Embed Size (px)
Citation preview

Postlarval Scombroid Fishes of the Genera Acanthocybium, Nealotus, andDiplospinus from the Central Pacific Ocean
DONALD w. STRASBURGl
EXCLUSIVE of the mackerels and tunas, whosecommercial importance has caused them to bestudied extensively, the early life history ofscombroid fishes is poorly known. This is particularly true of the families Gempylidae andTrichiuridae, even though they are the bases forfisheries in Australia , South Africa, Madeira, andparts of Asia. There is also a paucity of lifehistory information about the non-schoolingScombridae. This paper describes young stages ofthe scombroid Acanthocybium solandri (Cuvierand Valenciennes), the gempylid Nealotus tripesJohnson, and the trichiurid Diplospinus multistriatus Maul, all three belonging to monotypicgenera. The first has a slight commercial imp ortance (Iversen and Yoshida, 1957:370) , theothers may be considered rare species of nocommercial value.
Th e thr~~ famili es are taxonomically wellknown, the Scombri dae having been reviewedby Fraser-Brunner (1950), the Gemp ylidae byMatsubara and Iwai (1952) and Grey (1953),and the Tri chiuridae by Tucker ( 1956). All ofthese authors dealt essentia lly with adult fish.Postlarval Acanthocybium are hitherto unrecorded, the smallest specim en mentioned in theliteratur e measuring 27 cm (Kishinouye, 1923 :387). Similarly, the smallest Diplospinus is 125mm (Maul, 1948 :45) , although there is a possibility that a 69 mm Lepidopus may in realitybe a Diplospinus (Tucker, 1956 :81 ). Small N ealotus have been recorded by Liitken (188 0 :4 58),Giinther ( 1887 :3 5) , N orman (1930:35 1) , andVoss (1957 :304 ) . Liitken figured his specim ensbut regarded their identi ty as dubious. His largerspecimen (about 13 mm standard length ) has20 spines in the first dorsal .and is probably
1 Bur eau of Commercial Fisheries Biological Laboratory, Honolulu , Hawaii. Manusc ript received Novemb er 13, 1962.
N ealotus. H is smaller fish measures only about8 mm in standard length and cannot be identi fied with certainty.
All material reported upon here was collectedby "British Columbia" trawl dur ing cruise 47of the Bureau of Comm ercial Fisheries researchvessel "Hugh M. Smith ." This cruise took placein October-November 1958, in the central Pacific southeast of Hawaii. A description of thecollection method is included in Matsumoto 'sreport ( in press ) on the young tunas taken onthis cruise. Figure 1 locates the trawling stati onsand,capture sites of the species under consideration. Additional data may be found in Table 1and in the species diagnoses.
Following captu re, the fish were preserved informalin and subsequently transferred to 65 %ethyl alcohol. This particular concentrationproduced less wrinkling, hard ening, and distort ion than the customarily used 70% solut ion.Most specimens were stained with alizarin, butclearing the tissues, as with glycerin, was notdone.
Thanks are due Messrs. G. Palmer , N . B.Marshall ( British Museum ) , and G. E. Maul(Museu Municipal do Funchal ) for supplementary information on Diplospinus, and Mrs.Nancy A. Voss (University of Miami MarineLaboratory ) for data on Nealotus. The InterAmerican Tropical Tuna Commission (La Jolla,California) loaned young Scomberomorus. I amalso indebted to Tamotsu Nakata, Bureau ofCommercial Fisheries Biological Laboratory, Honolulu , for preparing the figures. .
Acanthocybium solandri (Cuvier andValenciennes)
Station 60 1 specimen : 23.7 mm
The single A canthocybium captured is rathereasily recognizable in spite of the fact that its
174
Porstlarval Scombroids from the Central Pacific-STRASBURG
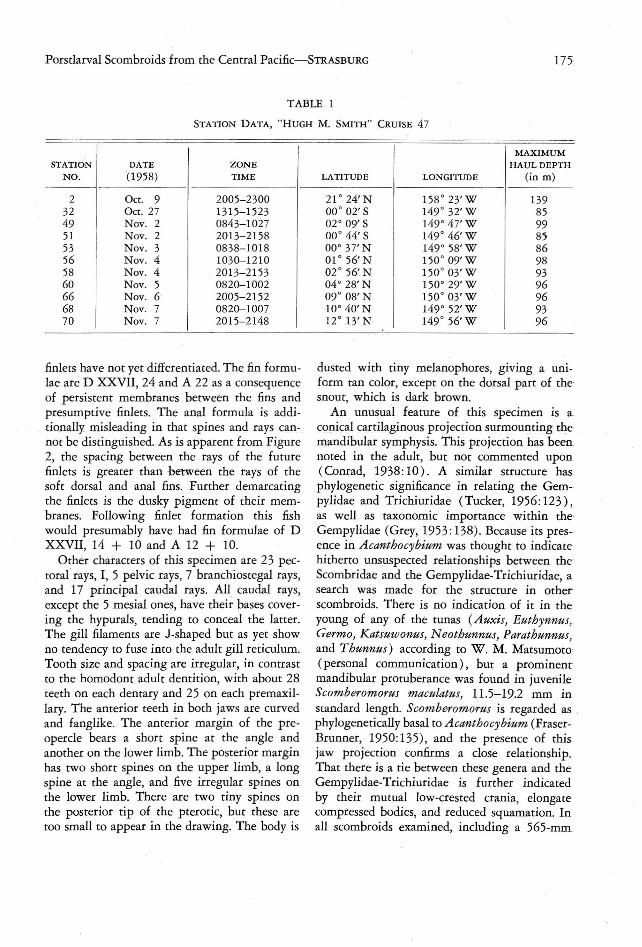
TABLE 1
STATION DATA, " H UGH M. SMITH" CRUISE 47
175
MAXIMUMSTATION DATE ZO NE HAUL DEPTH
NO. ( 1958) TIME LATITUDE LONGITUDE (i n m )
2 Oct. 9 2005- 2300 21° 24' N 158° 23' W 13932 Oct. 27 1315-1523 00 °02' S 149° 32' W 8549 N ov. 2 0843-1027 02° 09' S 149° 47' W 9951
INov. 2 2013- 2158 00° 44' S 149° 46' W 85
53 Nov. 3 0838-101 8 00° 37' N 149° 58'W 8656 I Nov. 4 1030-1210 01 ° 56' N 150 ° 09'W 98I
58I
Nov. 4 2013- 2153 02° 56' N 150 ° 03'W 9360 Nov. 5 0820- 1002 04° 28' N 150° 29' W 9666
I
Nov. 6 2005-2152 09 ° 08' N 150° 03' W 9668 Nov . 7 0820-1007 10° 40' N 149° 52' W 9370 Nov. 7 2015-2148 12° 13' N 149 ° 56'W 96
finlers have not yet differentiated. The fin formulae are D XXVII, 24 and A 22 as a consequenceof persistent membranes between the fins andpresumptive finlets. The anal formula is additionally misleading in that spines and rays cannot be distinguished. As is apparent from Figure2, the spacing between the rays of the futurefinlets is greater than b etw een the rays of thesoft dorsal and anal fins. Fur ther demarcatingthe finlers is the dusky pigment of their membranes. Following finlet formation this fishwould presumably have had fin formulae of DXXVII, 14 + 10 and A 12 + 10.
Other characters of this specimen are 23 pectoral rays, I, 5 pelvic rays, 7 branchiosregal rays,and 17 principal caudal rays. All caudal rays,except the 5 mesial ones, have their bases covering the hypurals, tending to conceal the latter.The gill filaments are J-shaped but as yet showno tendency to fuse into the adult gill reticulum.Tooth size and spacing are irregular, in contrastto the homodont adult dentition, with about 28teeth on each denrary and 25 on each premaxillary. The anterior teeth in both jaws are curvedand fanglike. The anterior margin of the pre opercle bears a short spine at the angle andanother on the lower limb. The posterior marginhas rwo short spines on the upper limb, a longspine at the angle, and five irregular spines onthe lower limb. There are two tiny spines onthe posterior tip of the pterotic, but these aretoo small to appear in the drawing. The body is
dusted with tiny melanophores, glvmg a uniform tan color, except on the dorsal part of thesnout, which is dark brow n.
An unusual feature of this specimen is aconical cartilaginous projection surmounting themandibular symphysis. This projection has beennoted in the adult, but not commented upon(Conrad, 1938 :10 ) . A similar structure hasphylogenetic significance in relating the Gempylidae and Trichiuridae (Tucker, 1956: 123),as well as taxonomic importance within theGempylidae (Grey, 1953 :138). Because its presence in A canthocybium was thought to indicatehitherto unsuspected relationships between theScombridae and the Gempylidae-Trichiuridae, asearch was made for the structure in otherscombroids . There is no indication of it in theyoung of any of the tunas (Auxis, Euthynnus,Germe, Katsuwonus, N eothunnus, Parathunnus,and Thunnus) according to W. M. Matsumoto(personal communication), but a promi nentmandi bular protuberance was found in juvenileScomberomorus maculatus, 11.5-19.2 mm instandard length. Scomberomo:'us is regarded as .phylogenetically basal to A canthocybium (FraserBrunner, 1950: 135), and the presence of thisjaw projection confirms a close relationship.That there is a tie between these genera and theGempylidae-Trichiuridae is further ind icatedby their mutual low-crested crania, elongatecompressed bodies, and reduced squamation. Inall scombroids examined, including a 565-mm

176 PACIFIC SCIENCE, Vol. XVIII, April 1964
Nealatus trip es Johnson
FIG. 1. Trawling stations on "H ugh M. Smith" cruise47, showing places of capture of Acanthocybium,Ne alotus, and Diplospinus.
Gempylus serpens, the jaw protuberance is flexible (and sometimes decurved in N esiarchusnasutus), precluding its use as a spear, and instead suggesting a hydrodynamic function (cfWisner, 1958 : 68) .
When first examined, the smaller specimenslisted above were thought to be N esiarchusnasutus Johnson, to which they bear a strongsuperficial resemblance (cf Voss, 1954). Subsequent staining revealed their salient taxonomicfeature, a reduction in the number of pelvicrays, showing that they belonged in a differentbranch of the Gempylidae, the R exea-Promethichthys-Nealatus group. According to Grey(1 953:139 ) , these genera are characterized bya reduction of the pelvic fins to a pair of spines ,or by the total absence of these fins. Only thelargest of the above specimens, a fish 176 mmin length, has the pelvies reduced to mere spinesand thus is referable to Nealatus tripes. Theremainder range from 9.0-41.5 mm, and havepelvic formulae of 1,2 or occasionally 1,1 (Table2). In this length range, as is shown in thefollowing paragraphs, the pelvic rays graduallybecome overgrown by abdominal tissues, withinwhich they are clearly visible after alizarin staining. At larger sizes the rays presumably are toodeep to be seen without dissection or staining.Norman (1930: 35:1+ . failed to find them inN. tripes specimens 53 and 88 mm long. Dissection of the pelvic region of my 176-mm fishrevealed fragments of raylike material in themusculature ventral to the pelvic bones. Thesefragments were fused to the pelvis anteriorly,and app eared to serve as anchoring struts.
In young N ealatus individuals there are twoprominent spines preceding the anal fin, whereasadults are generally regarded as having only one.My . 176-mm specimen conforms to the adultconcept externally, bur when dissected its peri anal region contained a small, flat spine, triangular in outline, embedded behind the first.Matsubara and Iwai (1952 :207) also noted asecond anal spine , bur in their 199-mm fish it isshown as being externally visible .
With an ontogenetic loss of pelvic rays andanal spines, the genus N ealatus (a nd perhapsothers) becomes more difficult to define. Characteristic counts are given in Table 2, along withdata for N esiarchus nasutus, a species of similarappearance.
Figure 3 depicts the smallest N ealatus examined, a fish 9.0 mm in standard length. At thissize the vertical fin rays have not all differentiated , the dorsal being XX-I,18 and the anal
145 I150
1 specimen: 176 mm10 specimens : 16.4-41.5 mm8 specimens: 19.6-25.8 mm
59 specimens: 10.0-20.2 mm3 specimens: 10.0-22.0 mm4 specimens: 9.0-24.0 mm6 specimens : 21.2-25.0 mm2 specimens : 21.4-28.0 mm1 specimen: 37.0 mm
155
Station 2Station 32Station 49Station 51Station 53Station 56Station 58Station 60Station 70
160
160' ISS' ISO' 145'
<J KAUAI ..l2Ao~u I
<>3::s'lAUI
20"1
~HAWAII 20'
I
I,
! I
IS' IS'
I • Acanlhocybium I
* Nealotus• Diplospinus "*70
II
i10' j 68
I l66 I
I,I
I· -*60, FANNING I. I
I *58I
1~CHRISTMAS I. *56
II I I10' *53
*3' O'I
*51
III -iT" * •• ,. . ..

Porstlarval Scombroids from the Central Pacific-STRASBURG 177
FIG. 2. Acanthocybium solandri 23.7 mm in standard length.
II-I,15, with both fins showing rudimentaryraylike structures in the membranes posteriorto the fins. The . principal caudal rays number9 -+ 8; the pectorals 13, and the pelvics 1,2.Each pelvic spine is long, rriradiate in crosssection, and spinigerous on all three edges. Theinner pelvic ray is not only much shorter thanthe outer, but is also membranously bound tothe belly anteriorly, whereas the outer ray is
.free. Like the pelvics, the dorsal spines are triradiate and spinigerous, with the degree ofspinulation decreasing posteriorly. The first twodorsal spines bear spinules on all three edges
. while the rest bear them only laterally; in addition, the number of spinules per spine decreases toward the rear of the fin. The analspines are like two spiny-edged, shore-handledspoons, attached by the bowls with the convexside forward, and the second nesting into thefirst.
The preopercle bears rwo strong denticulatespines at its angle and two shore smooth spineson its lower limb. The pteroric has two slenderspines almost perpendicular to the body surface.A row of 10 denriculations above the eye completes the head armament.
The mandible is prognathous and its tip issurmounted by a small cartilaginous pad. Themandibular symphysis is edentulous, but thereare two retrorse fangs on either side of it . Aboutseven shore upright teeth are borne on the
dentary . There is a large fang on each side ofthe premaxillary symphysis and a larger oneposterior and mesial to the first. About 13 otherteeth, some little more than denriculations, occuron the premaxillary. The palatines lack teeth,while the vomer bears a single prominent toothat each posterior tip.
Pigmentation consists of a brown streak fromthe top of the head to the rear dorsal base, plusdiffuse brown color around the dorso-anteriororbit and along the premaxillary. There is someinconsistency between specimens, probably reflecting differences in chromatophore contraction at death rather than the presence of morethan one species.
The 15.6-mm fish shown in Figure 4 differsfrom the 9-mm one principally in havingachieved the adult fin complement (D XX-I,19 + 2; A II-I, 16 + 2). The two dorsal and analfinlets are connected to each other and to thepreceding fins, being detectable only by a variation in spacing. The pelvic spines are as before,but the rays are now adnate to the belly forabout half their length. The anal spines havedeveloped a keel on their previously smooth
. anterior surfaces.The head armament consists of the two spines
at the angle of the preopercle (these now havinglost their denticulations), two shore spines onthe lower limb of the preopercle, a single perpendicular spine on the prerotic, and two pos-
178
rerior spinelike extensions of the strengtheningribs of the opercle. The supraorbital denticulations have been reduced to one or two.
Both the mandible and the upper jaw nowbear small cartilaginous symphysial pads, andthe dent ition is much advanced. Besides theanterior pair of fangs, each dentary has 11slightly retrorse teeth, and 12 similar teeth arefound along the edge of each premaxillary.Anteriorly the premaxillary has 3 or 4 largecurved fangs, the posterior ones longest, andall larger than the fangs on the dentary. Thevomer has a posterior tooth on each side, and
PACIFIC SCIENCE, Vol. XVIII, April 1964
there are about 4 slender teeth located on ornear each palatine. Those which are merelynear the palatines lack ossified bases. There areas yet no gill-rakers on the first arch .
Pigmentation is again inconsistent betweenspecimens, except for a pronounced dark sPOtbetween the first two dorsal spines. W ellmarked individuals are colored as is the 9-mmfish; others lack the dark pigmentation of thehead and dorsal fin base.
At 31.0 mm (Fig. 5 ) many of the larvalfeatures have been replaced by juvenile characters. The dorsal finlets are still interconnected
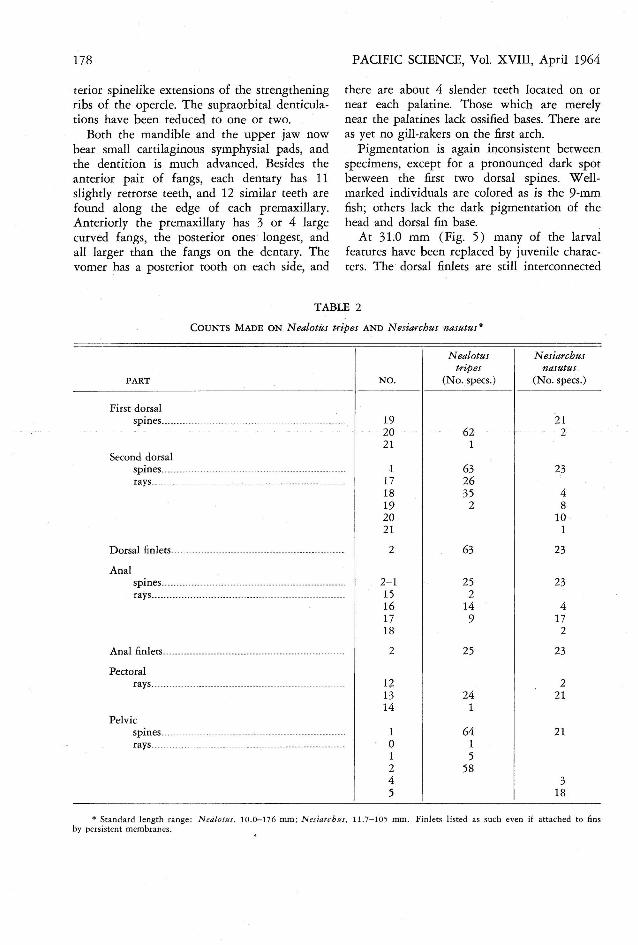
TABLE 2
COUNTS M ADE ON N ealotes tripes AND N esiarchus nasutus*
N ealotus N esiarchustripes nasutus
PART N O. (N o.specs.) (N o. specs.)
First dorsalspines.......................................... .................... 19 21
20 62 221 1
Second dorsa lspines... ....... ... ............ .............. ........ ...... ......... 1 63 23rays...... ......................................... ..... .............. 17 26
18 35 419 2 820 1021 1
D orsal finle ts.... ... ... ...... .. ... ............. ............. ........ ... 2 63 23
Analspines...... ..... ........................ ......... .................. 2- 1 25 23rays................ ..... .... ....... ................... ......... ... .. 15 2
16 14 417 9 1718 2
Anal finlets........ .......... ............... ...... ......... .......... ... 2 25 23
Pectoralrays.................... ... ....................... ... ........ ... ..... 12 2
13 24 2114 1
Pelvicspines ....... ....................... ....._......... ................ . 1 64 21rays........................... ...................................... 0 1
1 52 584 35 18
• Standard lengt h ra nge: Ne alotus , 10.0-176 mm ; N esiarcbes , 11. 7-1 ·05 mm . Finlets listed as such even if attached to finsby persistent membranes.

Porstlarval Scombroids from the Central Pacific-STRASBURG 179
FIG. 3. Nealotus tripes 9.0 mm in standard lengrh.
but are free from the last dorsal ray. The sameis true of the anal finlets, except that there is apersistent basal membrane between the last analray and the first finlet. The dorsal spines havelost the spinules from their anterior edges, andthe lateral spinules are less prominent on allfin spines, the space between the spinules havingbecome filled with bony material. The pelvicrays are both buried in epidermal tissues, withthe short mesial ray lying deeper than the longlateral one.
The upper spine at the preopercular angle isprominent, but the lower is broad and scarcelynoticeable. The lower preopercular limb issmooth, and the opercular spines are reduced insize. The cartilaginous protuberance at the tipof each jaw is conical and well developed. Thedentary has 2 anterior fangs and 13 variablysmaller teeth; the premaxillary bears 3 anteriorfangs and about 16 smaller teeth. The vomerhas 2 teeth on each posterior tip, and there areabout 6 teeth on or near each palatine. Asbefore, some of the latter are attached to unossified material on the roof of the mouth.There are 3 + 1 + 8 gill-rakers on the firstarch, the one at the angle being much largerthan the others.
A new feature is the lateral line, a few scalesof which are visible on the 31-mm fish. Theselie in the skin and are better developed andmore numerous anteriorly. In a 35-mm specimen (not shown) the lateral line is nearlycompletely developed . It is a tubelike structure
contaInIng 78 scales and extending from thehead to the caudal peduncle. Scale spacing isoccasionally irregular, suggesting incomplete development or loss through damage.
Pigmentation consists of rows of tiny, closeset, dark dots, giving an over-all tan color. Thisis intensified on top of the head, at the jawtips, along the dorsal base, on the sides of thecaudal peduncle, and around the anal spines. Thefins lack pigment except for a distal dark SPOt 'between the first three dorsal spines.
A 41.5-mm fish (Fig. 6) is essentially ajuvenile. Persistent larval features include theslightly denticulate fin spines, the absence ofscales except in the lateral line, and basal membranes between the finlets. The pelvic rays areembedded, the two preopercular and tWO opercular spines are reduced in size, and the symphysial jaw pads are less prominent (and nowdecurved in the case of the upper jaw). Theteeth have increased slightly in number, therenow being 3 anterior fangs and 19 lateralteeth on the premaxillary, 2 fangs and 16 lateralteeth on the dentary, 8 or 9 teeth on each palatine(all firmly ankylosed to the palatine and nonefree on the roof of the mouth) , and 1 or 2elongate teeth posteriorly on each side of thevomer . There are 94 scales in the lateral linebut none elsewhere . There are 3 + 1 + 8slender gill-rakers on the first arch. Pigmentationis as in the 31-mm fish, except that there arenow indications of distal dark spots betweenthe first four dorsal spines.

180 PACIFIC SCIENCE, Vol. XVIII, Apr il 1964
Diplospinus multistriatus Maul
The genus Diplospinus is known from Madeira, the West Indies , and the Phil ippine-EastIndi an. region (Tucker, 1956:79 ). The abovespecimens are the first from the central PacificOcean. Postlarvae are hitherto unknown, although they and earlier stages have been foundfor other rrichiurids (Delsman, 1927; Padoa,
It is unnecessary to figure my 176-mm fish,for it has essentially the same featur es as the199-mm specimen shown by Matsubara andIwai (1952:208) . As noted earlier, it differs inhaving the second anal spine embedded. Otherdata from my specimen are as follows. Thelower jaw has a rudimentary symphysial pad ,1 or 2 anterior fangs on each side, and a row of17 lateral teeth set closely together like the teethof a saw. On the pr emaxillary are 3 large ant erior fangs and a row of 21 closely-spaced lateralteeth. :The symphysial pad of the premaxillaryis blunt, decurved, and scarcely noticeable. Theroof of the mouth is overgrown by membraneswhich, when removed , reveal an edentulousvomer. There are 8 shan, blunt denticles oneach palatine. The gill-rakers consist of numerous spinules so variable in size, position, andspacing that they cannot be enumerated (cfMatsubara and Iwai, 1952 :209) . The specimenis partially skinned and the lateral line scalescannot be counted.
Station 66Station 68Station 70
2 specimens: 103 and 166 mm10 specimens: 10.0-17.4 mm1 specimen: 156 mm
1956: 508- 513; Regan , 1916 :144-145 ) . Thefollowing descriptions and figures illustrate development in which a short, high-bodied postlarva becomes an increasingly att enuate juvenile.
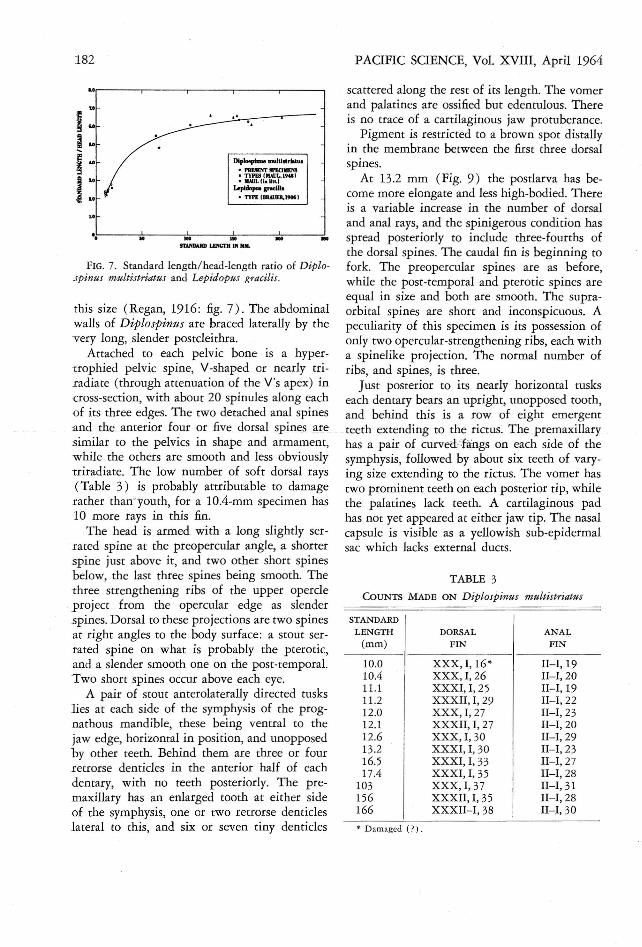
As pointed out by Tucker (1 956:81) , thereis a possibility that Diplospinus multistriatusMaul is a synonym of Lepidopus gracilis Brauer.Th e latter was described from a specimen 69mm in length (Brauer, 1906:291), which differsfrom multistriatus chiefly in characters whichchange with growth. Although it was imp ossibleto examine the type of gracilis, a brief study wasmade of a feature thought to distingu ish it frommultistriatus, the ratio of stand ard length to
head length. This ratio is plotted against standard length in Figure 7. The data repre sent thepresent specimens, the type series of multistriatus as described by Maul (1948 :45), a recentlycollected 65-mm multistriatus with a SL/HLratio of 5.5 (Maul, in Iirr. ) , and the type ofgracilis. The curve was derived from the regression of head length on standard length, Y =3.397 +0.1350X, calculated from the abovemultistriatns.
From Figure 7 it would seem that multistriatus and gracilis are separate species. This is confirmed by the fiducial limits of the head lengthof multistriatus 69 mm in standard length ( thelength of Brauer's gracilis). At this size one can
expect a head-length range of 12.3- 13.1 mm,using p.1l1 values of t and the formula
f'l1 ..~ x -
SY= - + -S"n x-
(Snedecor, 1946 : 120 ) . The 14.25-mm head
...F IG. 4. Nealotus tripes 15.6 mm in standard length.

Porstlarval Scombroids from the Central Pacific-S1RAsBURG 181
FIG. 5. Nealotus tripes 31.0 mm in standard lengrh.
length of gracilis lies significantly distant fromthis range, and the two . forms are thereforeconsidered distinct species.
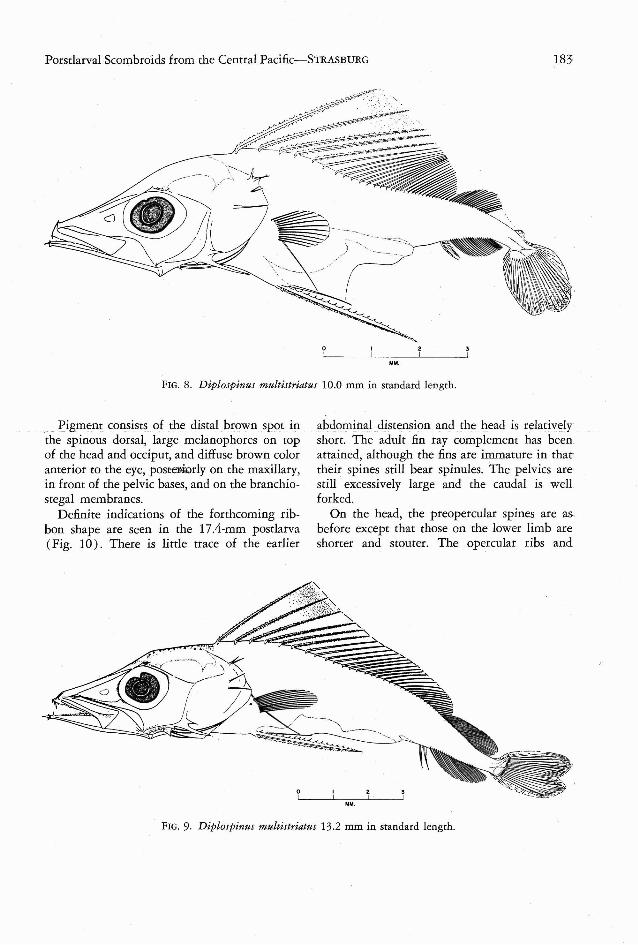
Because not all postlarvae in the 10.0-17.4mm length range have acquired the adult finray complement, it is necessary to list meristicand length data conjointly in Table 3. Thenumber of dorsal and anal fin rays increases irregularly with size. The pectoral, pelvic, andcaudal fins and the hranchiosregal rays showlittle or no meristic variation. Except for onespecimen with 13 rays on one side and 12 onthe other, the pectoral was constant at 12 rays.There were 7 branchiostegal rays and 9 + 8principal caudal rays, the latter being difficultto count because of damage or nonattainmentof definitive shape . In all 13 specimens thepelvic fins are represented by a pair of sroutdenticulated spines, in agreement with Maul'sfindings (1948:42) but not with Tucker's (1956:79-81). The latter, working with a "completely
skinned" paratype, observed "Ventral fin I-I;a narrow scale-like spine and an external splitray twice as long." Palmer and Marshall (inlitr.)have re-examined this para type but failed tofind the external split ray, observing only thescale-like spine and the pelvic bones. I suggestthat Tucker's "ray" was in reality the splintlikepelvis, perhaps loosened in its muscular bed bythe force which skinned the fish. In my specimens the attenuate pelvis bears a marked resemblance to a soft ray and is, coincidentally,just twice as long as the pelvic spine.
The most distinctive feature of the 1O.0-mmpostlarval Diplospinus is its relatively greatdepth at mid-length. As shown in Ftgure 8; thisis caused by a ventral distension of the belly bythe elongate pelvic bones, a condition reminiscent of the Balistidae and Monacanthidae, exceptthat in Diplospinus the pelvis extends posteriorly. The great depth of Diplospinus is quitedifferent from the narrow belly of Lepidopus at
MM.
FIG. 6. Nealotus tripes 41.5 mm in srandard length . The eyes are lacking in this specimen, but have beendrawn as rhey appear in another of abour the same size.
","
• D am aged ( ? ) .
TABLE 3
COUNTS MADE ON Diplospines multistriatus
PACIFIC SCIENCE, Vol. XVIII, April 1964
scattered along the rest of its length. The vomerand palatines are ossified but edentulous. Thereis no trace of a cartilaginous jaw protuberance.
Pigment is restricted to a brown spot distallyin the membrane between the first three dorsalspines.
At 13.2 mm (Fig. 9 ) the postlarva has become more elongate and less high -bodied. Thereis a variable increase in the number of dorsaland anal rays, and the spinigerous condition hasspread posteriorly to include three-fourths ofthe dorsal spin es. Th e caudal fin is beginning tofork. The preopercular spines are as before,while the post-temporal and prerotic spines areequal in size and both are smooth . The supraorbital spines are short and inconspicuous. Apeculiarity of this specimen is its possession ofonly two opercular-strengthening ribs, each witha spinelike projection. The normal numb er ofribs, and spines , is three .
Just posterior to its nearly horizontal tuskseach dentary bears an upright, unopp osed tooth,and behind this is a row of eight emergentteeth extending to the rictus . The premaxillaryhas a pair of curved-fangs on each side of thesymphysis, followed by about six teeth of varying size extending to the rictus. Th e vomer hastwo prominent teeth on each posterior tip, whilethe palatines lack teeth . A cartilaginous padhas not yet appeared at either jaw tip . The nasalcapsule is visible as a yellowish sub-epidermalsac which lacks external ducts.
STANDARD
LEN GTH DORSAL ANAL
(mm) FIN FIN
10.0 XX X, 1,16 * I1-I,1910.4 XXX, I, 26 I1-I,2011.1 XXXI ,I, 25 I1-I ,1911.2 XXXII, I, 29 I1-I, 2212.0 XXX, I, 27 I1-I, 2312.1 XXXII, I, 27 II-I, 2012.6 XXX, I, 30 II-I,2913.2 XXXI, I, 30 II-I, 2316.5 XXXI, I, 33 I1-I, 2717.4 XXX I, I, 35 II-I, 28
103 XXX, I, 37 I1-I , 31156 XXXII, I, 35 I1-I,28166 XXXII-I, 38 II-I, 30
......
Dip_piau. mulll.....IUl
• I'Ilf.SfNr SPEClMIM• TYPESI MAllL, 19181• MAUL II. IIllo)
Lopl""'" _111.· nn fBIUI.JER.I9061
... ".
IlA.l'mUD l.mCTB IN Mal.
..~---;----.;.---;;-----.:.---.:
..,,.---,-- - ----,- - -.,---...,.- - ---,
u
182
FIG. 7. Standard length /head-length ratio of Diplospinas multistriatus and Lepidopus gracilis.
this size (Regan, 1916 : fig. 7) . The abdominalwalls of Diplospinus are braced latera lly by thevery long, slender postcleirhra.
Attached to each pelvic bone is a hypertrophied pelvic spine, V-shaped or nearly tri radiate (through attenuation of the V's apex) incross-section, with about 20 spinules along eachof its three edges. The two detached anal spinesa nd the anterior four or five dorsal spines aresimilar to the pelvics in shape and armament,whil e the others are smooth and less obviouslytriradiate. The low number of soft dorsal rays( Table 3) is probably attributable to damagerather thanyouth, for a 10A-mm specimen has10 more rays in this fin.
The head is armed with a long slightly serrated spine at the preopercular angle, a shortersp ine just above it, and two other short spinesbelow, the last three spines being smooth. Thethree strengthening ribs of the upper opercle
. project from the opercu lar edge as slender.spines. Dorsal to these projections are two spinesat right angles to the body surface : a stout serrated spine on what is probably the pterotic,and a slender smooth one on the post-temporal.Two short spines occur above each eye.
A pair of stout ante rolaterally directed tuskslies at each side of the symphysis of the prognathous mandible, these being ventral to thejaw edge, horizontal in position, and unopposedby other teeth. Behind them are three or fourretrorse denticles in the anterior half of eachdentary, with no teeth posteriorly. The pre maxillary has an enlarg ed tooth at eith er sideof the symphysis, one or two retrors e denticleslateral to this, and six or seven tiny denticles
Porstlarval Scombroids from the Central Pacific-STRASBURG 183
oI
MM,
FIG. 8. Diplospinus multistriatus 10.0 mm in standard length.
. Pigment consists of the distal brown spot inthe spinous dorsal, large melanophores on topof the head and occiput, and diffuse brown coloranterior to the eye, poseerserly on the maxillary,in front of the pelvic bases, and on the branchiostegal membranes.
Definite indications of the forthcoming ribbon shape are seen in the 17.4-mm postlarva(Fig. 10) . There is little trace of the earlier
abdominal distension and the head is relativelyshort. The adult fin ray complement has beenattained, although the fins are immature in thattheir spines still bear spinules . The pelvics arestill excessively large and the caudal is wellforked.
On the head, the preopercular spines are asbefore except that those on the lower limb areshorter and stouter. The opercular ribs and
MM.
FIG. 9. Diplospinus multistriatus 13.2 mm in standard length.

184 PACIFIC SCIENCE, Vol. XVIII, April 1964
FIG. 10. Diplospinus multistriatus 17.4 mm in standard length.
spines and the post-temporal and pterotic spinesare likewise little changed. A single low spine,actually little more than a crest, occurs aboveeach eye. The upper of the two tusks on thedenrary is much the larger, and the upright toothposterior to the tusks equals it in size. About15 curved teeth, some occurring as pairs, occupythe length of the dentary. Anteriorly the premaxillary has 3 or 4 greatly enlarged fangs, theanterior 1 or 2 about half the size of the others .Behind the fangs is a row of about 10 shortteeth similar to those of the dentary . The vomerstill has 2 teeth near each posterior tip, andthese are attached on the inner surface of thebone rather than to its crown. Each palatinebears 6-8 short denticles. There are 3+ 1 + 11long, slender gill-rakers on the first arch. Thenasal capsule has 2 openings, an oval anteriornostril and an irregular posterior one. The nostrils must form suddenly, for they are not present in a 16.5-mm specimen. Small patches ofcartilage surmount the tips of both jaws, wherethey appear to do nothing more than smooth.over irregularities in the bones.
Large melanophores occur on top of the head,but give way to smaller ones which extend in anarrow line from the occiput along the entirebase of the spinous dorsal. Dark pigment overlies the nasal capsule and occurs as a ring aroundthe orbit. The maxillary, posterior denrary, andpre-pelvic and branchiostegal regions are alsodusky, and there is a dark spot in the spinousdorsal fin membranes. (These membranes aredamaged in the 17.4-mm fish and the spot isdrawn as it appears in a 16.5-mm specimen.)
REFERENCES
BRAUER, AUGUST. 1906. Die Tiefsee-Fische.Wiss . Ergebn. "Valdivia" 15 (1) :1-432, 18pIs., 176 figs.
CONRAD, G. MILES. 1938. The osteologyandrelationships of the wahoo (Acanthocybiumsolandri), a scombroid fish. Amer . Mus. Novirates (1000) :1-32,9 figs.
DELSMAN, H. C. 1927. Fish eggs and larvaefrom the Java Sea, 11. The genus Trichiurus.Treubia 9(4) :338- 35r , 19 figs.
FRASER-BRUNNER, A. 1950. The fishes of thefamily Scombridae. Ann. Mag. Nat. Hist. XII,3(26) :131-163,35 figs.
GREY, MARION. 1953. Fishes of the familyGempylidae, with records of Nesiarchus andEpinnula from the western Atlantic and descriptions of two new subspecies of Epinnulaorientalis. Copeia 3:135-141.
GUNTHER, A. 1887. Report on the deep-seafishes collected by H.M.S. Challenger duringthe years 1873-1876. Rept. Sci. Res. Voy.H.M.S. Challenger, Zool. 22 (57) : lxv + 335,73 pIs., 7 figs.
IVERSEN, EDWIN S., and HOWARD O. YOSHIDA.1957. Notes on the biology of the wahoo inthe Line Islands. Pacific Sci. 11 (4) :370-379,7 figs., 4 tabs.

Porstlarval Scombroids from the Central Pacific-STRASBURG . 185
KISHINOUYE, KAMAKICHI. 1923. Cont ributionsto the comparative study of the so-calledscombroid fishes. Jour. Coll, Agric. Imp. Univ.Tokyo 8 (3 ) :293-475, 26 figs.
LUTKEN, C. 1880. Spolia Atlant ica: K. Dansk.Selsk. Skrifr, V, 12 (6) :409- 613, 5 pis., 11figs., 1 tab.
MATSUBARA, KIYOMATSU, and TAMOTSU IWAI.1952. Studies on some Japanese fishes of thefamily Gempylidae. Pacific Sci. 6 (3) :193212, 12 figs.
MATSUMOTO, WALTER M. In press. Collectionand descriptions of juvenile tunas from thecentra l Pacific. Deep-Sea Res.
MAUL, G. E. 1948. Quarro peixes novos dosmares da Madeira. Bol. Mus. Munic. Funchal(3 ) :4 1-55,4 figs., 4 tabs.
N ORMAN, J. R. 1930. Oceanic fishes and flatfishes collected in 1925-1927. DiscoveryRepts. 2:261-:370, 1 pl., 47 figs.
PADOA, EMANUELE. 1956. Uova, larve e stadigiovanili di Teleostei, Divisione: Scombriformes. Fauna Flora Golfo N apoli 38 (3 ) :471- 547, 90 figs., 7 tabs.
REGAN, C. T. 1916. Larval and post-larval fishes.Brit. Antarctic ("Terra Nova") Exped., 1910,Zool. 1: 125-156, 10 pls., 5 figs.
SNEDECOR, GEORGE W. 1946. Statistical Methods. 4th ed. Iowa State College Press, Ames,Iowa. xvi + 485 pp.
TUCKER, DENYS W . 1956. Stud i es on thetrichiuro id fishes, 3. A pr eliminary revision ofthe family Trichiuridae. Bull. Brit . Mus. N at.Hist., Zool. 4 (3) :73- 130, 1 pl., 23 figs., 6tabs.
VOSS, NANCY A. 1954. The postlarval development of the fishes of the family Gempylidaefrom the Florida Current, I. Nesiarchus John son and Gemp ylus Cuv. and Val. Bull. Mar.
, Sci. Gulf Carib. 4 (2 ) :120-159, 15 figs., 9tabs.
--- 1957. Fishes of the family Gemp ylidaecollected by the Bermuda Oceanographic Expeditions 1929, 1930, 1931 and 1934. Copeia4 :304-305.
WISNER, ROBERT 1. 1958. Is the spear of istiophorid fishes used in feeding ? Pacific Sci.12 (l ) :60-70, 3 figs.





























