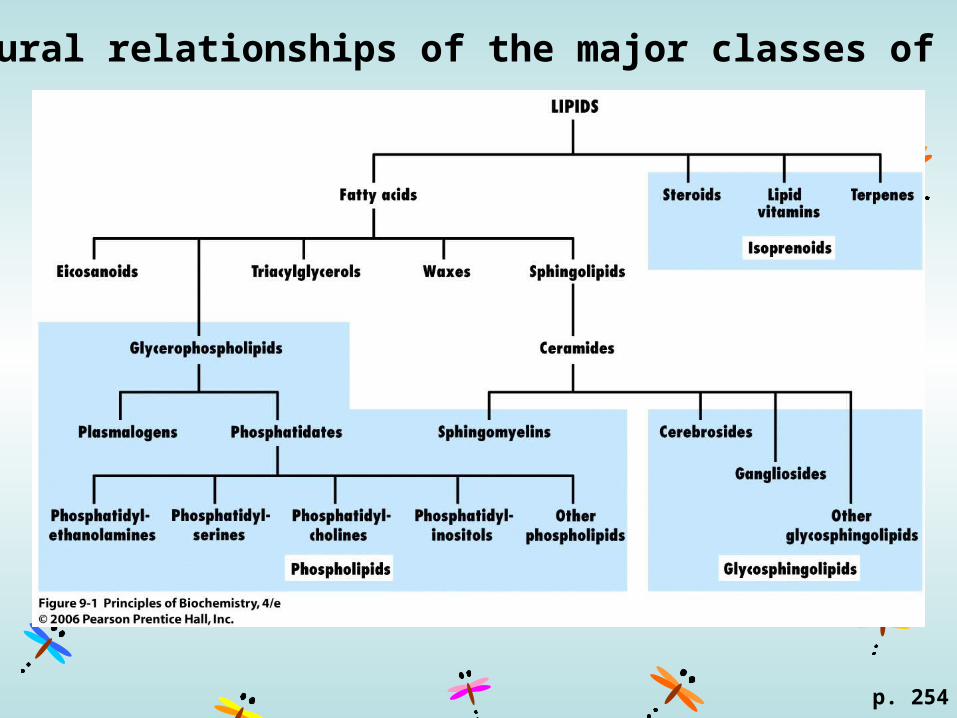
Structural relationships of the major classes of lipids
p. 254
Fatty acid
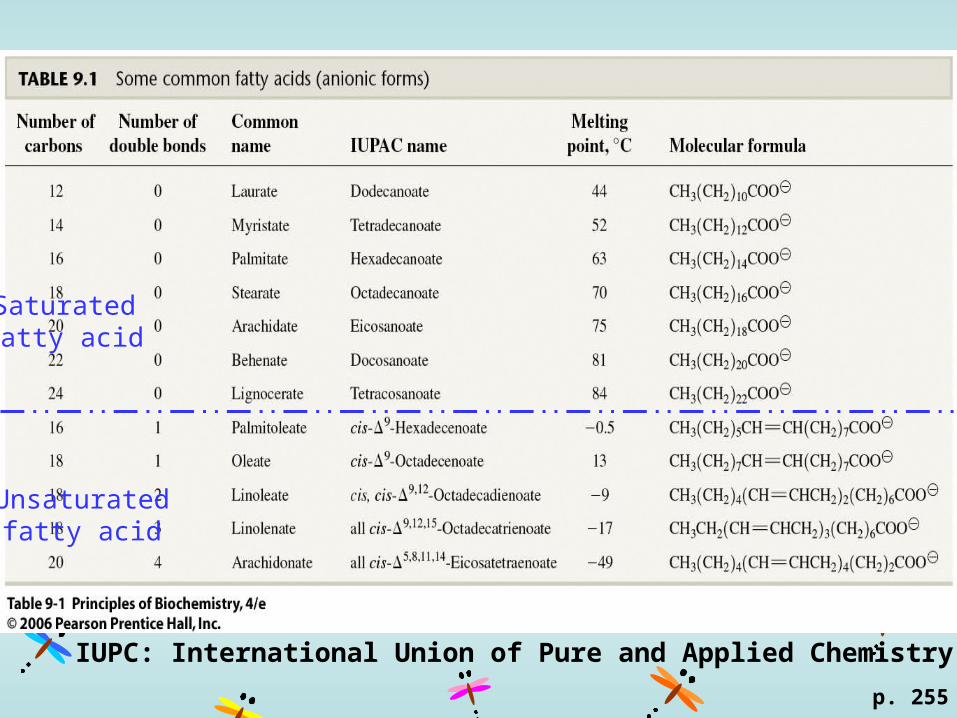
p. 255
Saturatedfatty acid
Unsaturatedfatty acid
IUPC: International Union of Pure and Applied Chemistry
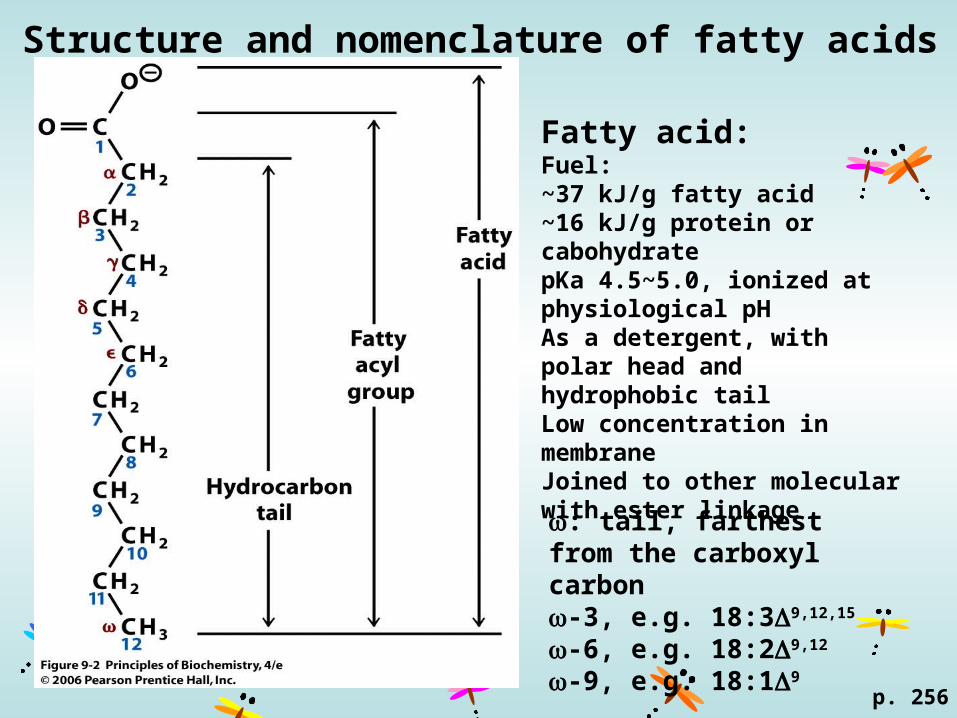
Structure and nomenclature of fatty acids
p. 256
: tail, farthest from the carboxyl carbon-3, e.g. 18:39,12,15
-6, e.g. 18:29,12
-9, e.g. 18:19
Fatty acid: Fuel:~37 kJ/g fatty acid~16 kJ/g protein or cabohydratepKa 4.5~5.0, ionized at physiological pHAs a detergent, with polar head and hydrophobic tailLow concentration in membraneJoined to other molecular with ester linkage
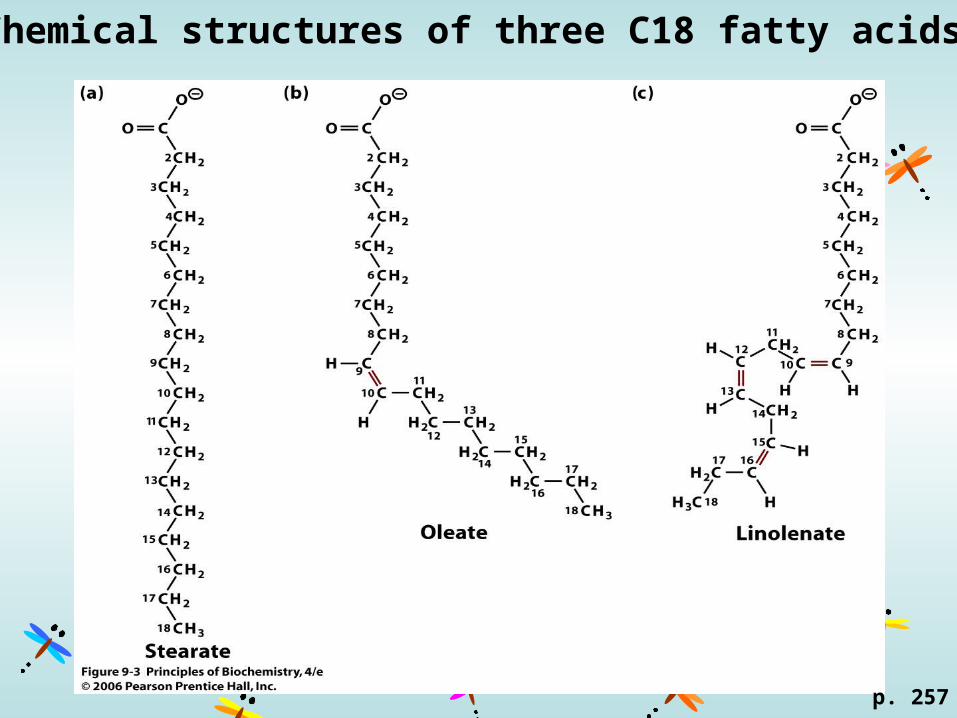
Chemical structures of three C18 fatty acids
p. 257

Triacylglycerol
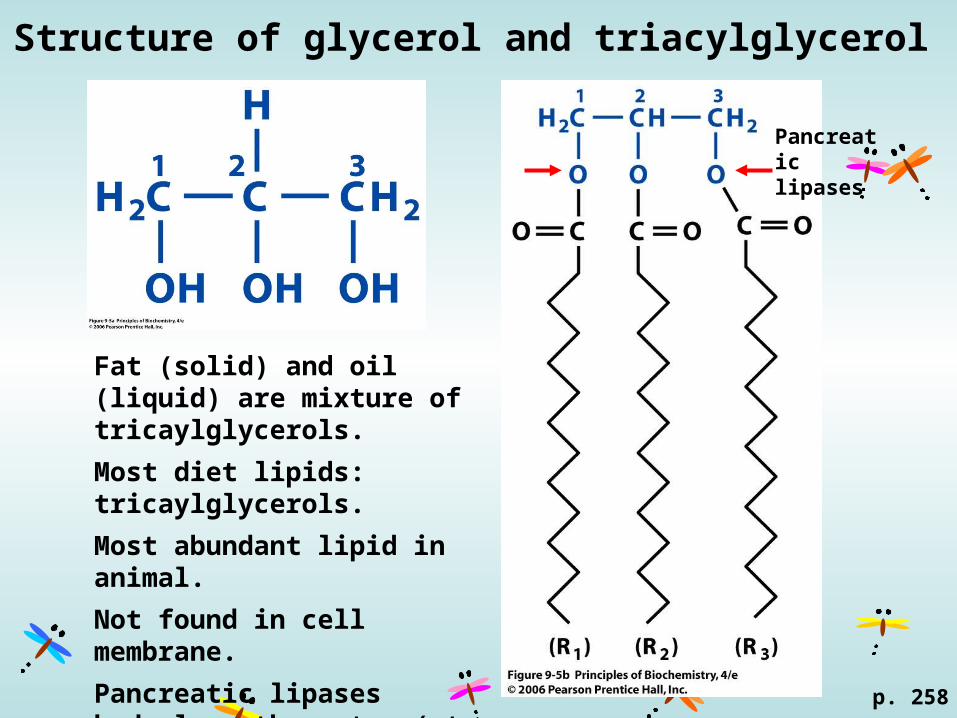
Structure of glycerol and triacylglycerol
p. 258
Fat (solid) and oil (liquid) are mixture of tricaylglycerols.
Most diet lipids: tricaylglycerols.
Most abundant lipid in animal.
Not found in cell membrane.
Pancreatic lipases hydrolyse the ester (at C1 and C3) of gtracylglycerol.
Pancreatic lipases

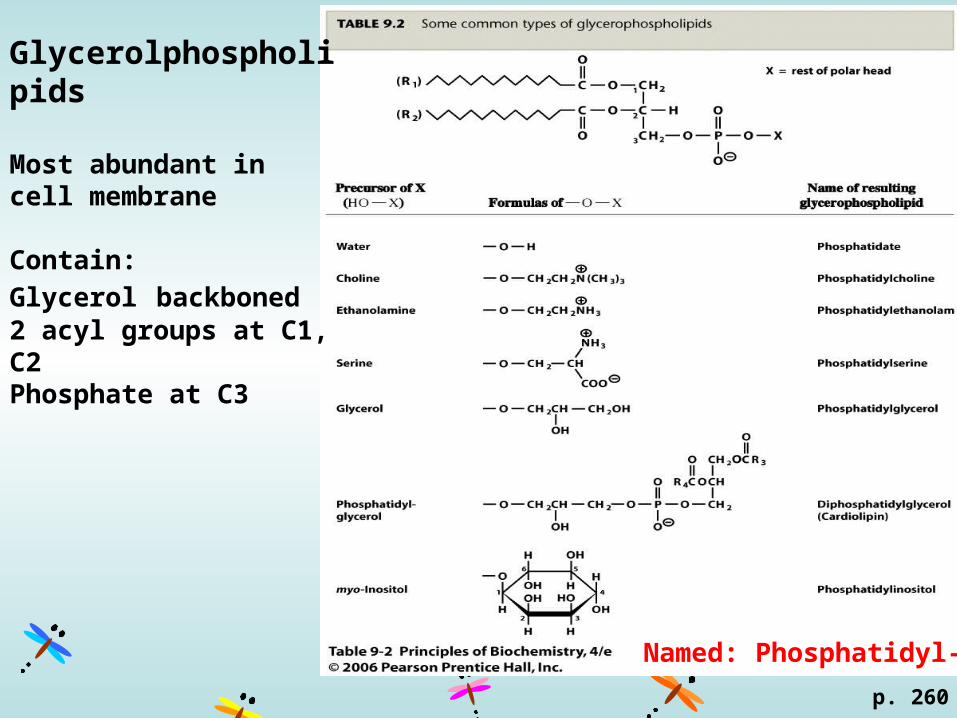
Glycerolphospholipids
p. 260
Glycerolphospholipids
Most abundant in cell membrane
Contain:
Glycerol backboned2 acyl groups at C1, C2Phosphate at C3
Named: Phosphatidyl-X

p. 261
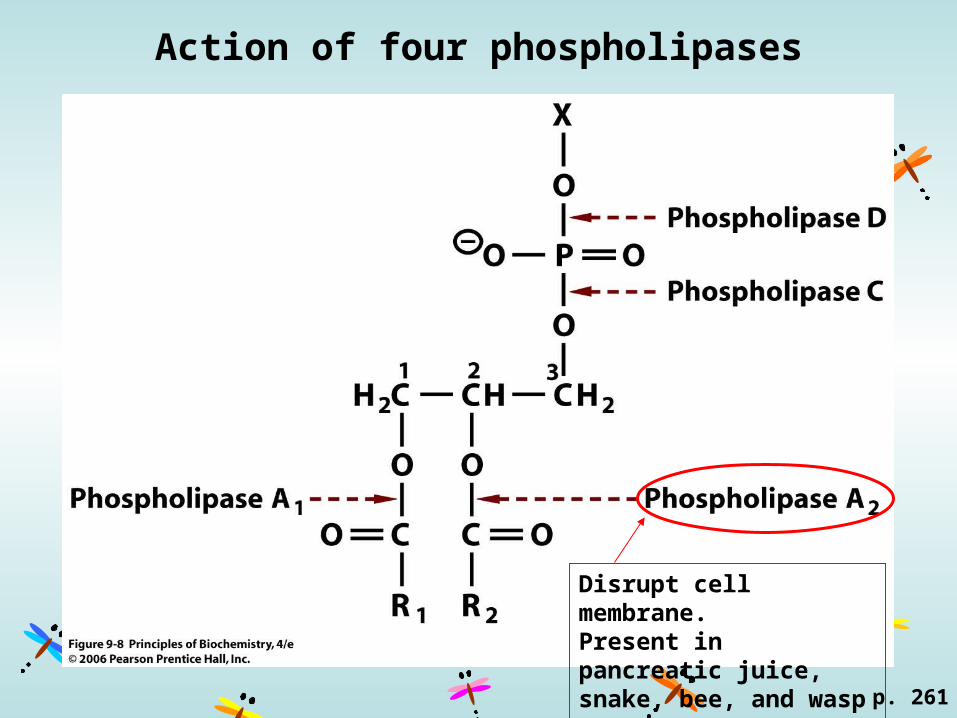
Action of four phospholipases
p. 261
Disrupt cell membrane.Present in pancreatic juice, snake, bee, and wasp venom.

Structure of an ethanolamine plasmalogen
p. 262
Ethanolamine and coline are the commonly esterified to plasmalogen.
About 23% of the glycerophospho-lipid in the human central nerve system, also found in the membrane of peripheral nerve and muscle.
Ethanolamine

Sphingolipids

Structures of sphingosine, ceramide, and sphingomyelin.
p. 262
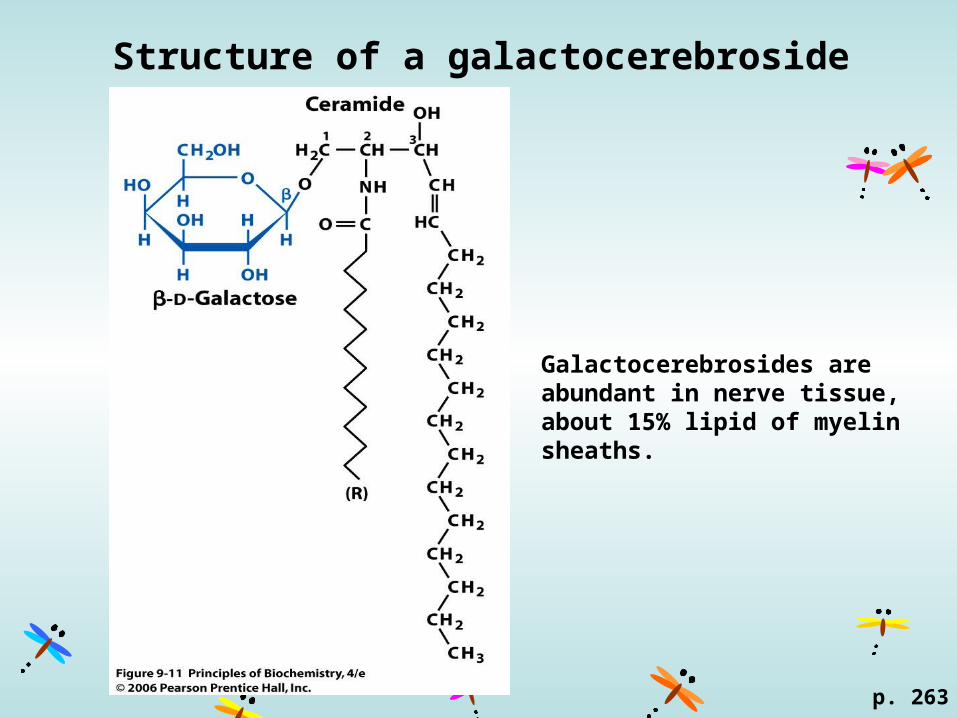
Structure of a galactocerebroside
p. 263
Galactocerebrosides are abundant in nerve tissue, about 15% lipid of myelin sheaths.
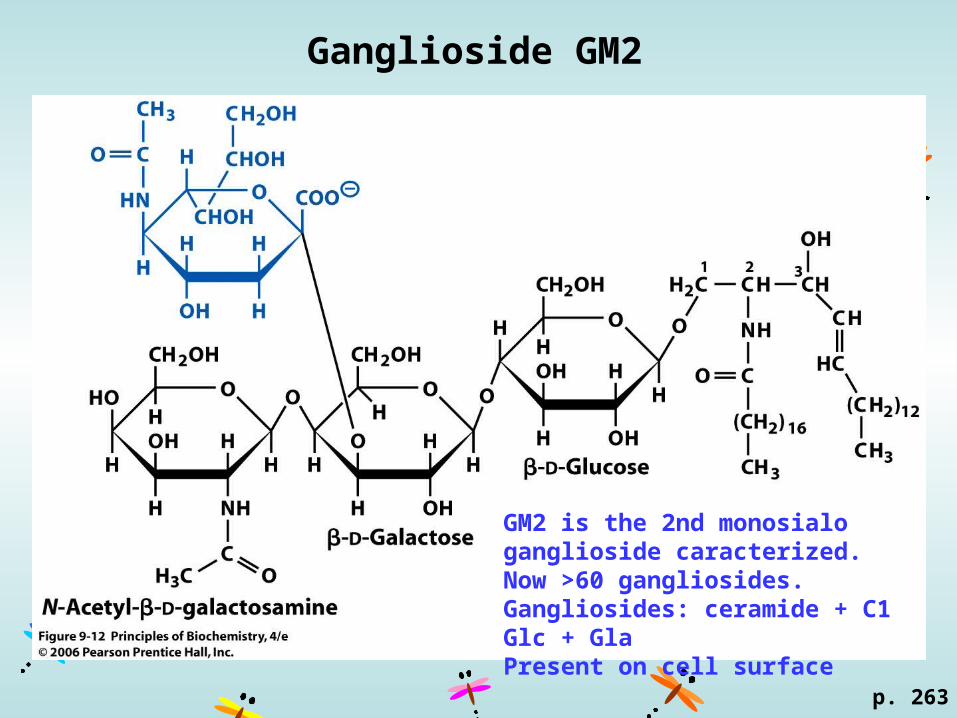
Ganglioside GM2
p. 263
GM2 is the 2nd monosialo ganglioside caracterized.Now >60 gangliosides.Gangliosides: ceramide + C1 Glc + GlaPresent on cell surface
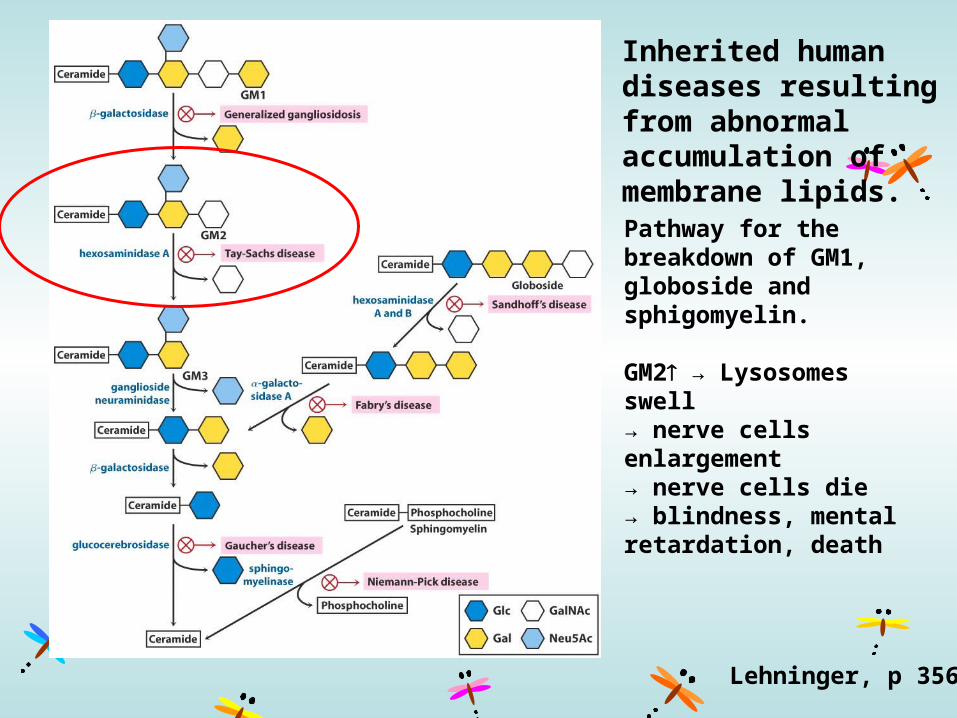
Inherited human diseases resulting from abnormal accumulation of membrane lipids.
Pathway for the breakdown of GM1, globoside and sphigomyelin.
GM2 → Lysosomes swell→ nerve cells enlargement→ nerve cells die→ blindness, mental retardation, death
Lehninger, p 356
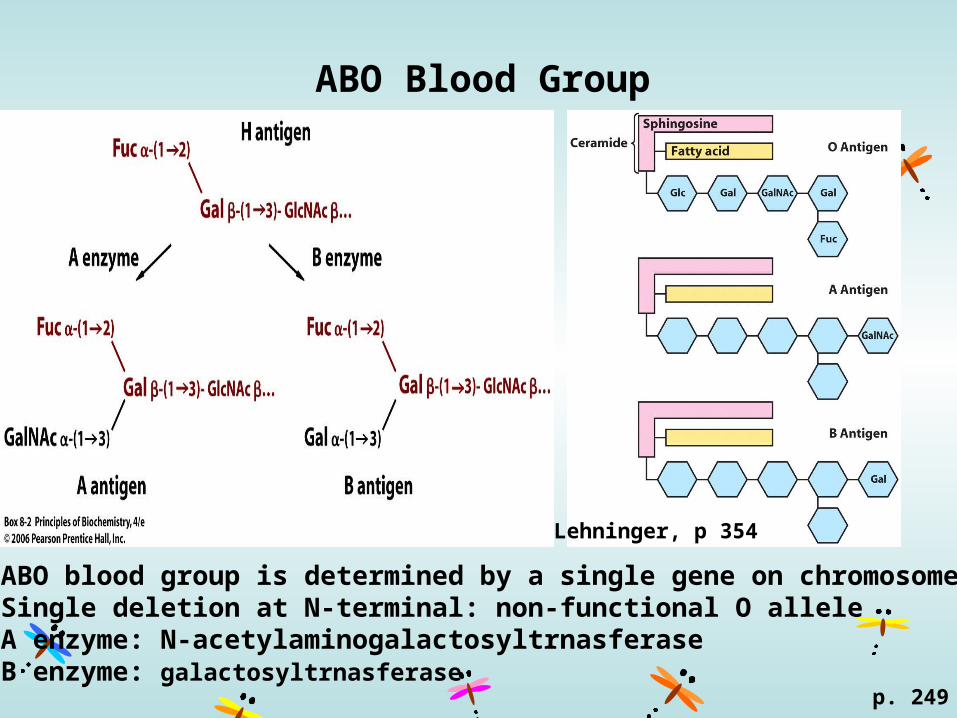
ABO Blood Group
p. 249
ABO blood group is determined by a single gene on chromosome 9.Single deletion at N-terminal: non-functional O alleleA enzyme: N-acetylaminogalactosyltrnasferaseB enzyme: galactosyltrnasferase
Lehninger, p 354

Steroids
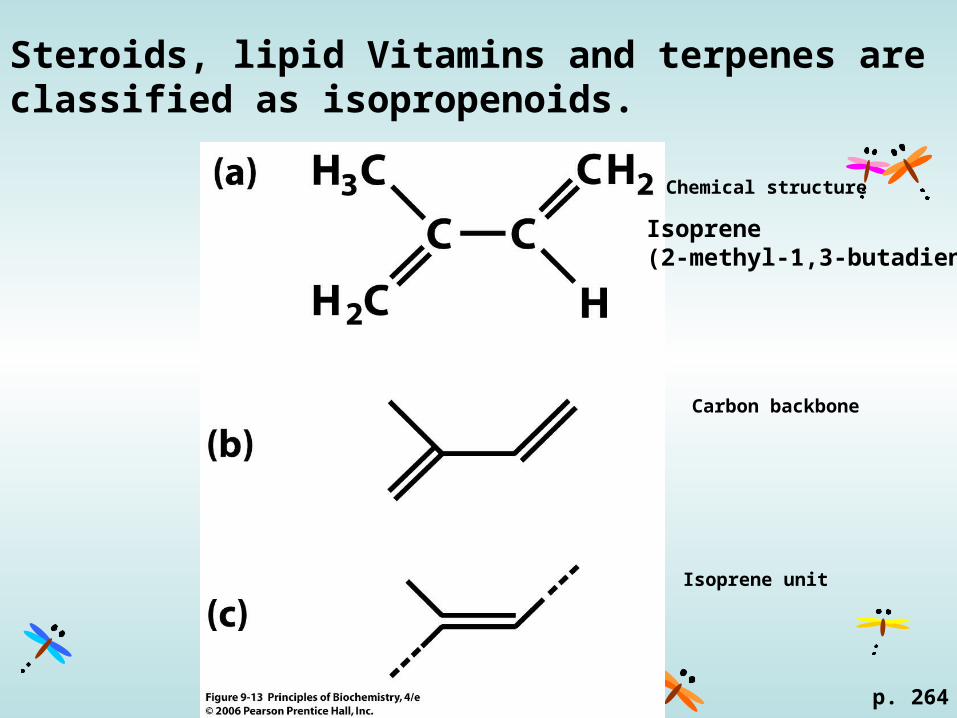
Steroids, lipid Vitamins and terpenes are classified as isopropenoids.
Chemical structure
Carbon backbone
Isoprene unit
p. 264
Isoprene(2-methyl-1,3-butadiene)
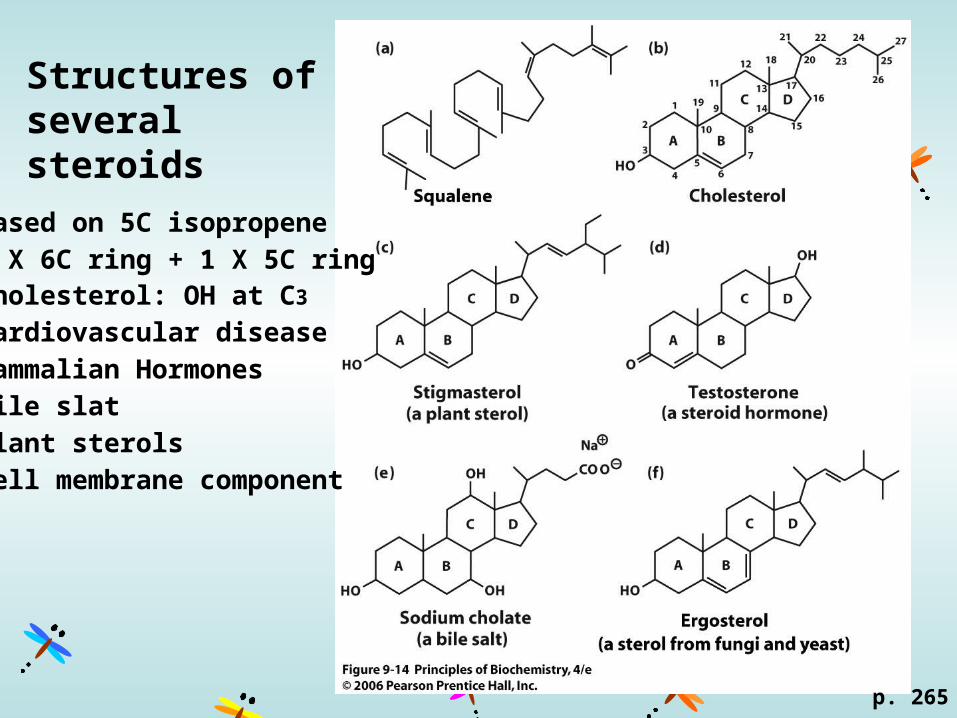
Structures of several steroids
p. 265
Based on 5C isopropene
3 X 6C ring + 1 X 5C ringCholesterol: OH at C3
Cardiovascular disease
Mammalian Hormones
Bile slat
Plant sterols
Cell membrane component
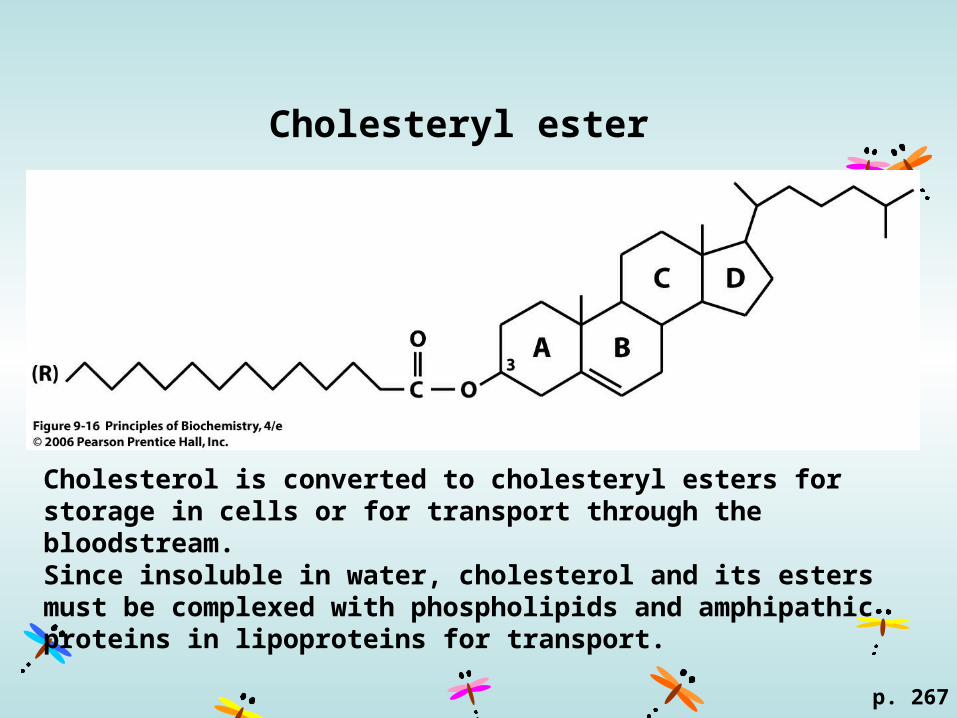
Cholesteryl ester
p. 267
Cholesterol is converted to cholesteryl esters for storage in cells or for transport through the bloodstream. Since insoluble in water, cholesterol and its esters must be complexed with phospholipids and amphipathic proteins in lipoproteins for transport.
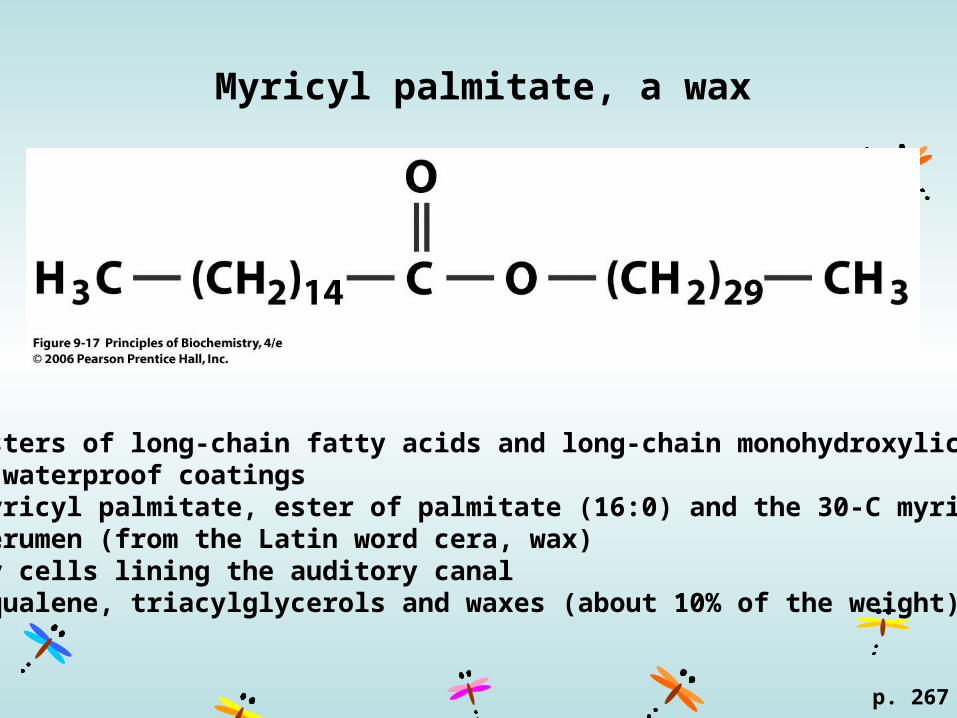
Myricyl palmitate, a wax
p. 267
Waxesnonpolar esters of long-chain fatty acids and long-chain monohydroxylic alcoholsprotective waterproof coatings Beeswax, myricyl palmitate, ester of palmitate (16:0) and the 30-C myricyl alcohol Ear wax, cerumen (from the Latin word cera, wax)secreted by cells lining the auditory canalcontains squalene, triacylglycerols and waxes (about 10% of the weight).
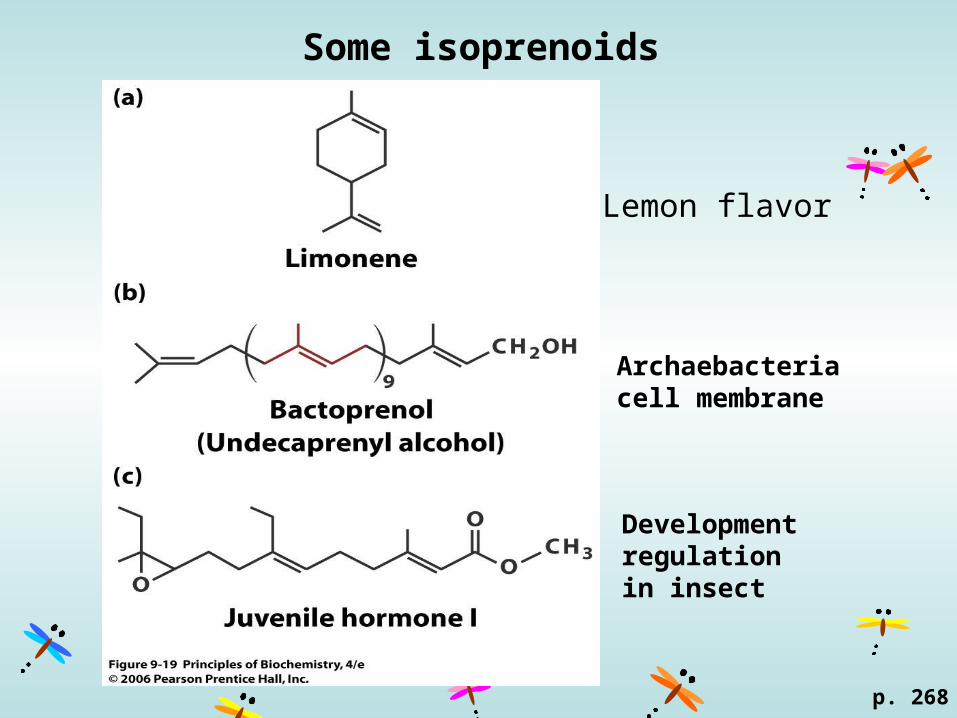
Some isoprenoids
p. 268
Lemon flavor
Archaebacteria cell membrane
Development regulation in insect
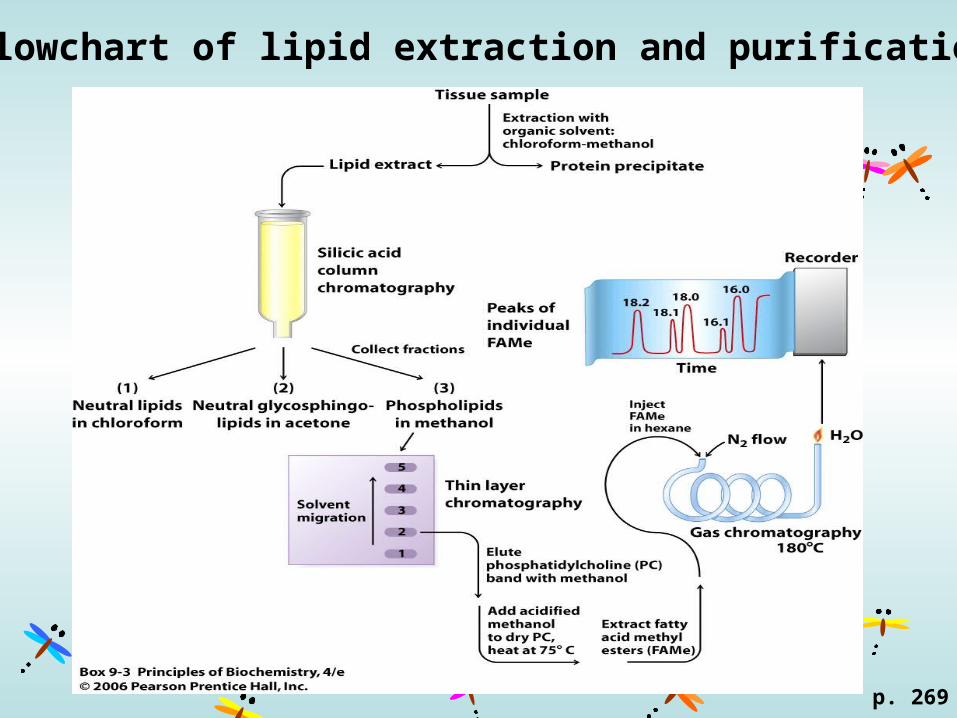
Flowchart of lipid extraction and purification
p. 269

Biological membrane
•Lipid bilayers and proteins •External boundaries of cells•Separate compartments within cells•Selective pumps: transport of ions and small molecules•Generating and maintaining the proton concentration gradients for ATP production•Recognize extracellular signals and communicate them to the cell interior.
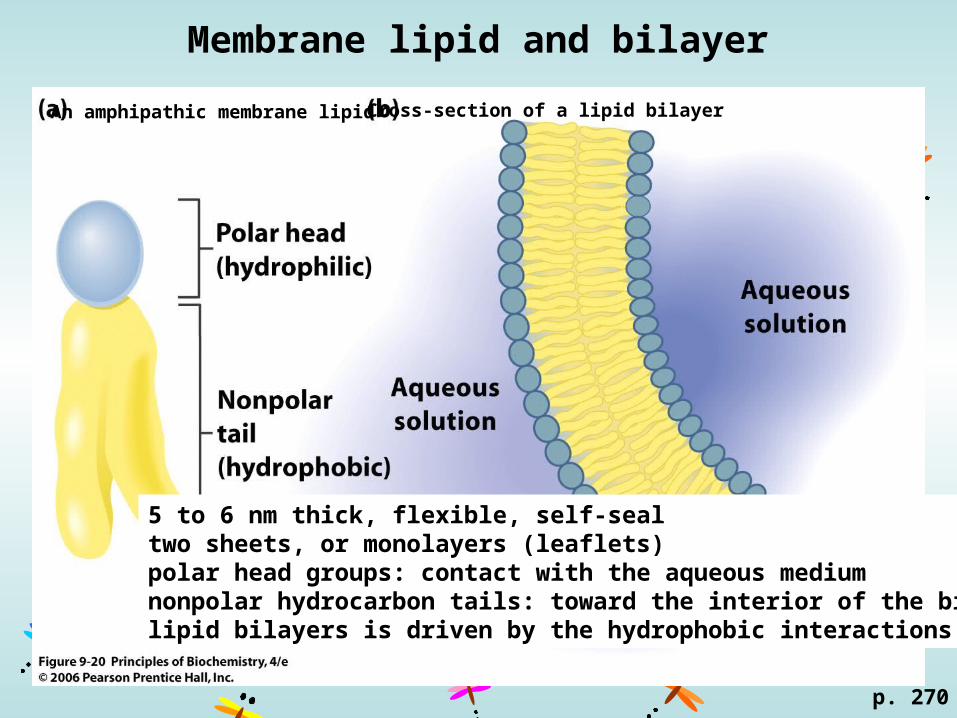
Membrane lipid and bilayer
An amphipathic membrane lipid Cross-section of a lipid bilayer
p. 270
5 to 6 nm thick, flexible, self-seal two sheets, or monolayers (leaflets)polar head groups: contact with the aqueous mediumnonpolar hydrocarbon tails: toward the interior of the bilayerlipid bilayers is driven by the hydrophobic interactions

Typical biological membrane
6-10 nmLipids: 25%~50%Proteins: 50%~75%Carbohydrate: 10%
Cholesterol and some other liplds: ~30% of total lipid
Components of inner and outer membranes are different. Red blood cells: rich in proteinsBrain cells: rich in phosphotidylserinesE. coli: inner-70% phosphatidylethanolamines
outer-lipopolysaccharidesMammalian: outer surface-90% sphingomylin
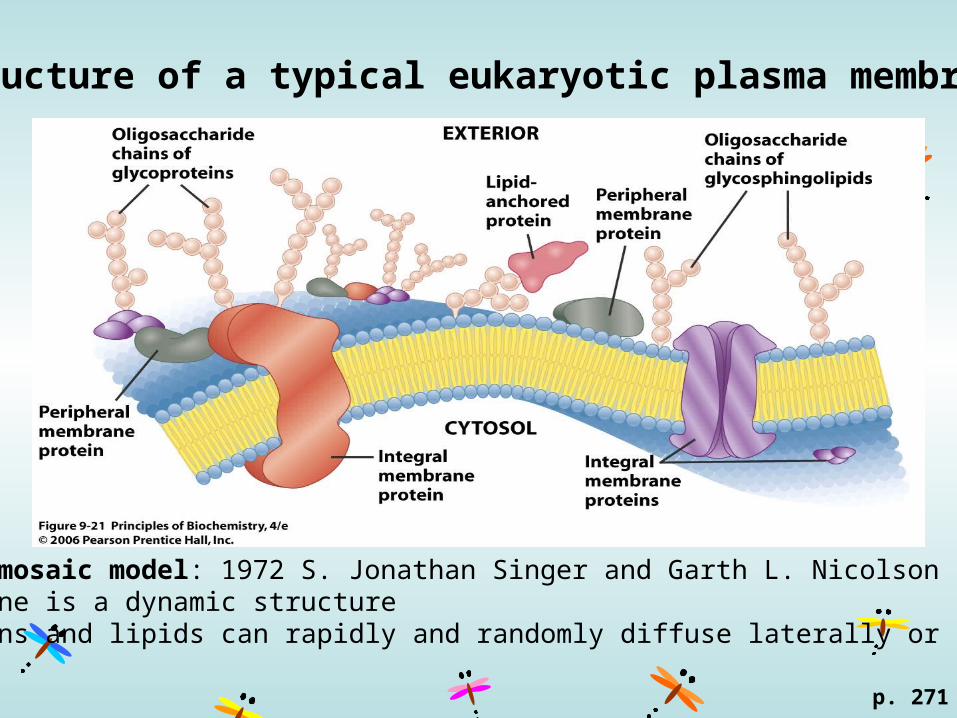
Structure of a typical eukaryotic plasma membrane
p. 271
fluid mosaic model: 1972 S. Jonathan Singer and Garth L. Nicolson membrane is a dynamic structure proteins and lipids can rapidly and randomly diffuse laterally or rotate
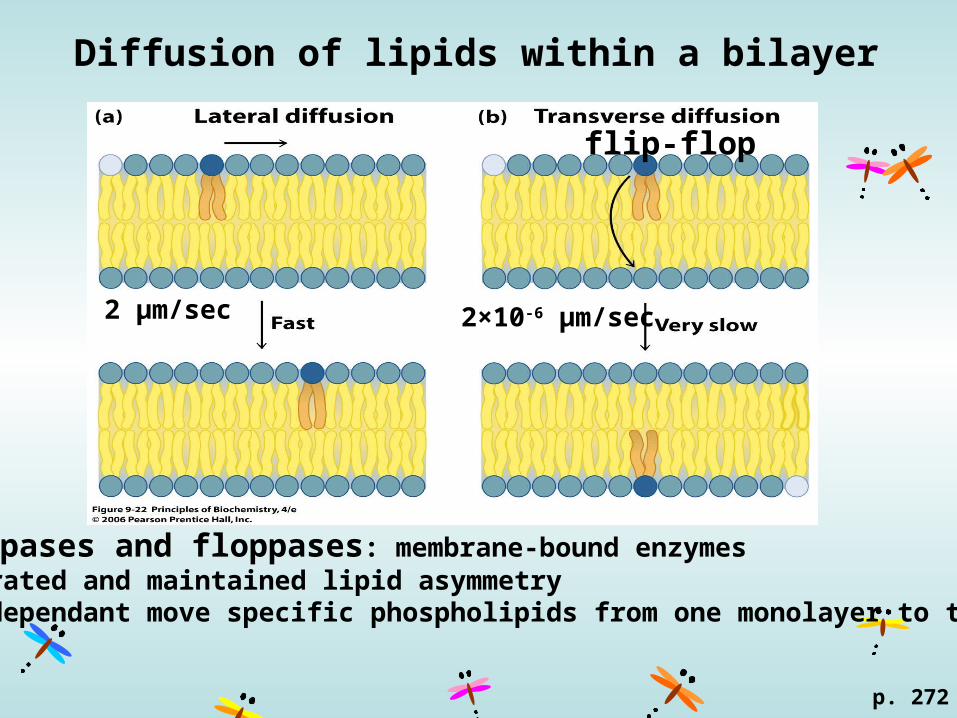
Diffusion of lipids within a bilayer
p. 272
2 μm/sec
flip-flop
2×10-6 μm/sec
flippases and floppases: membrane-bound enzymesgenerated and maintained lipid asymmetry ATP-dependant move specific phospholipids from one monolayer to the other
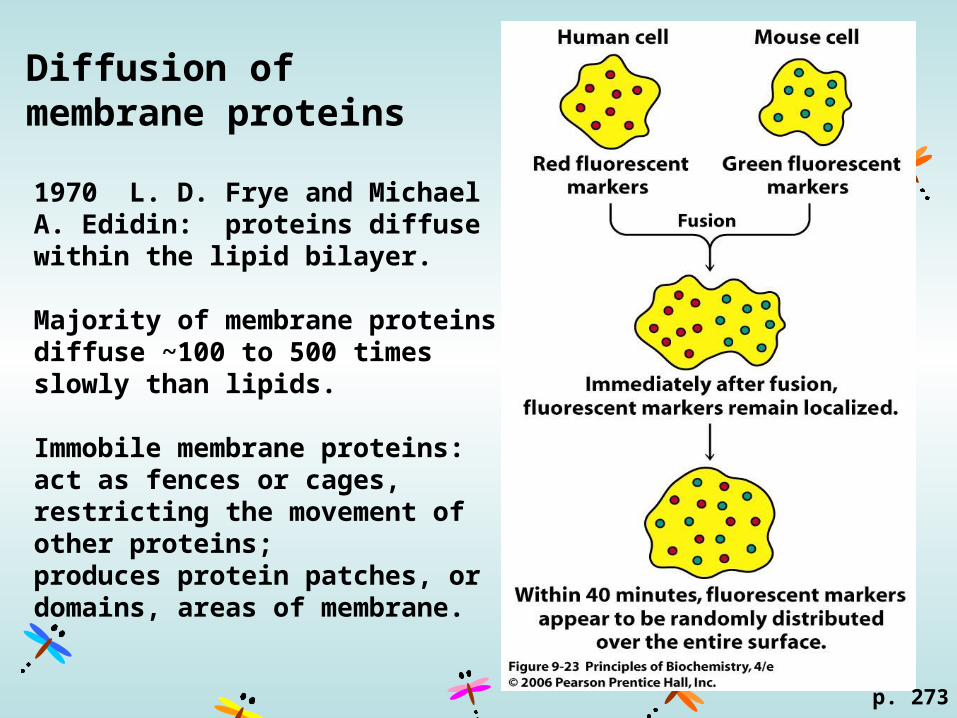
Diffusion of membrane proteins
p. 273
1970 L. D. Frye and Michael A. Edidin: proteins diffuse within the lipid bilayer.
Majority of membrane proteins diffuse ~100 to 500 times slowly than lipids.
Immobile membrane proteins: act as fences or cages, restricting the movement of other proteins; produces protein patches, or domains, areas of membrane.
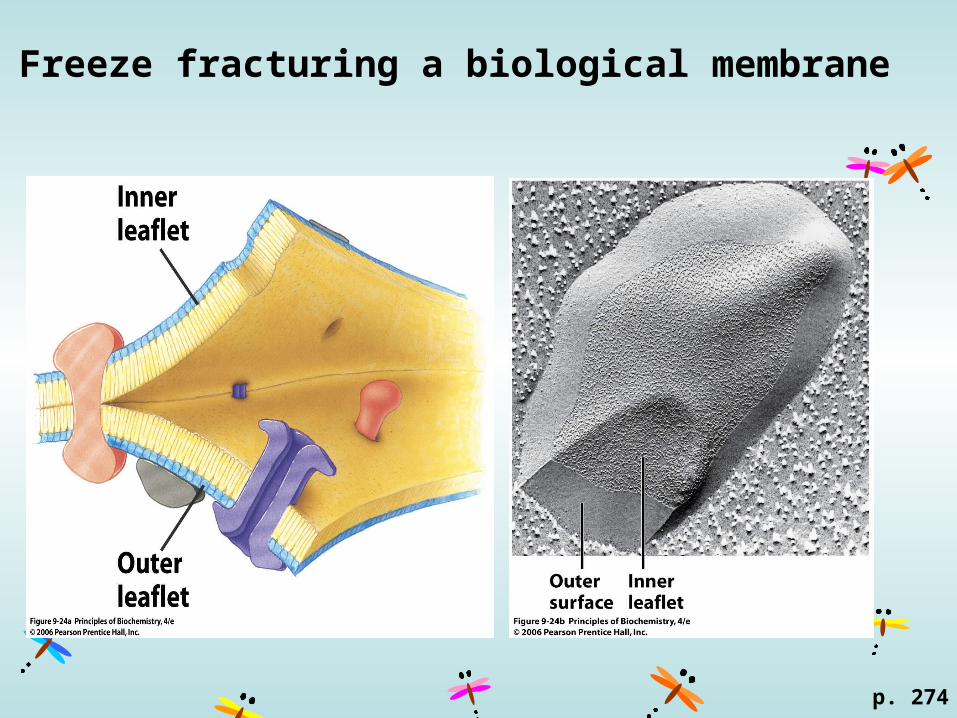
Freeze fracturing a biological membrane
p. 274
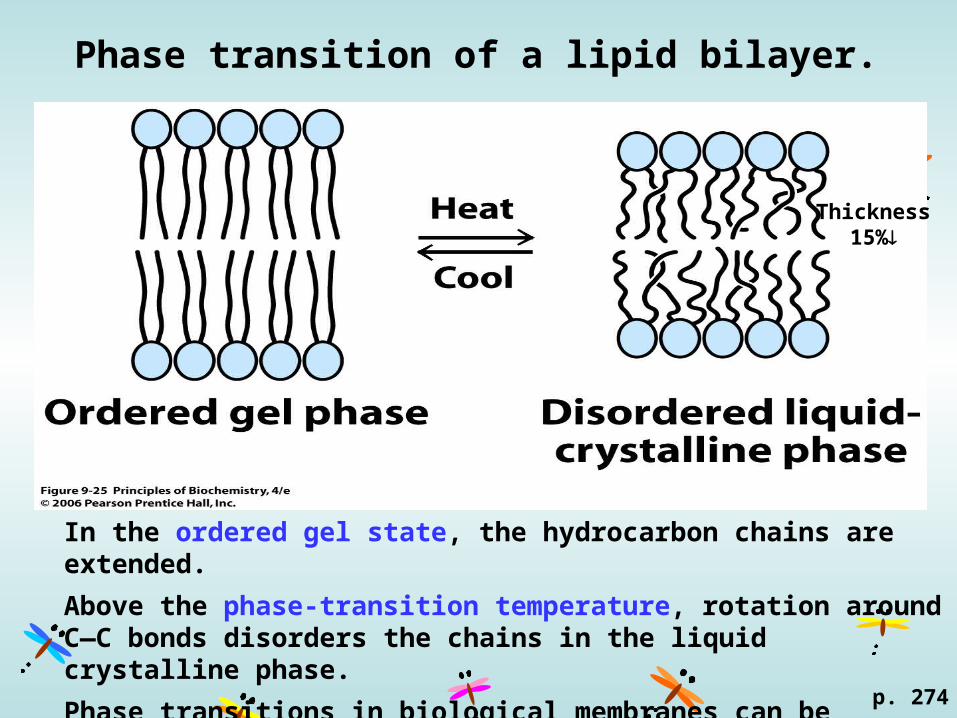
Phase transition of a lipid bilayer.
p. 274
In the ordered gel state, the hydrocarbon chains are extended.
Above the phase-transition temperature, rotation around C—C bonds disorders the chains in the liquid crystalline phase.
Phase transitions in biological membranes can be localized, so fluid- and gel-phase regions can coexist at certain temperatures.
Thickness15%

Fluidity and phasetransition temperature
The structure of a phospholipid: incorporating an unsaturated fatty acyl group into a phospholipid lowers the phase-transition temperature.
Cholesterol accounts for 20% to 25% of the mass of lipids in a typical mammalian plasma membrane and significantly affects membrane fluidity.lipid rafts:Cholestreol and sphingolipidMaintained by membrane proteins
p. 274
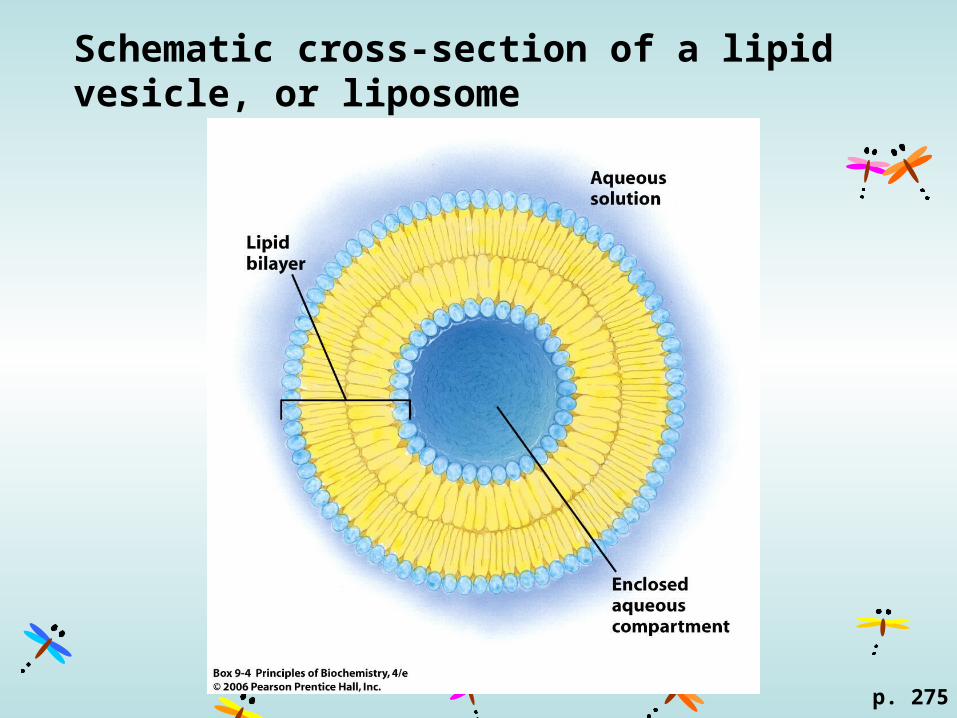
Schematic cross-section of a lipid vesicle, or liposome
p. 275
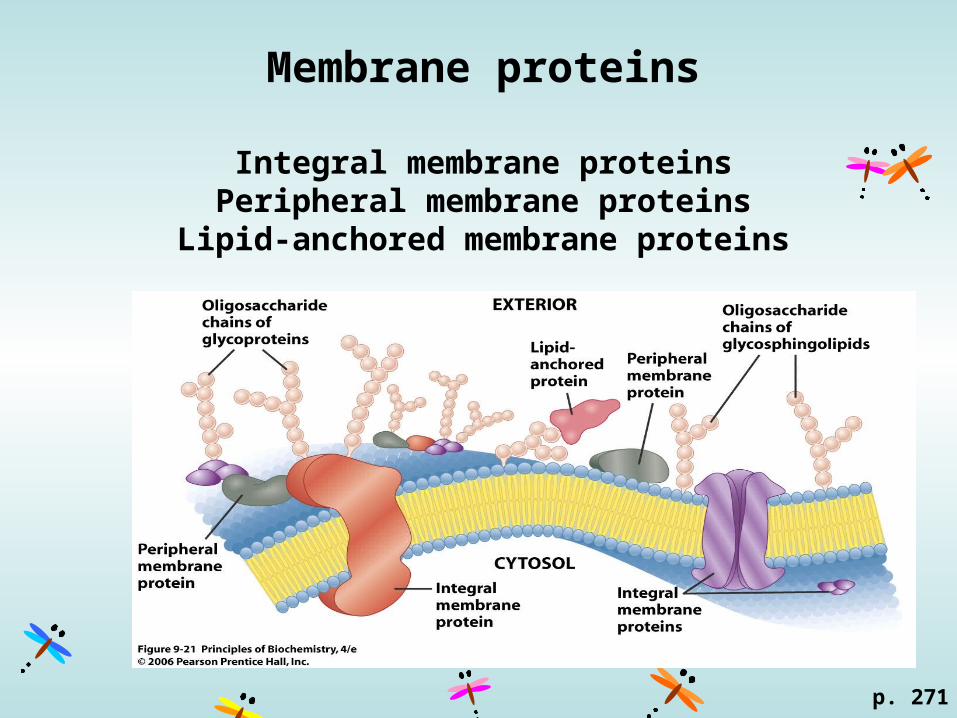
Membrane proteins
Integral membrane proteinsPeripheral membrane proteins
Lipid-anchored membrane proteins
p. 271

Bacteriorhodopsin from Halobacterium salinarumHarness light energy for ATP synthesis
p. 276
Integral membrane proteins (transmembrane proteins)Hydrophobic core (transmembrane region) + exposed proteins on the surface.
-helix~20 aa
loops
light-harvesting prosthetic group (in yellow)
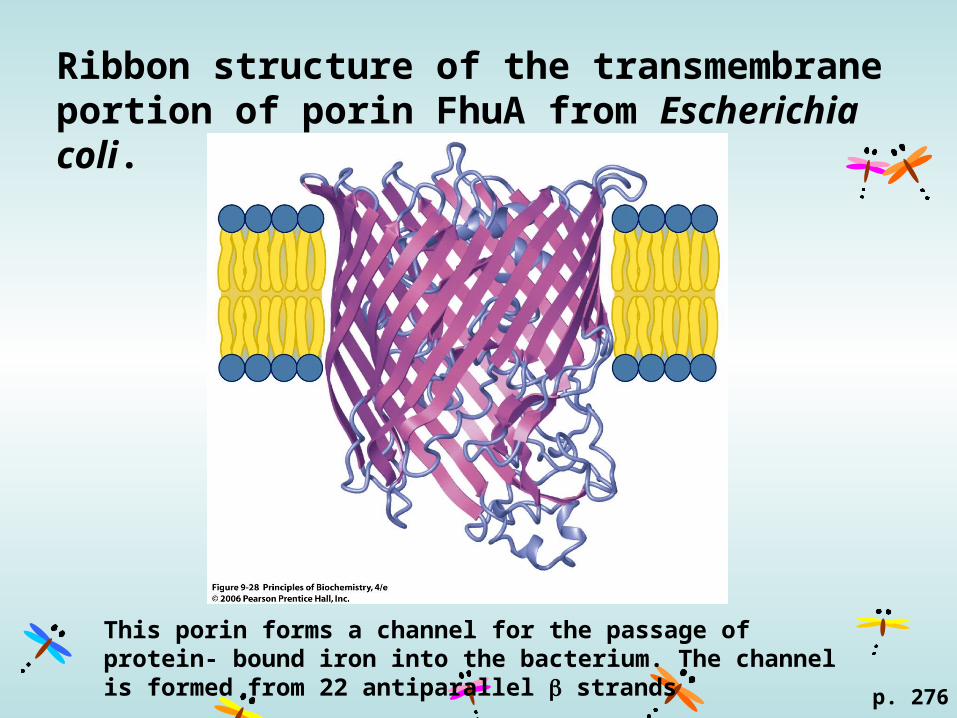
Ribbon structure of the transmembrane portion of porin FhuA from Escherichia coli.
p. 276
This porin forms a channel for the passage of protein- bound iron into the bacterium. The channel is formed from 22 antiparallel strands

Peripheral membrane proteins
Associated with one face of the membrane through charge–charge interactions and hydrogen bonding with integral membrane proteins or with the polar head groups of membrane lipids.
A change in pH or ionic strength is often sufficient to remove these proteins from the membrane.
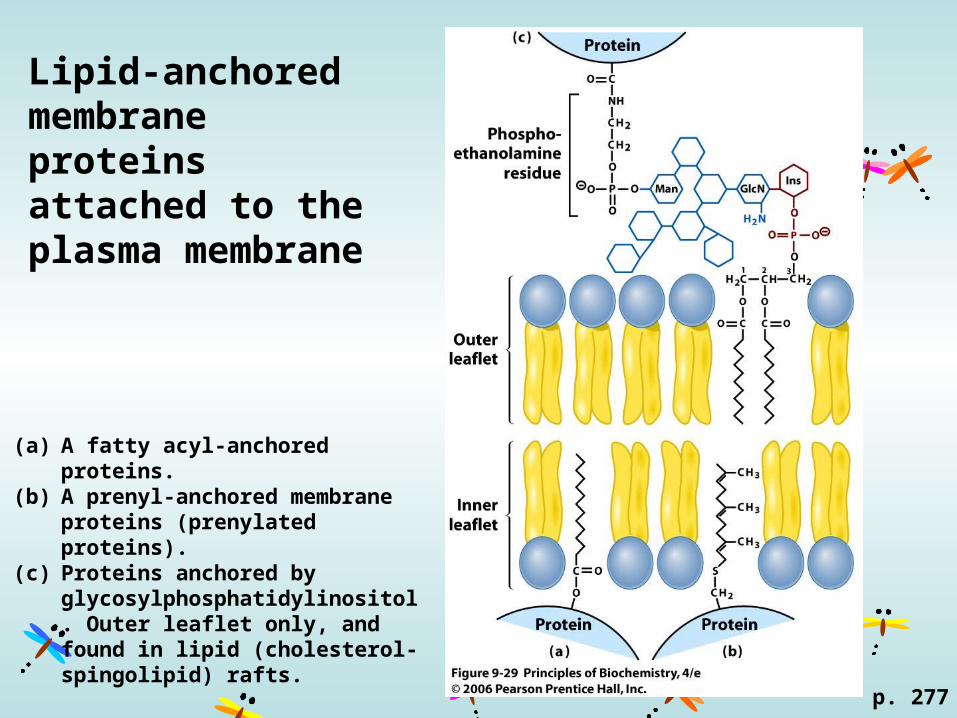
Lipid-anchored membrane proteins attached to the plasma membrane
(a) A fatty acyl-anchored proteins. (b) A prenyl-anchored membrane
proteins (prenylated proteins).(c) Proteins anchored by
glycosylphosphatidylinositol. Outer leaflet only, and found in lipid (cholesterol-spingolipid) rafts.
p. 277

p. 278
Membrane Transport
(Non-polar gases, Hydrophobic molecules)

Thermodynamics of Membrane Transport
Aout → Ain at 25°C, [Ain]=1mM, [Aout]=100mM

Thermodynamics of Membrane TransportCharge compounds
: membrane potential (in volts)F : Faradays’s constant (96.485 kJV-1 mol-1)z: charge on the molecule being transported

Membrane transport through a pore or channel
p. 280
pore: for bacteriachannel: for animals
No energy neededNot substrate saturableRate id diffusion-controlled
Some always open.Some open or close response to signal.
In nerve tissue: gated potassium channel rapid outward transport of K+ 10,000 times faster than transport of Na+.
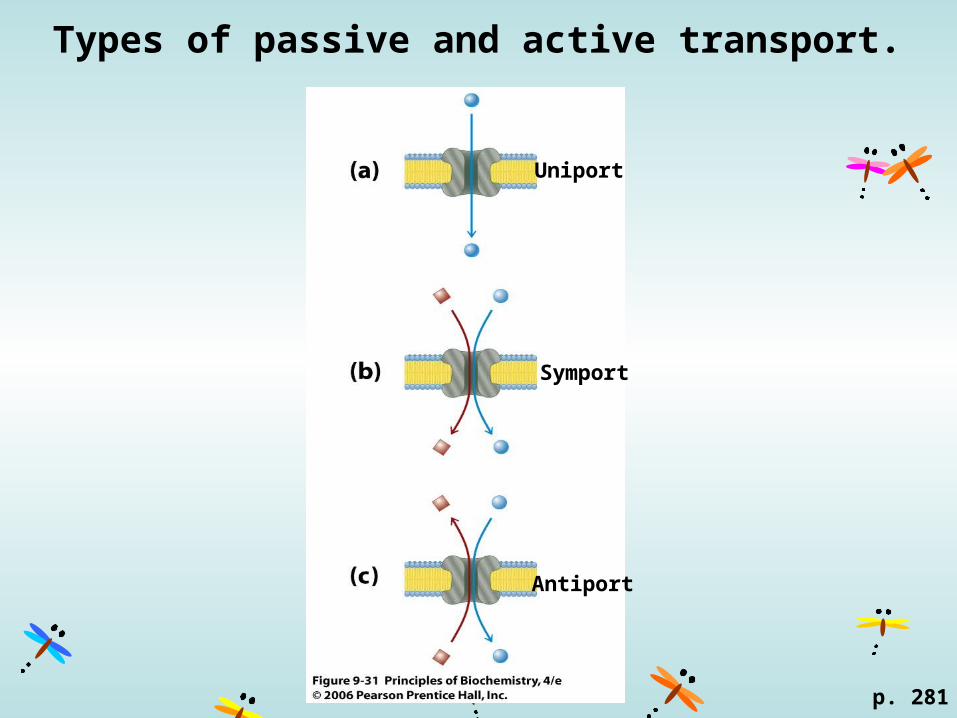
Types of passive and active transport.
Uniport
Symport
Antiport
p. 281
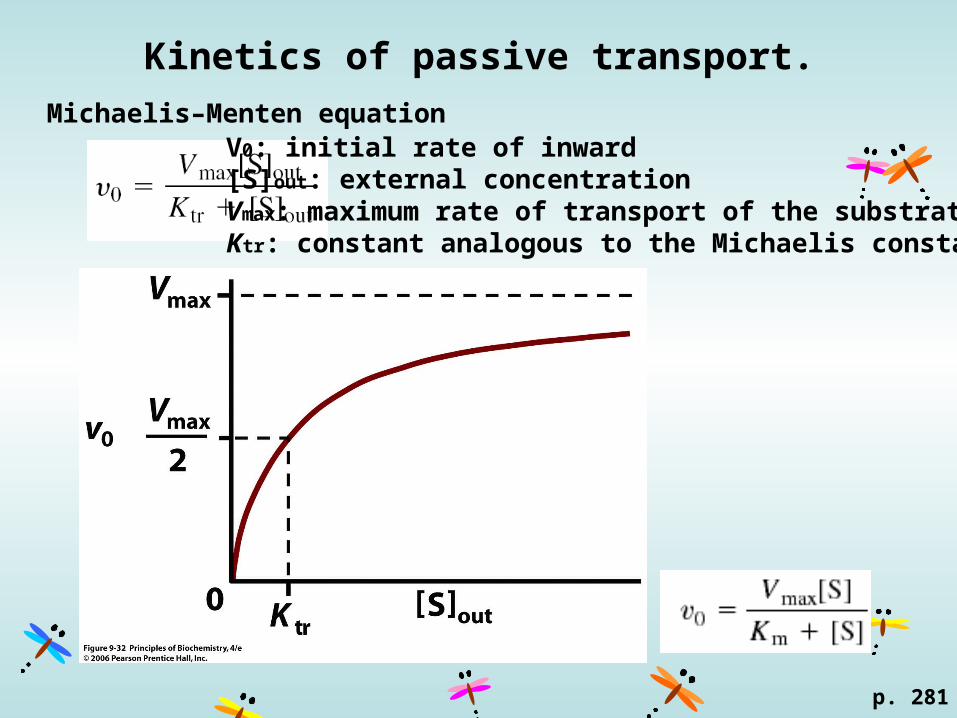
Kinetics of passive transport.
p. 281
Michaelis–Menten equationV0: initial rate of inward [S]out: external concentration Vmax: maximum rate of transport of the substrateKtr: constant analogous to the Michaelis constant Km
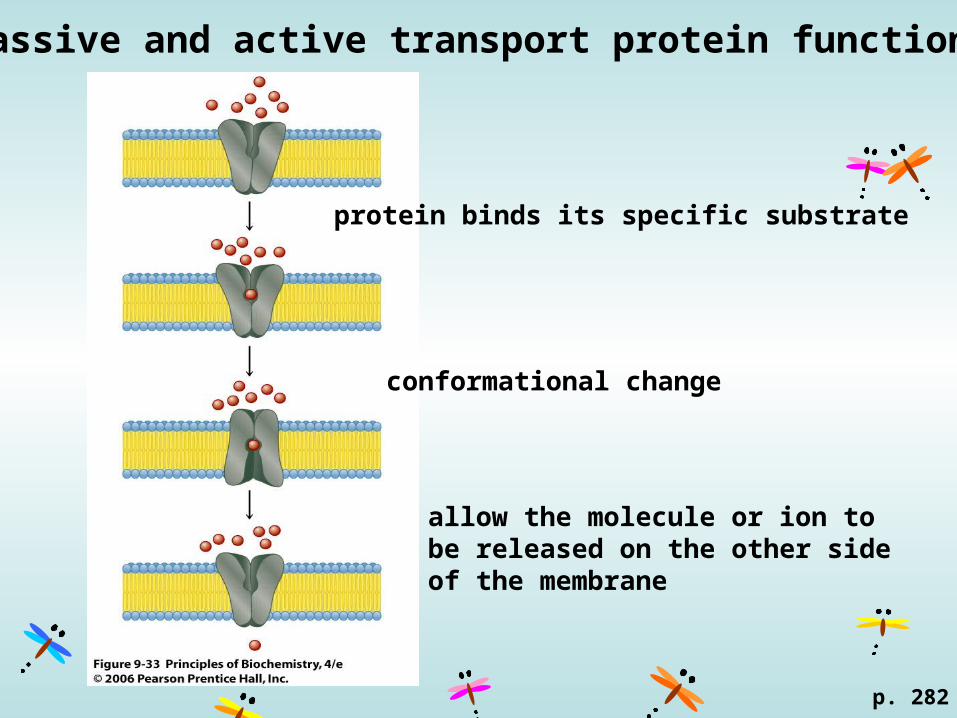
Passive and active transport protein function.
p. 282
protein binds its specific substrate
conformational change
allow the molecule or ion to be released on the other side of the membrane
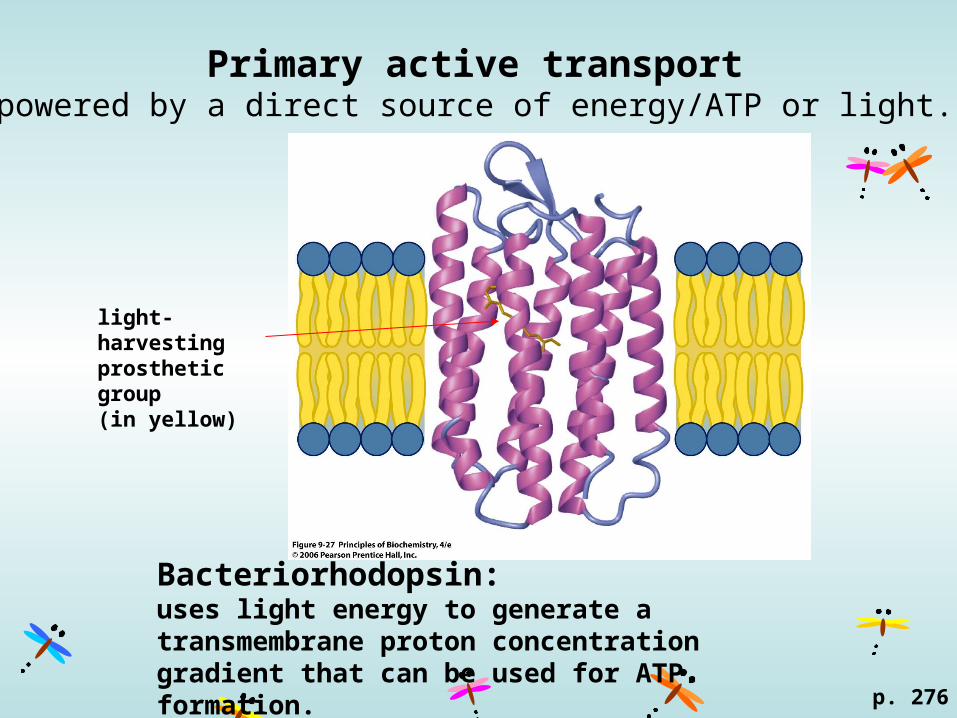
Primary active transportpowered by a direct source of energy/ATP or light.
p. 276
light-harvesting prosthetic group (in yellow)
Bacteriorhodopsin: uses light energy to generate a transmembrane proton concentration gradient that can be used for ATP formation.
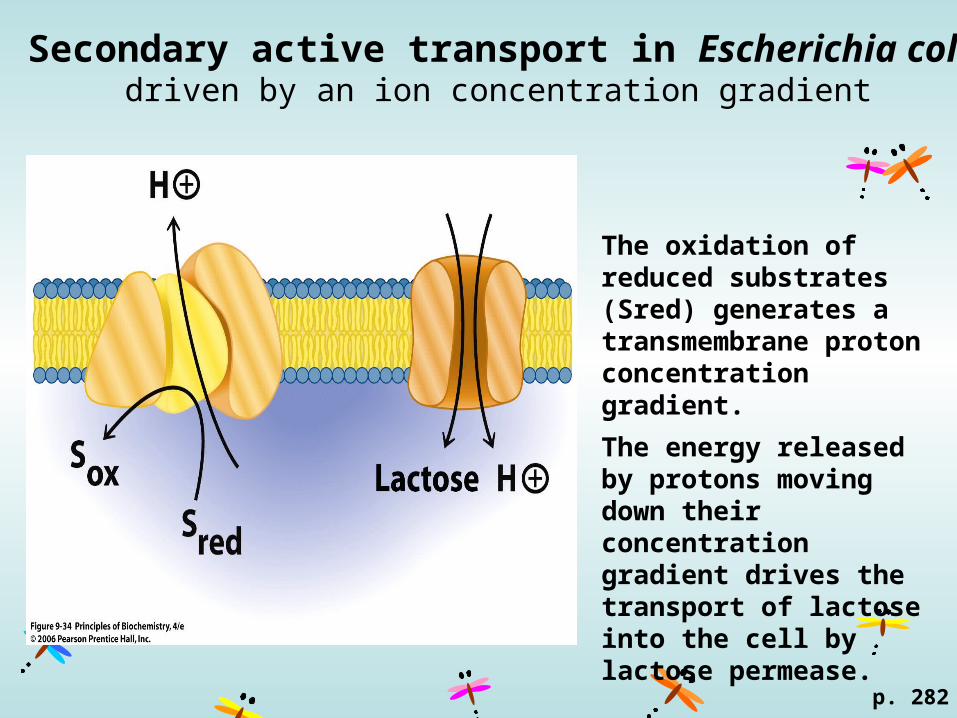
Secondary active transport in Escherichia colidriven by an ion concentration gradient
p. 282
The oxidation of reduced substrates (Sred) generates a transmembrane proton concentration gradient.
The energy released by protons moving down their concentration gradient drives the transport of lactose into the cell by lactose permease.
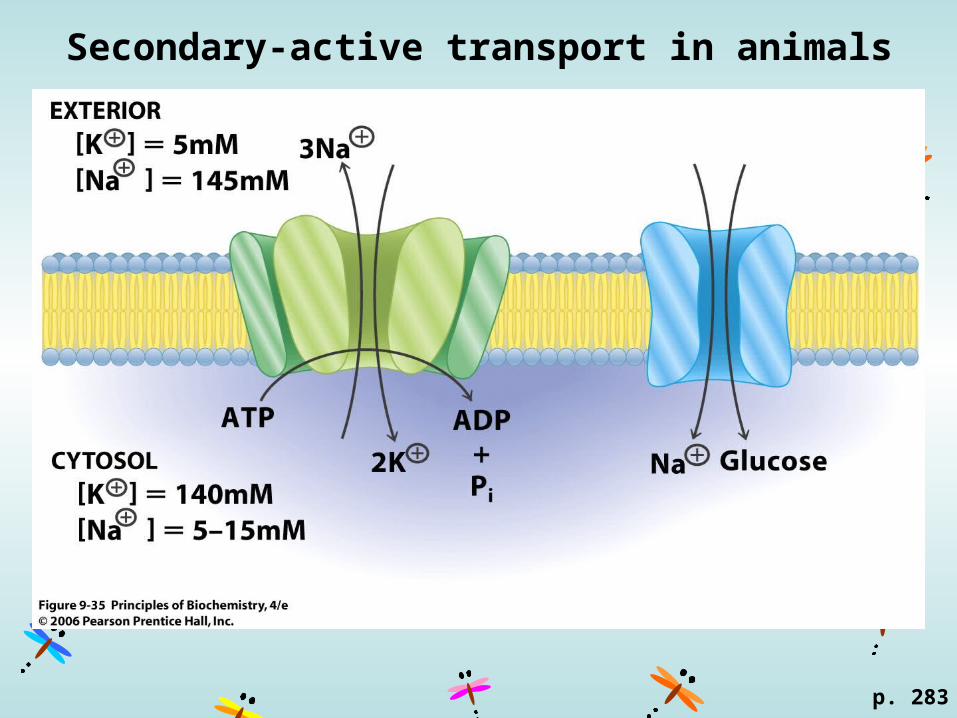
Secondary-active transport in animals
p. 283

Endocytosis and Exocytosis
p. 283
Movement for large molecule, such as proteins.
Receptor-mediated endocytosis: binding of macromolecules to specific receptor proteins in the plasma membrane of the cell, fusing with endosome then with lysosome.
Exocytosis: materials for secretion are enclosed in vesicles by the Golgi apparatus, then fuse with the plasma membrane, releasing the vesicle contents into the extracellular space.

Transduction of Extracellular Signals
Signals:
chemotaxis
hormones
neurotransmitters
growth factors
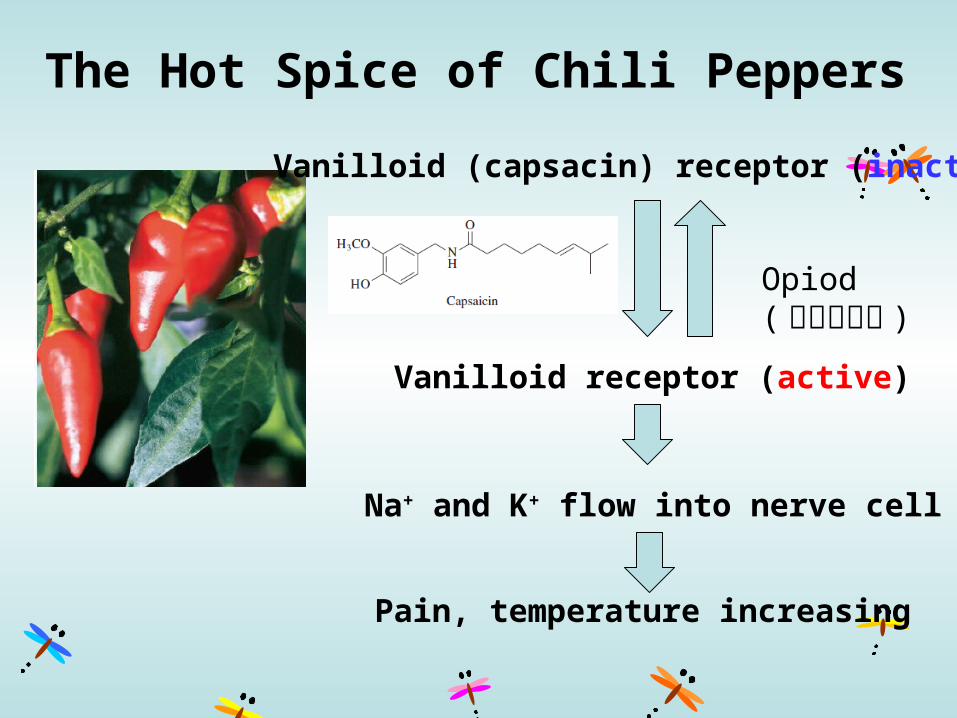
The Hot Spice of Chili Peppers
Pain, temperature increasing
Vanilloid (capsacin) receptor (inactive)
Vanilloid receptor (active)
Na+ and K+ flow into nerve cell
Opiod (內生型鴉片 )
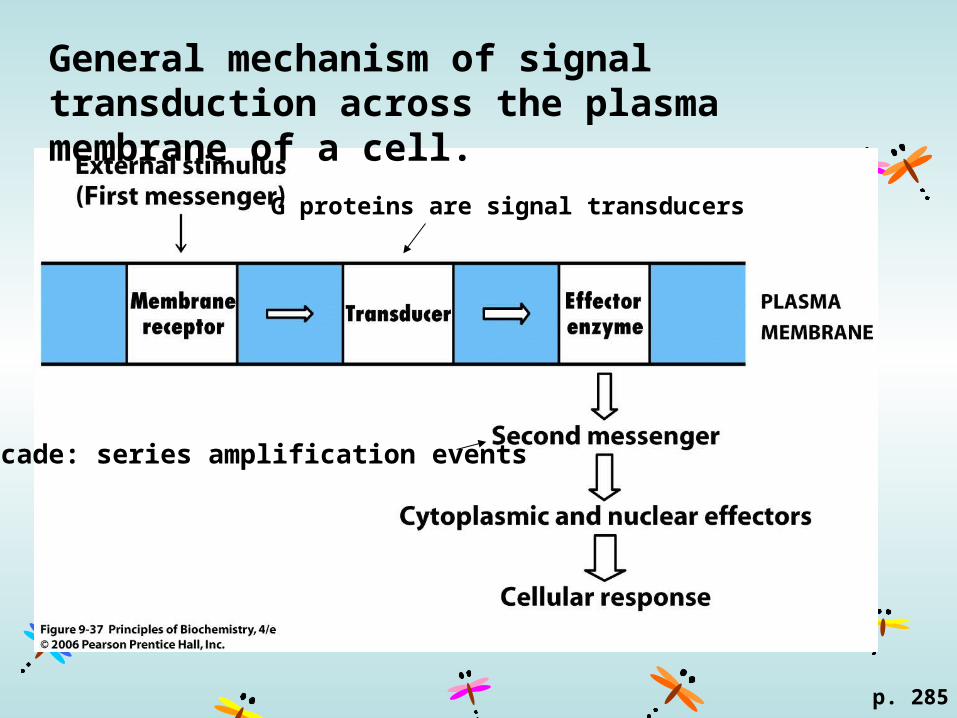
General mechanism of signal transduction across the plasma membrane of a cell.
p. 285
Cascade: series amplification events
G proteins are signal transducers
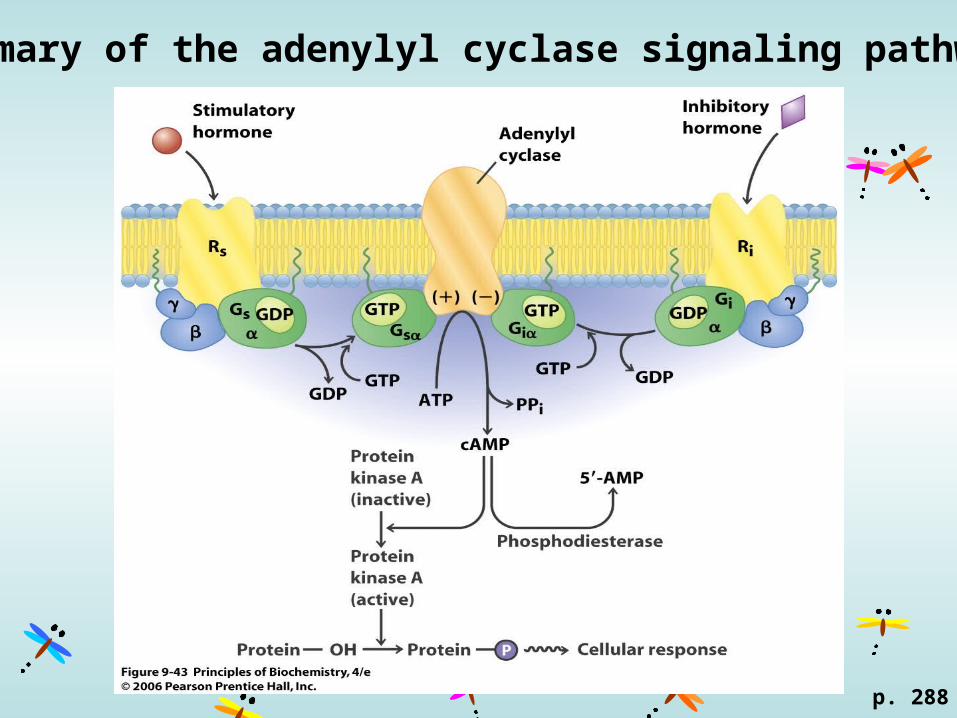
Summary of the adenylyl cyclase signaling pathway
p. 288
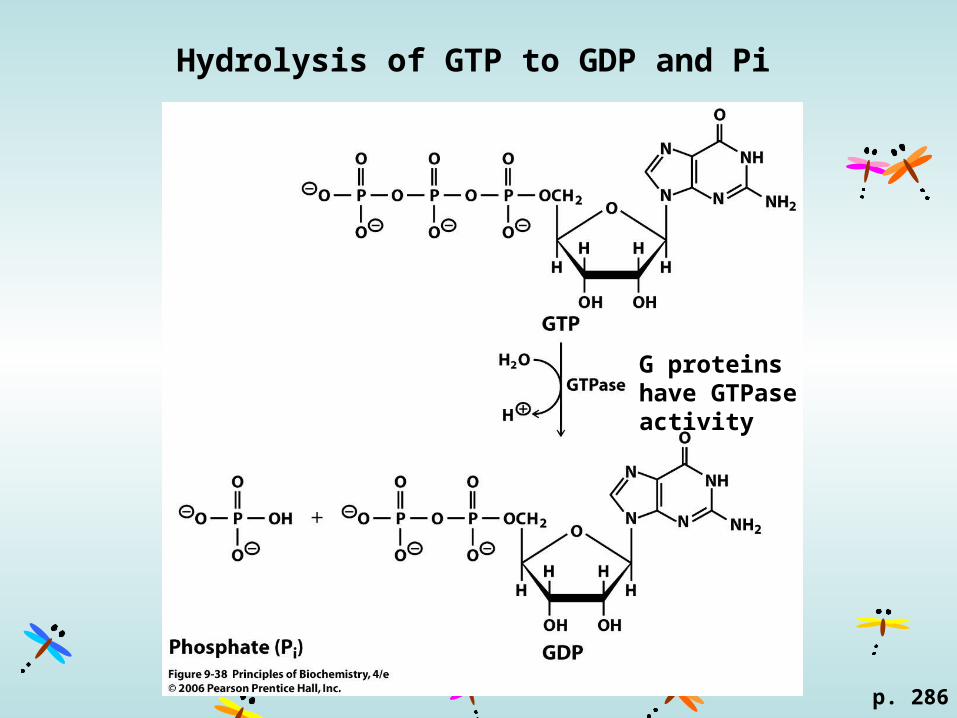
Hydrolysis of GTP to GDP and Pi
p. 286
G proteins have GTPase activity

G-protein cycle
p. 286
: fatty-acyl anchored protein: prenyl-anchored protein
http://upload.wikimedia.org
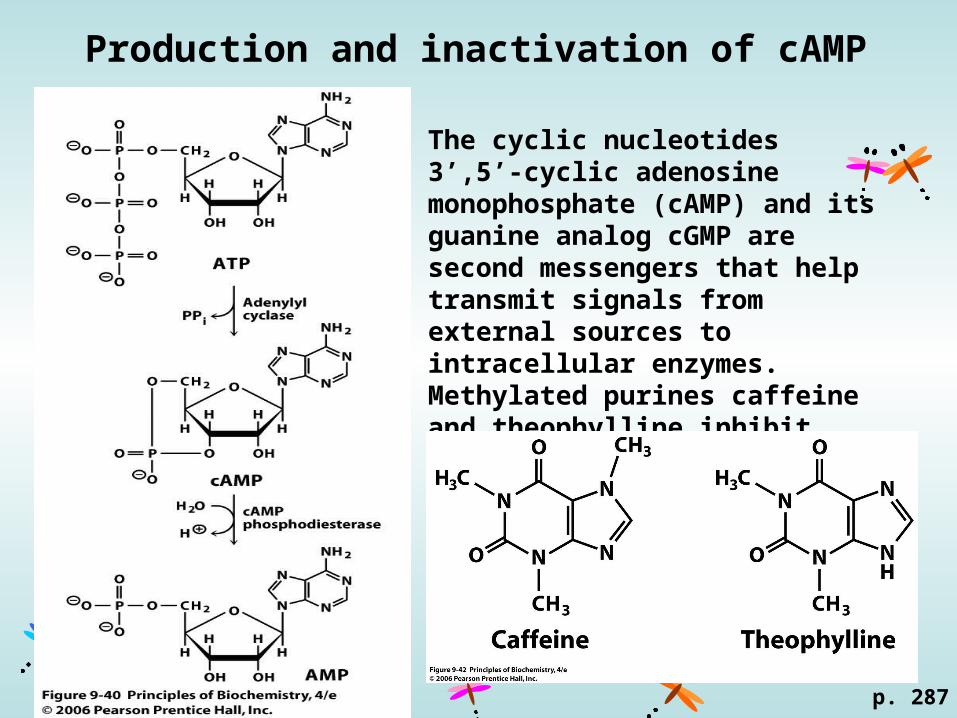
Production and inactivation of cAMP
p. 287
The cyclic nucleotides 3’,5’-cyclic adenosine monophosphate (cAMP) and its guanine analog cGMP are second messengers that help transmit signals from external sources to intracellular enzymes.Methylated purines caffeine and theophylline inhibit cAMP phosphodiesterase.
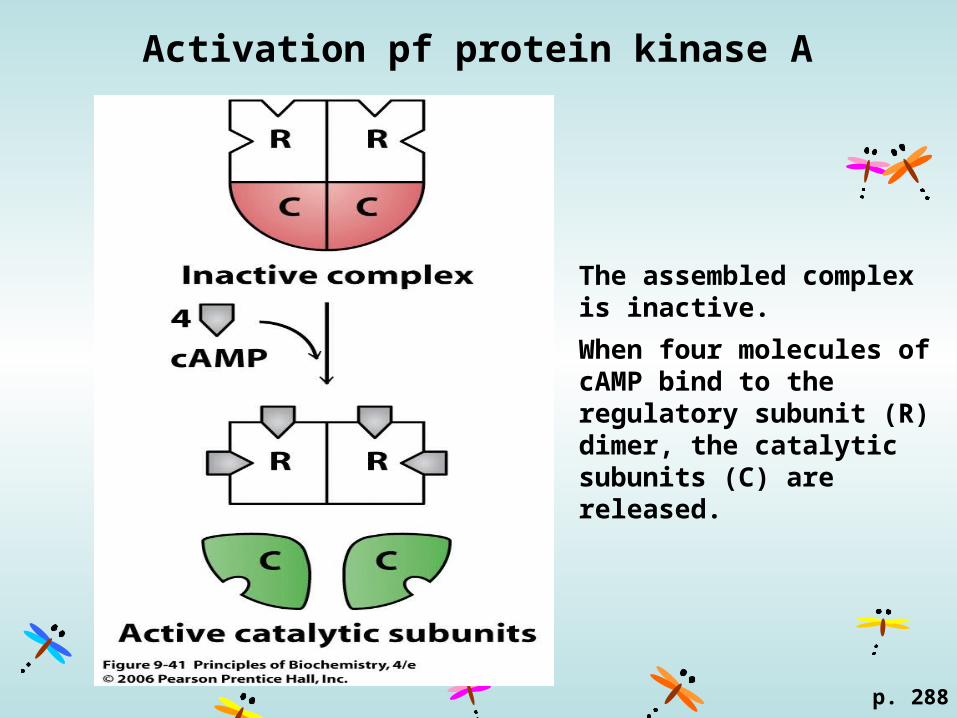
Activation pf protein kinase A
p. 288
The assembled complex is inactive.
When four molecules of cAMP bind to the regulatory subunit (R) dimer, the catalytic subunits (C) are released.
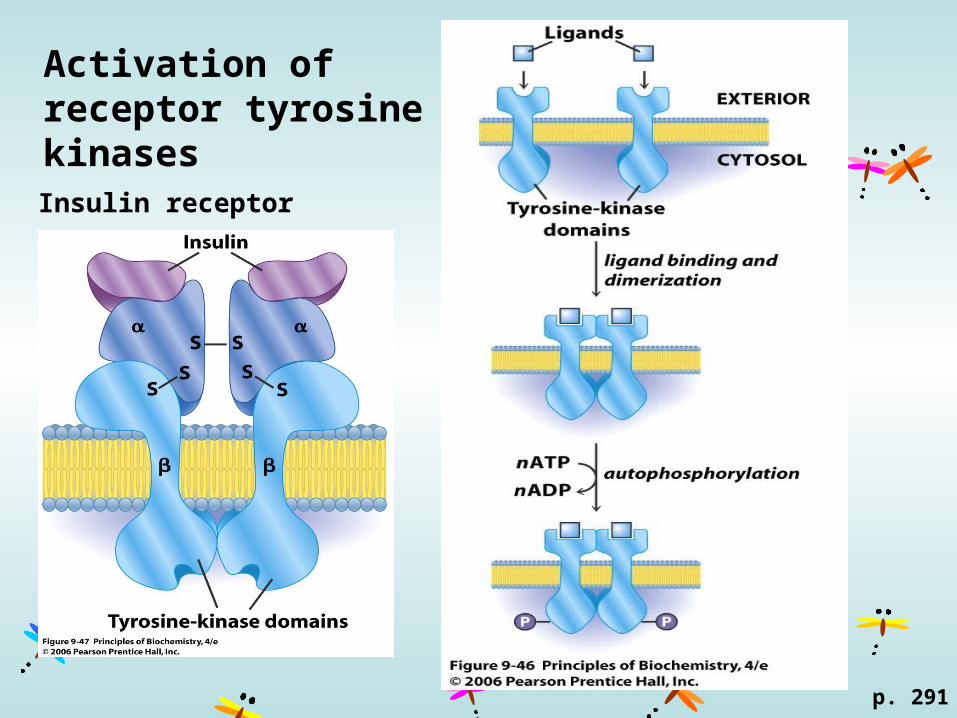
Activation of receptor tyrosine kinases
p. 291
Insulin receptor
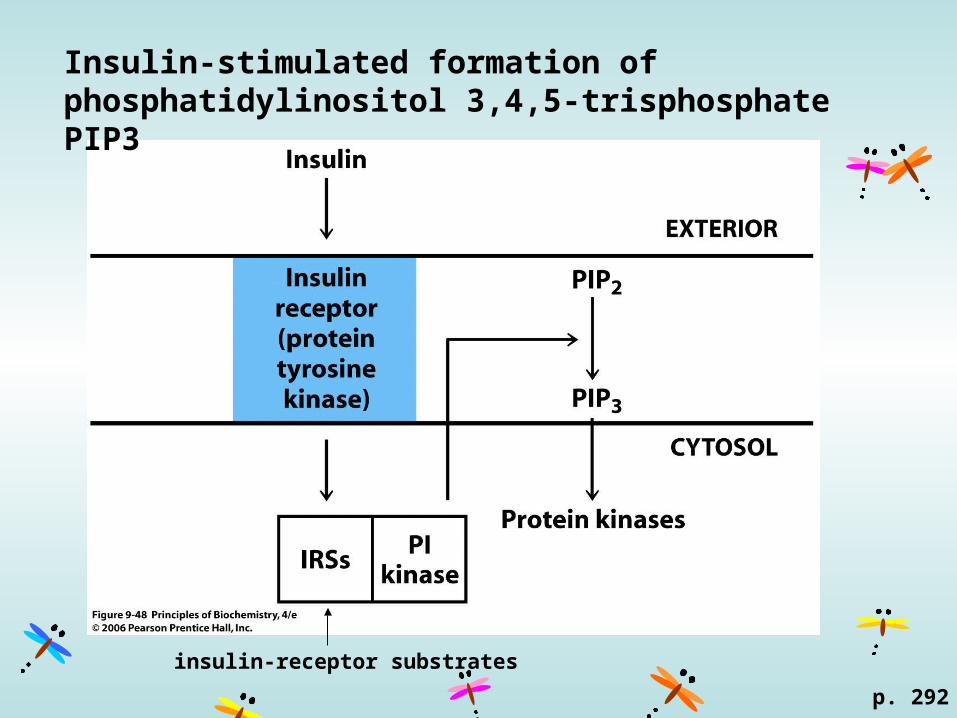
Insulin-stimulated formation of phosphatidylinositol 3,4,5-trisphosphate PIP3
p. 292
insulin-receptor substrates
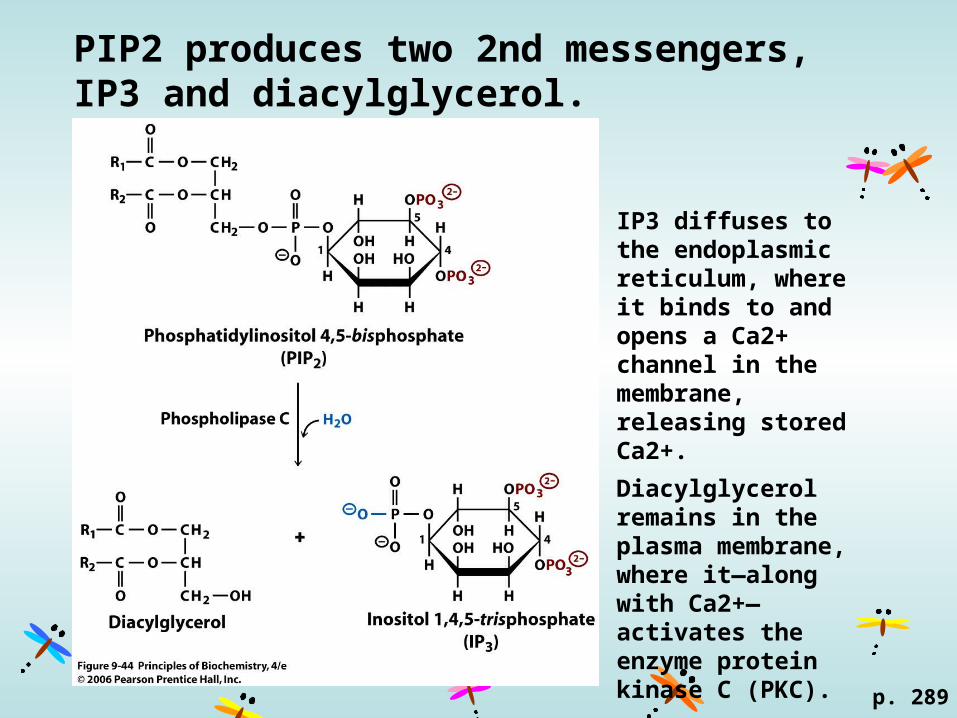
PIP2 produces two 2nd messengers, IP3 and diacylglycerol.
p. 289
IP3 diffuses to the endoplasmic reticulum, where it binds to and opens a Ca2+ channel in the membrane, releasing stored Ca2+.
Diacylglycerol remains in the plasma membrane, where it—along with Ca2+—activates the enzyme protein kinase C (PKC).
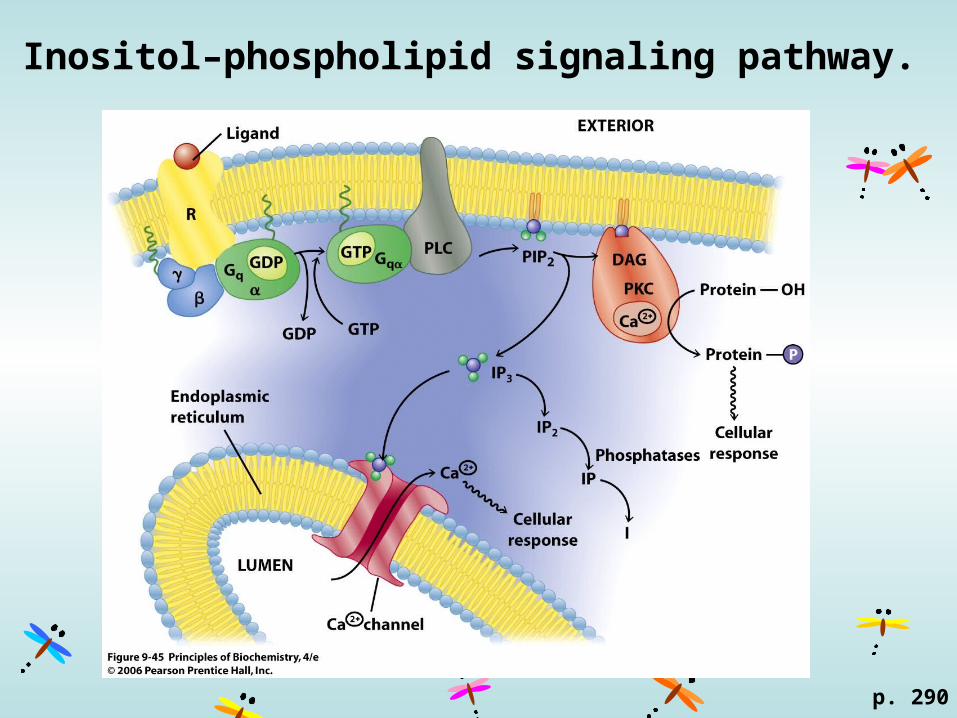
Inositol–phospholipid signaling pathway.
p. 290





























