Embed Size (px)
Citation preview

Phenotypic plasticity of pigmentation andmorphometric traits in Pnigalio soemius
(Hymenoptera: Eulophidae)
U. Bernardo1*, P.A. Pedata1 and G. Viggiani2
1CNR – Istituto per la Protezione delle Piante – Sezione di Portici,Via Universita 133, 80055 Portici, Italy: 2Dipartimento di Entomologia
e Zoologia Agraria, Universita degli Studi di Napoli ‘Federico II’,Via Universita 100, 80055 Portici, Italy
Abstract
Species of the genus Pnigalio Schrank are ectoparasitoids on several pest insects.Most species are polyphagous parasitoids of lepidopteran and dipteran leafminers.Despite their potential economic importance, information on intraspecific pheno-typic variability is insufficient. Pnigalio soemius (Walker) was reared at fivedifferent temperatures (10, 15, 20, 25, 30�C) on mature larvae of one of itsnatural hosts, Cosmopterix pulchrimella Chambers (Lepidoptera: Cosmopterigidae),to investigate the influence of temperature on size, colour and other morphologicaltraits, and to measure the range of variation of several characters. Thermaldevelopmental reaction norms, which represent the effect of temperature duringgrowth and development on the value of some adult traits, were produced. Theresults confirmed the influence of temperature on numerous characters and thatthese characters had a larger range of variation than realized previously in theconstruction of taxonomic keys to species. In particular, the number and positionof the costulae on the propodeum and colour of the gaster were affected by rearingtemperature.
Keywords: developmental reaction norm, gastral pigmentation, morphology,Pnigalio soemius, size variation, taxonomy
Introduction
Taxonomy of the Chalcidoidea (Hymenoptera) is almostexclusively based on the comparison of morphological traits,but these can be related to fitness and thus are a possibletarget of natural selection. As a consequence, morphologicaldifferences are shaped by both speciation divergence andecological adaptations. In particular, expression of numerousmorphological traits may be strongly influenced by thelarval environment and, especially, by developmentaltemperature (Gibert et al., 2004). Thus, errors are possiblewhen species are differentiated based on variable morpho-logical traits whose real variance is unknown. Morphological
and biological diversity is well known in the Chalcidoidea,and their classification is complicated by variation insize and morphology as a function of the host, seasonaldimorphism and dichroism (Askew, 1971).
In the Chalcidoidea, Pnigalio Schrank (Eulophidae) is ataxonomically problematic genus (Barrett et al., 1988).Taxonomists dealing with this genus have often useddifferences in colour, propodeal morphology and positionof the costulae to distinguish species, but have only rarelyassessed intraspecific variation. Moreover, numerous specieshave been described from only a few individuals froma restricted geographic area. High levels of phenotypicplasticity in Pnigalio have already been reported for severaltraits of P. minio (Walker) (as P. flavipes (Ashmead)) (Barrettet al., 1988). It is therefore important to determine thepotential magnitude of intraspecific variation in these traitsbefore they are used for species recognition.
*Fax: +39 081 7758122E-mail: [email protected]
Bulletin of Entomological Research (2007) 97, 101–109 DOI: 10.1017/S0007485307004816

The understanding of intraspecific variation in Pnigaliocan be facilitated by investigating a specific system ofplant–phytophage–parasitoid. Hitherto, these data have notbeen available, partly because of the difficulties in rearingspecies of this genus. For the present study we employedParietaria diffusa M. & K. (Urticales: Urticaceae), Cosmopterixpulchrimella Chambers (Lepidoptera: Cosmopterigidae), andPnigalio soemius (Walker) (Hymenoptera: Eulophidae).Pnigalio soemius is a polyphagous ectoparasitoid of lateinstar leafminer larvae belonging to different orders ofinsects, and of gall-forming sawflies.
In this study we evaluated the influence of rearingtemperature on several quantitative traits using develop-mental reaction norms (DRN). A DRN is the response curveof a phenotype as a function of an environmental gradient,using a character state approach (Via et al., 1995).
Materials and methods
Rearing
A culture of C. pulchrimella was maintained in anenvironmental chamber at 25+1�C, 60%+10% r.h. and12L:12D on P. diffusa. Plants with a height of about 15 cmwere collected in the field, and each placed in a pot of10 cm diameter that was then covered by a plastic isolator.After a quarantine period in a greenhouse, the plants wereexposed to about 20 adults of the leafminer by introducingthem through the isolator. The isolator was cylindrical,with a diameter of 10.5 cm and a height of 19 cm, andwas closed by a net on the top. After 24 h, the adults werecollected to infest other plants. After reaching maturity in14 days, the leafminer larvae were exposed to the para-sitoids.
A culture of P. soemius was established using adultsthat emerged from parasitized larvae of C. pulchrimellacollected in Parco Gussone, Portici (Na, Italy), in April, 2001.Identification of this parasitoid population as P. soemius wasmade by comparison with specimens from The NaturalHistory Museum, London, that had been identified asP. soemius by Dr Z. Boucek. However, the specific limits ofP. soemius have to be better defined because high variabilityin colour and other morphological traits, correlated withtemperature and host species differences, have been demon-strated in this genus (Viggiani, 1963; Askew, 1971; Barrettet al., 1988; Ateyyat, 2002; Bernardo et al., 2005).
Three pairs of parasitoids were offered a plant ofP. diffusa with full grown leafminer larvae for 24 h. Parasitoidfemales were at least 72 h-old and were allowed to feed onhoney and to host-feed (Bernardo et al., 2006).
Tests
Influence of temperature on gastral tergite pigmentationof P. soemius females was assessed at five temperatures(10�, 15�, 20�, 25�, 30�+1�C), whereas influence on size andother traits was evaluated at four temperatures (15�, 20�, 25�,30�+1�C). Parasitoid pairs (10 replicas of 3 pairs) wereoffered for five consecutive days a single infested plant ateach temperature, thus testing each insect at all the testedtemperatures.
Six days after rearing at 20–30�C, and 15 days at 10–15�C,leaves with leafminer larvae were collected and put inaerated boxes (65 mm diameter and 85 mm high). Newly
emerged parasitoid adults were counted, isolated and sexed.Colour comparison was performed on a reduced data set of15 females for each temperature, except for 10�C becauseonly 11 females were obtained at this temperature. Formorphological comparison, 50 newly emerged females wereisolated in glass vials (5.5r1.2 cm). After death, specimenswere mounted on card rectangles and used to measure thefollowing features and ratios: length of metatibia, a widelyused comparative measure of body size (Bezemer et al.,2005; Chow & Heinz, 2005), length of clava/first segment offunicle, length/width of fourth segment of funicle, length offore wing, length of marginal and stigmal veins, lengthof marginal vein/stigmal vein, length/width of propodeum,width of median carina near anterior margin of propodeum,insertion of costulae on median carina/length of pro-podeum, and number of costulae. The number of costulaewas established by giving an integer value to each costulathat reached the lateral margin of the propodeum (plicae),and a decimal value (0.25; 0.5; 0.75) given for each costulathat did not, based on the distance it was separated from theplicae or median carina; width of the median carina atthe anterior margin was ascribed to one of four qualitativeclasses we defined for our sample (fig. 1). The length/widthratio of the gaster is sometimes considered an importanttaxonomic character, but we did not take these measure-ments because the gaster quite often is collapsed in deadspecimens and its use is therefore impractical. Gastralpigmentation was evaluated within 24 h after death. The
Fig. 1. Propodeum of female Pnigalio soemius, illustrating widthrange of median carina at the anterior margin of propodeum.A, straight base (class 1); B, slightly widened base with smalltriangular inner cell (class 2); C, intermediate widened base withmedium triangular inner cell (class 3); D, widely enlargedbase with large triangular inner cell (class 4). c, costula; mc,median carina; p, plica; w1, w2, w3, w4, anterior base of mediancarina.
102 U. Bernardo et al.
percentage of black on the gastral tergites was calculatedusing the PDSview program.
Voucher specimens are deposited in the insect collectionof the Dipartimento di Entomologia e Zoologia Agraria‘Filippo Silvestri’, Universita degli Studi di Napoli‘Federico II’.
Statistical analysis
Data satisfying conditions of normality and homoscedas-ticity, both untransformed or after appropriate transforma-tion, were analysed by ANOVA and the means wereseparated at the 0.05 level of significance by a multiplerange test (Tukey HSD). In all other cases a non-parametrictest (Kruskal-Wallis) was used and medians were graphi-cally separated by a Box-and-Whisker plot analysis (Stat-graphics plus, 1997).
All data are presented non-transformed with theirstandard error within brackets.
Results
Pigmentation of gastral tergites
For gastral tergite pigmentation, a convex reaction normwas observed. The percentage of black pigmentation rangedbetween 5 and 100%, with all females reared at 10�Chaving the tergites completely black, and with a minimumof pigmentation around 25�C (Kruskal-Wallis test, P< 0.01;
df = 4) (fig. 2). At 10 and 15�C, the legs also showed a largerproportion of dark colour, although this trait was notevaluated quantitatively.
Size
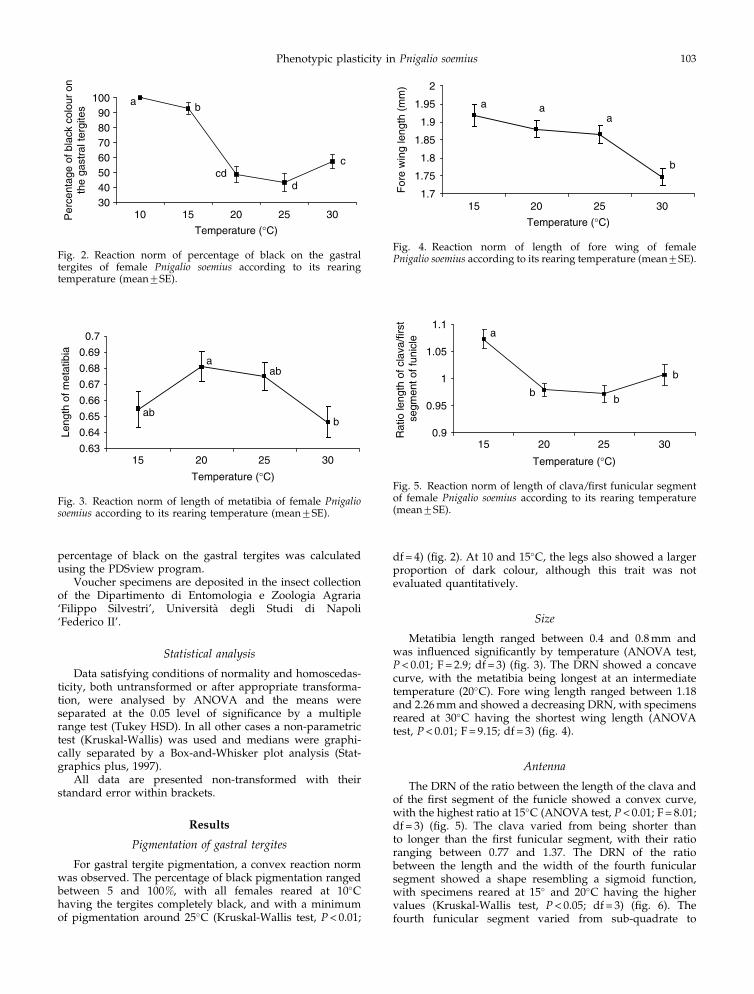
Metatibia length ranged between 0.4 and 0.8 mm andwas influenced significantly by temperature (ANOVA test,P< 0.01; F = 2.9; df = 3) (fig. 3). The DRN showed a concavecurve, with the metatibia being longest at an intermediatetemperature (20�C). Fore wing length ranged between 1.18and 2.26 mm and showed a decreasing DRN, with specimensreared at 30�C having the shortest wing length (ANOVAtest, P< 0.01; F = 9.15; df = 3) (fig. 4).
Antenna
The DRN of the ratio between the length of the clava andof the first segment of the funicle showed a convex curve,with the highest ratio at 15�C (ANOVA test, P< 0.01; F = 8.01;df = 3) (fig. 5). The clava varied from being shorter thanto longer than the first funicular segment, with their ratioranging between 0.77 and 1.37. The DRN of the ratiobetween the length and the width of the fourth funicularsegment showed a shape resembling a sigmoid function,with specimens reared at 15� and 20�C having the highervalues (Kruskal-Wallis test, P< 0.05; df = 3) (fig. 6). Thefourth funicular segment varied from sub-quadrate to
30405060708090
100 a b
cdd
c
10 15 20 25 30
Temperature (°C)
Per
cent
age
of b
lack
col
our
onth
e ga
stra
l ter
gite
s
Fig. 2. Reaction norm of percentage of black on the gastraltergites of female Pnigalio soemius according to its rearingtemperature (mean+SE).
0.63
0.64
0.65
0.66
0.67
0.68
0.69
0.7
15
ab
aab
b
20 25 30
Temperature (°C)
Leng
th o
f met
atib
ia
Fig. 3. Reaction norm of length of metatibia of female Pnigaliosoemius according to its rearing temperature (mean+SE).
1.7
1.75
1.8
1.85
1.9
1.95
2
15 20 25 30
Temperature (°C)
For
e w
ing
leng
th (
mm
)
a aa
b
Fig. 4. Reaction norm of length of fore wing of femalePnigalio soemius according to its rearing temperature (mean+SE).
0.9
0.95
1
1.05
1.1
15
a
bb
b
20 25 30
Temperature (°C)
Rat
io le
ngth
of c
lava
/firs
t se
gmen
t of f
unic
le
Fig. 5. Reaction norm of length of clava/first funicular segmentof female Pnigalio soemius according to its rearing temperature(mean+SE).
Phenotypic plasticity in Pnigalio soemius 103

rectangular, with the length:width range varying between1.12 and 2.33.
Fore wing venation
The marginal and stigmal veins showed different DRNtrends. The marginal vein had a concave curve, with itsgreatest length at intermediate temperatures (ANOVA test,P< 0.01; F = 5.42; df = 3) (fig. 7), whereas length of the stigmalvein was inversely proportional to the rearing temperature(ANOVA test, P< 0.01; F = 19.53; df = 3) (fig. 8). The ratiobetween the two measures increased with temperature(ANOVA test, P< 0.01; F = 24.1; df = 3) (fig. 9). Minimumand maximum values of this ratio were 3.05 and 5.58;marginal and stigmal vein length ranged between 0.40 to0.77 mm and 0.10 to 0.20 mm, respectively.
Propodeum, median carina and costulae
Females reared at 15�C had the highest average value ofthe length:width ratio of the propodeum (Kruskal-Wallistest, P< 0.01; df = 3) (fig. 10), with the range of variationbeing between 0.77 and 1.67. The mean number of costulaeincreased when rearing temperature increased (Kruskal-Wallis test, P< 0.01; df = 3) (fig. 11). Even though correlationbetween the size of a female and the number of costulae wasweak (Y= 0.611+0.022 X, F = 6.61, P< 0.05, R2 = 0.032), about
77% of females with more than three costulae were largerthan the average size of a female at that rearing temperature.An influence of rearing temperature on the position of thecostulae relative to the median carina was also observed.Females reared at 30�C had the costulae situated moreproximally on the propodeum than when reared at lowertemperatures (ANOVA test, P< 0.01; F = 12.44; df = 3) (fig. 12).The number of costulae ranged between 2 and 4, and theposition of their junction with the median carina rangedbetween its middle and proximal 1/6 length. The shapeof the costulae was very variable because they were oftenincomplete (fig. 1).
The median carina varied in structure close to theanterior margin of the propodeum, being straight andnarrow (fig. 1a) or widened anteriorly and then forminga small (fig. 1b), medium (fig. 1c) or large triangular cell(fig. 1d). The number of individuals with a larger triangularcell increased with the rearing temperature (ANOVA test,P< 0.01; F = 20.57; df = 3), and specimens assigned to class 4(n= 6) were reared only at 30�C (fig. 13).
Discussion
Pigmentation of gastral tergites
Temperature showed a strong influence on pigmentationof the gastral tergites and other body parts, with both thetergites and tarsi tending to be darker at lower temperatures.
1.56
1.58
1.6
1.62
1.64
1.66
1.68
1.7
15
a
a
bb
20 25 30
Temperature (°C)
Rat
io le
ngth
/wid
th o
f fou
rth
segm
ent o
f fun
icle
Fig. 6. Reaction norm of length/width of fourth funicularsegment of female Pnigalio soemius according to its rearingtemperature (mean+SE).
0.590.6
0.610.620.630.640.650.660.67
15
ab
b
b
a
20 25 30
Temperature (°C)
Leng
th o
f mar
gina
l vei
n (m
m)
Fig. 7. Reaction norm of length of marginal vein of femalePnigalio soemius according to its rearing temperature (mean+SE).
0.1250.13
0.1350.14
0.1450.15
0.1550.16
0.165
15
a
bb
c
20 25 30
Temperature (°C)
Leng
th o
f stig
mat
ic v
ein
(mm
)
Fig. 8. Reaction norm of length of stigmal vein of female Pnigaliosoemius according to its rearing temperature (mean+SE).
3.7
3.9
4.1
4.3
4.5
4.7
4.9
15
a
bbc
c
20 25 30
Temperature (°C)
Rat
io le
ngth
mar
gina
l vei
n /
stig
mat
ic v
ein
Fig. 9. Reaction norm of length of marginal vein/stigmal vein offemale Pnigalio soemius according to its rearing temperature(mean+SE).
104 U. Bernardo et al.

Results for the gastral tergites gave a convex DRN with aminimum around 25�C, which is similar in pattern toresponses for body pigmentation that have been recordedfor other insects (Capy et al., 1988; Laudonia & Viggiani,1993; Petavy et al., 2002; Mound, 2005).
The classical interpretation of the adaptive function ofsuch variation is the thermal budget hypothesis, i.e. a darkercolour absorbs more solar radiation, visible or infrared, atlow temperatures (David et al., 1985, 1990; Kingsolver &Wiersnaz, 1991; Goulson, 1994; Solensky & Larkin, 2003;Gibert et al., 2004). In species that are polymorphic fordorsal colour pattern, individuals that belong to darkmorphs generally warm up more rapidly and attain higherbody temperatures than paler individuals (De Jong et al.,1996; Forsman, 1997, 2000). For example, black individualsof the pygmy grasshopper, Tetrix subulata (Linnaeus)(Orthoptera: Tetrigidae), attain a mean temperature exceed-ing that of grey individuals by 49% (Forsman, 1997).Furthermore, darker forms may have the thermal optimumat a lower level than light-coloured ones; for example,dark coloured (nigra) specimens of Harmonia axyridis (Pallas)(Coleoptera: Coccinellidae) have the thermal optimum 3.7�Clower than light coloured (aulica) specimens (Soares et al.,2003). It also seems that geographical distribution affects thecolour of specimens, with darker individuals at higher
latitudes probably due to lower average temperatures(de Oliveira et al., 2004). Variation in thermal capacity islikely to have pervasive implications for individual fitness,affecting activity period, energy budget, escape capability,dispersal, mating success, and fecundity (De Jong et al., 1996;Gilchrist, 1996; Kingsolver, 1996; Forsman & Appelqvist,1999; Forsman, 2000).
Temperature had less influence on P. soemius males forcolour of the gaster because at 10�C males did not havecompletely black gastral tergites (data not shown). A lessereffect of temperature on males has been observed also forother insects (David et al., 1997; Marriott & Holloway, 1998;Gibert et al., 2004). Forsman (2000) found a lower preferredtemperature for males than for females of pygmy grass-hoppers, which partially reflected the sexual difference inbody size and variation in colour. Males are smaller thanfemales and because of the proportionally larger surface areacould be more susceptible to desiccation, but compensatebehaviourally for this disadvantage by maintaining lowerbody temperatures, thereby reducing evaporative water loss.The preference for a higher body temperature in femalesin pygmy grasshoppers could reflect the strong effectof temperature on reproductive performance (Forsman,2000). The latter author pointed out the strong associationbetween thermal preferences and the ability to attain high
1.04
1.06
1.08
1.1
1.12
1.14
1.16
1.18
15
a
bb
b
20 25 30
Temperature (°C)
Rat
io le
ngth
/wid
th o
f pro
pode
um
Fig. 10. Reaction norm of length/width of propodeum of femalePnigalio soemius according to its rearing temperature (mean+SE).
2
2.2
2.4
2.6
2.8
3
15
aa
ab b
20 25 30
Temperature (°C)
Num
ber
of c
ostu
lae
Fig. 11. Reaction norm of the number of costulae of femalePnigalio soemius according to its rearing temperature (mean+SE).
0.27
0.29
0.31
0.33
0.35
0.37
0.39
0.41
15
a aa
b
20 25 30
Temperature (°C)
Rat
io in
sert
ion
of c
ostu
lae
on
med
ian
carin
a/le
ngth
of p
ropo
deum
Fig. 12. Reaction norm of the intersection of the costulae withthe median carina/length of propodeum of female Pnigaliosoemius according to its rearing temperature (mean+SE).
1.41.61.8
22.22.42.62.8
3
15
a ab
b
c
20 25 30
Temperature (°C)
Wid
th o
f med
ian
carin
a ne
ar
ante
rior
edge
of p
ropo
deum
Fig. 13. Reaction norm of the anterior width of the mediancarina of female Pnigalio soemius according to its rearingtemperature (mean+SE).
Phenotypic plasticity in Pnigalio soemius 105

temperatures across colour morphs of females of pygmygrasshoppers. However, even though colour may have aprofound effect on the capacity for thermoregulation, thismay not necessarily translate into morphospecific differ-ences in performance and fitness (Gilchrist, 1996; Forsman,2000).
Seasonal dichroism in Eulophidae is well documented(Askew, 1971; Barrett et al., 1988) and chromatic variation hasbeen recorded in several other families of Hymenoptera(Viggiani, 1963, 1999; Laudonia & Viggiani, 1993; Zaviezo &Mills, 1999). Thus, the present results suggest that lessimportance should be placed on colour in the construction ofidentification keys for species of Pnigalio.
The present results showed that extent of gastral tergitepigmentation did not have a linear correlation withdecreased temperature because females reared at 30�C weredarker than those reared at 20 and 25�C (fig. 2). This effecthas also been recorded for other insects (Laudonia &Viggiani, 1993; Petavy et al., 2002). It is possible that stressmay be responsible for the development of pigmentation andit is the amplitude of the stress rather than the quality (coldor heat) that determines the magnitude of the effect (Petavyet al., 2001, 2002).
Size
The size of insects is often measured as total body length,but in some cases this measurement is not easily takenbecause it is influenced by several variables (head position,inclination of junctures, segments of the gaster telescoping tovarying degrees in different individuals) (Gauld & Fitton,1987). The choice of a trait that is simple to measureobjectively is not easy; however, fore wing or tibia lengthsare often used to estimate body size variation (Gauld &Fitton, 1987; Bezemer et al., 2005; Chow & Heinz, 2005;Lalonde, 2005). The present results showed that the choice ofa representative size trait is critical because different traitsshowed very different DRNs.
Pnigalio soemius is an idiobiont and, for this kind ofparasitoid, host resources either are static or slowly diminishin quality over time because the host does not feed andgrow. Thus, idiobiont parasitoid offspring size is usuallycorrelated with the size of the host when it is parasitized(Harvey, 2005). Temperature may have influenced thefinal body size of P. soemius adults by two different andsomewhat opposing ways: at higher rearing temperatureslarval hosts could have attained a larger body size beforebeing parasitized, but host degrading processes may befaster at higher temperatures during the growth of theparasitoid larva.
Antenna
Calculated ratios between the length of the clava and firstsegment of the funicle and between the length and width ofthe fourth funicular segment were both highly variable. Theclava varied from being shorter to longer than the firstsegment of the funicle, and the fourth funicular segmentranged from sub-quadrate to rectangular. Because the firstratio and the shape of fourth funicular segment are bothused in taxonomic keys (Graham, 1959; Askew, 1968), theirvariability could result in the ranges of different speciesoverlapping.
Fore wing
Fore wing length decreased with increased rearingtemperature, which is consistent with the pattern typicalof ectothermic animals, and as has been recorded for twodifferent species of Drosophila (Diptera: Drosophilidae)(Atkinson, 1994; David et al., 1997; Morin et al., 1999; Gibert &De Jong, 2001; Petavy et al., 2001). A possible adaptiveexplanation for longer wings at lower temperatures is for alower wing loading when flying in colder temperatures, aswas suggested for Drosophila by Petavy et al. (1997).
Taxonomists often use relative measurements, andespecially ratios between the length of wing veins, becausethese are often considered a means of discriminating species,but the present data showed that wing veins are stronglyinfluenced by overall size variations, and that differentlengths and ratios exhibited very different DRNs. The onlylinear relationship we found was between stigmal vein andfore wing lengths. Moretau et al. (1998) and Gibert et al.(2004) demonstrated similar results for Drosophila melanoga-ster Meigen and D. simulans Sturtevant, with several partsof the wings of these species exhibiting different DRNs.Hoffmann et al. (2005) also showed differences in sizeand shape of the wings for different insects reared underconditions of different stresses that involved low foodquality, addition of ethanol, and cold shock.
Wing morphology is subject to sexual dimorphism(Gauld & Fitton, 1987; Pretorius, 2005; Teder & Tammaru,2005); therefore, analogous studies should be performed formales before extending the present results to this sex.
Propodeum, median carina and costulae
The number of costulae and where these intersectthe median carina are considered important characters inall keys of Pnigalio species (Boucek, 1958; Graham, 1963;Askew, 1968; Miller, 1970; Yoshimoto, 1983). The intra-specific variation of these traits has been poorly investigated(Barrett et al., 1988), but the high variability shown by thepresent results suggests that they should be used carefullyto discriminate among species. Differences in propodealsculpturing of reared males were less pronounced, butsmaller specimens showed a reduced number of costulae inaccordance to what has been reported by other authors(Miller, 1970; Barrett et al., 1998).
Conclusions
Several studies have suggested previously that insects arecharacterized by physiological and behavioural traits thatare temperature dependent, and that the optimal tempera-ture for these traits may vary (Forsman, 2000). When plottedas a function of temperature, several traits we measuredshow a curvilinear relationship with a peak in value at someintermediate rearing temperature. The phenotype of almostall the traits we tested was affected by temperature, and thevariation in these was larger than previously known. Asa consequence, a high number of our specimens couldhave not been correctly keyed on the available keys for theEuropean Pnigalio species (Boucek, 1958; Graham, 1963;Askew, 1968). Consequently, any future revision of Pnigalioshould more carefully assess variation in these traits forspecies recognition.
106 U. Bernardo et al.

It is important to emphasize that we analysed only onepotential source of morphological variability because weused a single host species and the same larval instar ofthat species as host. Several authors have shown for otherinsects that very dissimilar morphs can be correlated withsize or physiological condition of the host (Pinto et al.,1989; Salvo & Valladares, 1995). Polyphagous parasitoids,which utilize a wide range of host species and sizes,could be most markedly affected, even though Salvo &Valladares (1995) did not show a definite trend betweenparasitoid size and host size of polyphagous parasitoids ofleafminers.
Furthermore, the present study evaluated the influenceof only a single environmental factor (temperature) usingparasitoids cultured from a single population from onelocality. Generally, there is a broad parallelism betweengeographic genetic variation and morphological plasticity,and this parallelism is often considered as adaptive(Atkinson, 1994; Gibert et al., 2004). Moreover, with respectto size, it is likely that insects developing under naturalconditions are much more variable than laboratory-rearedinsects, as has been reported for Drosophila (Moretau et al.,1995; David et al., 1997; Gibert et al., 1998; Petavy et al., 2001).For example, Petavy et al. (2001) showed that the adults ofD. melanogaster and D. simulans reared under alternatingconditions were smaller than adults grown under a constanttemperature, and that the magnitude of this effect wasproportional to the amplitude of the thermoperiod. Petavyet al. (2002) also showed that variability in body size of wildflies sometimes can be 10 times that of laboratory-rearedflies. Therefore, the variance of the different features wemeasured for P. soemius could be even larger if a greaternumber of host species and other environmental factors(i.e. humidity) had been included.
This study demonstrates the risk of using any singlefeature when distinguishing species, and that ratios,although certainly more reliable also can be problematicbecause reaction norms based on ratio calculations can havedifferent shapes. Consequently, not only is phenotypicplasticity of great importance to the ecologist and evolu-tionary biologist, it is of basic importance to the taxonomist(Ananthakrishnan, 2005).
Acknowledgements
The authors are grateful to an anonymous referee forextensive revision of the manuscript, Dr Andrew Polaszek(The Natural History Museum, London, UK) for his valuablecomments and Lucio Bernardo for his technical help inevaluating gastral pigmentation on specimens used in thisstudy.
References
Ananthakrishnan, T.N. (2005) Perspective and dimension ofphenotypic plasticity in insects. pp. 1–23 in Anantha-krishnan, T.N. & Whitman, D. (Eds) Insect phenotypic
plasticity diversity of responses. Enfield, New Hampshire,Science Publishers, Inc.
Askew, R.R. (1968) Hymenoptera 2. Chalcidoidea Section (b).Handbooks for the identification of British insects 8(2)b. 39 pp.
Askew, R.R. (1971) Parasitic insects. 316 pp. New York, AmericanElsevier.
Ateyyat, M.A. (2002) Parasitoid complex of citrus leafminer,Phyllocnistis citrella, on lemon in the central Jordan Valley.BioControl 47, 33–43.
Atkinson, D. (1994) Temperature and organism size – abiological law for ectotherms? Advances in Ecological
Research 25, 1–58.Barrett, B.A., Brunner, J.F. & Turner, W.J. (1988) Variations
in colour, size, and thoracic morphology of Pnigaliospecies (Hymenoptera: Eulophidae) parasitizing Phyllonor-
ycter elmaella (Lepidoptera: Gracillariidae) in Utah andWashington. Annals of the Entomological Society of America
81, 516–521.Bernardo, U., Pedata, P.A. & Viggiani, G. (2005) Influenza della
temperatura su alcuni caratteri morfologici e cromatici inPnigalio soemius (Walker) (Hymenoptera: Eulophidae). AttiXX Congresso Nazionale Italiano di Entomologia, Perugia,Assisi 12–18 June 2005, p. 30.
Bernardo, U., Pedata P.A. & Viggiani, G. (2006) Life history ofPnigalio soemius (Walker) (Hymenoptera: Eulophidae) andits impact on a leafminer host through parasitization,destructive host-feeding and host-stinging behavior. Bio-
logical Control 37, 98–107.Bezemer, T.M., Harvey, J.A. & Mills, N.J. (2005) Influence of
adult nutrition on the relationship between body size andreproductive parameters in a parasitoid wasp. Ecological
Entomology 30, 571–580.Boucek, Z. (1958) A study of central European Eulophidae,
I: Eulophinae (Hymenoptera). Sbornık Entomologickeho
Oddeleni Narodnıho Muzea v Praze 33, 117–170.Capy, P., David, J.R. & Robertson, A. (1988) Thoracic trident
pigmentation in natural populations of Drosophila simulans:a comparison with Drosophila melanogaster. Heredity 61,263–268.
Chow, A. & Heinz, K.M. (2005) Using hosts of mixed sizes toreduce male-biased sex ratio in the parasitoid wasp,Diglyphus isaea. Entomologia Experimentalis et Applicata 117,193–199.
David, J.R., Capy, P., Payant, V. & Tsakas, S. (1985) Thoracictrident pigmentation in Drosophila melanogaster: differentia-tion of geographical populations. Genetics, Selection and
Evolution 17, 211–223.David, J.R., Capy, P. & Gautier, J.P. (1990) Abdominal
pigmentation and growth temperatures in Drosophila
melanogaster: similarities and differences in the norms ofreaction of successive segments. Journal of EvolutionaryBiology 3, 429–445.
David, J.R., Gibert, P., Gravot, E., Petavy, G., Morin, J.P.,
Karan, D. & Moreteau, B. (1997) Phenotypic plasticity anddevelopmental temperature in Drosophila: analysis andsignificance of reaction norms of morphometrical traits.Journal of Thermal Biology 22, 441–451.
De Jong, P.W., Gussekloo, S.W.S. & Brakefield, P.M. (1996)Differences in thermal balance, body temperature andactivity between non-melanic and melanic two-spot lady-bird beetles (Adalia bipunctata) under controlled conditions.Journal of Experimental Biology 199, 2655–2666.
de Oliveira, C.M., Lopes, J.R.S., Dias, C.T.D.S. & Nault, L.R.
(2004) Influence of latitude and elevation on polymorphismamong populations of the corn leafhopper, Dalbulus maidis
(DeLong and Wolcott) (Hemiptera: Cicadellidae), in Brazil.Environmental Entomology 33, 1192–1199.
Forsman, A. (1997) Thermal capacity of different colour morphsin the pygmy grasshopper Tetrix subulata. Annales ZoologiciFennici 34, 145–149.
Phenotypic plasticity in Pnigalio soemius 107

Forsman, A. (2000) Some like it hot: intra-population variation inbehavioral thermoregulation in colour-polymorphic pygmygrasshoppers. Evolutionary Ecology 14, 25–38.
Forsman, A. & Appelqvist, S. (1999) Experimental manipulationreveals differential effects of colour pattern on survivalin male and female pygmy grasshoppers. Journal of
Evolutionary Biology 12, 391–401.Gauld, I.D. & Fitton, M.G. (1987) Sexual dimorphism in
Ichneumonidae: a response to Hurlbutt. Biological Journal
of the Linnean Society 31, 291–300.Gibert, P. & De Jong, G. (2001) Temperature dependence of
development rate and adult size in Drosophila species:biophysical parameters. Journal of Evolutionary Biology 14,267–276.
Gibert, P., Moreteau, B., David, J.R. & Scheiner, S.M. (1998)Describing the evolution of reaction norm shape: bodypigmentation in Drosophila. Evolution 52, 1501–1506.
Gibert, P., Capy, P., Imasheva, A., Moreteau, B., Morin, J.P.,
Petavy, G. & David, J.R. (2004) Comparative analysisof morphological traits among Drosophila melanogaster andD. simulans: genetic variability clines and phenotypicplasticity. Genetica 120, 165–179.
Gilchrist, G.W. (1996) Quantitative genetic analysis of thermalsensitivity in the locomotor performance curve of Aphidiuservi. Evolution 50, 1560–1572.
Goulson, D. (1994) Determination of larval melanization in themoth, Mamestra brassicae, and role of melanin in thermo-regulation. Heredity 73, 471–479.
Graham, M.W.R. de V. (1959) Keys to the British generaand species of Elachertinae, Eulophinae, Entedontinae andEuderinae (Hym., Chalcidoidea). Transactions of the Society
for British Entomology 13, 169–204.Graham, M.W.R. de V. (1963) Additions and corrections to the
British list of Eulophidae (Hym., Chalcidoidea). Transac-tions of the Society for British Entomology 15, 167–275.
Harvey, J.A. (2005) Factors affecting the evolution of develop-ment strategies in parasitoid wasps: the importanceof functional constraints and incorporating complexity.Entomologia Experimentalis et Applicata 117, 1–13.
Hoffmann, A.A., Woods, R.E., Collins, E., Wallis, K., White, A.
& McKenzie, J.A. (2005) Wing shape versus asymmetryas an indicator of changing environmental conditions ininsects. Australian Journal of Entomology 44, 233–243.
Kingsolver, J.G. (1996) Experimental manipulation of wingpigment pattern and survival in western white butterflies.American Naturalist 147, 296–306.
Kingsolver, J.G. & Wiernasz, D.C. (1991) Seasonal polyphenismin wing-melanin pattern and thermoregulatory adaptationin Pieris butterflies. American Naturalist 137, 816–830.
Lalonde, R.G. (2005) Egg size variation does not affect offspringperformance under intraspecific competition in Nasonia
vitripennis, a gregarious parasitoid. Journal of Animal Ecology74, 630–635.
Laudonia, S. & Viggiani, G. (1993) Effetto della temperaturasulla colorazione degli adulti di Encarsia partenopea
Masi (Hymenoptera: Aphelinidae). Bollettino del Laboratoriodi Entomologia Agraria Filippo Silvestri 50, 141–146.
Marriott, C.G. & Holloway, G.J. (1998) Colour pattern plasticityin the hoverfly, Episyrphus balteatus: the critical immaturestage and reaction norm on developmental temperature.Journal of Insect Physiology 44, 113–119.
Miller, C.D. (1970) The Nearctic species of Pnigalio and Sympiesis
(Hym. Eulophidae). Memoirs of the Entomological Society of
Canada 68, 1–121.
Moretau, B., Capy, P., Alonso-Moraga, A., Munoz-Serrano, A.,
Stockel, J. & David, J.R. (1995) Genetic characterization ofgeographic populations using morphometrical traits inDrosophila melanogaster : isogroups versus isofemale lines.Genetica 96, 207–215.
Moretau, B., Imasheva, A.G., Morin, J.P. & David, J.R. (1998)Wing shape and developmental temperature in twoDrosophila sibling species: different wing regions exhibitdifferent norms of reaction. Russian Journal of Genetics 34,183–192.
Morin, J.P., Moreteau, B., Petavy, G. & David, J.R. (1999)Divergence of reaction norms of size characters betweentropical and temperate populations of Drosophila melanoga-
ster and D. simulans. Journal of Evolutionary Biology 12,329–339.
Mound, L.A. (2005) Fighting, flight and fecundity: behaviouraldeterminants of Thysanoptera structural diversity.pp. 81–105 in Ananthakrishnan, T.N. & Whitman, D. (Eds)Insect phenotypic plasticity diversity of responses. Enfield,New Hampshire, Science Publishers, Inc.
Petavy, G., Morin, J.P., Moreteau, B. & David, J.R. (1997)Growth temperature and phenotypic plasticity in twoDrosophila sibling species: probable adaptive changesin flight capacities. Journal of Evolutionary Biology 10,875–887.
Petavy, G., Moreteau, B., Gibert, P., Morin, J.P. & David, J.R.
(2001) Phenotypic plasticity of body size in Droso-phila: effects of a daily periodicity of growth tempera-ture in two sibling species. Physiological Entomology 26,351–361.
Petavy, G., Moreteau, B., Gibert, P. & David, J.R. (2002)Phenotypic plasticity of body size in Drosophila: influence ofa developmental thermoperiodic regime in two siblingspecies. Physiological Entomology 27, 124–135.
Pinto, J.D., Velten, R.K., Platner, G.R. & Oatman, E.R. (1989)Phenotypic plasticity and taxonomic characters in Tricho-
gramma (Hymenoptera: Trichogrammatidae). Annals of the
Entomological Society of America 82, 414–425.Pretorius, E. (2005) Using geometric morphometrics to investi-
gate wing dimorphism in males and females of Hymenop-tera – a case study based on the genus Tachysphex Kohl(Hymenoptera: Sphecidae: Larrinae). Australian Journal of
Entomology 44, 113–121.Salvo, A. & Valladares, G. (1995) Intraspecific size variation
in polyphagous parasitoids (Hym.: Parasitica) of leafminers and its relation to host size. Entomophaga 40,273–280.
Soares, A.O., Coderre, D. & Schanderl, H. (2003) Effect oftemperature and intraspecific allometry on predation bytwo phenotypes of Harmonia axyridis Pallas (Coleoptera:Coccinellidae). Environmental Entomology 32, 939–944.
Solensky, M.J. & Larkin, E. (2003) Temperature-inducedvariation in larval coloration in Danaus plexippus (Lepido-petra: Nymphalidae). Annals of the Entomological Society of
America 96, 211–216.Statgraphics plus (1997) Version 3.0. Manugistics. Maryland,
USA.Teder, T. & Tammaru, T. (2005) Sexual size dimorphism within
species increases with body size in insects. Oikos 108,321–334.
Via, S., Gomulkiewicz, R., De Jong, G., Scheiner, S.M.,
Schlichting, C.D. & Van Tienderen, P.H. (1995) Adaptivephenotypic plasticity: consensus and controversy. Tree 10,212–217.
108 U. Bernardo et al.

Viggiani, G. (1963) Osservazioni sulla morfo-biologia delPnigalio mediterraneus Ferr. & Del. (Hym.: Eulophidae).Entomophaga 8, 191–198.
Viggiani, G. (1999) Variations and biological traits of Coccopha-gus gossypariae Gahan (Hymenoptera: Aphelinidae). Bio-
logical Control 16, 43–46.Yoshimoto, C.M. (1983) Review of the North American Pnigalio
Schrank (Hymenoptera: Eulophidae). Canadian Entomologist115, 971–1000.
Zaviezo, T. & Mills, N. (1999) Aspects of the biology of Hyssopuspallidus (Hymenoptera: Eulophidae), a parasitoid of thecodling moth (Lepidoptera: Olethreutidae). Environmental
Entomology 28, 748–754.
(Accepted 12 October 2006)� 2007 Cambridge University Press
Phenotypic plasticity in Pnigalio soemius 109




![GINGIVAL PIGMENTATION · Oral Pigmentation Oral pigmentation is a discolouration of the oral mucosa or gingiva associated with several exogenous and endogenous factors.[1-3] This](https://img.dokumen.tips/doc/110x75/5f82dc5ea46ef73d4a1ef172/gingival-pigmentation-oral-pigmentation-oral-pigmentation-is-a-discolouration-of.jpg)
























