Embed Size (px)
Citation preview

1
Oxytocin and Vasopressin in Treating
Autism: Results of Comparative Research
by Daniel M. Hitzfield
Abstract
Using animal models to understand the mechanisms underlying human social behavior
reveals otherwise inaccessible neurophysiological processes. There are three purposes to this
paper. First, to review the findings of prominent comparative research on oxytocin and
vasopressin in regards to their effect on adult social bonding. Second, to summarize
advancements made in treating autism, a social disorder. Third, to discuss the limits of
comparing animal models to human research.

2
Introduction
Long-term, healthy social affiliations (e.g., relationships) have been clinically correlated
with the peak mental health of human individuals (Lim & Young, 2006). The latter century has
yielded the necessary equipment for scientific studies to pinpoint the areas of the human brain
associated with social behavior, such as the amygdala and medial prefrontal cortex (Fossati,
2012). Unfortunately, the neuroendocrinology of human social behavior remains under research
without a wholly consistent explanation for its effects; it is considered a critical area of
neuroscience to explore because understanding the neurophysiological mechanisms will
hopefully highlight possible treatments for human social disorders. This ongoing work is still
largely in the stages of correlating specific pathways with subsets of behavior (e.g., Zink et al.,
2011). At best, analyzing just the human brain and relevant behavior in case studies only allows
for a superficial understanding of the adult mental and behavioral processes governing social
interaction. Animal models, on the other hand, have been previously shown to shed light on the
mental and behavioral processes of humans without overstepping the bounds of ethics that would
be violated in similar experiments actually involving humans (Modi & Young, 2012). Moreover,
animal correlates to human behavior are practically necessary to uncover the the most effective
size, time period and method of treatment in many cases. Defining the effect of oxytocin (OT)
and vasopressin (VP) treatments on animals' adult social affiliative behavior will be the former
purpose of this paper. The latter will be review and summary of case studies and other
experiments performed on humans regarding OT and VP treatment as a potential cure for autism
spectrum disorder (ASD). ASD covers a range of dysfunctional behaviors from almost
unnoticeable to crippling degrees of social impairment.

3
Adult Social Affiliation
Affiliative behavior is defined as any action that fosters social relationships, or bonds.
Regarding humans, this set of behaviors dominates life and relationships with peers of either sex
(Lim & Young, 2006). As peer relationships are the most common, they will be the sole
subsection of affiliate behavior discussed. The most long-lasting of these relationships are often
those of pair-bonds (i.e., sexual partners), as evidenced by the research of Young & Wang (2004).
Pair bonds are typically judged by recording the quantity of particular behaviors in non-human
experiments, such as the work done by Garber & Leigh (1997) on eleven different species of
New World primates. These can include social grooming (i.e., allogrooming), sharing resources
and huddling together (Garbe & Leigh, 1997; Adiseshan et al., 2011). Other experiments on
primates, particularly a study done on macaques by Adiseshan et al. (2011), demonstrated much
longer-term social relationships on a scale beyond pair bonds; these varied by kin (i.e., extended-
family alliances) or simple living-proximity group alliances (Adiseshan et al., 2011).
Furthermore, maintenance of these larger-scale alliances has been observed as conciliatory
behavior soon after aggression occurs by one or both parties involved (Cords and Aureli, 1997).
The importance of this social activity as a whole is confirmed by the majority of time spent in
affiliative behavior by primates in general: anywhere from 50-98% (Sussman et al., 2005). This
behavior (ie making and maintaining friendship) is also observed alongside and blunting
aggressive or competitive behavior in a variety of non-human species (Sussman et al., 2005).
These studies of animals and particularly of primates correlate well with studies of equivalent
human social affiliative behavior (Ebstein et al., 2012), making comparison the most promising
method of determining the most efficacious treatments of ASD.

4
Mechanisms Influencing Social Affiliation
In the history of neurophysiology in relation to social disorders, research on humans in
the past century has been limited to individuals with social impairments or pre-existing brain
damage (as opposed to lesioning performed on non-humans); in the latter part of this century
brain-imaging techniques possible because of developing technology have become the forefront
methods in human studies (Kennedy & Adolphs, 2012). While such studies provide a chance to
determine ultimate cause-and-effect, regulatory pathways and their intermediates are still not
coherently understood to a satisfactory degree. Moreover, the subject of social relationships and
its set of behaviors is a difficult one, because it relies upon the processing of information on
multiple levels of consciousness. In order to make progress, one level of these processing
pathways in the brain is examined at a time and for a particular system. In this case, that system
is the pathway(s) by which OT and VP regulate social cognition and emotion. Fortunately, non-
human social behaviors recorded in a lab setting correlate well with observation of natural non-
human social behavior (Lim & Young, 2006) and with human brain-imaging results (Zink &
Meyer Lindenberg, 2012).
Regarding the influence of OT and VP on social affiliative behavior, researchers have
particularly focused on the prairie vole (Microtus Ochrogaster) and experimental manipulation
of its naturally strong monogamous traits. Earlier experiments were only performed on a
relatively short-term basis (e.g., Carter et al., 1995; Cho et al., 1999; Young & Wang, 2004),
while experiments in the last year or so have expanded experimental parameters to the entire
lifetime of the prairie vole (e.g., Bales et al., 2013). Another species that has proven useful is the
Wistar rat (Rattus Norvegicus), used to determine specific oxytonergic and vasopressinergic

5
pathways and observe multiple social behaviors (e.g., De Vries & Bujis, 1983). Similarly, mice,
whether bred as transgenic comparisons to other species or tested as wild-types, have provided
additional data on social behavior (e.g., Young et al., 1999).
Neuropeptides Implicated in Regulating Social Behavior
According to work done by De Vries & Bujis (1983) on rats, OT is originally
manufactured by the paraventricular nuclei (PVN) or supraoptic nuclei (SON) of the
hypothalamus in order to act on its corresponding oxytocin receptors (OTR) scattered in the
brain; this was proven in the experiment by lesioning the PVN, which almost completely
crippled the oxytocinergic system. In a review of OT and VP, Sofroniew (1983) supported the
conclusions of De Vries & Bujis, adding that both the PVN and SON project toward the posterior
pituitary (i.e., neurohypophysis) and both OT and VP are secreted from that point as peptides
into peripheral circulation; OT in particular was associated with numerous projection sites from
the PVN: the hippocampus, striatum, nucleus accumbens and spinal cord. Like OT, VP is
produced in the PVN and SON; it is additionally produced in the suprachiasmatic nucleus, bed
nucleus of the stria terminalis and medial amygdala (De Vries & Miller, 1998). Either VP or OT
that is released centrally instead of peripherally has been determined for some time to have a
strong influence on social behavior; evidence that has been uniform across numerous rodent and
other mammalian experimental subjects (Lim & Young, 2006). These two have been the
promising candidates for research in later decades because of their additional evidence for
altering neuronal activity.
Adult Affiliative Behavior: Oxytocin
Rats are ordinarily social animals with several distinctive non-sexual affiliative behaviors

6
which makes them one of the more tested subjects to determine the role of OT. In a study very
recently performed on them by Calcagnoli et al. (2013), it was determined that acute, short-term,
centrally-injected doses of OT resulted in two distinctive behavior changes: reduced levels of
anti-social intruder aggression, and marked increase of time spent in social contact. Note, “social
contact” simply denotes exploration among and proximity with other individuals, not necessarily
the observation of specific behaviors. In the same experiment, another experimental group was
centrally-injected with an OT antagonist, resulting in a non-significant increase of aggression
among naturally low-aggression rats, and almost no difference in ordinarily high-aggression rats.
In short, observation for one hour after the intracerebroventricular OT injections did not yield
any significant increase in the behaviors correlated with social affiliation, instead merely
lowering aggression. While most other experiments over the last two decades have studied the
immediate affiliative behavioral effects of experiments on males rats such as the above, Witt et
al. (1992) performed continuous observation for six hours after centrally administering OT to
male rats. All the experimental and control males still had their gonads intact and hormonal
secretion uninterrupted, paired equally between estrous and ovariectomized females. The only
significant difference were two times the number of non-sexual, specifically affiliative social
behaviors—such as grooming and anogenital sniffing—in the experimental group compared to
the control group. The conclusion is that constant exposure to OT heavily influences the
tendency to engage in social affiliative interactions given a relatively long time to exert its
influence (e.g., six hours or more).
Later research on adult male house mice (Mus Musculus) by Murakami et al. (2011)
supported the associations made in rats; social behavior was linked by simple correlations with

7
mRNA and neuropeptide receptor expression. The medial amygdala and PVN were the primary
sites of activity for mRNA expression of OT and VP receptors and their corresponding
neuropeptides, respectively. Interestingly, the levels of OT and VP mRNA were associated with
estrogen receptors as well as increased social interaction, suggesting that estrogen receptor
activity has a strong regulatory influence on both hormones' expression.
Interestingly, the results from Carter et al. (1995) in an experiment on monogamous
prairie voles (Microtus Ochrogaster) yielded somewhat different results than the aforementioned
research with male rats. Pair-bonding took place to different degrees, measured by observation of
affiliated behaviors under the influence of various test variables, such as the injection of VP,
injection of OT, and being allowed to mate. Unlike in male rats, centralized injection of OT into
male voles did not produce a significantly stronger effect on pair-bonding, while it did in females
even in the absence of mating (Carter et al., 1995). In a similar experiment years later which
involved hour-long OT and/or VP doses alongside mating, similar results were obtained with an
increased rate of pair-bonding only with the presence of mating alongside injections (Cho et al.,
1999). Unfortunately, OT injections that last from childhood to adulthood appear to have a far
less positive effect than much more short-term injections. According to the latest research
performed by Bales et al. (2013) on prairie voles, long-term developmental treatment causes a
lack of discernment in social interactions. The frequency of expected behaviors increased, but
not in accordance with normal partner-preference. That is, affiliative behaviors will increase to
the point that the subject will not bond with any other; the subject will effectively act
polygamous in a typically monogamous species.
Attempting to apply both rat and vole models to humans at face value is problematic.

8
However, the key for interpreting these results is the baseline behavior. In the case of rats, there
is no long-lived or lifelong pair-bonding. As a result, it was expected that an OT injection would
increase nonspecific social behavior. It did. In the case of prairie voles, the baseline behavior is
vastly different: lifelong monogamy. It was unknown whether an OT injection would strengthen
or erode a pair-bond in its effect. The results, as discussed, were intriguing. Acute injections
temporarily increased partner preference only when mating also occurred. Chronic injections
crippled partner preference, increasing social behavior to the point of almost complete non-
specificity. This is the common link between the two models, and the application to humans. OT
injections can be said to increase non-specific social affiliative behavior, the results differing by
time period of injections and the baseline of behavior. If the baseline is pure monogamy, chronic
injections can cripple that baseline. In the case of humans, our baseline is far from clear; we can
be monogamous, polygamous, promiscuous, etc. There is no direct correlation we can make in
applying the rat and vole models to humans by dosage and time period of OT treatment.
Humanity cannot be neatly lined up against a monogamous or promiscuous animal species; it is
somewhere in between. Hence, the value of both animal models.
This “somewhere in between” postulation can be seen played out in primates. In a study
on macaques done by Rosenblum et al. (2002), OT levels significantly fluctuated between
species; in the relatively social Bonnet macaques (Macaca Radiate), centralized OT
concentrations located in the cerebrospinal fluid was far higher than in comparatively asocial
Pigtail macaques (Macaca Nemestrina). This research implies that there is a range of effect for
OT, dependent on genetically-determined receptor density and external variables, such as mating.
Or in other words, the baseline social behavior tied to OT can vary even within a species!

9
Adult Affiliative Behavior: Vasopressin
The effect of centralized VP infusions in voles recorded by Young et al. (1999) had
intriguing results, differing in whether the species of vole tested was ordinarily monogamous or
promiscuous; the monogamous prairie voles, normally social to begin with, had their affiliative
behavior significantly increased while promiscuous montane voles had little to no change in the
same subset of behavior (Young et al., 1999). The researchers involved postulated that this
difference in effect was a result of the natural and far lower density of vasopressin receptors
(VPR) in montane voles. To test this, transgenic house mice (Mus Musculus) were bred by the
same researchers with a genetic alteration in VPR expression to mimic the densities of prairie
voles; in testing with OT infusions the same results were obtained as with the voles—higher
social afffiliative behavior (Young et al., 1999). From this, the researchers were confirmed in
their hypothesis: VP, like OT, has a range of effect dependent on receptor density.
In some of the most recent research on rats done by Veneema et al. (2013), juvenile rats
were tested in which social affiliative play behavior was recorded while under the effect of
centralized (eg intracerebroventricular) injections of VP a1 receptor antagonist. Compared to
frequency of social play exhibited by control juveniles, male experimental subjects had their
frequency drastically reduced, while female subjects actually increased their frequency. When
the VP antagonist specifically targeted the lateral septum, the opposite results occurred: males
demonstrated a higher frequency of social play while females displayed less (Veneema et al.,
2013). This showcases how VP's effect on social affiliative behavior is dependent on both sex
and brain region.
Adult Affiliative Behavior: Vasopressin and Oxytocin Interactions

10
In the case of Microtus Ochrogaster tested by Cho et al. (1999), it was determined that
OT or VP injection alone (without any external stimulus, e.g., mating) only results in the
occurrence of more pro-social behavior unrelated to forming individual social bonds. On the
other hand, the presence of both OT and VP with functioning receptors was enough to produce a
mild increase in partner-preference even without the added stimulus of mating, suggesting that
OT and VP must act in concert to positively influence pair-bonding. Likewise, OT and VP
injections had no effect when either OTR or VPR antagonists were present. This was confirmed
also by the work of Veneema et al. (2013) in their testing of OT receptor antagonist apart from
the VP antagonist, which had no appreciable effect alone.
Human Social Brain and Abnormalities
As a result of advancing technology (i.e., neuroimaging, fMRI, etc) studies have been
able to pinpoint areas of the brain in relation to social cognition. These have largely validated the
physiological discoveries in model animals, as shall be discussed.
For instance, in both rats and mice the medial amygdala is the primary site of expression
for OTR and VPR; this was also confirmed in humans by Kirsch et al. (2005) using fMRI
techniques. In their clinical study of fifteen adult males, OT was intranasally administered and
then each man was tested with negatively emotional stimuli (e.g., fear). The experimental group
displayed drastic reduction of amygdala activity, muting the negative social effect of fear (Kirsch
et al., 2005). A similar study using intranasally-administered OT in conjunction with fMRI done
by Baumgartner et al. (2008) associated a reduction of activity in the amygdala, midbrain, and
dorsal striatum with a suppression of fear and corresponding increase in trust. These results also
confirm the activity of the striatum seen in rats as a result of secretions from the PVN and SON.

11
From these, it can be said that OT reduces negative social emotions (e..g, fear) by directly or
indirectly suppressing activity in the amygdala. The same ultimate results were determined for
VP in one of the first studies done for it by Zink et al. (2010), in which adult men were given
intranasal VP and tested for emotional processing of faces. The fMRI results showed greater
activity in the amygdala and reduced activity in the medial prefrontal cortex (mPFC) for the
experimental group, correlated with reduced fear. The interpretation of these results holds true
with the other experiments described—the connections between the amygdala, the fear center,
and the mPFC are more frequently activated in the presence of negative stimuli (e.g., angry
faces), but the presence of VP mutes the resulting activity in the mPFC.
These studies discussed not only confirmed general areas of OT and VP activity also
found in animal models, but pinpointed an additional area in humans linked to complex social
cognition: the prefrontal cortex. Research is still ongoing on this portion of the human brain, and
conclusions about functional connections are very incomplete, mostly relying on derivation from
primate models such as the macaque (Petrides & Pandya, 1999). Unfortunately, these models are
limited in that most primates have a far less developed PFC than humans. In the last decade,
direct neuro-imaging of the human brain has revealed far more of the mPFC's functions which
OT and VP affect. To summarize over a decade's worth of such direct research and not delve into
topics beyond the scope of this paper, the mPFC has been divided into three major portions, each
activated by a particular type of social cognition, outlined in Figure 1.1. While these discoveries
are undoubtedly of use in many areas of therapy, there have not been enough at this point to base
any direct injections of hormones into the mPFC for treatment of ASD.
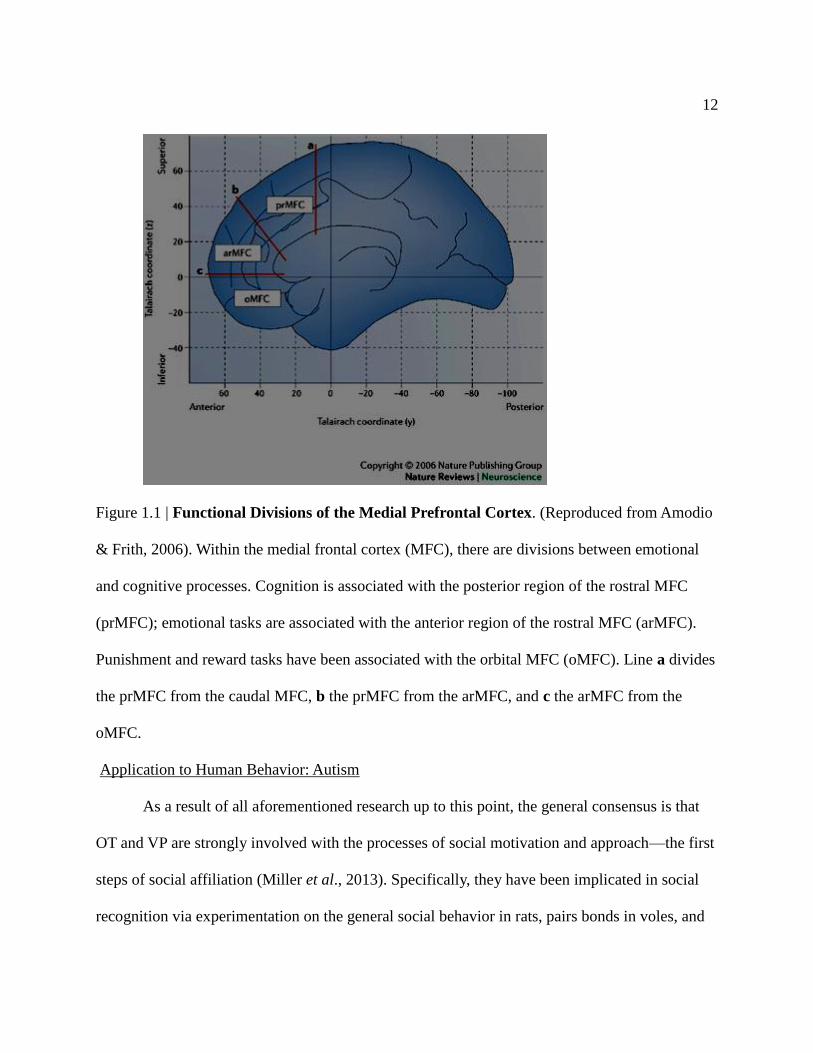
12
Figure 1.1 | Functional Divisions of the Medial Prefrontal Cortex. (Reproduced from Amodio
& Frith, 2006). Within the medial frontal cortex (MFC), there are divisions between emotional
and cognitive processes. Cognition is associated with the posterior region of the rostral MFC
(prMFC); emotional tasks are associated with the anterior region of the rostral MFC (arMFC).
Punishment and reward tasks have been associated with the orbital MFC (oMFC). Line a divides
the prMFC from the caudal MFC, b the prMFC from the arMFC, and c the arMFC from the
oMFC.
Application to Human Behavior: Autism
As a result of all aforementioned research up to this point, the general consensus is that
OT and VP are strongly involved with the processes of social motivation and approach—the first
steps of social affiliation (Miller et al., 2013). Specifically, they have been implicated in social
recognition via experimentation on the general social behavior in rats, pairs bonds in voles, and

13
development of widespread social alliances in primates. Considering how strong this implication
is across lower mammals, they are likewise strong candidates for explaining modulation of
human social affiliative behavior and therefore similar or even identical treatment may improve
disorders such as autism.
Given that humans are naturally social animals and form pair-bonds in addition to wider
social structures, it is assumed that any abnormal amount of asocial behavior is linked with a
deficit in the oxytocinergic and vasopressinergic systems (Green et al., 2001; Modi & Young,
2012). That is to say, genetic defects hindering or crippling such systems creates a tendency for
the same behavior seen in an animal species such as Pigtail macaques, which has a lower
expression of the relevant hormone receptors to begin with. This assumption has been solidly
confirmed in multiple forms. According to the research of Green et al. (2001), the defect was an
unusually high concentration of amidated and C-terminal extended OT prohormones to OT in
blood plasma. This suggests an enzymatic malfunction without any error in the genes that code
for OT's precursors, but that does not mean autism cannot also be linked to other genetic
malfunctions.
Some of the most recent research of the human OTR gene and genetic variation in it has
been implicated to be a contributor to ASD in a case-control study (MA et al., 2013). Moreover, a
specific genetic difference was identified to correlate with ASD in these cases: rs35062132.
Expressed OTRs with this variation exhibited increased rate of endocytosis of bound OT and
decreased levels of free Calcium ions; increased cycling by this particular OTR variant may
decrease overall levels of circulating OT and therefore diminish the positive influence on social
behavior. This step forward implies that autism may not only be eventually aided by OT or OTR

14
treatment, but also by genetic manipulation.
Before any non-genetic treatments are discussed below, the value of combining intranasal
OT or VP administration with fMRI techniques must be discussed. It is hypothesized that
administering peptides intranasally lets them move through cerebral spinal fluid to the brain from
nasal epithelium clefts; this ultimate result has been supported by the recorded effects of such
treatment, which will be later discussed. fMRIs have been critical in confirming increased
activity in the brain after administration, clinching ultimate cause and effect. However, the exact
mechanism by which OT and VP affect receptors in the brain—directly or indirectly—is not
clear.
A two-month treatment of transcutaneous electrical acupoint stimulation (TEAS) was
recently explored as a possible therapeutic solution for ASD by Zhang et al. (2012) because it
increases release of both OT and VP in the human brain. The basis for this experiment was past
experimentation on rats by Yang et al. (2007) which increased measured levels of VP and OT in
the brain via acupuncture stimulation, and later studies on the improvement of social behavior in
children with ASD via similar electro-acupuncture (Wong & Chen, 2010). While the exact
mechanisms of action by which acupuncture increases activity of OT and VP systems in the brain
is unknown, the previous results mentioned were encouraging enough to warrant further testing,
and peripheral hormone levels could be implied to correlate with central levels given a
significant deviation from normal concentrations at the time of sampling (e.g., while asleep and
compared to a control group). Sampling of plasma by enzyme immunoassay by Zhang et al.
(2012) yielded higher concentrations of both OT and VP in the experimental group, correlated
with significant reduction of asocially-linked emotions—such as fear and anxiety—along with

15
firmer control over them when they did occur. Interestingly, the treatment was only effective for
those that were withdrawn and/or passive, not those already aggressive but still asocial (Zhang et
al., 2012).
In a shorter-term experiment performed by Guastalla et al. (2008), a twenty-four hour
intranasal administration of OT to fifty-two human males preceded a thorough test of an
subconscious behavior—eye gaze—tied to social cognition. The experimental group displayed
both increased frequency of locking eyes onto the recorder and overall time of doing so
(Guastalla et al., 2008). Likewise, another double-blind study done by Mikolajczak et al. (2010)
determined that the measure of trust displayed under the effect of OT with either of two different
values at stake—currency and confidential information—was higher, confirming previous
studies' results of the same point and eliminating a confounding factor of that same experiment.
On the other side of the scale, intranasally-administered OT to eighteen human males in a
double-blinded experiment was found to significantly decrease the frequency of aversion
displayed to an angry facial expression (Evans et al., 2010). Moreover, this reduction of negative
social behavior (aversion) was determined to be independent of other financial input, inferring
that OT specifically affects social behavior without direct interaction of others.
Brunnlieb et al. (2013) performed a very similar experiment with intranal administration
of VP on adult men; activation of the right amygdala shown via fMRI was correlated with greater
emotional empathy while the subjects viewed a socially negative scene. Compared to the control
group, the experimental group showed increased activity alongside increased connectivity with
the medial prefrontal cortex (mPFC). Not only was VP confirmed to modulate social emotions,
but also the means of doing so: by reducing suppression of mPFC activity, which in turn

16
stimulated the right amygdala. This experiment was unfortunately unable to confirm whether VP
had a direct impact as well.
The effects of OT and VP are more complicated than a simple abnormal concentration
across all humans, unfortunately. In one of the most recent clinical studies on the OT and VP
levels in blood plasma of children by Miller et al. (2013), significant differences were found by
sex and behaviors associated with severe ASD. Females had higher abnormal concentrations of
OT associated with greater anxious behavior and lower VP than males associated with repeated,
rigid mannerisms. Males on the other hand had no significant amount of behaviors that set them
apart from females, despite their higher relative concentrations of VP. These results would
suggest that future treatment of ASD may need to be altered according to sex and desired
behavioral patterns.
Because of the effectiveness of treatments to date, short-term treatments would be
limited to hormonal injections and/or stimulation, and a long-term treatment that hopefully lasts
for life would be genetic recombination.
Limitations on Research and Application
The insights provided by comparative research between humans and animals is relatively
undisputed in reference to multiple generalities (Modi & Young, 2012). Most research has been
focused on the more promising model species: voles, rats, primates, and cats, to name a few.
Unfortunately, with the development of techniques such as functional magnetic resonance
imaging (fMRI), significant variations are being found between even human and primate brains,
which are often the prime correlate utilized in experiments (Sinha, 2005). In recent research by
Zink et al. (2011), fMRI located the left temporoparietal junction in humans to be a critical

17
binding site for VP. As research such as this opens new possibilities of treatment, it begins to
make comparative research obsolete. Moreover, any assumption of the same neurophysiological
mechanisms driving behavior between correlate species is suspect, unless it falls under those
well-documented generalities mentioned previously in the paper (e.g., the amygdala and mPFC).
For instance, there is increasing evidence that Bonnet macaques have a wide range of social
tendencies even within tight-knit social groups as a result of a myriad of factors, including
cultural learning (Sinha, 2005). In order to make comparative research more applicable, a wider
range of species tested is necessary to uncover further generalities. Once discovered, these can
provide material from which to extrapolate the function of various structures, as utilized in the
review by Modi & Young (2012).
Conclusion
The majority of the research to date in social neurophysiology for application to disorders
like ASD has been comparative. While this work has laid the foundation for general aims in
future studies and given a number of fruitful insights, caution is necessary; there is simply too
much inter-species and inter-individual variation in the details of structure and function. Recent
advancements in technology and methods also provide direct evidence from which to derive
treatments, subduing the practical use of comparison to model animals (Zink et al., 2011; Zhang
et al., 2012; Zink & Meyer-Lindenberg, 2012). As the primary goal of this field is to treat
disorders, the frequency of experiments will undoubtedly veer toward direct methods in the
future. Treatments for ASD, such as TEAS, have already been proven to have limited value
(Zhang et al., 2012). The value of comparative research remains, though; it is a guide and source
of information from which to aim direct research on humans instead of inevitably inaccurate

18
correlations.

19
Literature Cited
1. Adiseshan, A., Adiseshan, T. & Isbell, L.A. 2011. Affiliative Relationships and
Reciprocity among Adult Male Bonnet Macaques at Arunachala Hill, India. American
Journal of Primatology. 73, 1107-1113.
2. Bales, K.L., Perkeybile, A.M., Conley, O.G., Lee, M.H., Guoynes, C.D., Downing, G.M.,
Yun, C.R., Solomon, M., Jacob, S. & Mendoza, S.P. 2013. Chronic Intranasal Oxytocin
causes Long-Term Impairments in Partner Preference Formation in Male Prairie Voles.
Biol. Psychiatry, 74(3), 180-188.
3. Baumgartner, T., Heinrichs, M., Vonlanthen, A., Fischbacher, U. & Fehr, E. 2008.
Oxytocin Shapes the Neural Circuitry of Trust and Trust Adaption in Humans. Neuron.,
58, 639-650.
4. Brunnlieb, C., Munte, T.F., Tempelmann, C. & Heldmann, M. 2013. Vasopressin Modules
Neural Responses Related to Emotional Stimuli in the Right Amygdala. Brain Res., 7,
29-42.
5. Calcagnoli, F., de Boer, S.F., Althaus, M., den Boer, J.A. & Koolhaas, J.M. 2013.
Antiaggressive Activity of Central Oxytocin in Male Rats. Psychopharmacology, (Apr
28) [Epub ahead of print].
6. Carter, C.S., DeVries, A.C. & Getz, L.L. 1995. Physiological Substrates of Mammalian
Monogamy: the Prairie Vole Model. Neuroscience & Biobehavioral Reviews, 19, 303-
314.
7. Cho, M.M., DeVries, A.C., Williams, J.R. & Carter, C.S. 1999. The Effects of Oxytocin
and Vasopressin on Partner Preferences in Male and Female Prairie Voles (Microtus
Ochrogaster). Behavioral Neuroscience, 113, 1071-1079.
8. Cords, M. & Aureli, F. 1997. Reasons for Reconciling. Evolutionary Anthropology, 5, 42-
45.
9. De Vries, G.J. & Miller, M.A. 1998. Anatomy and Function of Extrahypothalamaic
Vasopressin Systems in the Brain. Progress in Brain Research, 119, 3-20.
10. De Vries, G.J. & Buijs, R.M. 1983. The Origin of the Vasopressinergic and Oxytocinergic
Innervation of the Rat Brain with Special Reference to the Lateral Septum. Brain Res.,
273(2), 307–317.
11. Ebstein, R.P., Knafo, A., Mankuta, D., Chew, S.H. & Lai, P.S. 2012. The Contributions of
Oxytocin and Vasopressin in Pathway Genes to Human Behavior. Hormones and
Behavior, 61, 359-379.

20
12. Evans, S., Shergill, S.S. & Averbeck, B.B. 2010. Oxytocin Decreases Aversion to Angry
Faces in an Associative Learning Task. Neurpsychopharmacology, 35(13), 2502-9.
13. Fossati, P. 2012. Neural Correlates of Emotion Processing: from Emotional to Social
Brain. Eur. Neuropsychopharmacol., 22(3), S487-91.
14. Garber, P.A. & Leigh, S.R. 1997. Ontogenetic Variation in Small-Bodied New World
Primates: Implications for Patterns of Reproduction and Infant Care. Folia
Primatologica, 68, 1-22.
15. Green, L., Fein, D., Modahl, C., Feinstein, C., Waterhouse, L. & Morris, M. 2001.
Oxytocin and Autistic Disorder: Alterations in Peptide Forms. Biol. Psychiatry, 50, 609-
13.
16. Guastalla, A.J., Mitchell, P.B. & Dadds, M.R. 2008. Oxytocin Increases Gaze to the Eye
Region of Human Faces. Biological Psychiatry, 63, 305.
17. Kennedy, D.P. & Adolphs, R. 2012. The Social Brain in Psychiatric and Neurological
Disorders. Trends Cogn. Sci., 16(11), 559-72.
18. Kirsch, P., Esslinger, C., Chen, Q., Mier, D., Lis, S., Siddhanti, S., Gruppe, H., Mattay, V.,
S., Gallhofer, B., Meyer-Lindenberg, A. 2005. Journal of Neuroscience, 25(49), 11489-
11493.
19. Lim, M.M. & Young, L.J. 2006. Neuropeptidergic Regulation of Affiliative Behavior and
Social Bonding in Animals. Hormones and Behavior, 50, 506-517.
20. Ma, W.J., Hashii, M., Munesue, T., Havashi, K., Yagi, K., Yamagishi, M., Higashida, H.
& Yokoyama, S. 2013. Non-synonymous Single-nucleotide Variations of the Human
Oxytocin Receptor Gene and Autism Spectrum Disorders: A Case-control Study in a
Japanese Population and Functional Analysis. Mol. Autism, 4(1), 22.
21. Amodio, D.M. & Frith, C.D. 2006. Meeting of Minds: The Median Frontal Cortex and
Social Cognition. Nat. Rev. Neuroscience, 7(4), 268-277.
22. Mikolajczak, M., Pinon, N., Lane, A., de Timary, P. & Luminet, O. 2010. Oxytocin not
only Increases Trust when Money is at Stake, but also when Confidential Information is
in the Balance. Biol. Psychol., 85(1), 182-4.
23. Miller, M., Bales, K., L., Tayler, S., L., Yoon, J., Hostetler, C.M., Carter, C., S. &
Solomon, M. 2013. Oxytocin and Vasopressin in Children and Adolescents with Autism
Spectrum Disorders: Sex Differences and Associations with Symptoms. Autism Res.,
6(2), 91-102.

21
24. Modi, M.E. & Young, L.J. 2012. The Oxytocin System in Drug Discovery for Autism:
Animal Models and Novel Therapeutic Strategies. Hormones and Behavior, 61, 340-350.
25. Murakami, G., Hunter, R.G., Fontaine, C., Ribiero, A. & Pfaff, D. 2011. Relationships
among Estrogen Receptor, Oxytocin and Vasopressin Gene Expression and Social
Interaction in Male Mice. European Journal of Neuroscience, 34(3), 469-477.
26. Petrides, M. & Pandya, D.N. 1999. Dorsolateral Prefrontal Cortex: Comparative
Cytoarchitectonic Analysis in the Human and Macaque Brain and Corticocortical
Connection Patterns. Eur. J. Neurosci., 11, 1011-1036.
27. Rosenblum, L.A., Smith, E.L., Altemus, M., Scharf, B.A., Owens, M.J., Nemeroff, C.B.,
Gorman, J.M. & Coplan, J.D. 2002. Differing Concentrations of Corticotropin-releasing
Factor and Oxytocin in the Cerebrospinal Fluid of Bonnet and Pigtail Macaques.
Psychoneuroendocrinology, 27, 651-660.
28. Sinha, A. 2005. Not in Their Genes: Phenotypic Flexibility, Behavioral Traditions and
Cultural Evolution in Wild Bonnet Macaques. Journal of Biosciences, 30, 51-64.
29. Sofroniew, M.V. 1983. Morphology of Vasopressin and Oxytocin Neurons and their
Central and Vascular Projections. Prog. Brain Res., 60, 101-114.
30. Sussman, R.W., Garber, P.A. & Cheverud, J.M. 2005. The Importance of Cooperation and
Affiliation in the Evolution of Primate Sociality. American Journal of Physical
Anthropology, 128, 84-97.
31. Veenema, A.H., Bredewold, R. & De Vries, G.J. 2013. Sex-specific Modulation of
Juvenile Social Play by Vasopressin. Psychoneuroendocrinology, (July 6) [Epub ahead of
print].
32. Walum, H., Lichtenstein, P., Neiderhiser, J.M., Reiss, D., Ganiban, J.M., Spotts, E.L.,
Pederson, N.L., Anckarsater, H., Larsson, H. & Westberg, L. 2012. Variation in the
Oxytocin Receptor Gene is Associated with Pair-Bonding and Social Behavior. Biol.
Psychiatry, 71(5), 419-426.
33. Witt, D.M., Winslow, J.T. & Insel, T.R. 1992. Enhanced Social Interactions in Rats
following Chronic, centrally infused Oxytocin. Pharmacology, Biochemistry & Behavior,
43, 855-861.
34. Wong, V.C. & Chen, W., X. 2010. Randomized Controlled Trail of Electro-acupuncture
for Autism Spectrum Disorder. Alternative Medicine Review, 15, 136-146.
35. Yang, J., Yang, Y., Chen, J., M., Liu, W., Y., Wang, C., H. & Lin, B., C. 2007. The Effect

22
of Oxytocin on Acupuncture Analgesia in The Rat. Neuropeptides, 41, 285-292.
36. Young, L.J., Nilsen, R., Waymire, K.G., MacGregor, G.R. & Insel, T.R. 1999. Increased
Affiliative Response to Vasopressin in Mice Expressing the V1a Receptor from a
Monogamous Vole. Nature, 400, 766-768.
37. Young, L.J. & Wang, Z. 2004. The neurobiology of pair bonding. Nature Neuroscience, 7,
1048-1054.
38. Zhang, R., Jia, M.X., Zhang, J.S., Xu, X.J., Shou, X.J., Zhang, Z.T., Li, L., Li, N., Han,
S.P. & Han J.S. 2012. Transcutaneous Electrical Acupoint Stimulation in Children with
Autism and its Impact on Plasma Levels of Arginine-Vasopressin and Oxytocin: A
Prospective Single-Blinded Controlled Study. Res. Dev. Disabil., 33(4), 1136-46.
39. Zink, C.F., Kempf, L., Hakimi, S., Rainey, C.A., Stein, J.L. & Meyer Lindenberg, A.
2011. Vasopressin Modulates Social Recognition-Related Activity in the Left
Temporoparietal Junction in Humans. Transl. Psychiatry, 1(4), e3.
40. Zink, C.F. & Meyer-Lindenberg, A. 2012. Human Neuroimaging of Oxytocin and
Vasopressin in Social Cognition. Hormones and Behavior, 61, 400-09.
41. Zink, C.F., Stein, J.L., Kempf, L., Hakimi, S. & Meyer-Lindenberg, A. 2010. Vasopressin
Modulates Medial Prefrontal Cortex-Amygdala Circuity During Emotion Processing in
Humans. Journal of Neuroscience, 30(20), 7017-7022.