Embed Size (px)
Citation preview

RESEARCH ARTICLE
Presence of bacteria in aqueous solution influencesvirus adsorption on nanoparticles
Bingzi Zhao & Jiabao Zhang & Yan Jiang
Received: 21 March 2013 /Accepted: 2 May 2013# Springer-Verlag Berlin Heidelberg 2013
Abstract Virus contamination in wastewater is usually ac-companied by the existence of various bacteria. Nanoparticles(NPs) have been shown to efficiently remove virus. In thisstudy, bacterial cells, supernatants, and cultures wereharvested separately from three strains at the culture ages of6 and 24 h, corresponding to the log and stationary phases,respectively. The aim is to investigate how their presenceaffects virus adsorption on the three Fe and Al oxide NPs(α-Fe2O3, γ-Fe2O3-B, and Al2O3) and how these effectschange with bacterial growth phase. Bacteriophage phiX174was used as a virus model. Results showed that bacterial cells,supernatants, and cultures harvested at 6 h generally reducedvirus adsorption by an average of 0.75±0.84, 7.7±9.0, and10.3±8.6 %, respectively, while those harvested at 24 h re-duced virus adsorption by an average of 2.1±0.93, 21.5±6.6,and 24.6±6.9 %, respectively. Among the NPs, α-Fe2O3
showed more sensitivity to bacteria than the other two, prob-ably because of its relatively higher value of point of zerocharge. It was found that cell-induced and supernatant-induced reductions were combined to achieve added results,in which the supernatants contributed much more than thecells, implying that the bacterial exudates might be morecrucial in the reduced virus adsorption than the bacterial cells.These results strongly demonstrated that the bacteria-inducedreduction in virus adsorption became more significant withculture age. It is suggested that studies conducted in theabsence of bacteria may not accurately evaluate the potentialof virus removal efficiency of the NPs in bacteria-containingenvironments.
Keywords Virus adsorption . Bacterial cultures .
Supernatants . Cells . Growth phase
Introduction
Virus can be removed from aqueous solution through ad-sorption and/or inactivation (Zhang et al. 2010).Understanding the factors influencing virus adsorption is,therefore, important for evaluating virus removal efficiencyby an adsorbent. Natural aquatic systems consist of complexassemblages of organic macromolecules (natural dissolvedorganic matter [DOM] and bacterial exudates), inorganiccolloids (clay minerals and oxides), and microbial cells(Omoike et al. 2004). In addition, wastewaters such asmunicipal wastewaters and animal waters typically containbacteria, viruses, intestinal parasites, and a complex mixtureof partially digested organic matter simultaneously(Bradford et al. 2006; Koivunen and Heinonen-Tanski2005). This indicates that viruses usually exist simulta-neously with organic macromolecules, inorganic colloids,and microbial cells in a wastewater, and thus, their effectsshould be investigated when removing viruses from thesecomplex systems.
How DOM, inorganic colloids, as well as solution ioniccomposition and ionic strength influence virus adsorptionand virus fate has received significant attention (e.g., Chu etal. 2000; Jin et al. 2000; Kim et al. 2003; Zhuang and Jin2003; Gutierrez et al. 2009; Shen et al. 2010), which usuallyleads to the conclusion that these environmental factors mayeither facilitate and reduce or have no effect on virus ad-sorption. Additionally, the few previous investigations re-garding the significance of bacteria on virus adsorption, e.g.,Kim et al. (2003) showed that contaminant is enhancedmarkedly in the presence of DOM and bacteria using nu-merical simulations but without any support from any ex-perimental results, whereas Lipson and Stotzky (1985) only
Responsible editor: Robert Duran
B. Zhao (*) : J. Zhang :Y. JiangState Key Laboratory of Soil and Sustainable Agriculture, Instituteof Soil Science, Chinese Academy of Sciences, P.O. Box 821, 71East Beijing Road,Nanjing 210008, People’s Republic of Chinae-mail: [email protected]
Environ Sci Pollut ResDOI 10.1007/s11356-013-1802-y

concerned the effect of culture supernatants that containedbacterial exudates from one growth phase and Zhang et al.(2010) demonstrated, by comparing virus adsorption onunsterilized soils with that on sterilized soils, that sterilization-induced virus adsorption was virus type dependent. Thus, thesignificance of the effect of bacteria on virus adsorption as wellas the contribution of cells and exudates has never been testedby direct comparison of virus adsorption onto an adsorbent inthe presence of cells with those in the presence of exudates fromthe same strain and same growth phase. It is noteworthy thatbacteria are living organisms that show metabolic and physio-logical changes (Walker et al. 2005) and the amount of cells andexudates change with growth phases, which consist of lag, log,stationary, and death phases.
Nanoparticles (NPs) have a typical size of 5–100 nm(Link et al. 2007), which may result in high surface area.Inorganic metal oxide NPs have been shown to be able toremove various viruses from aqueous solutions, with thebinding capacity of Al2O3 NPs reaching more than0.26 μg DNA (6×1010 molecules of DNA)/μg NPs (Linket al. 2007). Gutierrez et al. (2009) reported a high removalof bacteriophage MS2 and rotavirus at a low concentrationof hematite NPs in solution. Our recent study showed thatNPs of α-Fe2O3 and γ-Fe2O3-N adsorbed more than 99 %of phiX174, in which α-Fe2O3 was shown to be moreefficient, with an adsorption percentage of phiX174reaching at about 100 % when the initial virus concentrationwas around 1×103 plaque-forming units (PFU)ml−1 (Shenet al. 2010). The surface charge on Fe2O3 in aqueous solu-tions results from the dissociation of surface hydroxylgroups that efficiently remove viruses from water by elec-trostatic interactions (Ryan et al. 2002). A number of abio-logical factors have been proposed to govern virusadsorption on NPs, such as DOM (Gutierrez et al. 2009),bivalent and monovalent cations (Gutierrez et al. 2009; Shenet al. 2010; Link et al. 2007), anions (Gutierrez et al. 2009),NP material (Liga et al. 2011; Nangmenyi et al. 2011; Hu etal. 2012), and NP content (Liga et al. 2011). However, fewinvestigations have studied the effect of bacteria.
The present work aims at determining how the presenceof two Gram-negative bacteria (Pseudomonas putida andPseudomonas aeruginosa) and one Gram-positive bacteri-um (Bacillus subtilis) of environmental interests affectsvirus adsorption on three Fe and Al oxide NPs and assessingthe relative contribution of different bacterial components(i.e., intact cells and bacterial exudates in the supernatants)harvested at culture ages of 6 and 24 h. The specific objectivesare to (1) investigate virus adsorption on NPs as influenced bythe cells, supernatants, and cultures and (2) evaluate whetherthe response of virus adsorption to the presence of bacteria isaltered by the growth phase. Possible mechanisms of virusadsorption on NPs as influenced by bacteria are discussed. Toour knowledge, these results have not been reported.
Materials and methods
Bacterial strains, culture conditions, and growth curves
Bacterial species used in the present study consisted of bothGram-negative and Gram-positive bacteria and wereobtained from the Chinese Center for Type CultureCollection. They were selected because they are ubiquitous-ly distributed in water and soil. They are (1) P. putida: aGram-negative bacterium; cultivated at 30 °C in LB medi-um [1.0 g tryptone (LP0042, Oxoid Ltd., England), 0.5 gyeast extract (LP0021, Oxoid Ltd., England), and 1.0 gNaCl in 100 ml], which was used for liquid cultivation,while for solid medium cultivation, 13.5 g agar L−1 wasadded; (2) P. aeruginosa: a Gram-negative bacterium; cul-tivated at 35 °C in LB medium; and (3) B. subtilis: a gram-positive bacterium; cultivated at 35 °C in PYG medium[3.0 g glucose, 1.0 g tryptone, 0.5 g yeast extract, 0.45 gKH2PO4, and 0.2 g CaCO3 in 100 ml], which was used forliquid cultivation, while for solid medium cultivation, 13.5 gagar L−1 was added. The number of colony-forming units(CFU) was used to quantify bacteria by heterotrophic platecount.
Flasks were inoculated from 24-h-old bacteria subculturedfrom a single colony for measurement of bacterial growthcurves. These cultures were allowed to grow at 30 °C (P.putida) or 35 °C (P. aeruginosa and B. subtilis) and shakenat 150 rpm for 48 h. At the requisite intervals, 1-ml sampleswere withdrawn for optical density (OD) measurement at600 nm (P. putida) or 585 nm (P. aeruginosa and B. subtilis)using a 722 visible spectrophotometer (JingHua Instruments,Shanghai, China) against the growth medium as a control tofollow bacterial cell multiplication and identify the growthphases. The number of viable cells was determined and wasexpressed as log10 CFU per milliliter. Experiments were rep-licated three times. Duplicate measurements were taken foreach replicate experiment.
Nanoparticles
NPs of α-Fe2O3, γ-Fe2O3-B, and Al2O3 were used in thisstudy. They were purchased from Beijing Nachen TechnologyCo., Ltd. (Beijing, China) with purity >99%. Besides the highefficiency of virus adsorption of NPs of Fe and Al oxides(Shen et al. 2010), another reason for selecting them was thatthey present low risks for adding a further pollutant such asdisinfection by-products via chlorination to the system. Beforeuse, NPs were extracted with sterilized physiological saline(SPS) and quantified for bacteria, fungi, and actinomycetes.We found that almost no microorganisms were detected. Afterthat, they were examined by (1) transmission electron micros-copy (JEM-200CX, JEOL, Japan) and X-ray diffraction(D/MAX 2000PC, Rigaku Corporation, Japan) for particle
Environ Sci Pollut Res

size estimation; (2) nitrogen sorption for specific surface area,total pore volume, and mesopore size (BET method using anASAP 2010 surface area analyzer (Micromeritics, Norcross,GA, USA)); and (3) potentiometric titration method (Van Raijand Peech 1972) for point of zero charge. Their results arelisted in Table 1.
Virus and plaque assay
Bacteriophage phiX174 was chosen as a model virus for allexperiments in this study because it has been used as asurrogate in many studies and because of its structure resem-blance to many human enteric viruses (Schijven andHassanizadeh 2000; Jin and Flury 2002). phiX174 wasobtained from the American Type Culture Collection (ATCC13706B1). It is an icosahedral single-stranded DNA phagewith a diameter of about 23.0 nm (Yates et al. 1990) and has anisoelectric point of 6.6 (Dowd et al. 1998). BEP medium [5 gNaCl, 3 g ‘Lab-Lemco’ powder (LP0029, Oxoid Ltd.,England) and 5 g tryptone in 1,000 ml] was used for liquidcultivation, and 13.5 or 4 g agar L−1 was added for solidmedium or soft medium cultivation, respectively. phiX174was grown on bacterial lawns of host Escherichia coli(ATCC 13706) on BEP medium agar and then harvested.
Propagation of phiX174 and preparation of its stock solu-tion have been described in our previous studies (Shen et al.2010; Zhang et al. 2010). Typical stock concentrations rangedbetween 108 and 109 PFU ml−1. phiX174 was assayed by thedouble-layer overlay method (Adams 1959). As for the de-tails, refer to Zhang et al. (2010) and Shen et al. (2010). Viablevirus concentration was determined by counting the numberof plaques in the host lawn and reported as PFU per milliliter.Only dilutions that resulted in 10–300 plaques per plate wereaccepted for quantification. Each reported virus concentrationis the average of three replicate plates.
Experimental design
Preparation of bacterial cultures, culture supernatants,and bacterial cell suspensions
To investigate the influence of each bacterium and its compo-nents (i.e., cells and exudates) at different growth phases onvirus adsorption on NPs, the fresh cultures were harvested at
culture ages of 6 and 24 h, using the same cultivation conditionsas described in the “Bacterial strains, culture conditions, andgrowth curves” section. The cultures were divided into twofractions at the time of harvest. The first fraction was directlykept at 4 °C until the start of the virus adsorption experiment.The second fraction was transferred to a sterilized 50-ml poly-propylene centrifuge tube, followed by centrifugation for10 min at 12,096×g at 4 °C (Avanti J-30I, Beckman Coulter,USA). The cell-free culture supernatants were collected andstored at 4 °C until needed. Bacterial cells in the precipitates inthe tube were washed in SPS for two to three times andresuspended to its original volume using SPS, which formedbacterial cell suspension and was kept at 4 °C until needed.
Virus adsorption on the NPs as influenced by the bacterialcells
The equilibrium adsorption experiments were conducted at4 °C to avoid thermal inactivation of the virus. One milliliterof virus solution with concentration of about 107 PFUml−1 and1.0 ml of 6 or 24 h cell suspension was added to a sterilized 15-ml polypropylene centrifuge tube containing 0.0800 g of α-Fe2O3 and γ-Fe2O3-B or 0.0400 g of Al2O3. The used virussolutions were diluted from the same stock solution with SPS.The virus–NP–cell suspension was immediately sealed andmixed for 3 h at 4 °C, which was continuously shaken at300 rpm. The equilibration time of 3 h was selected based onthe results of our previous study that showed 3 h to be sufficientto reach the equilibrium of virus adsorption (Zhao et al. 2008).The suspension was then centrifuged for 20 min at 12,096×g(Avanti J-30I, Beckman Coulter, USA) and viable virus con-centration in the supernatant was assayed. The controlsconsisted of 1.0 ml of the virus solution plus NPs plus 1.0 mlof the SPS, and the blanks consisted of 1.0 ml of the virussolution plus 1.0 ml of the SPS. Both control and blank tubeswere treated in the same manner as the experimental tubes.There were three replicates for each treatment. Virus adsorptionpercentage was calculated with the following formula:
R ¼ ½ Ci–Cð Þ � 100=Ci�where Ci,C, and R are, respectively, the concentrations of virusin the blank liquid phase (in PFU per milliliter), the concentra-tions of virus in the experimental liquid phase or in the control
Table 1 Some properties of the NPs used in this study
Average particle size(nm)
BET specific surface area(m2 g−1)
Pore volume(m3 g−1)
Average pore size(nm)
Point of zerocharge
α-Fe2O3 35 51.9 0.24 18.2 8.36
γ-Fe2O3-B
10 107.0 0.30 11.0 6.73
Al2O3 15 87.5 0.15 7.0 6.55
Environ Sci Pollut Res
treatment (in PFU per milliliter), and the virus adsorptionpercentage (in percent).
At the same time, viable bacteria were assayed at end ofthe batch experiment. For comparison, viable bacteria in thetubes without viruses and NPs were also assayed.
Virus adsorption on the NPs as influenced by the culturesupernatants
The experimental procedure in this section was quite similarto that described in the “Virus adsorption on the NPs asinfluenced by the bacterial cells” section, except that the cellsuspension was replaced by the culture supernatants.Consequently, the controls consisted of 1.0 ml of the virussolution plus NPs plus 1.0 ml of the liquid growth mediumof LB (P. putida and P. aeruginosa) or PYG (B. subtilis),and the blanks consisted of 1.0 ml of the virus solution plus1.0 ml of the liquid growth medium.
Virus adsorption on the NPs as influenced by the bacterialcultures
The experimental procedure in this section was quite similarto that described in the “Virus adsorption on the NPs asinfluenced by the culture supernatants” section, except thatthe culture supernatants were replaced by the bacterial cul-tures. Consequently, the controls and blanks were complete-ly the same as that indicated in the “Virus adsorption on theNPs as influenced by the culture supernatants” section.Additionally, viable bacteria were assayed at the end of thebatch experiment. For comparison, viable bacteria in thetubes without viruses and NPs were also assayed.
Statistical analysis
Analysis of variance (ANOVA) was used to examine theeffects of bacterial cells, culture supernatants, and cultures ofeach bacterial species harvested at different growth phases onvirus adsorption percentage. ANOVAwas also used to exam-ine the changes in the number of viable bacteria before andafter the batch experiments. Significant differences of meanswere judged by Duncan multiple comparison tests, with thechosen statistical difference level of 5 %. All statistical anal-yses were performed with SPPS version 17.0.
Results
Bacterial growth curves
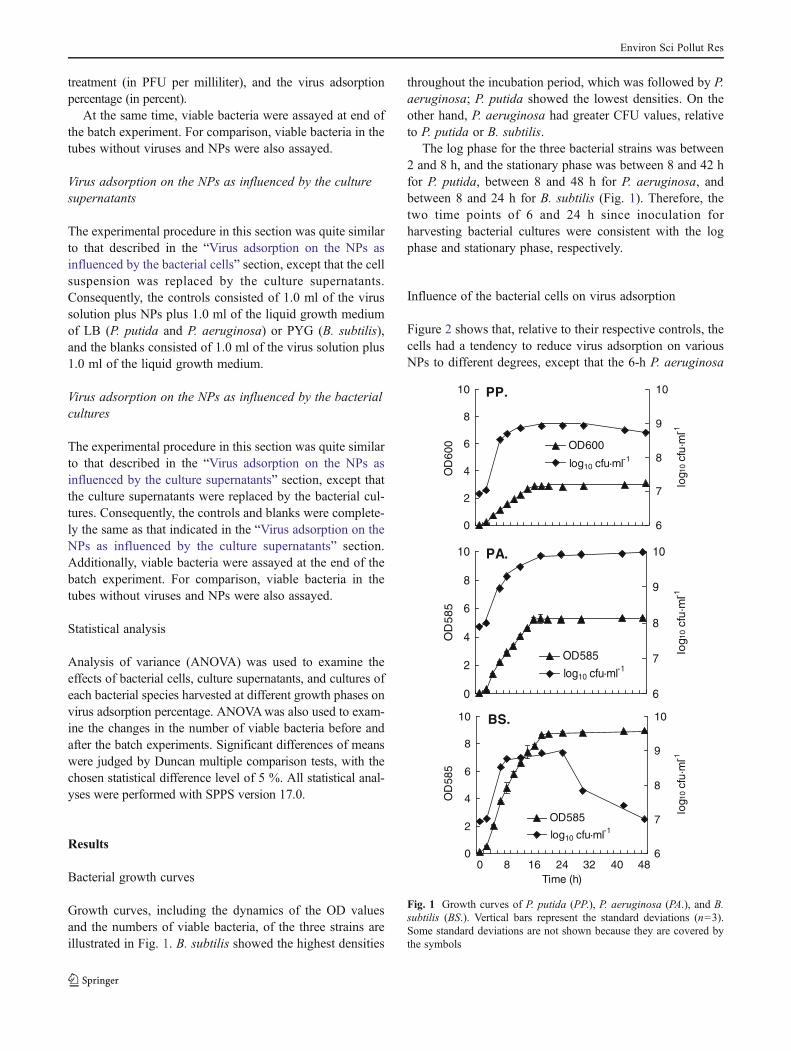
Growth curves, including the dynamics of the OD valuesand the numbers of viable bacteria, of the three strains areillustrated in Fig. 1. B. subtilis showed the highest densities
throughout the incubation period, which was followed by P.aeruginosa; P. putida showed the lowest densities. On theother hand, P. aeruginosa had greater CFU values, relativeto P. putida or B. subtilis.
The log phase for the three bacterial strains was between2 and 8 h, and the stationary phase was between 8 and 42 hfor P. putida, between 8 and 48 h for P. aeruginosa, andbetween 8 and 24 h for B. subtilis (Fig. 1). Therefore, thetwo time points of 6 and 24 h since inoculation forharvesting bacterial cultures were consistent with the logphase and stationary phase, respectively.
Influence of the bacterial cells on virus adsorption
Figure 2 shows that, relative to their respective controls, thecells had a tendency to reduce virus adsorption on variousNPs to different degrees, except that the 6-h P. aeruginosa
PP.
0
2
4
6
8
10
OD
600
6
7
8
9
10
log1
0 cf
u·m
l-1
OD600
log10 cfu·ml-1
PA.
0
2
4
6
8
10
OD
58
5
6
7
8
9
10
log1
0 cf
u·m
l-1
OD585
log10 cfu·ml-1
BS.
0
2
4
6
8
10
0 8 16 24 32 40 48Time (h)
OD
585
6
7
8
9
10
log1
0 cf
u·m
l-1
OD585
log10 cfu·ml-1
Fig. 1 Growth curves of P. putida (PP.), P. aeruginosa (PA.), and B.subtilis (BS.). Vertical bars represent the standard deviations (n=3).Some standard deviations are not shown because they are covered bythe symbols
Environ Sci Pollut Res

cells significantly promoted virus adsorption on both γ-Fe2O3-B and Al2O3. The 24-h cells consistently significantlyinhibited virus adsorption on all the studied NPs, irrespectiveof the cell species, and showed more significant inhibitiveeffect than the 6-h cells, with the average virus adsorptionbeing reduced by 2.1±0.93% in the presence of 24-h cells andby 0.75±0.84 % in the presence of 6-h cells (Fig. 2).
Following the 3-h batch adsorption, the concentration ofviable cells was usually reduced in the virus–NP–cell sys-tems, relative to their respective controls, though not allreduction was significant (Table 2). Culturable cells in thevirus–Al2O3–6 h P. putida cell and virus–Al2O3–6 h B.subtilis cell systems, for example, did not significantly differfrom those in their controls (Table 2).
90
92
94
96
98
100
Viru
s ad
sorp
tion
perc
enta
geR
(%)
Control PP.a
b
a
bb
a
2O3 2O3-B Al2O3
24 h Control PA.
ab
abb
a
2O3 2O3-B Al2O3
24 h Control BS.
b
aba b
a
2O3 2O3-B Al2O3
24 h
a
-Fe -Fe -Fe -Fe -Fe
-Fe
90
92
94
96
98
100
Viru
s ad
sorp
tion
perc
enta
geR
(%)
Control PP.a
ba b
ab
2O3 2O3-B Al2O3
6 h
-Fe -Fe
Control PA. a b b a b a
2O3 2O3-B Al2O3
6 h
-Fe -Fe
Control BS.a
b
ab
ab
2O3 2O3-B Al2O3
6 h
-Fe
-Fe
Fig. 2 The presence of bacterial cells affects virus adsorption on NPs.PP., PA., and BS. stand for P. putida, P. aeruginosa, and B. subtilis,respectively. Vertical bars represent the standard deviations (n=3).
Different letters within each NP indicate significant differences basedon Duncan0.05
Table 2 Concentration of culturable bacterial cells following the 3-h batch experiment in the virus–NP–cell system
Inputconcentration(CFU ml−1)
Concentration in blankwithout viruses and NPs(CFU ml−1)
Concentration invirus–α-Fe2O3–cellsystem (CFU ml−1)
Concentration in virus–γ-Fe2O3-B–cell system(CFU ml−1)
Concentration in virus–Al2O3–cell system(CFU ml−1)
6 h
P. putida 3.13E+08 (3.04±0.18)E+08 a (1.68±0.09)E+08 c (2.47±0.12)E+08 b (3.02±0.16)E+08 a
P.aerugin-osa
9.46E+08 (9.53±0.67)E+08 a (5.64±0.15)E+08 c (4.38±0.20)E+08 d (7.55±0.80)E+08 b
B. subtilis 3.33E+08 (3.26±0.20)E+08 a (9.64±0.64)E+07 b (9.56±0.76)E+07 b (3.18±0.30)E+08 a
24 h
P. putida 8.05E+08 (7.98±0.42)E+08 a (4.93±0.17)E+08 c (5.72±0.36)E+08 b (5.16±0.24)E+08 bc
P.aerugin-osa
8.20E+09 (8.13±0.55)E+09 a (3.85±0.14)E+09 c (4.09±0.23)E+09 c (7.08±0.60)E+09 b
B. subtilis 8.38E+08 (8.30±0.72)E+08 a (5.44±0.26)E+08 b (5.57±0.24)E+08 b (7.30±0.61)E+08 b
The concentrations are presented as the means±standard deviations (n=3). Different letters within a row indicate significant difference based onDuncan 0.05
Environ Sci Pollut Res

Influence of the culture supernatants on virus adsorption
Similar to the bacterial cells, the culture supernatants signif-icantly reduced virus adsorption on the NPs in most cases,except that the 6-h B. subtilis supernatants had no significanteffect on the virus adsorption on γ-Fe2O3-B and Al2O3
(Fig. 3). The 24-h supernatants consistently and significantlyinhibited virus adsorption onto all three NPs and generallyshowed more significant inhibitive effect than the 6-h super-natants, with the average virus adsorption being reduced by21.5±6.6 % in the presence of the 24-h supernatants and by7.7±9.0 % in the presence of the 6-h supernatants.
Influence of the bacterial cultures on virus adsorption
As with the effects of the culture supernatants, the bacterialcultures significantly reduced virus adsorption on the NPs,except that the 6-h B. subtilis cultures had no significanteffect on virus adsorption on γ-Fe2O3-B and Al2O3 (Fig. 4).The 24-h cultures significantly inhibited virus adsorption onall three NPs and showed more significant inhibitive effectthan the 6-h cultures (Fig. 4), with the average virus adsorp-tion being reduced by 24.6±6.9 % in the presence of the 24-h cultures and by 10.3±8.6 % in the presence of the 6-hcultures.
At the end of the 3-h batch experiment, viable cells weresignificantly reduced in the systems of virus–α-Fe2O3–cultureand virus–γ-Fe2O3-B–culture, regardless of the cell speciesand growth phases, relative to their respective controls(Table 3). However, in the virus–Al2O3–culture system, the6-h P. putida and B. subtilis and 24-h P. aeruginosa and B.subtilis cultures did not lead to any significant change in theculturable cells at the end of the batch experiments (Table 3).
Discussion
The main aim of this study was to investigate whether virusadsorption on NPs was altered by the presence of bacteria inan aqueous solution. Our main finding is that bacterial cells,supernatants, and cultures had a tendency to inhibit virusadsorption on the three studied NPs. However, this inhibitiveeffect was greater in the presence of the 24-h bacteria than the6-h bacteria, as Figs. 2, 3, and 4 demonstrated that the 24-hbacteria led to consistent reduction in virus adsorption in allthe cases, while the effects of the 6-h bacteria varied with thetypes of NPs and bacterial species (Figs. 2, 3, and 4). Amongthe three NPs used in the present study,α-Fe2O3 was probablythe most susceptible to the presence of bacteria in terms of theinhibition of virus adsorption, as revealed by the consistently
0
15
30
45
60
Viru
s ad
sorp
tion
perc
enta
ge R
(%)
Control PP.24 h
2O3 2O3-B Al2O3
a
b
a
b
a
Control PA.24 h
2O3 2O3-B Al2O3
a
b
a a
b
Control BS.
Al2O32O3-B2O3
a
b
a
b
a
b
24 h
b
b
Control PA.
-Fe2O3 2O3-B Al2O3
6 h
a
b a
b
a
b
0
15
30
45
60
Viru
s ad
sorp
tion
perc
enta
geR
(%)
Control PP.
2O3 2O3-B Al2O3
6 h
a ba
b
a b
Control BS.
2O3 2O3-B Al2O3
a
ba
a a a
6 h
b
-Fe
-Fe-Fe
-Fe
-Fe-Fe
-Fe
-Fe
-Fe-Fe
-Fe
Fig. 3 The presence of bacterial supernatants affects virus adsorptionon NPs. PP., PA., and BS. stand for P. putida, P. aeruginosa, and B.subtilis, respectively. Vertical bars represent the standard deviations
(n=3). Different letters within each NP indicate significant differencesbased on Duncan0.05
Environ Sci Pollut Res
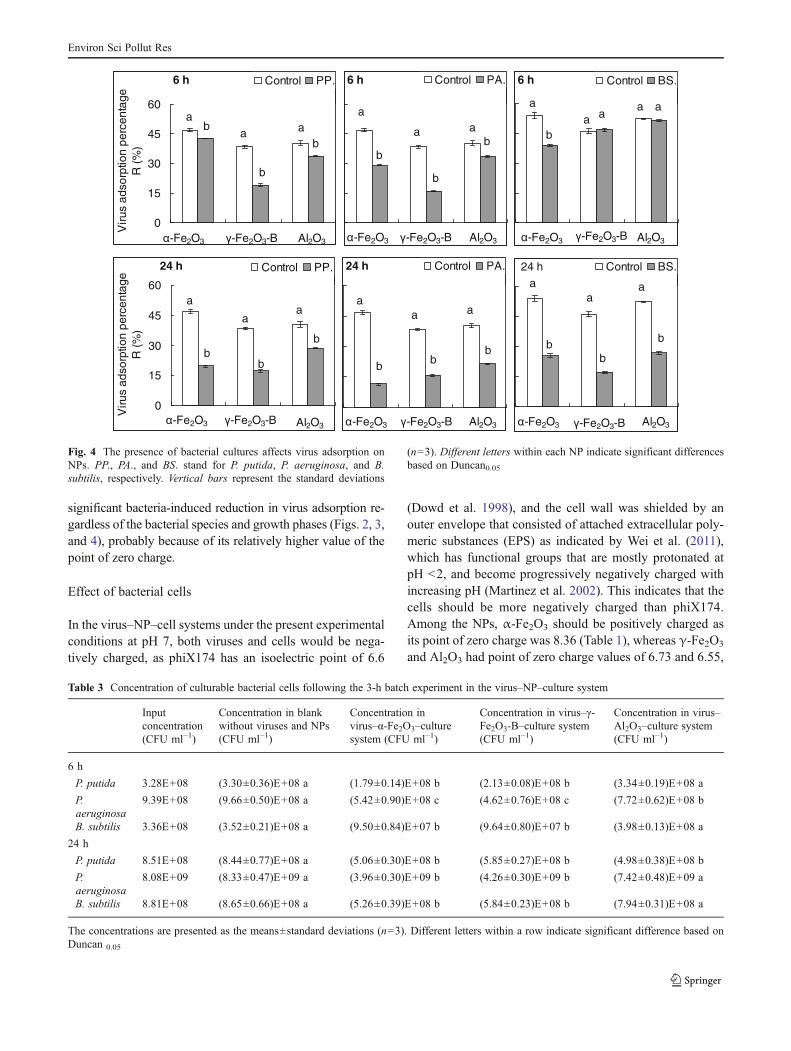
significant bacteria-induced reduction in virus adsorption re-gardless of the bacterial species and growth phases (Figs. 2, 3,and 4), probably because of its relatively higher value of thepoint of zero charge.
Effect of bacterial cells
In the virus–NP–cell systems under the present experimentalconditions at pH 7, both viruses and cells would be nega-tively charged, as phiX174 has an isoelectric point of 6.6
(Dowd et al. 1998), and the cell wall was shielded by anouter envelope that consisted of attached extracellular poly-meric substances (EPS) as indicated by Wei et al. (2011),which has functional groups that are mostly protonated atpH <2, and become progressively negatively charged withincreasing pH (Martinez et al. 2002). This indicates that thecells should be more negatively charged than phiX174.Among the NPs, α-Fe2O3 should be positively charged asits point of zero charge was 8.36 (Table 1), whereas γ-Fe2O3
and Al2O3 had point of zero charge values of 6.73 and 6.55,
Table 3 Concentration of culturable bacterial cells following the 3-h batch experiment in the virus–NP–culture system
Inputconcentration(CFU ml−1)
Concentration in blankwithout viruses and NPs(CFU ml−1)
Concentration invirus–α-Fe2O3–culturesystem (CFU ml−1)
Concentration in virus–γ-Fe2O3-B–culture system(CFU ml−1)
Concentration in virus–Al2O3–culture system(CFU ml−1)
6 h
P. putida 3.28E+08 (3.30±0.36)E+08 a (1.79±0.14)E+08 b (2.13±0.08)E+08 b (3.34±0.19)E+08 a
P.aeruginosa
9.39E+08 (9.66±0.50)E+08 a (5.42±0.90)E+08 c (4.62±0.76)E+08 c (7.72±0.62)E+08 b
B. subtilis 3.36E+08 (3.52±0.21)E+08 a (9.50±0.84)E+07 b (9.64±0.80)E+07 b (3.98±0.13)E+08 a
24 h
P. putida 8.51E+08 (8.44±0.77)E+08 a (5.06±0.30)E+08 b (5.85±0.27)E+08 b (4.98±0.38)E+08 b
P.aeruginosa
8.08E+09 (8.33±0.47)E+09 a (3.96±0.30)E+09 b (4.26±0.30)E+09 b (7.42±0.48)E+09 a
B. subtilis 8.81E+08 (8.65±0.66)E+08 a (5.26±0.39)E+08 b (5.84±0.23)E+08 b (7.94±0.31)E+08 a
The concentrations are presented as the means±standard deviations (n=3). Different letters within a row indicate significant difference based onDuncan 0.05
0
15
30
45
60
Viru
s ad
sorp
tion
perc
enta
geR
(%)
Control PP.
Al2O32O3-B2O3
24 h
a
b
a
b
a
b
Control PA.
Al2O32O3-B2O3
24 h
a
b
a
b
a
b
Control BS.
2O3 2O3-B Al2O3
a
b
a
b
a
b
24 h
-Fe0
15
30
45
60V
irus
adso
rptio
n pe
rcen
tage
R (%
)
Control PP.6 h
Al2O32O3 2O3-B
ab
a
b
ab
Control PA.6 h
Al2O32O3 2O3-B
a
b
a b
a
b
Control BS.
2O3-B Al2O32O3
a
ba a
a a
6 h
-Fe -Fe
-Fe -Fe
-Fe
-Fe
-Fe -Fe
-Fe -Fe
-Fe
Fig. 4 The presence of bacterial cultures affects virus adsorption onNPs. PP., PA., and BS. stand for P. putida, P. aeruginosa, and B.subtilis, respectively. Vertical bars represent the standard deviations
(n=3). Different letters within each NP indicate significant differencesbased on Duncan0.05
Environ Sci Pollut Res

respectively (Table 1), with their values approximating thevalue of the isoelectric point owned by phiX174. Virus andcell sorption on clay minerals and iron oxides is largelygoverned by electrostatic interactions, and van der Waalsforces also contribute to virus sorption (Jin and Flury 2002;Cao et al. 2011). This suggested that the cells might greatlyfavor the electrostatic interactions between cells and NPsand, thus, reduce the chance for viruses interacting withNPs, in which α-Fe2O3 should be more pronounced, mainlybecause of its high point of zero charge value.
In addition, it is highly likely that some fractions of NPswould be adsorbed by bacterial cells or may deposit on thecell walls, which results in changes in bacterial cell proper-ties (Adams et al. 2006; Zhang et al. 2007). Damage of cellwalls by NPs has not been investigated directly on thepresently used cells or NPs. Nonetheless, the contact of E.coli cells and ZnO NPs has led to extensive damage of cellmembrane in E. coli cells and cause the increase of E. colimembrane permeability, leading to the accumulation of ZnONPs in the cell membrane and NP internalization (Brayner etal. 2006). Brayner et al. (2006) further speculated thatleakage of the intracellular content, following passing ofNPs through the membrane, was possible. Moreover, formetallic oxide NPs, the generation of reactive oxygen spe-cies may be responsible for cell death (Su et al. 2009). Thishas been partly confirmed by the general reduction in theculturable cells following the 3-h batch adsorption, aspresented in Table 2.
Taking the previously discussed findings into consider-ation, it is indicated that the general reduction in virusadsorption on the NPs, as influenced by the presence ofcells illustrated in Fig. 2, may be attributed to (1) directdisturbance of the interaction between viruses and NPs bythe cells, (2) less amount of NPs available for virus adsorp-tion resulting from the adsorption of NPs on the outside orinside of the bacterial cells, and (3) indirect disturbance ofinteraction between viruses and NPs by more dissolvedorganic carbon (DOC) resulting from the leakage of theintracellular organic solutes or from the lysis of the deadcells, as many studies have proven that DOC may inhibitphiX174 adsorption by competing for binding sites (e.g.,Schijven and Hassanizadeh 2000; Zhuang and Jin 2003;Zhang et al. 2010). This seems to be not applicable to thepromoted adsorption of viruses on γ-Fe2O3-B and Al2O3 inthe presence of the 6-h P. aeruginosa cells (Fig. 2). A possibleexplanation is that these two NPs might have enhanced theadsorption of the virus to the surface of the 6-h P. aeruginosacells. We inferred this from a recent study that shows thefacilitation of virus uptake into the host bacteria by silverand zinc oxide NPs via enhancing their adsorption (You etal. 2011). This likely concentrates the viruses around the cellsand facilitates the contact between the NPs and viruses forsubsequent adsorption and/or inactivation.
Compared to the 6-h cells, the 24-h cells had more viablecells as well as dead cells (Fig. 1), which may have accu-mulated more NPs and resulted in more DOC content and,subsequently, more significant direct and indirect distur-bance of interaction between viruses and NPs and lessamount of NPs available for virus adsorption in the presenceof the 24-h cells relative to the 6-h cells (Fig. 2). In addition,Walker et al. (2005) found that the cells in the stationaryphase were more adhesive to quartz surfaces than that in themid-log phase because the former had a more heterogeneousdistribution of charged functional groups on the bacterialsurface than the later, as surface charge heterogeneity candecrease electrostatic repulsion (Song et al. 1994; Elimelechet al. 2000). This suggests that, in the present study, the 24-hcells would be more favorably adsorbed by the NPs than the6-h cells and, thus, more pronounced to reduce virus ad-sorption on the NPs.
Effect of culture supernatants
During the growth period, the bacteria can exude EPSoutside the cell. The EPS is either bound to the cell surface(i.e., attached or capsular) or secreted outside the cell andreleased into solution (i.e., dissolved) (Omoike et al. 2004;Subramanian et al. 2010). In practice, the dissolved EPS isthe fraction that is contained in the supernatant and that canbe washed out from cells of microorganisms during centri-fugation, while the attached EPS is stable and is attached onthe cell walls of microorganisms during this process (Liuand Fang 2002; Subramanian et al. 2010). Therefore, underthe present experimental conditions, the supernatants mainlycontained EPS, relative to the controls.
Adsorption of viruses on the three NPs was significantlyreduced by the supernatants from all three bacterial strainsharvested at both culture ages of 6 and 24 h, except that the6-h B. subtilis supernatants had no significant effect on virusadsorption on γ-Fe2O3-B and Al2O3 (Fig. 3), suggesting theexudates-induced reduction in virus adsorption in the virus–NP–supernatant systems relative to their respective controls.Our results were contrary to those reported by Lipson andStotzky (1985), in which they found that reovirus adsorptionon kaolinite and montmorillonite was enhanced by the su-pernatants from both Serratia marcescens and E. coli. Thisdiscrepancy may have been the result of differences in theexperimental conditions (e.g., adsorbent type, bacterial spe-cies, and bacterial growth phase).
As with the attached EPS in the virus–NP–cell systemmentioned in the “Effect of bacterial cells” section, the EPSin the virus–NP–supernatant systems under the present ex-perimental conditions should also be negatively charged andbe preferably adsorbed by the NPs through electrostaticinteractions. Cao et al. (2011) also demonstrated that EPSadsorbed to goethite and clay minerals mainly through
Environ Sci Pollut Res

electrostatic interaction, hydrogen bonding, and chemicalbonding interactions (ligand exchange). In natural aqueousenvironments, pristine mineral surfaces become coated rap-idly by biogenic organic films (Bos et al. 1999). This mayreduce the chance for viruses interacting with the NPs byblocking sites for virus sorption and, consequently, inhibitedvirus adsorption (Zhuang and Jin 2003; Bradford et al.2006). On the other hand, virus inactivation as a result ofenzymatic attack is also possible (John and Rose 2005).Protease and elastase secreted by P. aeruginosa can cleaveinfluenza virus hemagglutinin (Callan et al. 1997). Nasser etal. (2002) reported that the inactivation of coxsackievirusand hepatitis A in soil saturated with suspensions of extra-cellular exudates from P. aeruginosa cultures increased,compared to buffered saline controls, suggesting the en-hancement of virus inactivation by microbial activity.Lipson and Stotzky (1985) contributed an antiviral compo-nent(s) to the enhanced adsorption of the viruses on kaolin-ite suspended in the supernatants from B. subtilis.
We, therefore, inferred that the overall virus adsorptionunder the presence of supernatants depended on the relativecontribution of the two competing processes: decrease invirus adsorption from site blocking of exudates in the su-pernatants versus increase in virus adsorption from virusinactivation by enzymatic attack. The significantly reducedvirus adsorption in most cases illustrated in Fig. 3 indicatedthat decrease in virus adsorption from site blocking of EPSmostly dominated over the increase in virus adsorption fromvirus inactivation by enzymatic attack and the trend becamemore significant with culture age or growth phase, as indi-cated by the more considerable reduction in virus adsorptionunder the presence of the 24-h supernatants compared to the6-h supernatants. The contribution of these two competingprocesses was likely comparative on virus adsorption ontoγ-Fe2O3 and Al2O3 under the presence of the 6-h P.aeruginosa or B. subtilis rather than the 24-h P. aeruginosaor B. subtilis (Fig. 3), probably because the exudates mayneed time to produce and accumulate before it becomeseffective to inhibit virus adsorption.
Effect of bacterial cultures
As cultures contain both cells and supernatants, the ob-served effect of cultures on virus adsorption on NPs waslikely a combined effect of cells and supernatants. The factthat virus adsorption percentages in the virus–NP–cultureand virus–NP–supernatant systems were significantly corre-lated (r=0.9924, P<0.001), while that in the virus–NP–culture and virus–NP–cell systems did not significantlycorrelate (r=0.2782, P>0.05), indicating that the effect ofsupernatants might have contributed mainly to the effect ofcultures.
On average, the presence of the 6-h cultures reducedvirus adsorption by 10.25±8.56 % (Fig. 4), which exceededthe sum of the reduction caused by the 6-h cells (0.74±0.84 %) and the 6-h culture supernatants (7.71±8.99 %)(Figs. 2 and 3). Likewise, more reduction in virus adsorption(24.57±6.89 %) resulting from the presence of the 24-hcultures than the sum of the reduction caused by the 24-hcells (2.13±0.93 %) and the 24-h supernatants (21.46±6.62 %) was found (Figs. 2, 3, and 4). The results furtherindicated the dominant contribution of supernatants to thecultures and partly confirmed our speculation that the effectof the cultures on virus adsorption resulted from the additiveeffect of supernatants and cells. Some observed discrepancymentioned previously can be attributed to the possible cellpropagation resulting from the unused nutrients in the cul-tures during the 3-h batch experiment, relative to the pres-ence of cells suspended with physiological saline (Table 3).The increasing indigenous bacterial numbers over time wasfound to be significantly correlated to a more rapid declineof bacteriophage MS2 (Yates et al. 1990). It is also possiblethat these discrepancies are attributable to the substantialrelative standard deviations associated with different typesof NPs or bacterial species.
In conclusion, our results showed that the presence ofcultures, as well as supernatants and cells, had a tendency toreduce virus adsorption on NPs, with the reduction extentvarying with the types of NPs and bacterial species andbecoming significant with culture age. We believe that theeffect of cultures was attributed to the additive effects ofsupernatants and cells, with the former contributing muchmore than the latter, implying that the bacterial exudatesmight be more crucial in the reduced virus adsorption thanthe bacterial cells. The results suggest that the presence ofsome specific bacteria may shift the fate of viruses in anopen environment with various underlying substrates, inwhich the exudates produced by the bacteria play a majorrole, and also suggest that estimates of virus removal effi-ciency by the NPs must always take into account the effectof bacteria which may be responsible for an overestimationof viral load in certain studies. The results from this studyare based on only three bacterial species, which certainlygives some insights into the behavior of NPs for removingviruses in natural systems. However, these studies should beextended to a wider and more representative range of con-ditions, such as microorganism with different exudates se-cretion capability and composition, and other constituents inthe aqueous solutions. Moreover, to avoid a possible distur-bance from the bacterial growth media, we need to conductfurther studies in their absence.
Acknowledgments The work was financially supported by the Na-tional Nature Science Foundation of China (40971129).
Environ Sci Pollut Res

References
Adams LK, Lyon DY, Alvarez PJJ (2006) Comparative eco-toxicity ofnanoscale TiO2, SiO2, and ZnO water suspensions. Water Res40(19):3527–3532
Adams MH (1959) Bacteriophages. Wiley Intersciences, New York, pp443–457
Bos R, van der Mei HC, Busscher HJ (1999) Physico-chemistry ofinitial microbial adhesive interactions—its mechanisms andmethods for study. FEMS Microbiol Rev 23(2):179–229
Bradford SA, Tadassa YF, Jin Y (2006) Transport of coliphage in thepresence and absence of manure suspension. J Environ Qual35:1692–1701
Brayner R, Ferrari-Iliou R, Brivois N, Djediat S, Benedetti MF, FiévetF (2006) Toxicological impact studies based on Escherichia colibacteria in ultrafine ZnO nanoparticles colloidal medium. NanoLett 6(4):866–870
Callan JR, Hartmann FA, West SE, Hinshaw VS (1997) Cleavage ofInfluenza Avirus H1 hemagglutinin by swine respiratory bacterialproteases. J Virol 71(10):7579–7585
Cao Y, Wei X, Cai P, Huang Q, Rong X, Liang W (2011) Preferentialadsorption of extracellular polymeric substances from bacteria onclay minerals and iron oxide. Colloid Surf B-Biointerfaces83(1):122–127
Chu Y, Jin Y, Yate MV (2000) Virus transport through saturated sandcolumns as affected by different buffer solutions. J Environ Qual29:1103–1110
Dowd SE, Pillai SD, Wang S, Corapcioglu MY (1998) Delineating thespecific influence of virus isoelectric point and size on virusadsorption and transport through sandy soils. Appl EnvironMicrobiol 64(2):405–410
Elimelech M, Nagai M, Ko CH, Ryan JN (2000) Relative insignificanceof mineral grain zeta potential to colloid transport in geochemicallyheterogeneous porous media. Environ Sci Technol 34:2143–2148
Gutierrez L, Li X, Wang JW, Nangmenyi G, Economy J,Kuhlenschmidt TB, Kuhlenschmidt MS, Nguyen TH (2009) Ad-sorption of rotavirus and bacteriophage MS2 using glass fibercoated with hematite nanoparticles. Water Res 43(20):5198–5208
Hu X, Mu L, Wen J, Zhou Q (2012) Covalently synthesized grapheneoxide-aptamer nanosheets for efficient visible-light photocatalysisof nucleic acids and proteins of viruses. Carbon 50:2772–2781
Jin Y, Flury M (2002) Fate and transport of viruses in porous media.Adv Agron 77:39–102
Jin Y, Pratt E, YatesMV (2000) Effect of mineral colloids on virus transportthrough saturated sand columns. J Environ Qual 29:532–539
John DE, Rose JB (2005) Review of factors affecting microbial sur-vival in groundwater. Environ Sci Technol 39(19):7345–7356
Kim S, Corapcioglu MY, Kim D (2003) Effect of dissolved organicmatter and bacteria on contaminant transport in riverbank filtra-tion. J Contam Hydrol 66:1–23
Koivunen J, Heinonen-Tanski H (2005) Inactivation of enteric micro-organisms with chemical disinfectants, UV irradiation and com-bined chemical/UV treatments. Water Res 39:1519–1526
Liga MV, Bryant EL, Colvin VL, Li Q (2011) Virus inactivation bysilver doped titanium dioxide nanoparticles for drinking watertreatment. Water Res 45:535–544
Lipson SM, Stotzky G (1985) Effect of bacteria on the inactivation andadsorption on clay minerals of reovirus. Can J Microbiol 31:730–735
Link N, Brunner TJ, Dreesen IAJ, Stark WJ, Fussenegger M (2007)Inorganic nanoparticles for transfection of mammalian cells andremoval of viruses from aqueous solutions. Biotechnol Bioeng98:1083–1093
Liu H, Fang HHP (2002) Characterization of electrostatic binding sitesof extracellular polymers by linear programming analysis of titra-tion data. Biotechnol Bioeng 80:806–811
Martinez RE, Smith DS, Kulczycki E, Ferris FG (2002) Determinationof intrinsic bacterial surface acidity constants using a Donnanshell model and a continuous pKa distribution method. J ColloidInterface Sci 253:130–139
Nasser AM, Glozman R, Nitzan Y (2002) Contribution of microbialactivity to virus reduction in saturated soil. Water Res36(10):2589–2595
Nangmenyi G, Li X, Mehrabi S, Mintz E, Economy J (2011) Silver-modified iron oxide nanoparticle impregnated fiberglass for dis-infection of bacteria and viruses in water. Mater Lett 65:1191–1193
Omoike A, Chorover J, Kwon KD, Kubicki JD (2004) Adhesion ofbacterial exopolymers to r-FeOOH: inner-sphere complexation ofphosphodiester groups. Langmuir 20:11108–11114
Ryan JN, Harvey RW, Metge D, Elimelech M, Navigato T, Pieper AP(2002) Field and laboratory investigations of inactivation of vi-ruses (PRD1 and MS2) attached to iron oxide-coated quartz sand.Environ Sci Technol 36: 2403–2413
Schijven JF, Hassanizadeh SM (2000) Removal of viruses by soilpassage: overview of modeling, processes, and parameters. CritRev Environ Sci Technol 30(1):49–127
Shen L, Zhao B, Zhang J, Chen J, Zheng H (2010) Virus adsorptiononto nano-sized iron oxides as affected by different backgroundsolutions. Environ Sci (in Chinese) 31(4):983–989
Song LF, Johnson PR, Elimelech M (1994) Kinetics of colloid depo-sition onto heterogeneously charged surfaces in porous media.Environ Sci Technol 28:1164–1171
Su HL, Chou CC, Hung DJ, Lin SH, Pao IC, Lin JH, Huang FL, DongRX, Lin JJ (2009) The disruption of bacterial membrane integritythrough ROS generation induced by nanohybrids of silver andclay. Biomaterials 30(30):5979–5987
Subramanian SB, Yan S, Tyagi RD, Surampalli RY (2010) Extracellu-lar polymeric substances (EPS) producing bacterial strains ofmunicipal wastewater sludge: isolation, molecular identification,EPS characterization and performance for sludge settling anddewatering. Water Res 44:2253–2266
Van Raij B, Peech M (1972) Electrochemical properties of someoxisols and alfisols of the tropics. Soil Sci Soc Am Proc36:587–593
Walker SL, Redman JA, Elimelech M (2005) Influence of growthphase on bacterial deposition: interaction mechanisms inpacked-bed column and radial stagnation point flow systems.Environ Sci Technol 39:6405–6411
Wei X, Fang L, Cai P, Huang Q, Chen H, Liang W, Rong X(2011) Influence of extracellular polymeric substances (EPS)on Cd adsorption by bacteria. Environ Pollut 159:1369–1374
Yates MV, Stetzenbach LD, Gerba CP, Sinclair NA (1990) The effectof indigenous bacteria on virus survival in ground water. JEnviron Sci Health Part A- Environ Sci Eng Toxicol 25(1):81–100
You J, Zhang Y, Hu Z (2011) Bacteria and bacteriophage inactivationby silver and zinc oxide nanoparticles. Colloid Surf B-Biointerfaces 85(2):161–167
Zhang H, Zhang J, Zhao B, Zhang C (2010) Removal of bacterio-phages MS2 and phiX174 from aqueous solutions using a redsoil. J Hazard Mater 180(1–3):640–647
Zhang L, Jiang Y, Ding Y, Povey M, York D (2007) Investigation intothe antibacterial behaviour of suspensions of ZnO nanoparticles(ZnO nanofluids). J Nanopart Res 9:479–489
Zhao B, Zhang H, Zhang J, Jin Y (2008) Virus adsorption and inacti-vation in soil as influenced by autochthonous microorganisms andwater content. Soil Biol Biochem 40(3):649–659
Zhuang J, Jin Y (2003) Virus retention and transport as influenced bydifferent forms of soil organic matter. J Environ Qual 32(3):816–823
Environ Sci Pollut Res
![Adsorption of lisinopril and chlorpheniramine from aqueous …carbonlett.org/Upload/files/CARBONLETT/[012-022]-02.pdf · 12 Adsorption of lisinopril and chlorpheniramine from aqueous](https://img.dokumen.tips/doc/110x75/5a87c54a7f8b9ac96a8dff3d/adsorption-of-lisinopril-and-chlorpheniramine-from-aqueous-012-022-02pdf12.jpg)




























