Embed Size (px)
Citation preview

Hum Genet (1989) 81 : 188-190
© Springer-Verlag 1989
Presymptomatic testing for Huntington's disease
A case complicated by recombination within the D4S10 locus
Ann Curtis, F. Millan, Susan Holioway, Moira Mennie, Aileen Crosbie, J .A. Raeburn, and D.J .H. Brock
Human Genetics Unit, University of Edinburgh, Western General Hospital, Edinburgh, EH4 2XU, UK
Summary. Presymptomatic testing for Huntington's disease (HD) is possible through the use of restriction fragment length polymorphisms (RFLPs) at the closely linked D4S10 locus. Recombination between the HD and D4S10 loci will occur in 4%-5% of meioses, and is a well-recognised complication of predictive testing. Recombination between RFLPs within the D4S10 locus is a rare event and can usually be ignored. We re- port a case where such an intra-locus recombination frustrated attempts to predict the chance of a high-risk individual inherit- ing the HD gene.
Introduction
Huntington's disease (HD) is an autosomal dominantly inher- ited genetic defect characterised by progressive chorea, psy- chological disturbance and ultimately dementia (Martin and Gusella 1986). The symptoms of the disease do not usually ap- pear until the third or fourth decade, and progression to death occurs within 15-20 years (Harper 1986). The penetrance of the gene is assumed to be 100%, so that unless they die of other causes, all carriers of the HD gene eventually manifest the disease (Conneally 1984). Individuals who have an affect- ed parent have a 50% risk of inheriting the HD gene and must live for many years with this uncertainty as their major preoc- cupation.
The discovery of DNA markers tightly linked to the HD locus (Gusella et al. 1983) has now made it possible to track the HD gene in suitable families and to offer presymptomatic testing to those who are at high risk. Recent surveys suggest that as many as two-thirds of all those with an affected parent may request such testing (Kessler et al. 1987; Mastromauro et al. 1987; Meissen and Berchek 1987; Markel et al. 1987). Since the inception of our predictive testing programme a year ago, 20 individuals have enrolled in the extensive psychiatric and genetic counselling processes, which necessarily precede labo- ratory analyses. Details of this programme will be reported when post-testing counselling and evaluation is completed.
One of the limitations of the use of D N A markers in pre- symptomatic testing is the possibility of recombination be- tween the D4S10 locus and the HD locus. This is estimated to be between 4% and 5% (Quarre l le t al. 1987; Meissen et al. 1988). A less well-recognised drawback is recombination with- in the D4S10 locus, now characterised by a number of different restriction fragment length polymorphisms (RFLPs) (Gusella
Offprint requests to: D. J. H. Brock
et al. 1983, 1985; Harper 1986; Hayden et al. 1987). In the first family that we analysed for predictive testing, a rare re- combinant of this kind was encountered.
Materials and methods
Family study
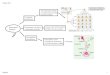
The consultand (Fig. 1, IV-l ) was a 28-year-old woman with an affected mother (III-2), maternal uncle (III-3) and mater- nal grandfather (II-1). Inspection of the extended pedigree in our Genetic Register showed two unaffected great-uncles, aged 70 and 68 (II-3, II-4), and an affected great-aunt (II-5). Blood and DNA samples were available from all the individu- als in generations II and III, including the deceased grand- father (II-1), but not from the two individuals in generation I. Since the family structure appeared suitable for presymptoma- tic testing, the consultand was enrolled in the programme and a preliminary work-up initiated.
DNA analysis
D N A was prepared from white blood cells by standard meth- ods. Samples of 5 gg genomic D N A were digested with the re- striction enzymes HindIII, EcoRI, PstI and BglI according to
ii / m
Hind n I sire 1 17.5/15.0 17.5/15.0 17.5/15.0 17.5/15.0 17.5/15.0 Hind In sire 2 3.7/ 3.7 3.7/ 3.7 5.7/ 3.7 3.7/ 3.7 3.7/ 3.7 EcoRI 14.5/145 14.5/1/+5 14.5/ 9.1 145/ 9.1 1L,.5/1~-.5 PstI 5.5/ 2,4 5.5/ 2.4 5.5/ 2,4 5.5/ 2,4 5.5/ 2,~- Bgll 2.0/ 2,0 2.0/ 2,0 2.0/ 2,0 2.0/ 2.0 2.0/ 2.0
,, .L2 .I.3 17.5 / 17.5 17.5 / 15.0 15.0 / 15.0 3.7 / 3.7 33 / 3.7 3.7 / 3.7
14.51 9.1 1&5 / 1~-.5 14.5/14.5 5.5/ 5.5 2.4/ 2,4 2,41 2.4 2,0/ 1.9 1 2.0/ 2.0 2.0/ 2.0
IV
17.5/15.0 3.7 / 3,7
14.5/ 9.1 5.5/ 2.4 2.0 / 1.9
Fig.1. Pedigree of the extended family and genotypes for the five re- striction fragment length polymorphisms

189
the maker's instructions. After electrophoresis on 0.8% agarose gels and transfer to nylon membranes (Genescreen), samples were hybridized with probes pR7 (HindIII, PstI, BglI) or pK083 (EcoRI).
Risk analysis
The probability of the consultand being a carrier of the HD gene was calculated using the MLINK option of the LINKAGE program (version 3.5). The recombination frequency between the HD and D4S10 loci was fixed at 5%, and between restric- tion sites at the D4S10 locus at (arbitrarily) 0.1%. A domi- nant model with equal recombination frequencies in males and females was assumed, and the HD gene frequency taken as 0.0001. The penetrance for the unaffected at-risk individu- als, 1I-3 and II-4, respective ages of 70 and 68, was assumed to be 0.94 (Newcombe 1981).
Results
Genotypes at the five polymorphic restriction sites for the ex- tended family are shown in Fig. 1. It is apparent that recombi- nation within the D4S10 locus had occurred, either in a pater- nal meiosis of II-1 or a maternal meiosis of II-2. The alterna- tive explanation of non-paternity is extremely unlikely, given that the HD gene has been transmitted from II-1 to both his children.
In attempting to determine the site of recombination the HindIII site 2, EcoRI and BglI polymorphisms can be ignored since they are not informative. The most probable orientation of the remaining restriction sites is HD-HindIII (site 1)-PstI (Gilliam et al. 1987a; Wexler et al. 1987; Skraastad et al. 1987). If this is the case recombination must have occurred in a maternal meiosis of II-2, with either III-2 or III-3 inheriting recombinant chromosomes.
Calculation of risks was made using the LINKAGE pro- gram. For the most likely gene order, HD-HindIII (site 1)- PstI, the risk of the consultand inheriting the HD gene was 78.7%. For the less likely order, HD-PstI-HindIII (site 1), the risk was 46.0%. The consultand was advised to await the in- troduction of the new generation of probes showing a lower recombination rate with the HD gene (Gilliam et al. 1987b; Wasmuth et al. 1988).
Discussion
The chance of recombination between different restriction sites in a short marker locus is small and can usually be ignored. This is the second published report of recombination within the D4S10 locus; Gilliam et al. (1987a) described a cross-over in maternal meiosis between the HindIII site i and the EcoR!
pK083 R7
HI BP H2 R p te r / I i i ~ i i Centrornere
HD
Fig. 2. Probable order of polymorphic restriction sites on chromosome 4. HI HindIII site 1, B BglI, P PstI, H2 HindIII site 2, R EcoRI
site. This report and others (Wexler et al. 1987; Skraastad et al. 1987) suggest the orientation of restriction sites shown in Fig. 2.
This case emphasises the importance of using multiple markers at the D4S10 locus in carrying out presymptomatic testing for HD. If only the HindIII polymorphism had been used the consultand would have been advised of a high risk of inheriting the HD gene. However, the use of the PstI poly- morphism cast doubt on this conclusion and made it sensible to avoid any diagnosis until more polymorphic and more close- ly linked probes were in routine use (Gilliam et al. 1987b).
As a consequence of this experience, we have revised our protocol for presymptomatic testing. It is now our practice to work up all available samples from the immediate family with a battery of probes before taking a blood sample from the con- sultand. The theoretical possible genotypes for the consultand are then constructed and the risk attaching to each are calcu- lated with the MLINK computer program. In this way it is possible to advise the consultand of the range of risks he or she is likely to encounter before entering on the final phase of presymptomatic testing. We have found that many individuals at 50% risk of inheriting the HD gene are disinclined to under- go testing unless their odds can be very substantially altered.
Acknowledgements. We are grateful to Dr. J. F. Gusella for supplying the probes used here. The work was supported by grants from the Scottish Hospital Endowments Research Trust and the Ludovici Trust of the University of Edinburgh.
References
Conneally PM (1984) Huntington disease: genetics and epidemiology. Am J Hum Genet 36 : 506-526
Gilliam TC, Tanzi RE, Haines JL, Bonner TI, Faryniarz AG, Hobbs WJ, McDonald ME, Cheng SV, Folstein SE, Conneally PM, Wexler NS, Gusella JF (1987a) Localization of the Huntington's disease gene to a small segment of chromosome 4 flanked by D4S10 and the telomere. Cell 50 : 565-571
Gilliam TC, Bucan M, McDonald ME, Zimmer M, Haines JL, Cheng SV, Pohl TM, Meyers RH, Whaley WL, Allitto BA, Faryniarz A, Wasmuth JJ, Frischauf A-M, Conneally PM, Lehrach H, Gusella JF (1987b) A DNA segment encoding two genes very tightly linked to Huntington's disease. Science 238 : 950-952
Gusella JF, Wexler NS, Conneally PM, Naylor SL, Anderson MA, Sakaguchi AY, Young AB, Shoulson I, Bonilla E, Martin JB (1983) A polymorphic DNA marker genetically linked to Hun- tington's disease. Nature 306 : 234-238
Gusella JF, Tanzi RE, Bader PI, Phelan MC, Stevenson R, Hayden M, Hofman KJ, Faryniarz AG, Gibbons K (1985) Deletion of Huntington's disease-linked G8 (D4S10) locus in Wolf-Hirschhorn syndrome. Nature 318 : 75-78
Harper PS (1986) The prevention of Huntington's chorea. J R Coll Physicians Lond 20 : 7-14
Hayden MR, Hewitt J, Kastelein JJP, Langlois S, Wilson RD, Fox S, Hilbert C, Bloch M (1987) First-trimester prenatal diagnosis for Huntington's disease with DNA probes. Lancet II: 1284-1285
Kessler S, Field T, Worth L, Mosbarger H (1987) Attitudes of per- sons at risk for Huntington disease toward predictive testing. Am J Med Genet 26 : 259-270
Markel DS, Young AB, Penney JB (1987) At-risk persons' attitudes toward presymptomatic and prenatal testing of Huntington dis- ease in Michigan. Am J Med Genet 26 : 295-305
Martin JB, Gusella JF (1986) Huntington's disease. Pathogenesis and management. N Engl J Med 315 : 1267-1276
Mastromauro C, Myers RH, Berkman B (1987) Attitudes toward pre- symptomatic testing in Huntington disease. Am J Med Genet 26 : 271-282

190
Meissen GJ, Berchek RL (1987) Intended use of predictive testing by those at risk for Huntington disease. Am J Med Genet 26:283- 293
Meissen GJ, Myers RH, Mastromauro CA, Koroshetz WJ, Klinger KW, Farrer LA, Watkins PA, Gusella JF, Bird ED, Martin JB (1988) Predictive testing for Huntington's disease with use of a linked DNA marker. N Engl J Med 318:535-542
Newcombe RG (1981) A life table for onset of Huntington's chorea. Ann Hum Genet 45: 375-385
Quarrell OWJ, Meredith AL, Tyler A, Youngman S, Upadhyaya M, Harper PS (1987) Exclusion testing for Huntington's disease in pregnancy with a closely linked DNA marker. Lancet II:1281- 1283
Skraastad MI, Lange LF de, Bakker E, Van Ommen GJB, Pearson PL (1987) Extension of the Huntington's disease marker D4S10 by cosmid walking. Cytogenet Cell Genet 46 : 693-694
Wasmuth JJ, Hewitt J, Smith B, Allard D, Haines JL, Skarecky D, Partlow E, Hayden MR (1988) A highly polymorphic locus very tightly linked to the Huntington's disease gene. Nature 332:734- 736
Wexler NS, Young AB, Tanzi RE, Travers H, Starosta-Rubinstein S, Penney JB, Snodgrass SR, Shoulson I, Gomez F, Ramos Arroyo MA, Penchaszadeh GK, Moreno H, Gibbons K, 'Faryniarz A, Hobbs W, Anderson MA, Bonilla E, Conneally MC, Gusella JF (1987) Homozygotes for Huntington's disease. Nature 326:194- 197
Received May 7, 1988 / Revised August 19, 1988