Embed Size (px)
Citation preview

Pre-mRNA Splicing
Lecture 1
Douglas Black
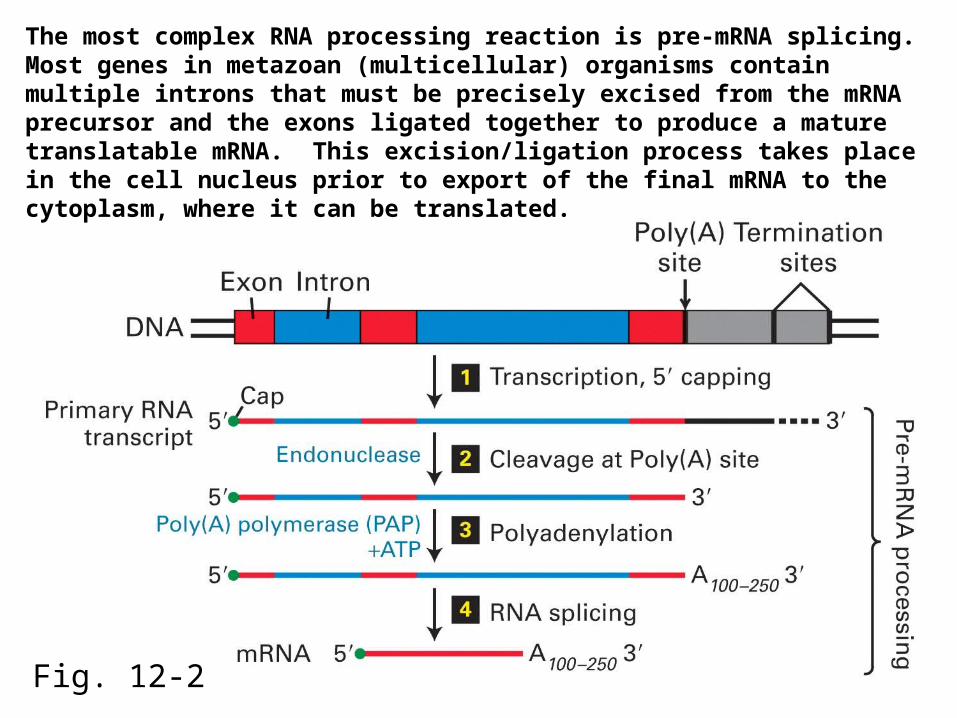
Fig. 12-2
The most complex RNA processing reaction is pre-mRNA splicing. Most genes in metazoan (multicellular) organisms contain multiple introns that must be precisely excised from the mRNA precursor and the exons ligated together to produce a mature translatable mRNA. This excision/ligation process takes place in the cell nucleus prior to export of the final mRNA to the cytoplasm, where it can be translated.
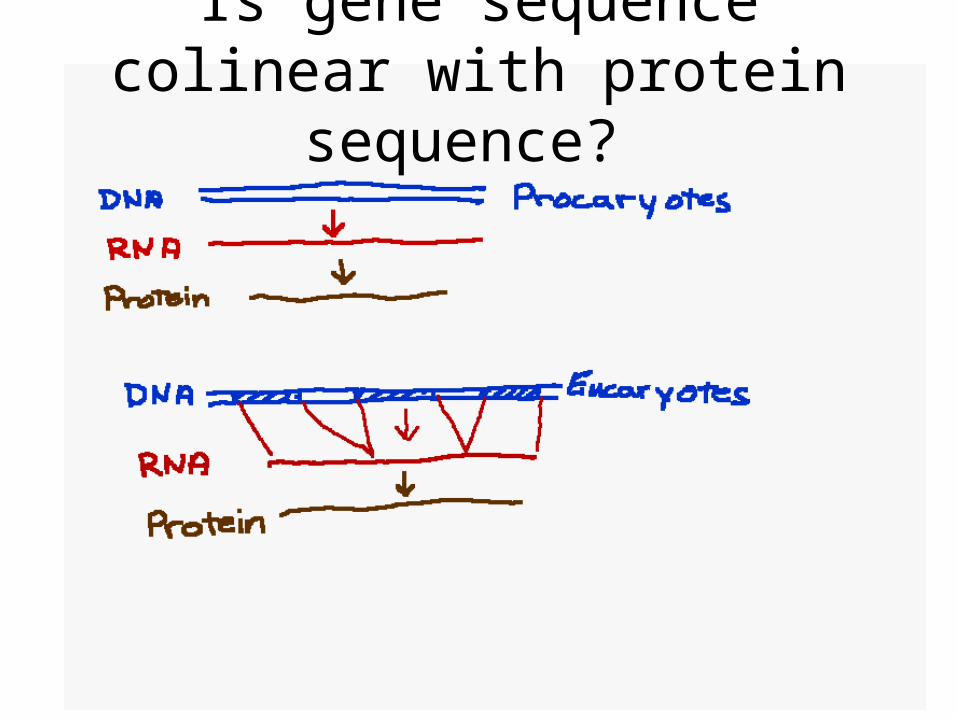
Is gene sequence colinear with protein sequence?
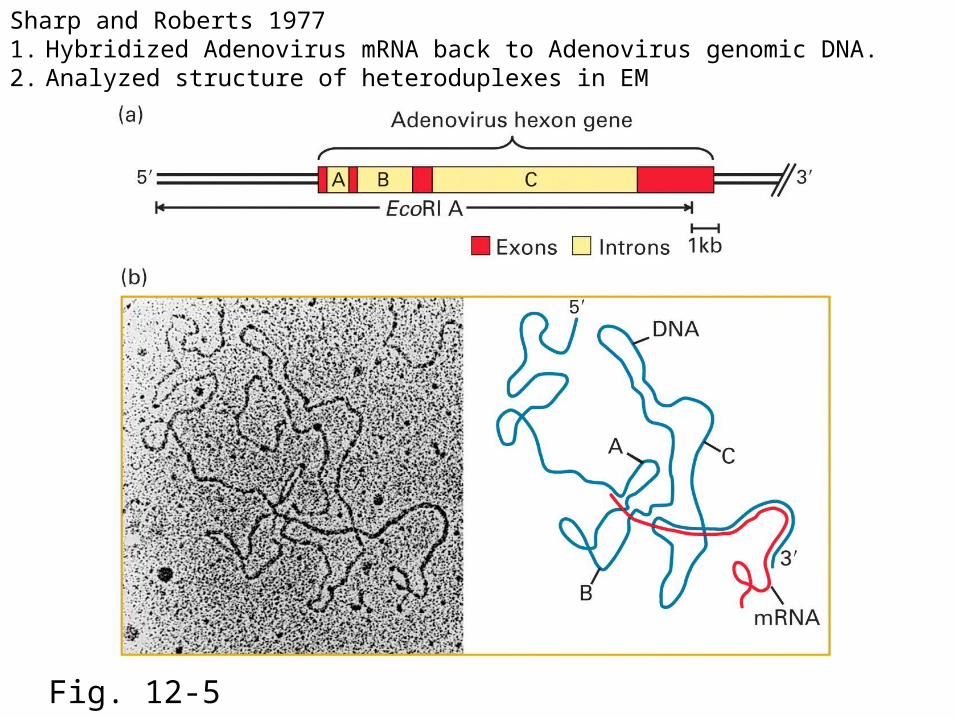
Fig. 12-5
Sharp and Roberts 19771. Hybridized Adenovirus mRNA back to Adenovirus genomic DNA.2. Analyzed structure of heteroduplexes in EM

Fig. 12-6
- Introns begin and end with splice sites that are not identical but conform to consensus sequences. - Introns always begin with GU encompassed within a larger 5’ splice site consensus.- Introns always end with AG, which is preceded by a polypyrimidine tract and the branch point sequence.- These sequences tell the splicing apparatus where to cut and join the RNA.
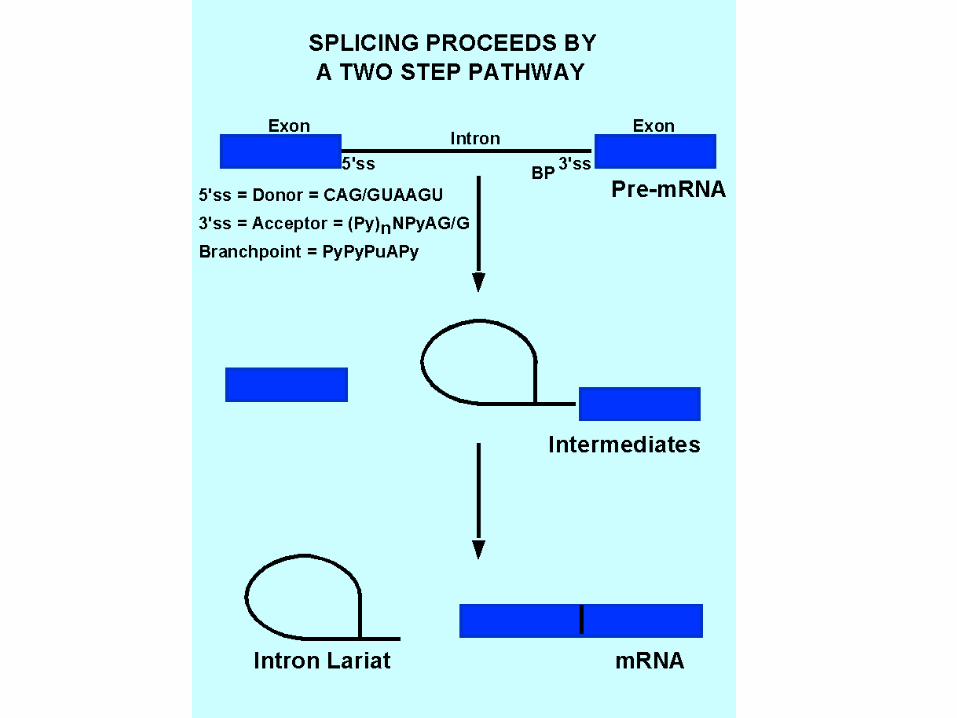
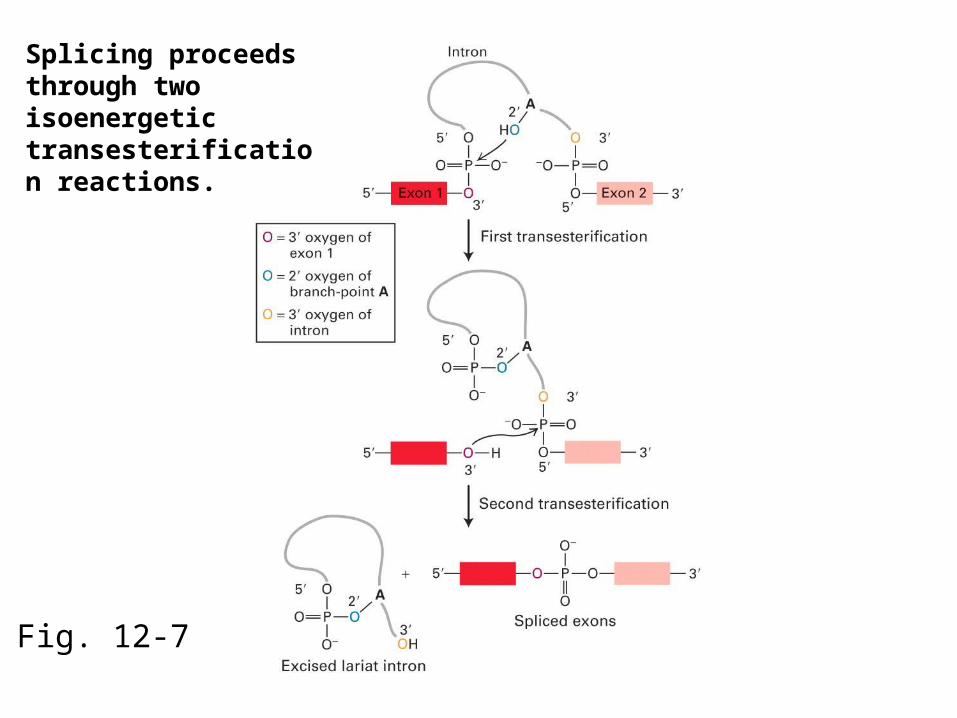
Fig. 12-7
Splicing proceeds through two isoenergetic transesterification reactions.
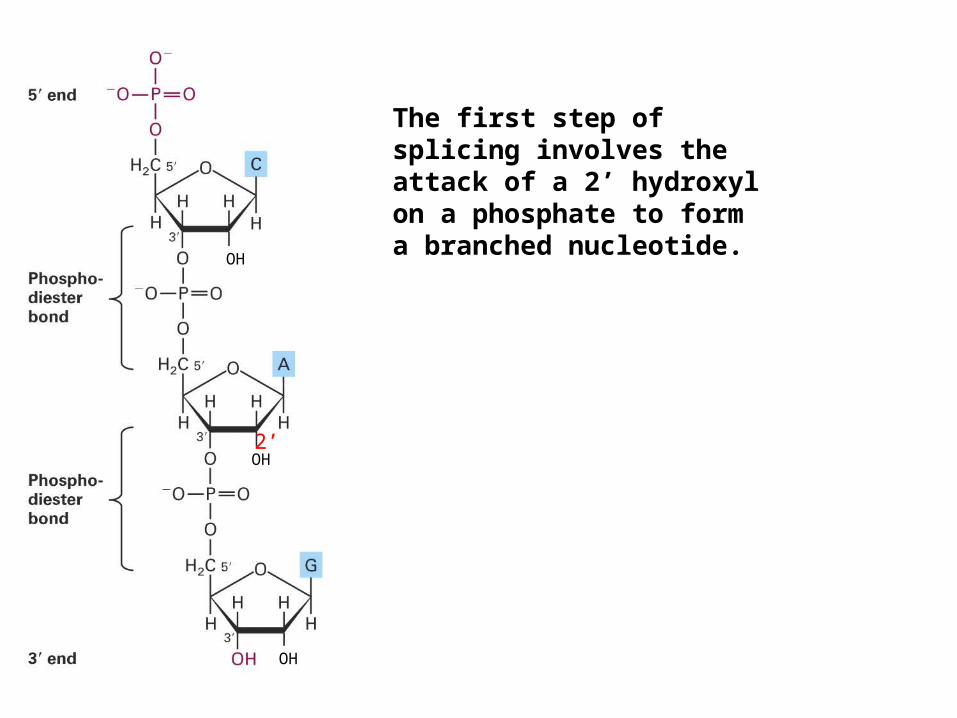
OH
OH
OH
2’
The first step of splicing involves the attack of a 2’ hydroxyl on a phosphate to form a branched nucleotide.
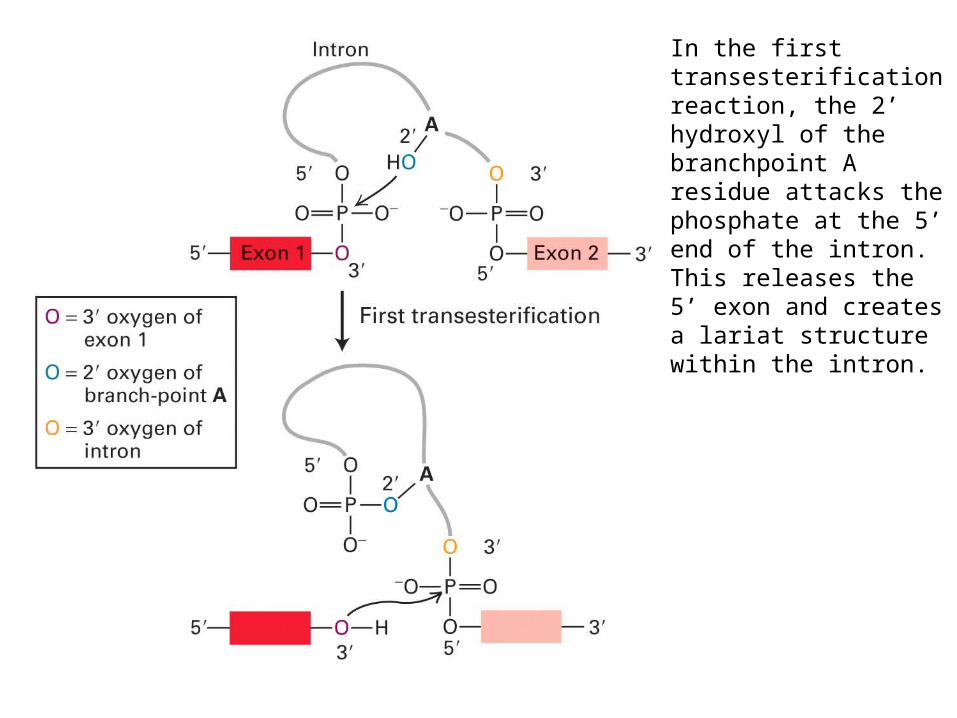
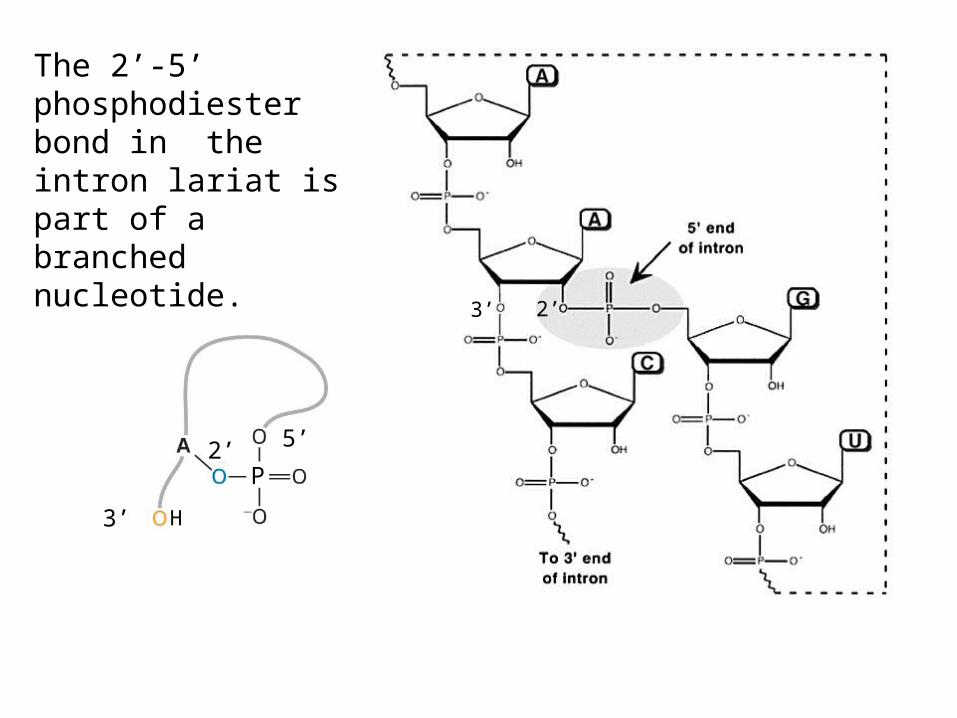
In the first transesterification reaction, the 2’ hydroxyl of the branchpoint A residue attacks the phosphate at the 5’ end of the intron.This releases the 5’ exon and creates a lariat structure within the intron.
3’
5’2’
P
P
3’
The 2’-5’ phosphodiester bond in the intron lariat is part of a branched nucleotide.
H
3’ 2’
OH
OH
OH
2’
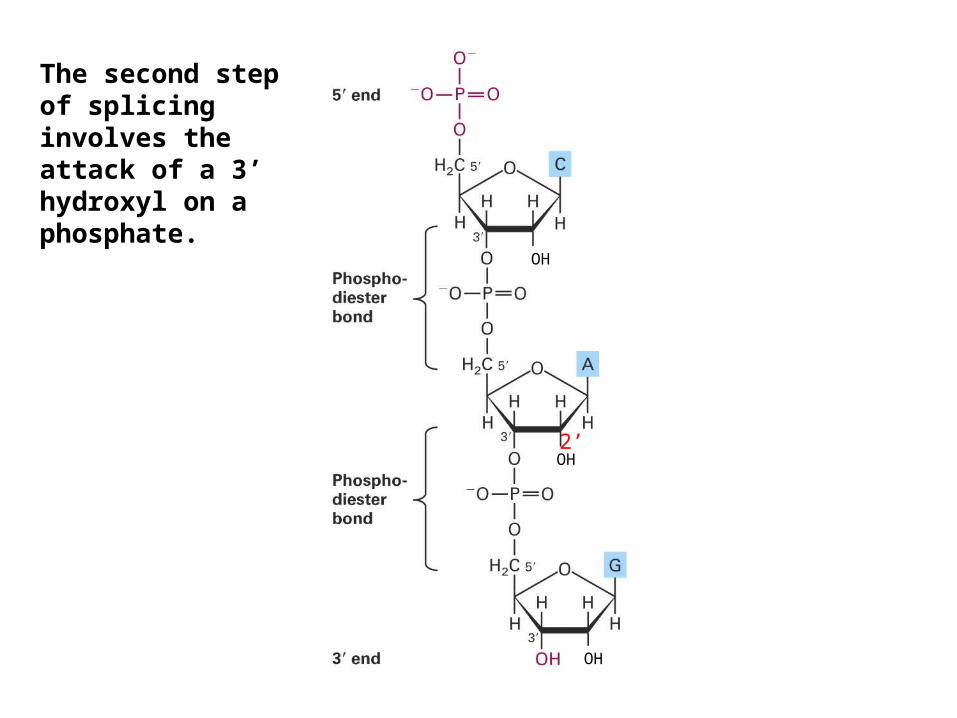
The second step of splicing involves the attack of a 3’ hydroxyl on a phosphate.
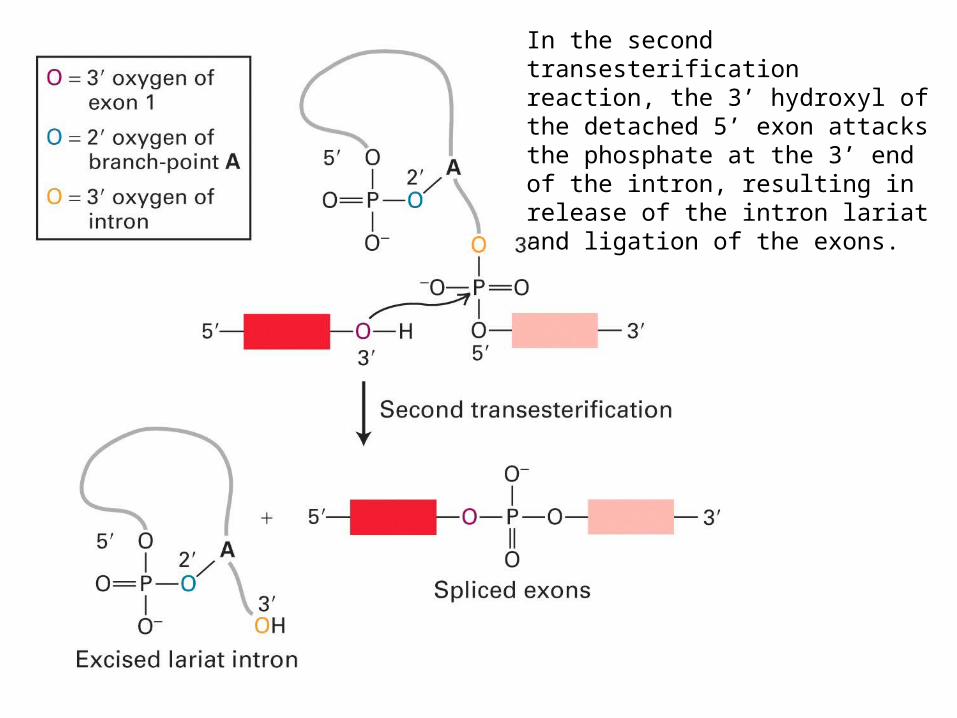
In the second transesterification reaction, the 3’ hydroxyl of the detached 5’ exon attacks the phosphate at the 3’ end of the intron, resulting in release of the intron lariat and ligation of the exons.
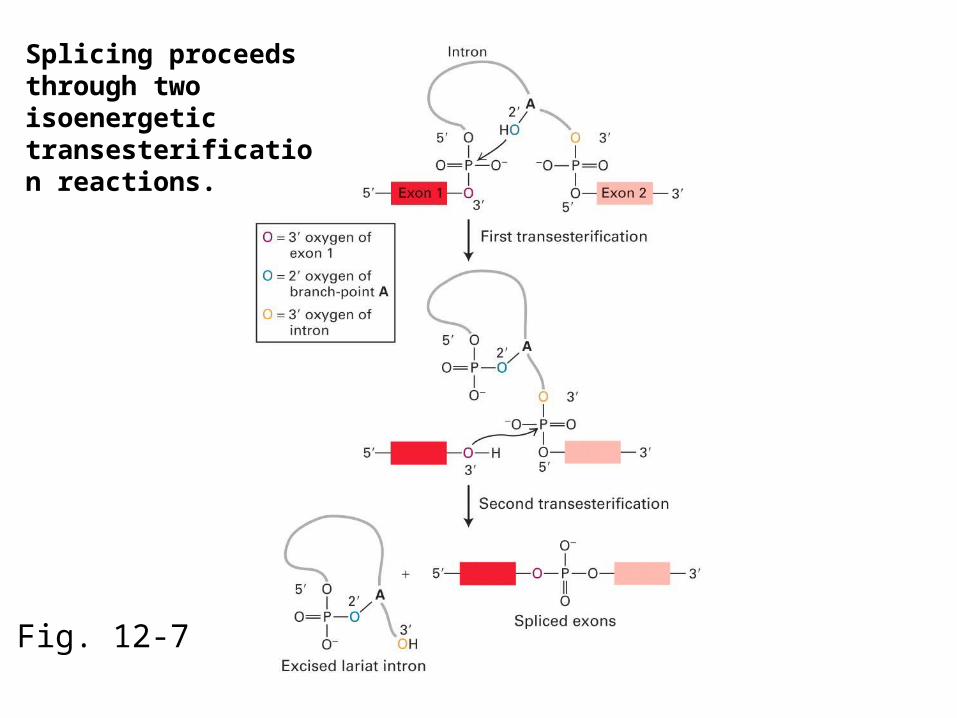
Fig. 12-7
Splicing proceeds through two isoenergetic transesterification reactions.
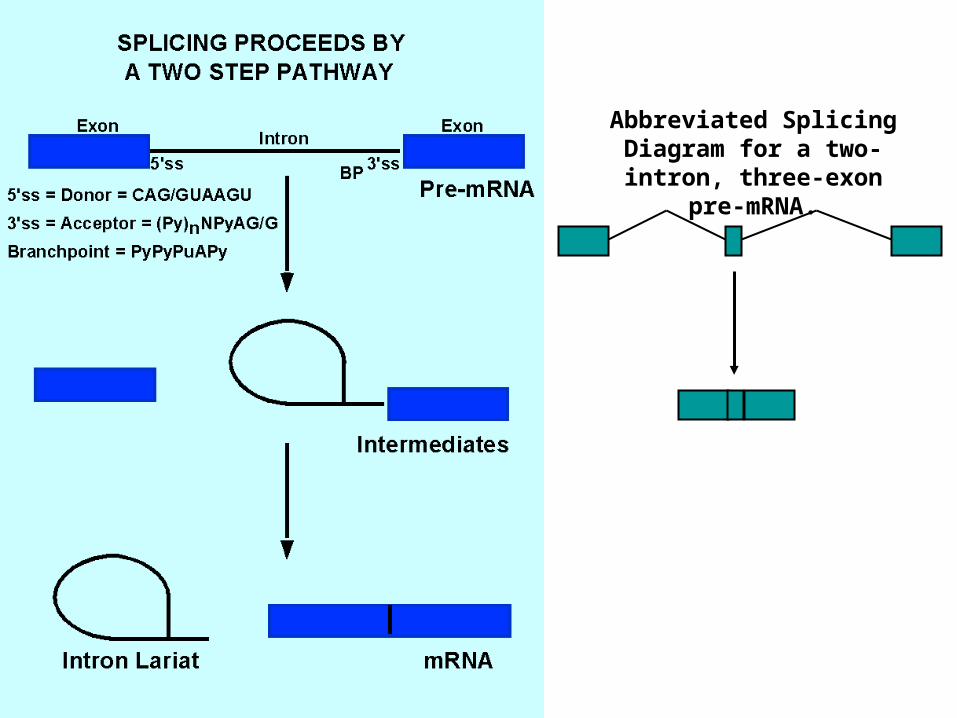
Abbreviated Splicing Diagram for a two-intron,
three-exon pre-mRNA.
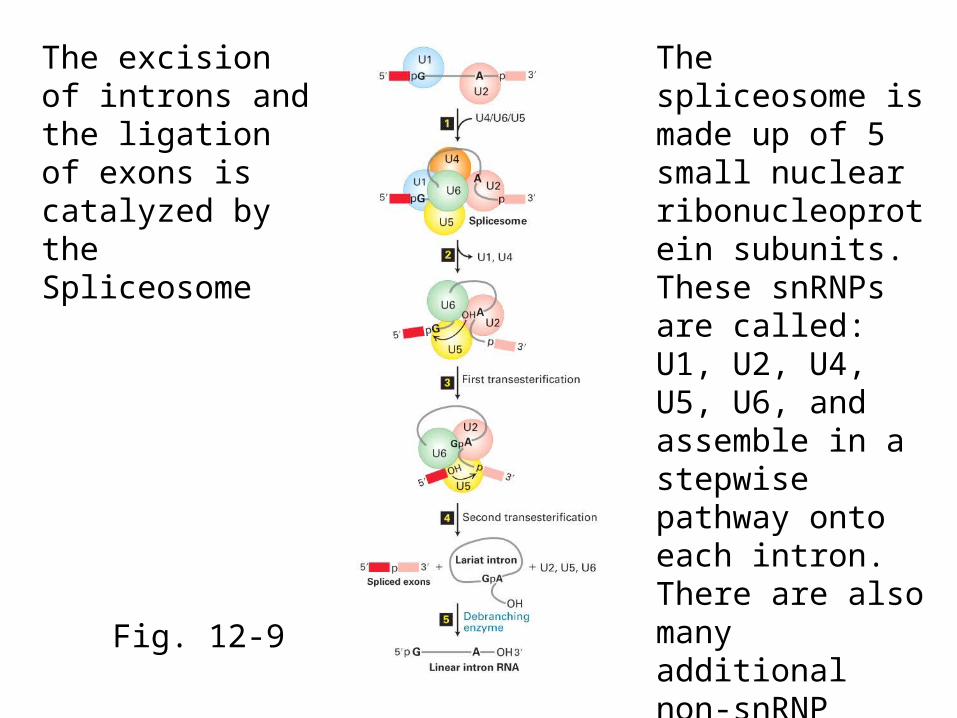
Fig. 12-9
The excision of introns and the ligation of exons is catalyzed by the Spliceosome
The spliceosome is made up of 5 small nuclear ribonucleoprotein subunits. These snRNPs are called: U1, U2, U4, U5, U6, and assemble in a stepwise pathway onto each intron. There are also many additional non-snRNP proteins in the spliceosome.
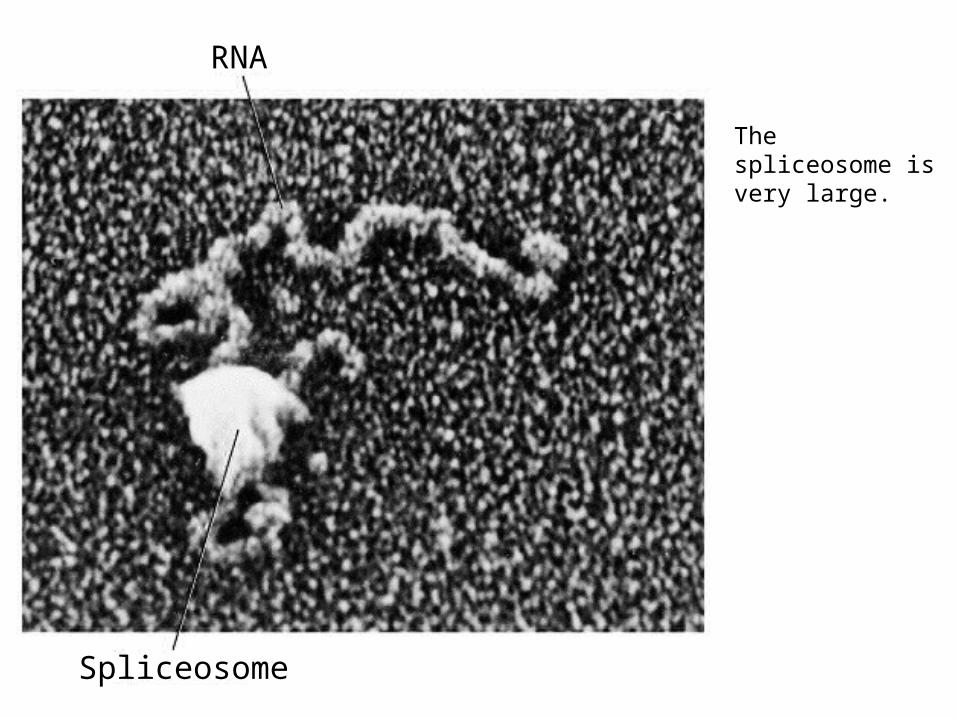
RNA
Spliceosome
The spliceosome is very large.

The spliceosome is made up of 5 small nuclear Ribonucleoproteins - snRNPs.
• named U1, U2, U4, U5 and U6
• Each snRNP is a small particle composed of a small nuclear RNA molecule bound by a specific set of proteins
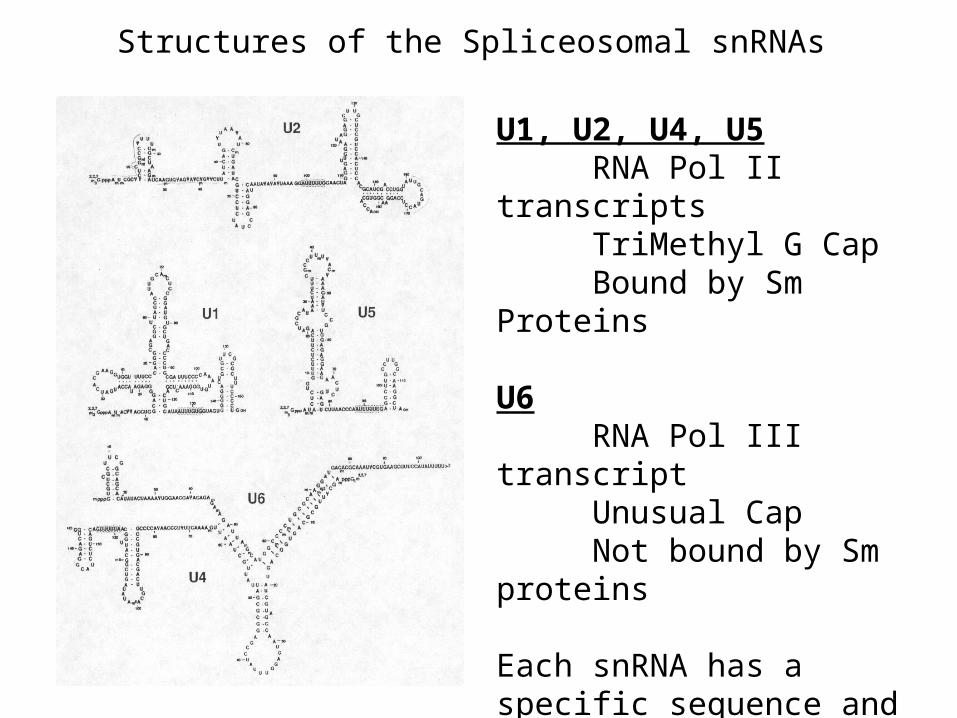
Structures of the Spliceosomal snRNAs
U1, U2, U4, U5RNA Pol II transcriptsTriMethyl G CapBound by Sm Proteins
U6RNA Pol III transcriptUnusual CapNot bound by Sm proteins
Each snRNA has a specific sequence and secondary structure and is bound by additional specific proteins
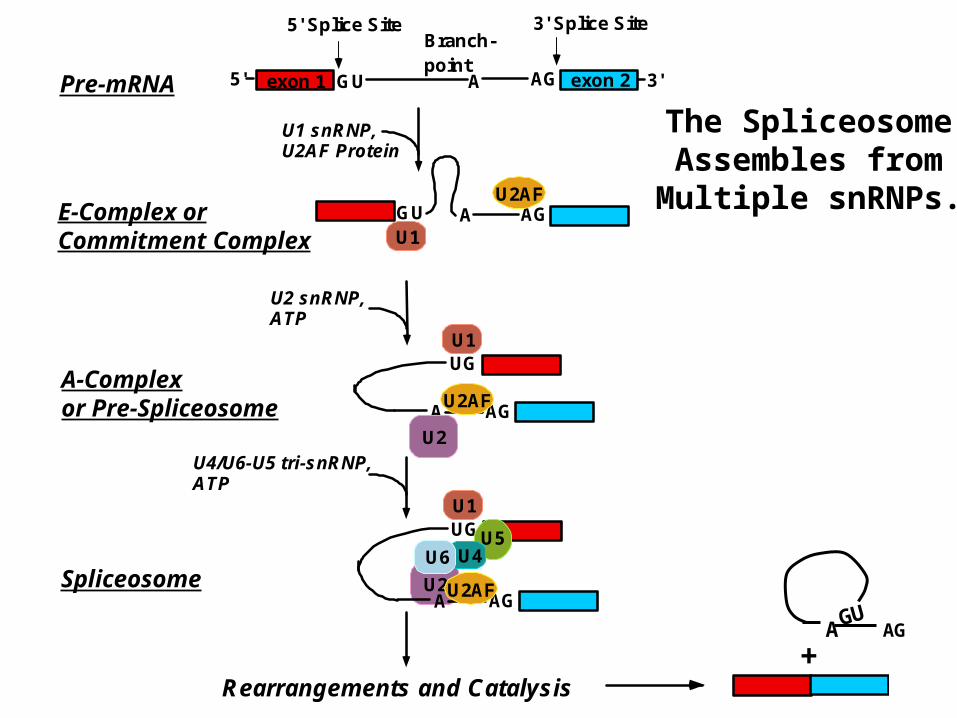
AG
UG
AU2AF
U1
U2
exon 15' GU 3'exon 2A AG
5' Splice Site 3' Splice SiteBranch-point
U1 snRNP,U2AF Protein
GU AGAU2AF
U1
U2 snRNP,ATP
U4/U6-U5 tri-snRNP,ATP
Rearrangements and Catalysis
U2
UG
AGAU2AF
U1
U5U4U6
Pre-mRNA
E-Complex orCommitment Complex
A-Complexor Pre-Spliceosome
Spliceosome
The SpliceosomeAssembles fromMultiple snRNPs.
+AGAGU
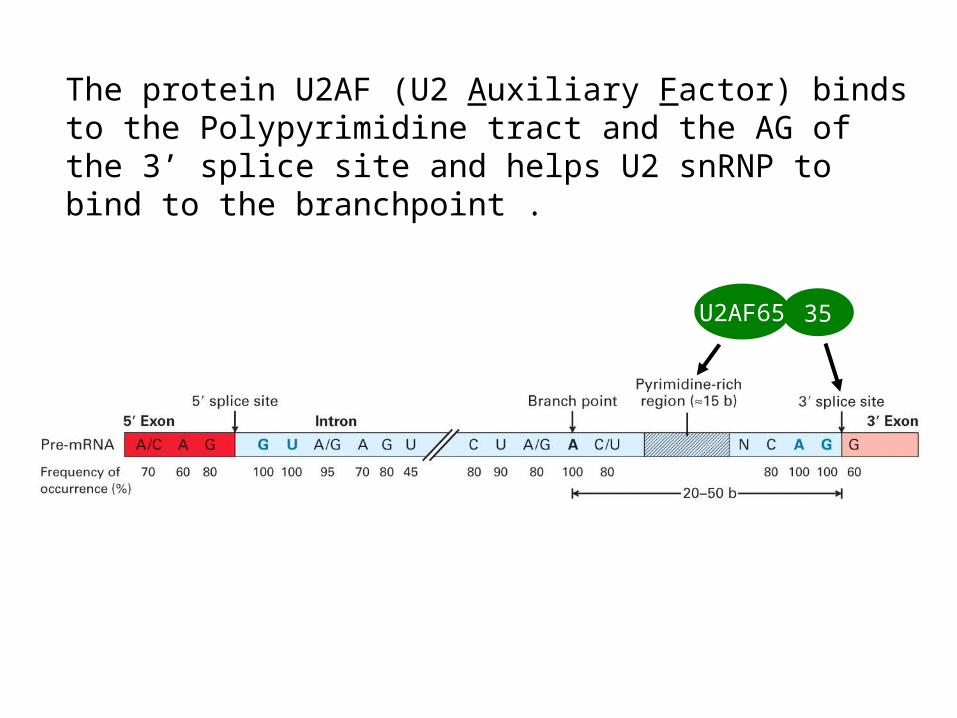
The protein U2AF (U2 Auxiliary Factor) binds to the Polypyrimidine tract and the AG of the 3’ splice site and helps U2 snRNP to bind to the branchpoint .
35U2AF65
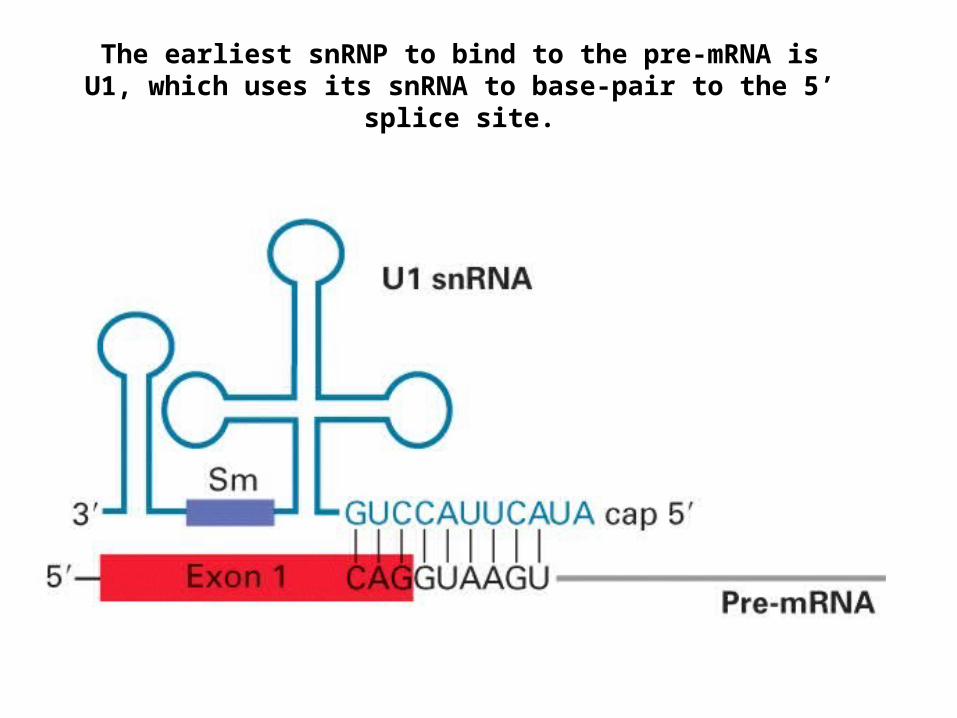
The earliest snRNP to bind to the pre-mRNA is U1, which uses its snRNA to base-pair to the 5’ splice site.
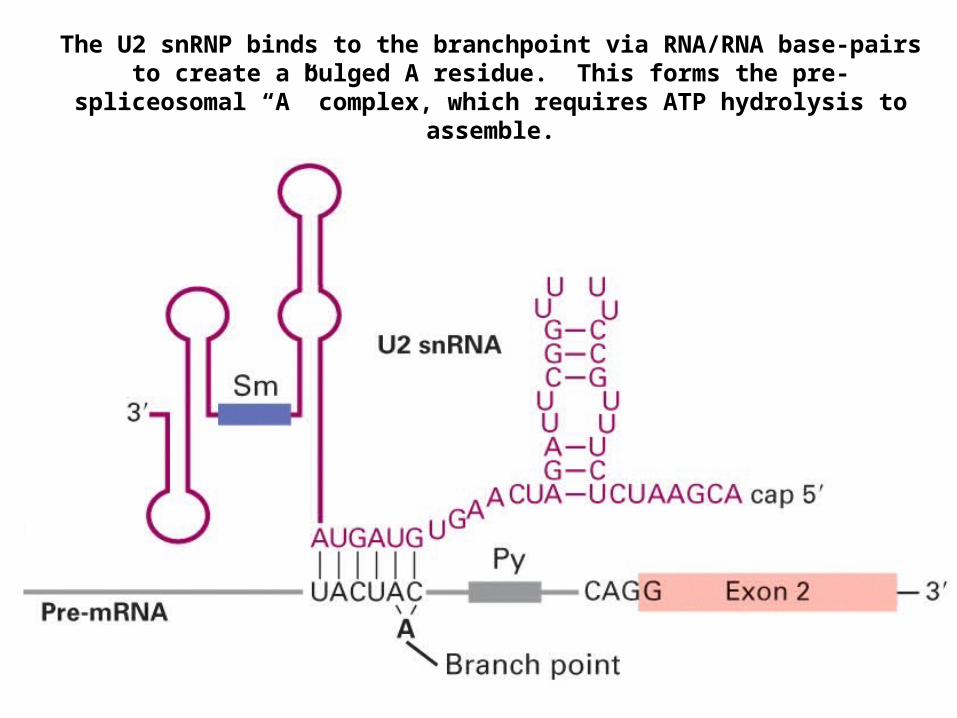
The U2 snRNP binds to the branchpoint via RNA/RNA base-pairs to create a bulged A residue. This forms the pre-spliceosomal “A” complex, which
requires ATP hydrolysis to assemble.
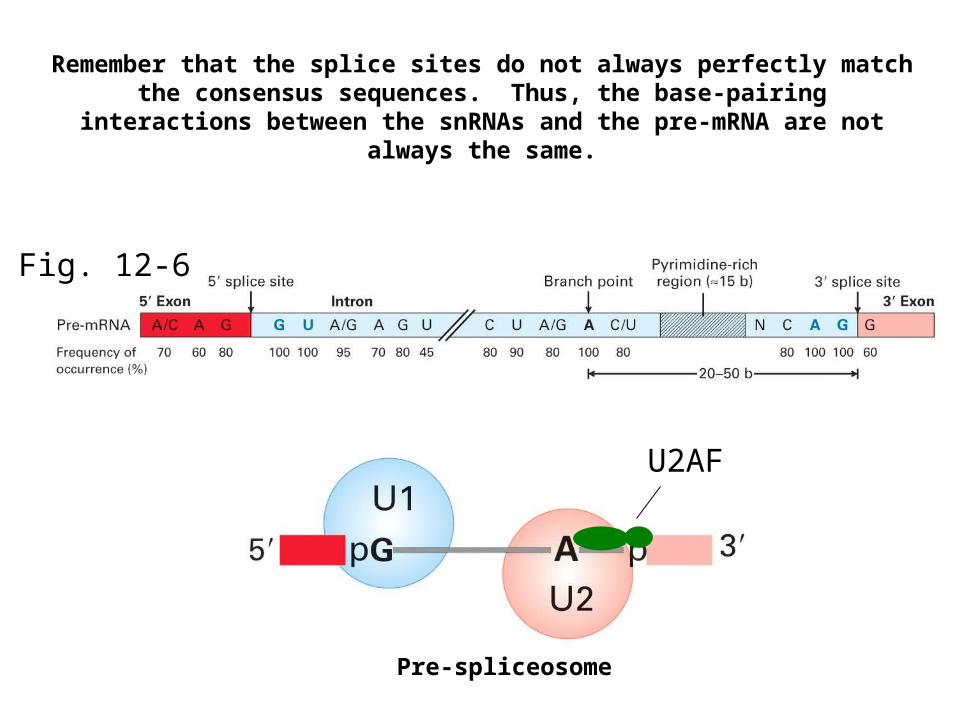
Fig. 12-6
U2AF
Remember that the splice sites do not always perfectly match the consensus sequences. Thus, the base-pairing interactions between the
snRNAs and the pre-mRNA are not always the same.
Pre-spliceosome
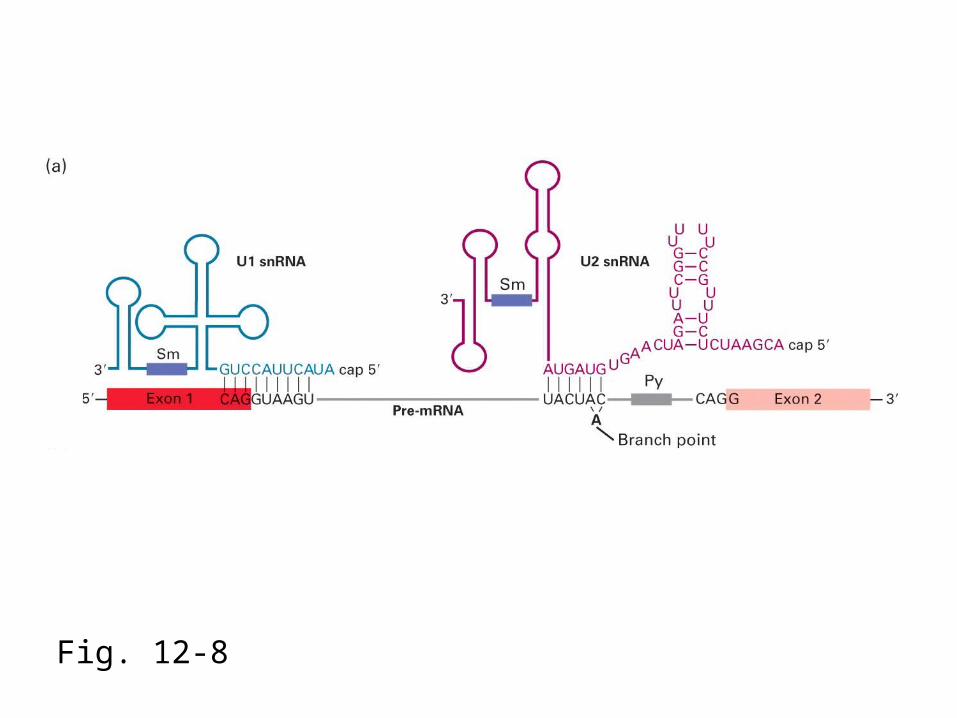
Fig. 12-8
Pre-mRNA
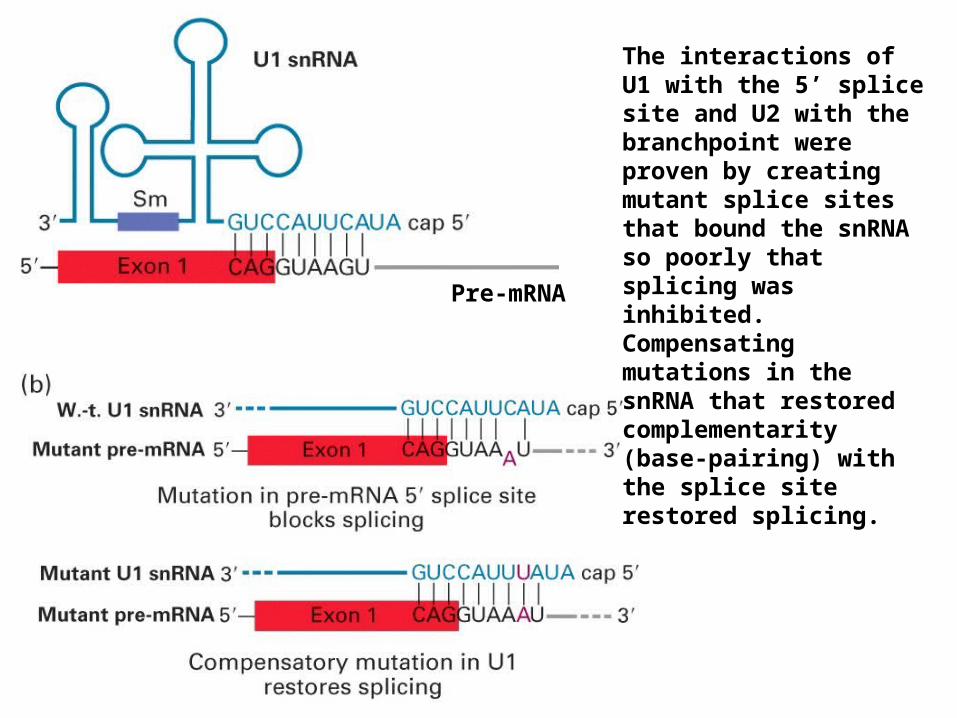
The interactions of U1 with the 5’ splice site and U2 with the branchpoint were proven by creating mutant splice sites that bound the snRNA so poorly that splicing was inhibited. Compensating mutations in the snRNA that restored complementarity (base-pairing) with the splice site restored splicing.
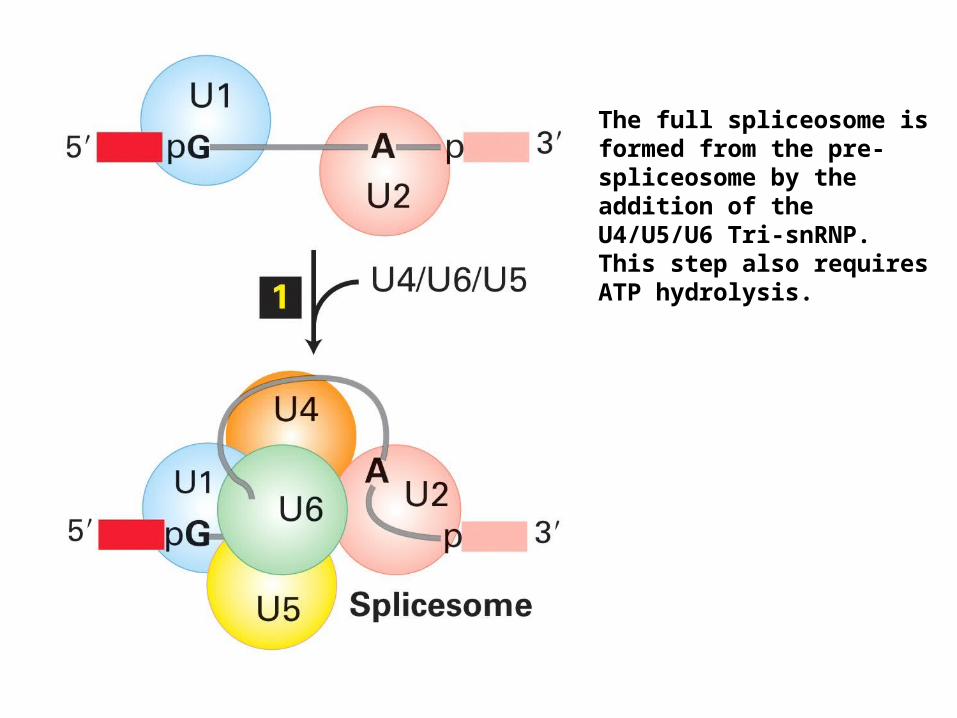
The full spliceosome is formed from the pre-spliceosome by the addition of the U4/U5/U6 Tri-snRNP. This step also requires ATP hydrolysis.
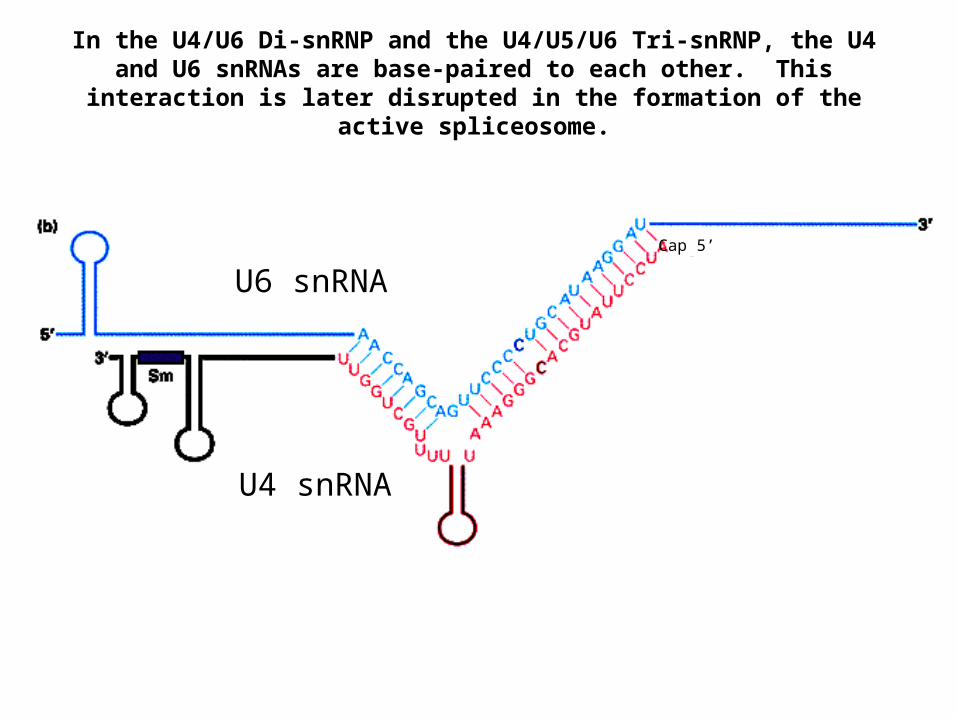
U6 snRNA
U4 snRNA
Cap 5’
In the U4/U6 Di-snRNP and the U4/U5/U6 Tri-snRNP, the U4 and U6 snRNAs are base-paired to each other. This interaction is later disrupted
in the formation of the active spliceosome.
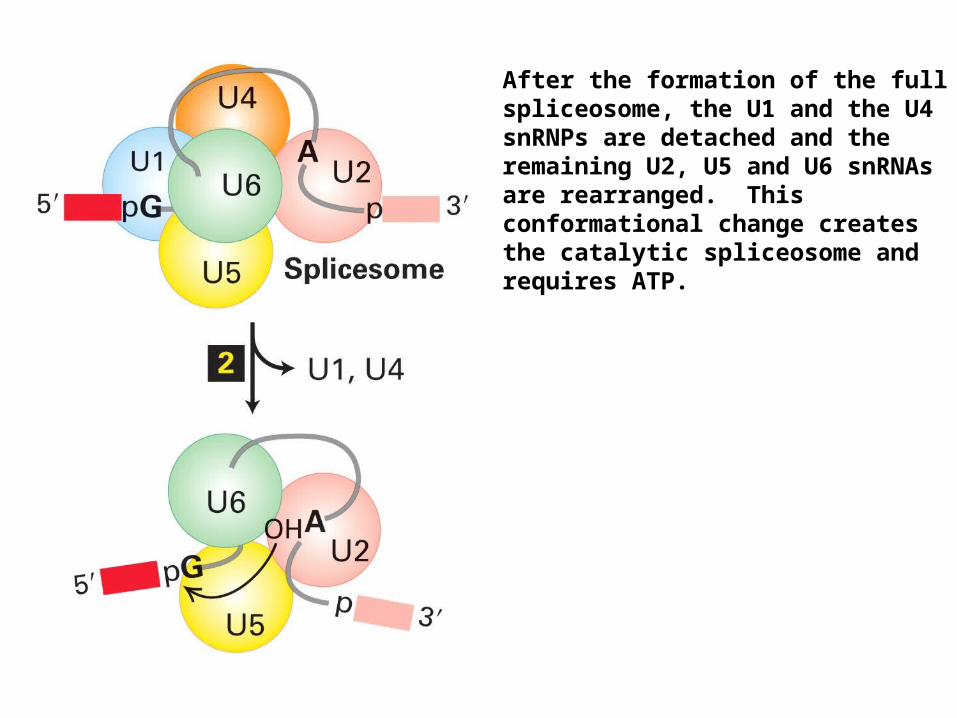
After the formation of the full spliceosome, the U1 and the U4 snRNPs are detached and the remaining U2, U5 and U6 snRNAs are rearranged. This conformational change creates the catalytic spliceosome and requires ATP.
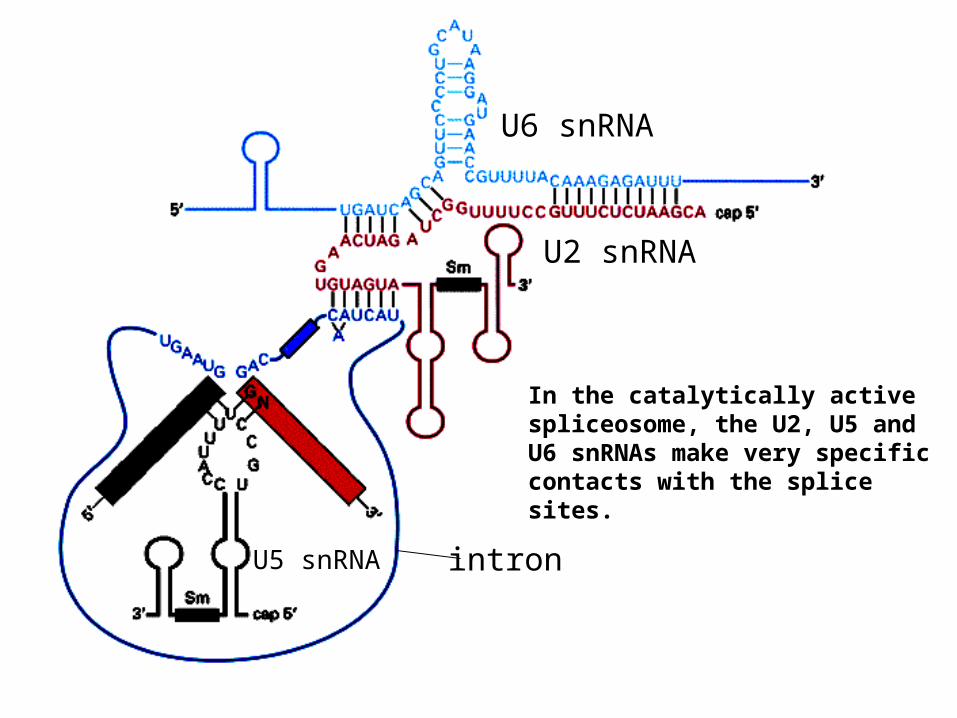
U2 snRNA
U6 snRNA
U5 snRNA intron
In the catalytically active spliceosome, the U2, U5 and U6 snRNAs make very specific contacts with the splice sites.
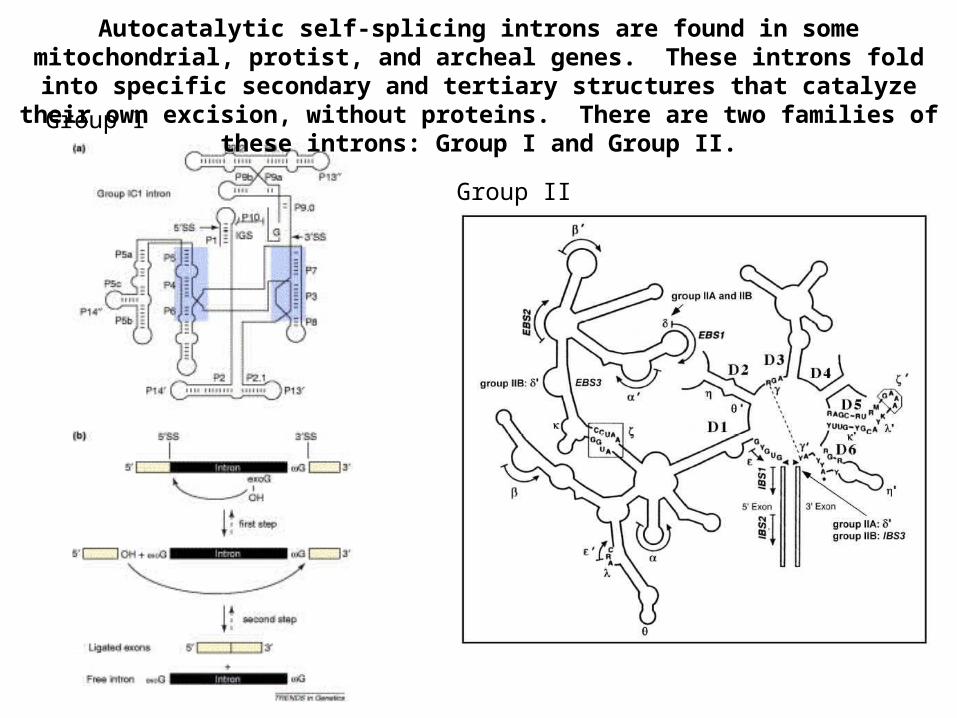
Autocatalytic self-splicing introns are found in some mitochondrial, protist, and archeal genes. These introns fold into specific secondary and tertiary structures
that catalyze their own excision, without proteins. There are two families of these introns: Group I and Group II.Group I
Group II
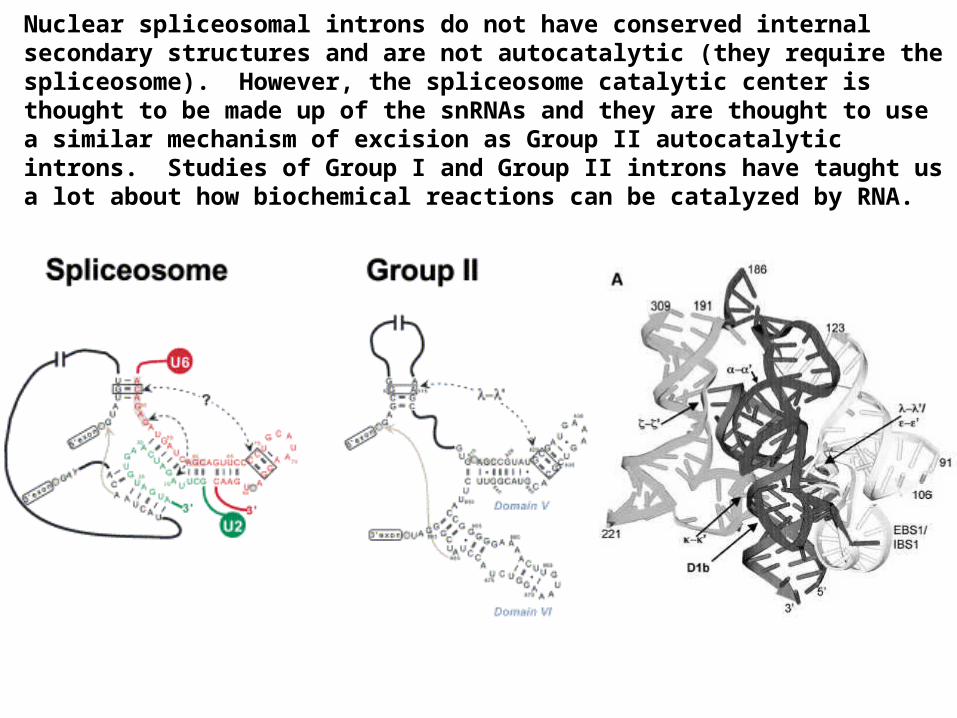
Nuclear spliceosomal introns do not have conserved internal secondary structures and are not autocatalytic (they require the spliceosome). However, the spliceosome catalytic center is thought to be made up of the snRNAs and they are thought to use a similar mechanism of excision as Group II autocatalytic introns. Studies of Group I and Group II introns have taught us a lot about how biochemical reactions can be catalyzed by RNA.
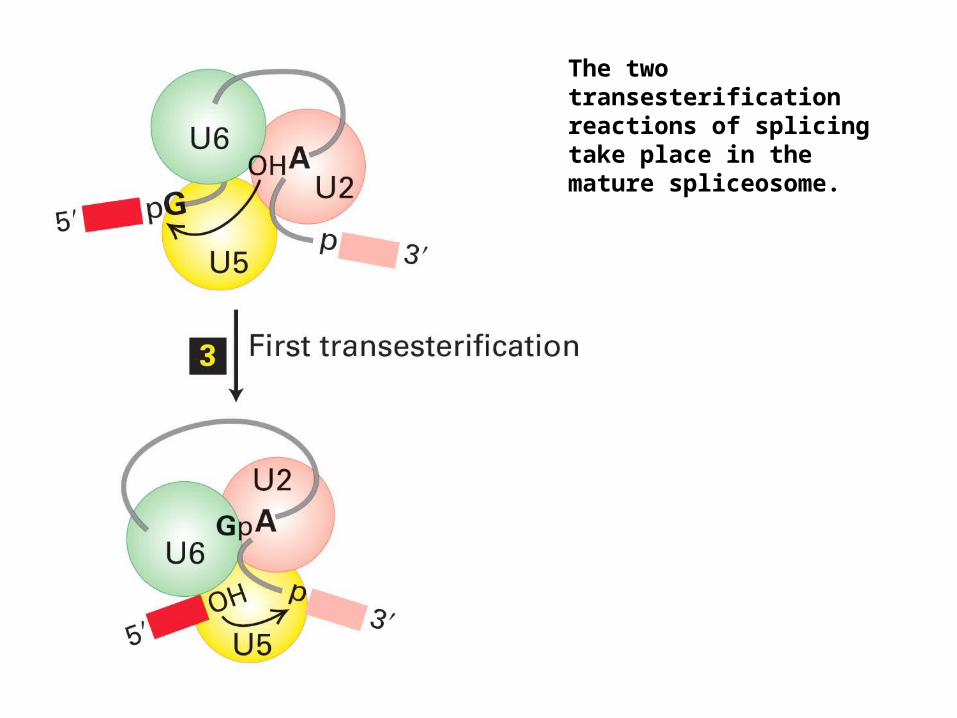
The two transesterification reactions of splicing take place in the mature spliceosome.
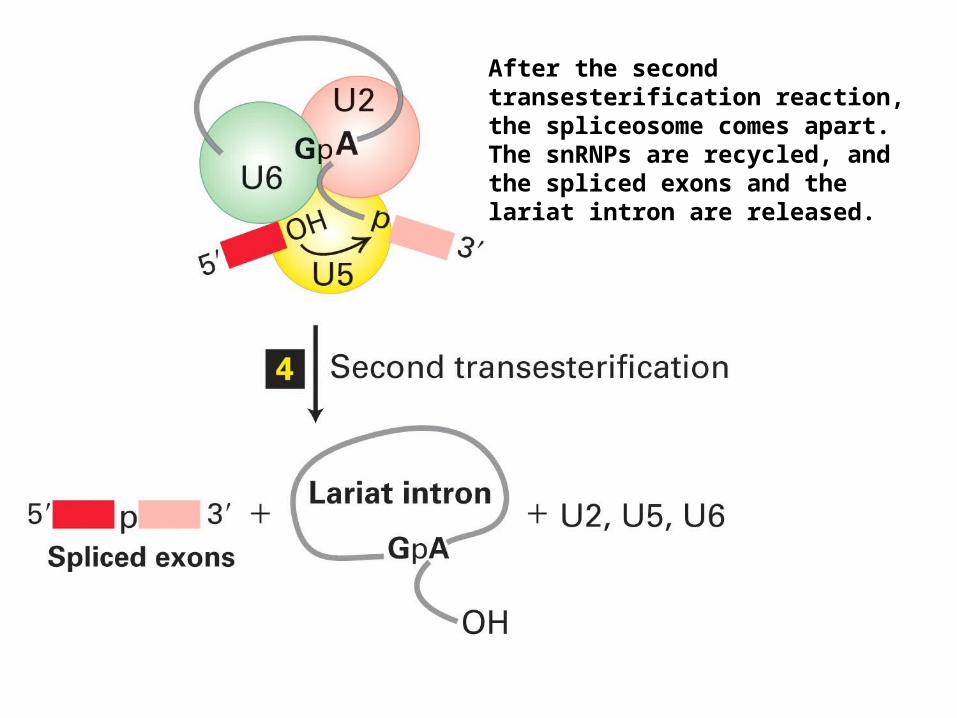
After the second transesterification reaction, the spliceosome comes apart. The snRNPs are recycled, and the spliced exons and the lariat intron are released.
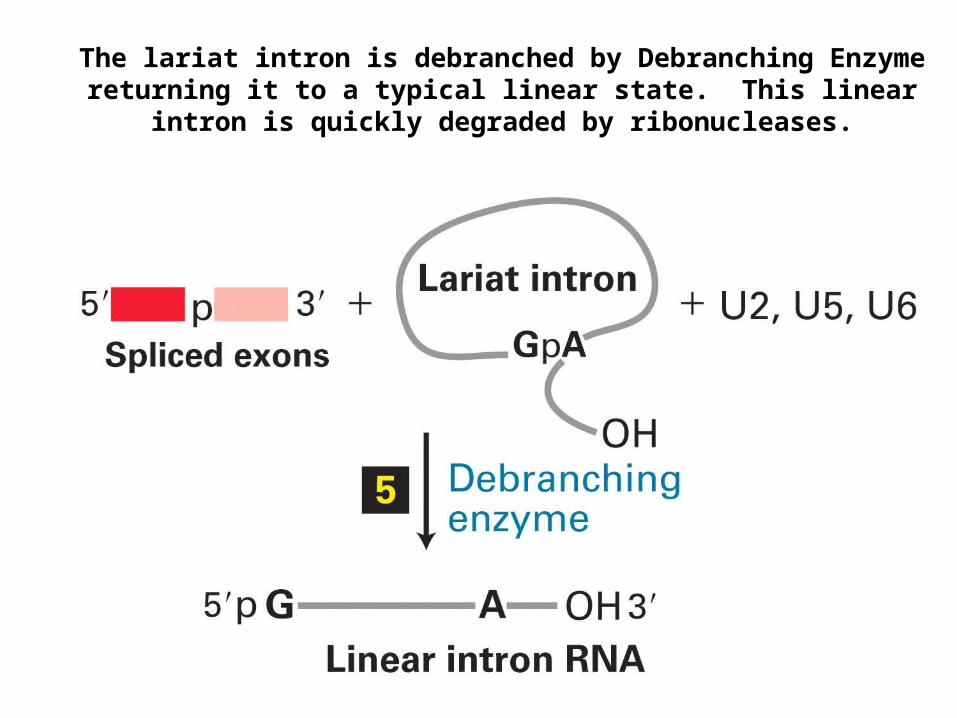
The lariat intron is debranched by Debranching Enzyme returning it to a typical linear state. This linear intron is quickly degraded by
ribonucleases.
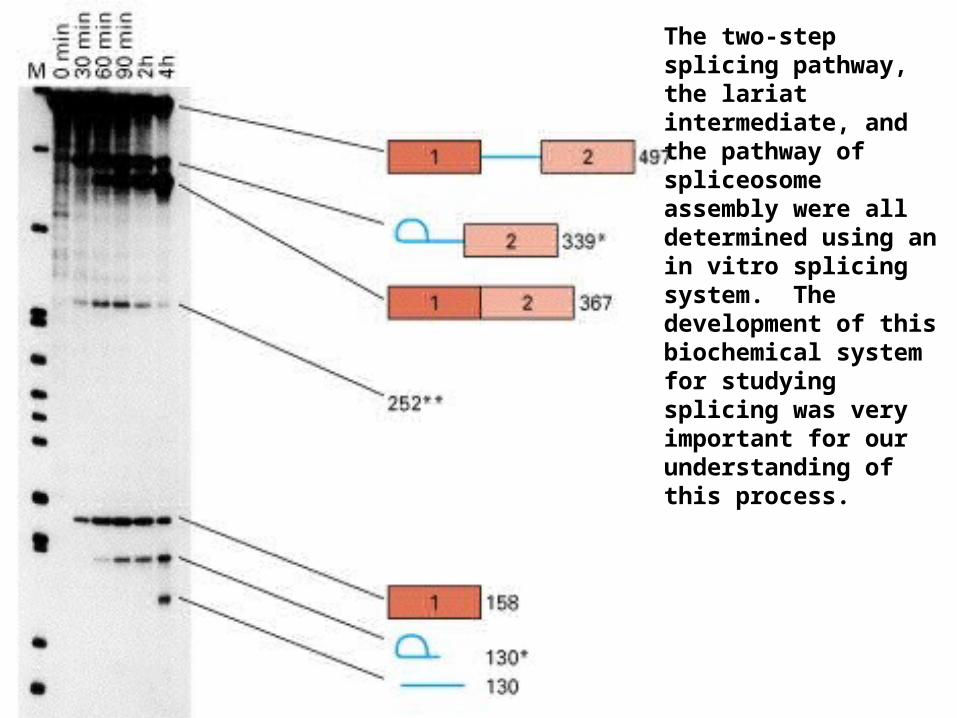
The two-step splicing pathway, the lariat intermediate, and the pathway of spliceosome assembly were all determined using an in vitro splicing system. The development of this biochemical system for studying splicing was very important for our understanding of this process.
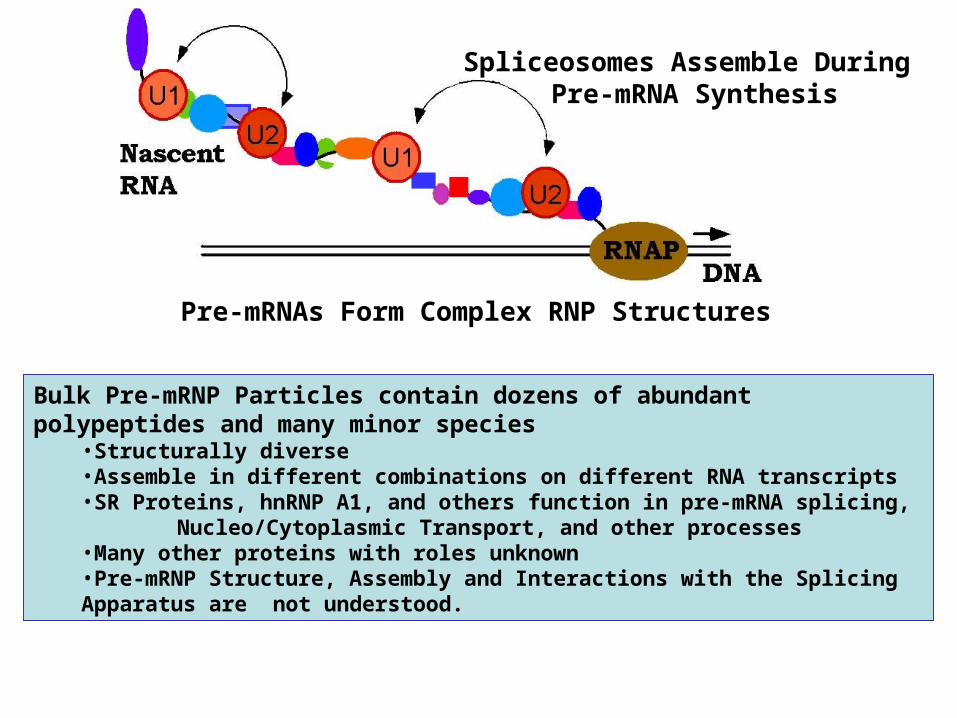
Spliceosomes Assemble During Pre-mRNA Synthesis
Pre-mRNAs Form Complex RNP Structures
Bulk Pre-mRNP Particles contain dozens of abundant polypeptides and many minor species
•Structurally diverse•Assemble in different combinations on different RNA transcripts•SR Proteins, hnRNP A1, and others function in pre-mRNA splicing, Nucleo/Cytoplasmic Transport, and other processes•Many other proteins with roles unknown•Pre-mRNP Structure, Assembly and Interactions with the Splicing Apparatus are
not understood.

Minor class AU/AC introns are spliced using alternative, low abundance snRNPs that perform the functions of U1, U2, U4 and U6. This alternative spliceosome uses the same U5 snRNP as the major class spliceosome that removes standard AG/GU introns.
5’SS 3’SSAU AC
U11, U12, U4atac, U6atac, U5
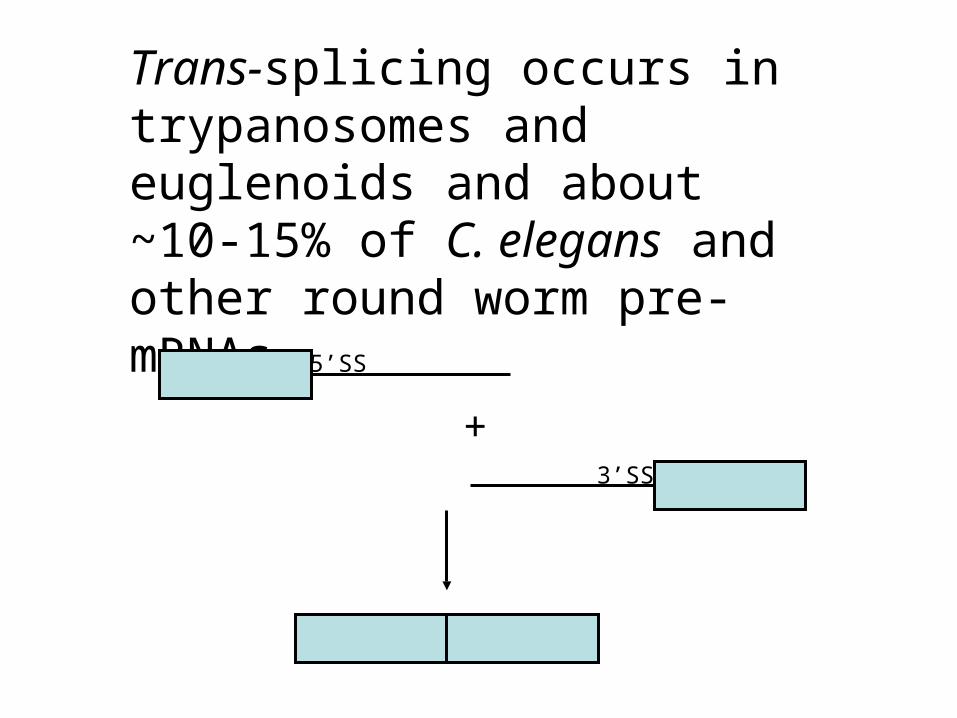
Trans-splicing occurs in trypanosomes and euglenoids and about ~10-15% of C. elegans and other round worm pre-mRNAs.
5’SS
3’SS
+
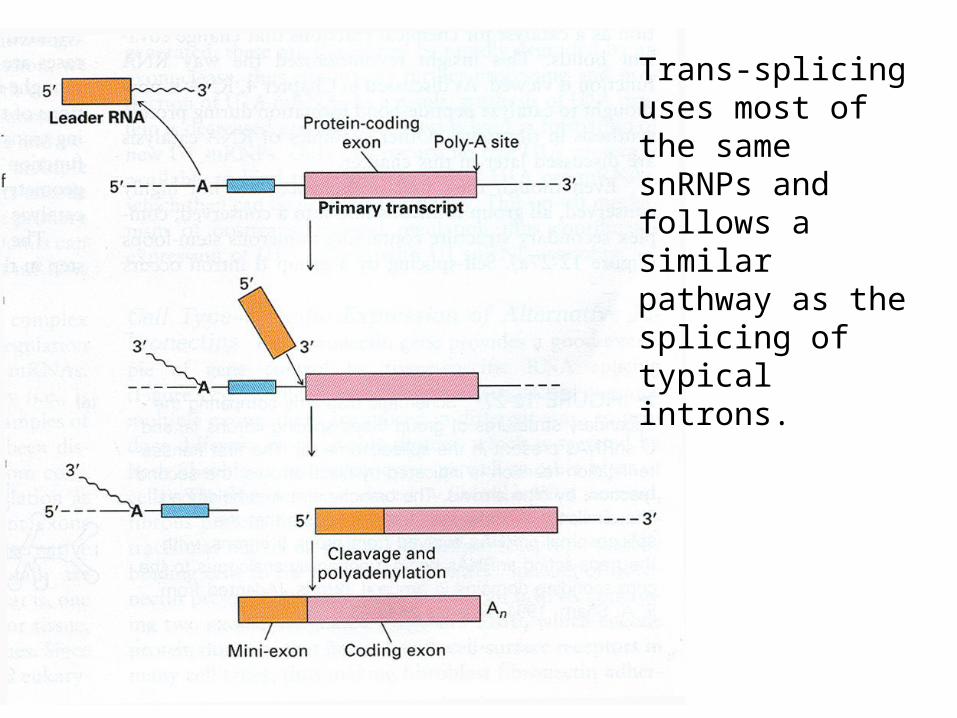
Trans-splicing uses most of the same snRNPs and follows a similar pathway as the splicing of typical introns.