Embed Size (px)
Citation preview

In: Principles of Free Radical Biomedicine. Volume 1 ISBN: 978-1-61209-773-2 Editors: K. Pantopoulos, H. M. Schipper, pp. © 2011 Nova Science Publishers, Inc.
Chapter II
Oxygen Radicals and Related Species
Ohara Augusto and Sayuri Miyamoto Departamento de Bioquímica, Instituto de Química, Universidade de São Paulo, Caixa
Postal 26077, 05513-970, São Paulo, SP, Brazil
1. Introduction1: Oxygen and its Metabolites Imprinted the Evolution of Life
As discussed in Chapter 1, around three billion years ago, life on Earth consisted of
anaerobic microbes subsisting on the energy provided by cycles of electron transfer between prevalent populations of electron donors and acceptors. Potential donors were molecular hydrogen (H2), hydrogen sulfide (H2S) and methane (CH4), whose electrons were transferred to acceptors such as carbon dioxide (CO2) and, to a lesser extent, sulfate (SO4
2-). The biggest electron-donor pool, water (H2O), remained biologically inaccessible until the first photosynthetic organisms evolved between ~3.2 and 2.4 billion years ago. Employing the energy of the sun and H2O as the reductant to fuel metabolism, these ancient organisms produced molecular oxygen (O2) as a waste product. Over a relatively short period of geologic time, around 100 million years, O2 atmospheric levels increased, indicating that the energy-transducing process of photosynthesis was replacing the old metabolic networks. As a consequence, the life forms that evolved in anaerobic conditions had to adapt to O2, hide or become extinct. With O2 overwhelming the Earth’s reducing atmosphere, these primitive microbes became restricted to hypoxic or anaerobic environments. However, some of them were able to escape and adapt to the oxidizing atmosphere. They either modified parts of their metabolism, or formed symbiotic associations that permitted the coupling of nutrient oxidation to reduction of O2 back to H2O. This respiratory pathway was far more efficient in extracting energy from nutrients, but it came with costs. The triplet ground state of O2 is prone to one-electron transfers, which yield species that are toxic to life (see section 6.3). This
1 Due to space constraints, we cited only a small fraction of the original investigations that contributed to the
current understanding of the multiple roles of radicals/oxidants in Biology. Whenever possible, we cited reviews that referred to the original papers.

Ohara Augusto and Sayuri Miyamoto 2
pushed the development of mechanisms to protect the genetic investment in the metabolic pathway that drive the “H2O-H2O” cycle upon which most life on Earth would come to depend. The protective pathways evolved further, driving the evolution of single-cell eukaryotes, from which all current plants and animals on Earth descend [1, 2].
Adaptation to O2, however, went beyond the creation of redox energy couples that allowed complex life forms to evolve. As a recent meta-metabolomic analysis demonstrated, it also pushed an evolutionary explosion of random, alternative and novel metabolic networks yielding a wide variety of gene products that increased fitness of the organisms. Although we have a long way to go to understand how life evolved, it is clear that O2 and its derived metabolites imprinted the evolution of complex life forms [1, 2]. Thus, the participation of oxygen-derived metabolites in cell regulatory and signaling pathways could be anticipated [2]. Nevertheless, until recently, oxygen-derived metabolites were mostly considered for their involvement in cell damaging mechanisms [3, 4].
In this Chapter, the chemical properties of O2 that cause its propensity to produce free radicals and oxidants as metabolites will be presented. These species, known by the general term reactive oxygen species (ROS) will be historically contextualized. The discoveries pointing to their roles as mediators of physiological and pathophysiological circuits will be summarized. Finally, the reactivity of specific ROS towards biomolecules will be presented in the context of health and disease - topics that will be extended in the other Chapters of this book.
Figure 1. Electronic distribution of the electrons of two oxygen atoms (8O) in atomic orbitals, which combine in molecular orbitals to form molecular oxygen (O2) (left side). O2 contains two unpaired electrons. This is because in distributing its electrons in molecular orbitals, the last two electrons have two molecular orbitals of the same energy to occupy (2*) and each electron occupies one (Hund´s rule). Because the presence of two electrons in antibonding orbitals (2*) energetically cancels out one of the bonding orbitals (2), the two oxygen atoms in O2 are bound by two covalent bonds (3 bond-occupied orbitals minus 2 antibound semi-occupied orbitals). B. Electronic distribution of the electrons of one nitrogen atom (7N) and one oxygen atom (8O) in atomic orbitals which combine in molecular orbitals to form nitric oxide (NO●) (right side). Distribution of the 17 electrons of NO● in molecular orbitals leaves an unpaired electron in the 2* orbital making it a free radical in the ground state. Nitrogen and oxygen in NO● are bound by 2.5 covalent bonds (3 bond-occupied orbitals minus 0.5 antibond orbitals).

Oxygen Radicals and Related Species 3
2. Molecular Oxygen, a Sluggish Oxidant Producer of Reactive Species
Long before the formation of molecules and ions was conceptualized by quantum
mechanics, the discoverers of molecular oxygen in the late 18th century - Priestley, Scheele and Lavoisier - reported its beneficial and toxic effects on living organisms. These opposing effects result from the properties of molecular oxygen, which is formed by the combination of two oxygen atoms in covalent bonds (O2). The characteristics of a covalent bond, such as strength, length and direction, depend on the occupied molecular orbitals. O2 is bound by two covalent bonds (Figure 1), and requires 402 kJ/mol to break into two oxygen atoms (O). This is roughly the amount of energy required to bring 1 liter of H2O to boiling, indicating that the bond strength between the two O atoms in O2 is strong. In other words, O2 is a considerably stable molecule, as attested by our daily experience. Most covalent bonded molecules are equally stable, possessing energy bonds from 150 to 950 kJ/mol.
However, O2 has its peculiarities. Although stable, a spark triggers its reaction with fuels (combustion) liberating energy to move our cars. Likewise, the oxidation of the foods we eat (respiration) produces energy to sustain our lives. Indeed, most reactions of O2 are slow but, once initiated or catalyzed, liberate a considerable amount of energy. In other words, reactions of molecular oxygen are favored by thermodynamics but not by kinetics. This is because O2 has two unpaired electrons in the ground state, which makes it a triplet molecule (Figure 1A). In contrast, most organic molecules possess all electrons paired, being singlet in the ground state. To react with them, O2 has to receive a pair of electrons, but this requires spin inversion, an event that is prohibited by the spin conservation rule (Figure 2). According to its structure, O2 tends to receive electrons by one-electron steps, reacting rapidly only with species capable of one-electron transfer. Examples of such species are biomolecules that produce stable free radicals, such as flavin enzymes and coenzymes, and species that contain unpaired electrons, such as other free radicals and transition metal ions (Figure 2). Thus, it is not surprising that iron artifacts are rapidly turned into scrap and that most enzymes that catalyze biological oxidations contain transition metal ions and/or flavin coenzymes in their active sites. O2, free radicals and transition metal ions are closely related. In fact, O2 and transition metal ions are also free radicals because they contain unpaired electrons. Nevertheless, ‘free radical’ is more frequently defined as a species (molecule, cation or anion) that contains one unpaired electron. Only in these cases is the unpaired electron denoted by a superscript dot to the right preceding any charge in the radical formula [5]. The term free radical is historical because organic radicals, which are used to refer to the chemical groups in a molecule, are bound as opposed to free. Currently, both terms free radical and radical are used in the literature and understood by their contexts. A classical free radical in the ground state is nitric oxide (NO) (Figure 1B), whose role in signal transduction pathways was established in the last decade of the 20th century [3, 4, 6].

Ohara Augusto and Sayuri Miyamoto 4
Figure 2. Schematic representation of the main reactions of O2. Due to its unpaired electrons, O2 does not react with most organic molecules because they have paired electrons. Such reactions are extremely slow because they would require spin inversion which is prohibited by the spin conservation rule. O2 reacts rapidly with molecules that produce stable radicals by one-electron transfer (BM●+) or have unpaired electrons, such as free radicals (R●) and transition metal ions (Mn+). If O2 receives energy (from light, excited molecules, etc.), it is excited to singlet oxygens (1gO2 or 1g
+O2) that are more reactive towards biomolecules because there is no spin restriction.
3. Free Radicals in Biology: An Historical Account In the first half of the 20th century, organic radicals were recognized as intermediates of
chemical reactions, and many of their properties were studied and explored in polymerization processes. Most investigators considered radicals irrelevant to biology, except for some pioneers. For instance, the known propensity of O2 to react by one-electron steps (Figure 2) led some investigators, such as Otto Warburg and Leonor Michaelis, to propose that during respiration O2 would produce one-electron intermediates, such as superoxide (O2
-) and the hydroxyl radical (HO) (Reaction 1).
Evidence that O2 produced radicals in animals was first provided by Rebeca Gerschman
and co-workers in 1954 while studying the toxic effects of high O2 pressures and X-ray irradiation in mice [7]. In the resulting paper, they concluded: “it would appear that irradiation and O2 poisoning produce some of their lethal effects through at least one common mechanism, possibly that of the formation of free radicals”. In addition, they showed that
O2 O2∙ H2O2 HO∙ H2O
e e /2H+ e /H+ H2O e /H+
(Reaction 1)
O2 + 4H+ + 4e 2 H2O

Oxygen Radicals and Related Species 5
protectors against irradiation, such as glutathione and ethanol, protected against oxygen toxicity, or as currently defined, they acted as antioxidants. Another pioneer was Denham Harman, who proposed in 1954 that aging was a consequence of free radical attack on biomolecules (see Chapter 35).
These views remained largely ignored up to 1970, while the properties of free radicals were extensively studied by radiation chemists. The employment of biochemical approaches to explore fundamental biological questions, such as the molecular basis of life, provided a turning point. In this background, McCord and Fridovich discovered the enzyme superoxide dismutase (SOD) in 1969 [8] (see also Chapter 17). They showed for the first time that mammals possess an enzyme whose function is to catalyze the dismutation of O2
●- (k= 1.6 x 109 M-1 s-1), which also occurs spontaneously at a slower rate (k= 5 x 105 M-1 s-1) (Reaction 2). Dismutation is a typical reaction of most free radicals; two molecules of the same radical (such as O2
●-) react in an electron transfer process to produce oxidized (O2) and reduced (H2O2) products.
The fact that evolution preserved an enzyme whose function is to dismutate a free radical
indicated that radicals were constantly produced during normal metabolic processes. This fundamental discovery inaugurated a new area of research, “free radicals in biology”, which continues to expand.
The discovery of SOD provided evidence that cells generate O2●-. Soon thereafter, it
became clear that during biological processes cells also produce additional related intermediates, generically termed reactive oxygen species (ROS), such as H2O2, HO● and hypochlorous acid (HOCl). ROS were shown to be produced during mitochondrial respiration, phagocyte-mediated killing of pathogens, and xenobiotic metabolism. These investigations brought new concepts into pathophysiology. One was the importance of reactions catalyzed by transition metal ions (Fenton chemistry; Chapter 5) to produce HO●, an extremely potent oxidant, from the less reactive O2
●- (Reactions 3 and 4).
Another was the notion that if not eliminated by the antioxidant defenses, ROS would
attack DNA, lipids and proteins (Chapters 6-9) causing cell and tissue injury. The most influential one was the concept of oxidative stress defined as an imbalance between free radicals/oxidants and antioxidants in favor of the former [9]. As a consequence, radicals and oxidants became associated with most human diseases and many intervention studies were designed to examine the effects of antioxidant vitamins on diseases, particularly cardiovascular and neurological.
O2∙ + O2
∙ + 2 H+ H2O2 + O2
(Reaction 2)
H2O2 + Fe2+ Fe3+ + HO∙ + HO
(Reactions 3 and 4)
Fe 3+ + O2∙ Fe2+ + O2

Ohara Augusto and Sayuri Miyamoto 6
By the last decade of the 20th century, however, most data collected from intervention studies with antioxidants were inconclusive, suggesting that the classical view of oxidative stress required revision [4, 6]. This was reinforced by the discovery of NO● (Figure 1), a gaseous free radical, as a major autocrine and paracrine mediator of vascular relaxation, immune regulation and many other physiological effects. In mammals, NO● is mainly produced from arginine oxidation, catalyzed by a family of enzymes, the nitric oxide synthases (NOS) (Reaction 5) [10, 11].
Soon thereafter, it was demonstrated that O2
●- reacts rapidly with NO● (Reaction 6) and not only regulates NO● bioavailability but also leads to potent oxidants such as peroxynitrite (ONOO-/ONOOH)†, HO●, nitrogen dioxide (NO2
●) and carbonate radical (CO3●-) (Reactions
7 and 8) [12-14] (see also Chapter 3).
Figure 3. Schematic representation of the roles of radicals and oxidants as mediators of physiological and pathophysiological circuits. Present evidence indicates that free radicals and oxidants are constantly produced in vivo from exogenous and endogenous sources and are, directly or indirectly, derived from O2. The main endogenous sources of free radicals/oxidants are: the mitochondrial electron transport chain and enzymatic reactions catalyzed by nitric oxide synthases (NOS), NADPH oxidases (NOX) (from phagocytes and other cell types), xanthine oxidase (XO) and hemeperoxidase enzymes, such as myeloperoxidase (MPO). Organisms evolved enzymatic antioxidant defenses and the capability to use antioxidants from the diet to control the physiological levels of free radicals/oxidants. High levels of some oxidants may cause dysfunction because they oxidize biomolecules that lead to cell and tissue injury if not repaired by the evolved repair systems. Low levels of certain free radicals/oxidants also compromise physiological functions that evolved to depend on them. On the other hand, transient and small increases of some oxidants trigger redox-sensitive signaling pathways.
† The term peroxynitrite refers to the sum of peroxynitrite anion (ONOO-, oxoperoxonitrate (-1)) and
peroxynitrous acid (ONOOH, hydrogen oxoperoxonitrate) unless otherwise specified. Other abbreviations are defined in the text.
Arginine + NADPH + O2 NO∙ + Citruline + NADP+ + H2ONOS
(Reaction 5)

Oxygen Radicals and Related Species 7
Furthermore, in the same period, several lines of evidence indicated that H2O2 can act as
a second messenger for receptor agonists, such as growth factor and hormones, prompting the development of the concept of redox signaling. This involves cellular signal transduction networks in which the integrative element is a series of interconnected electron transfer reactions. By exerting second messenger effects, oxidants can regulate major cellular pathways [15-17]. Such function is likely rooted in the oxygen-dependent evolution of complex life forms. Our current knowledge about redox signaling mechanisms is still in its infancy.
For decades, free radical research focused on understanding the formation of reactive species in vivo, and elucidating how proteins, lipids and DNA are damaged by them, resulting in cellular injury and disease. The complete picture is more complex, though. Exceeding levels of reactive species may cause dysfunction, but low levels of certain oxidants also compromise physiological functions that evolved to depend on them, such as microbicidal activity, proliferative responses and vasodilation. Moreover, transient and small increases in some oxidants trigger redox-sensitive signaling pathways. Thus, radicals and oxidants are presently considered to control signaling circuits involved in physiological and pathological responses (Figure 3) [4, 6]. Unraveling these interrelated processes requires a better understanding of cellular oxidative mechanisms, including the identification of the involved oxidants, the pathways regulating their generation, and their molecular targets. Further advances in the field will depend on an interdisciplinary effort that combines rigorous chemical thinking and tools with relevant biological data and insights [16, 17].
4. Oxygen Radicals and Related Species: General Aspects of the Reactivity of One-
and Two-electron Oxidants Although discrimination between different biologically relevant oxidants became critical,
the difficulties in detecting these short-lived species in biological media stimulate the use of general terms, such as ROS, in the literature. As previously noted by Christine Winterbourn [16, 17], terms such as ROS and antioxidants are appropriate to refer to general classes of compounds but are counterproductive for understanding mechanisms. This is because not all antioxidants act by the same mechanism, and the species encompassed by ROS have widely different reactivities. The same is true for another frequently used general term, RNS (reactive
O2∙ + NO∙ ONOO
ONOO + H+ ONOOH0.7 NO3
+ 0.7 H+
ONOO + CO2 [ONOOCO2]
0.65 NO3 + 0.65 CO2
0.35 NO2∙ + 0.35 CO3
∙
(Reactions 6-8)
pKa= 6.4
k= 1.9 x 109 M-1 s-1
k= 2.6 x 104 M-1 s-1
0.3 HO∙ + 0.3 NO2∙
k= 0,17 s-1

Ohara Augusto and Sayuri Miyamoto 8
nitrogen species). RNS comprise reactive metabolites derived from NO●, such as peroxynitrite and NO2
● (Chapter 3). However, RNS and ROS are closely interrelated (see, for instance, Reactions 6-8), and only one general term may be more appropriate. Indeed, radicals typically react rapidly with other species containing unpaired electrons (Figure 2), making it often difficult to establish which oxidant is involved in a particular physiological process. To make progress in this direction, it is necessary to recognize the widely different characteristics of the species encompassed by the term ROS (and RNS). Some are radicals and undergo one-electron reactions, whereas others are non-radicals and promote two-electron oxidations. Some are strong and others weak oxidants, and the second-order rate constant of their reactions with biomolecules vary considerably [16, 17]. For instance, the second order rate constant of some physiologically relevant oxidants with glutathione (GSH) varies from undetectable to diffusion-controlled (k ~ 109-1010 M-1 s-1 in aqueous media) (Table 1). The tripeptide GSH (-L-glutamyl-L-cysteinylglycine), which is present at millimolar concentrations in most cell types, is a major intracellular reductant because of its cysteine thiol group (-SH) (Chapter 13).
In this book, oxygen radicals and related species (this Chapter), nitrogen radicals and related species (Chapter 3) and sulfur radicals and related species (Chapter 4) are discussed separately. Nevertheless, it is useful to stress some general aspects concerning the reactivity of biologically relevant oxidants. The oxidizing strength of a radical is given by its one-electron reduction potential (Eo), with higher values corresponding to more potent oxidants. This is usually reflected in the rates at which radicals react because of the low activation energy of radical reactions (Table 1). Thus, it is not surprising that HO● reacts with most biomolecules with second-order rate constants close to the diffusion limit. That is, the reaction occurs as fast as the reagents encounter each other. As one-electron oxidants, radicals favor radical chain reactions as occurs during lipid peroxidation (Chapter 7). A remarkable exception is NO●, which rapidly interacts with other radicals interrupting radical chain reactions (Reaction 9; see also Chapter 8).
Although low NO● levels inhibit lipid peroxidation, the radical can produce potent
oxidants by reacting with O2●- (Reaction 5) and O2 (Reaction 10).
In the case of two-electron oxidants, their reduction potential also determines their
oxidizing strength; however, kinetics determines their reactivity because of the high activation energy involved in these oxidations. For instance, H2O2 has a higher reduction potential than peroxynitrous acid (ONOOH) but reactions of H2O2 have higher activation energies and slower rates (Table 1).
NO∙ + ROO∙ NOOOR
k~ 109 M-1 s-1
(Reaction 9)
2 NO∙ + O2 2 NO2∙
k= 2 106 M-2 s-1
(Reaction 10)
Oxygen Radicals and Related Species 9
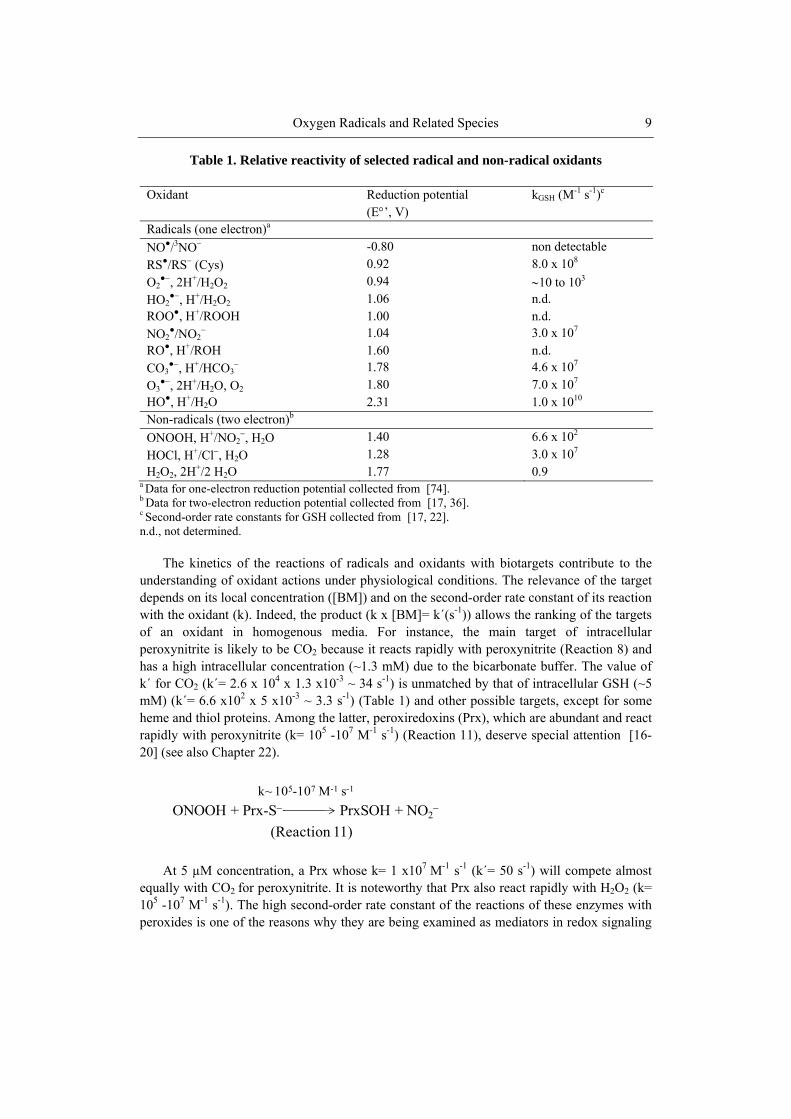
Table 1. Relative reactivity of selected radical and non-radical oxidants
Oxidant Reduction potential (E’, V)
kGSH (M-1 s-1)c
Radicals (one electron)a
NO●/3NO -0.80 non detectable
RS●/RS (Cys) 0.92 8.0 x 108
O2●, 2H+/H2O2 0.94 10 to 103
HO2●, H+/H2O2 1.06 n.d.
ROO●, H+/ROOH 1.00 n.d. NO2
●/NO2 1.04 3.0 x 107
RO●, H+/ROH 1.60 n.d. CO3
●, H+/HCO3 1.78 4.6 x 107
O3●, 2H+/H2O, O2 1.80 7.0 x 107
HO●, H+/H2O 2.31 1.0 x 1010 Non-radicals (two electron)b
ONOOH, H+/NO2, H2O 1.40 6.6 x 102
HOCl, H+/Cl, H2O 1.28 3.0 x 107 H2O2, 2H+/2 H2O 1.77 0.9
a Data for one-electron reduction potential collected from [74]. b Data for two-electron reduction potential collected from [17, 36]. c Second-order rate constants for GSH collected from [17, 22]. n.d., not determined.
The kinetics of the reactions of radicals and oxidants with biotargets contribute to the
understanding of oxidant actions under physiological conditions. The relevance of the target depends on its local concentration ([BM]) and on the second-order rate constant of its reaction with the oxidant (k). Indeed, the product (k x [BM]= k´(s-1)) allows the ranking of the targets of an oxidant in homogenous media. For instance, the main target of intracellular peroxynitrite is likely to be CO2 because it reacts rapidly with peroxynitrite (Reaction 8) and has a high intracellular concentration (~1.3 mM) due to the bicarbonate buffer. The value of k´ for CO2 (k´= 2.6 x 104 x 1.3 x10-3 ~ 34 s-1) is unmatched by that of intracellular GSH (~5 mM) (k´= 6.6 x102 x 5 x10-3 ~ 3.3 s-1) (Table 1) and other possible targets, except for some heme and thiol proteins. Among the latter, peroxiredoxins (Prx), which are abundant and react rapidly with peroxynitrite (k= 105 -107 M-1 s-1) (Reaction 11), deserve special attention [16-20] (see also Chapter 22).
At 5 µM concentration, a Prx whose k= 1 x107 M-1 s-1 (k´= 50 s-1) will compete almost
equally with CO2 for peroxynitrite. It is noteworthy that Prx also react rapidly with H2O2 (k= 105 -107 M-1 s-1). The high second-order rate constant of the reactions of these enzymes with peroxides is one of the reasons why they are being examined as mediators in redox signaling
ONOOH + Prx-S PrxSOH + NO2
k~ 105-107 M-1 s-1
(Reaction 11)
Ohara Augusto and Sayuri Miyamoto 10
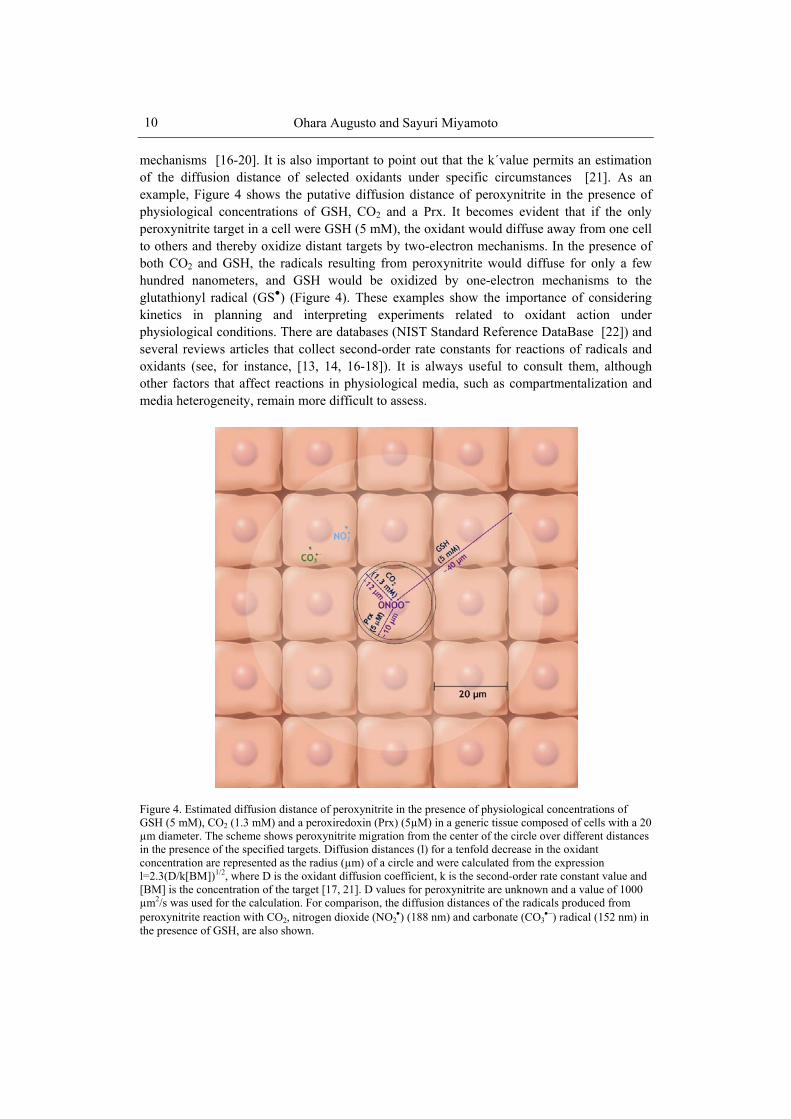
mechanisms [16-20]. It is also important to point out that the k´value permits an estimation of the diffusion distance of selected oxidants under specific circumstances [21]. As an example, Figure 4 shows the putative diffusion distance of peroxynitrite in the presence of physiological concentrations of GSH, CO2 and a Prx. It becomes evident that if the only peroxynitrite target in a cell were GSH (5 mM), the oxidant would diffuse away from one cell to others and thereby oxidize distant targets by two-electron mechanisms. In the presence of both CO2 and GSH, the radicals resulting from peroxynitrite would diffuse for only a few hundred nanometers, and GSH would be oxidized by one-electron mechanisms to the glutathionyl radical (GS●) (Figure 4). These examples show the importance of considering kinetics in planning and interpreting experiments related to oxidant action under physiological conditions. There are databases (NIST Standard Reference DataBase [22]) and several reviews articles that collect second-order rate constants for reactions of radicals and oxidants (see, for instance, [13, 14, 16-18]). It is always useful to consult them, although other factors that affect reactions in physiological media, such as compartmentalization and media heterogeneity, remain more difficult to assess.
Figure 4. Estimated diffusion distance of peroxynitrite in the presence of physiological concentrations of GSH (5 mM), CO2 (1.3 mM) and a peroxiredoxin (Prx) (5µM) in a generic tissue composed of cells with a 20 µm diameter. The scheme shows peroxynitrite migration from the center of the circle over different distances in the presence of the specified targets. Diffusion distances (l) for a tenfold decrease in the oxidant concentration are represented as the radius (µm) of a circle and were calculated from the expression l=2.3(D/k[BM])1/2, where D is the oxidant diffusion coefficient, k is the second-order rate constant value and [BM] is the concentration of the target [17, 21]. D values for peroxynitrite are unknown and a value of 1000 µm2/s was used for the calculation. For comparison, the diffusion distances of the radicals produced from peroxynitrite reaction with CO2, nitrogen dioxide (NO2
) (188 nm) and carbonate (CO3) radical (152 nm) in
the presence of GSH, are also shown.

Oxygen Radicals and Related Species 11
5. Chemistry of Biologically Relevant Oxygen-Derived Radicals
The biologically relevant oxygen-derived radicals include O2
●- and its conjugated acid (HO2
●), HO●, CO3●-, peroxyl (ROO●) and alkoxyl (RO●) radicals. The sources, properties and
main reactions of these species in physiological environments are discussed below.
5.1. Superoxide (O2●-) and Hydroperoxyl (HO2
●-) Radical Superoxide (O2
●-) is formed by the one-electron reduction of O2 (Reaction 1). In biological systems it is produced by enzymatic reactions catalyzed by oxidases, such as NADPH oxidases (NOX) and xanthine oxidase (XO), and non-enzymatically by redox active compounds, such as the semi-ubiquinones of the mitochondrial electron transport chain. O2
●
can act as oxidant or reductant, and the dismutation reaction is an example of this double action (Reaction 2). The second-order rate constant of the dismutation reaction is higher at acidic pH due to the increase in the concentration of the hydroperoxyl/perhydroxyl radical (HO2
●) (pKa = 4.8) in equilibrium with O2●- (Reaction 12). The determined second order rate
constant value for the spontaneous dismutation varies from 102 M-1 s-1 at pH 11 to 5 x 105 M-1 s-1 at pH 7. As noted above, the dismutation reaction catalyzed by SOD is much faster (k= 1.6 x 109 M-1 s-1).
O2
●- reactivity towards a variety of organic and inorganic targets has been studied. Despite its moderately high reduction potential (0.94 V) (Table 1), its reactivity with non-radical targets is limited [23]. However, O2
●-may induce harmful effects by reacting with radicals, such as NO● (Reaction 6), and species containing transition metal ions. For instance, O2
●- is highly reactive towards iron-sulfur ([Fe-S]) clusters [24] (see also Chapter 31), with the reaction occurring at near diffusion-limited rates (Reaction 13). O2
●- causes one-electron oxidation of the [4Fe-4S] clusters to form H2O2 and an unstable intermediate that decomposes losing iron (II). Relevantly, this free iron ion can react with H2O2 and catalyze the production of HO● by Fenton chemistry (Reactions 3 and 4).
Radical-radical reactions of O2
●- typically occur at near diffusion-controlled rates [25]. Among them, the reaction of O2
●- with NO· has received special attention due to the generation of peroxynitrite (Reaction 6), a strong biological oxidant (Chapter 3). Reactions of O2
●- with radicals formed on aromatic amino acids, such as the tyrosyl radical (Tyr●), have
O2∙ + H+ HO2
∙
(Reaction 12)
[4Fe-4S]2+ + O2∙+ 2 H+ + Fe2H2O2 +[4Fe-4S]3+ [3Fe-4S]+
k > 109 M-1 s-1
(Reaction 13)

Ohara Augusto and Sayuri Miyamoto 12
also been studied. The second-order rate constant of the reaction with Tyr● is about 3-fold higher than that of Tyr● dimerization to dityrosine [25]. Upon reaction with O2
●- the Tyr● radical can be repaired back to Tyr or be transformed into Tyr hydroperoxides depending on the position of the Tyr residue in the polypeptide chain.
Reactions of O2●- with biologically important thiol compounds have been investigated
[26]. Literature data on the second-order rate constant for these reactions vary from 10 to 105 M-1 s-1. This variation has been attributed to the use of inappropriate assays and the complexity of the chain reactions involving sulfinyl and thiyl radicals [27]. According to Winterbourn and colleagues, the best estimates are in the range of 30 to 103 M-1 s-1. These values indicate that biothiols do not react rapidly enough with O2
●- to compete with SOD. Therefore, proteins containing [Fe-S] clusters are likely to be O2
●- sensors in redox signaling (Reaction 13). In agreement, bacteria rely on the SoxR transcription factor, which contains [Fe-S] clusters, to upregulate resistance genes in response to sublethal O2
●- levels.
5.2. Hydroxyl Radical (HO●) HO● is the strongest oxidant produced in biological systems (E’ =2.31 V) (Table 1). It
reacts very rapidly and indiscriminately with most biological targets present at its site of formation (see, for comparison, Figure 4). Indeed, the rate constants for its reactions with most of biomolecules, including, lipids, proteins, carbohydrates and DNA are very close to the diffusion-controlled limit. HO● can be generated in vivo by four major processes: ionizing-radiation (e.g. UV, X-rays, -rays); transition metal ion-catalyzed reactions (Reaction 4); proton-catalyzed decomposition of peroxynitrite (Reaction 6); and decomposition of ozone (O3) (see section 6.5). Irradiation with high-energy radiations generates HO● by homolytic fission of water molecules (Reaction 14). HO● produced by this process is thought to be responsible for DNA damage and tumor development (e.g. skin cancer), while irradiation can also be employed for targeted killing of tumor cells (e.g. radiotherapy).
Reactions of the HO● can be classified into three main types: hydrogen abstraction,
addition and electron transfer reactions. HO● is extremely fast in abstracting hydrogen atoms from organic compounds, especially from those that are weakly bound. The reaction produces H2O and a carbon-centered radical (RC●), which, in the presence of O2 generates peroxyl radicals (ROO●), such as in the case of carbon-centered lipid radicals. In the absence of O2, a covalent bond can be formed between two RC● producing a crosslink. An example is the reaction between two Tyr● to produce dityrosine. Reactions of HO● with aromatic compounds usually involve the addition of the radical to produce hydroxylated radical adducts. An important example is the addition of HO● to the guanine moiety of DNA/RNA to produce 8-hydroxyguanine and 2,6-diamino-4-hydroxy-5-formamidopyrimidine (Chapter 9). Reactions of HO● with anions produce radicals by electron transfer mechanisms. For instance, the
H2O HO∙ + H∙
(Reaction 14)

Oxygen Radicals and Related Species 13
abundant Cl- and the NO●-metabolite, NO2-, are oxidized to chlorine (Cl●) and NO2
●, respectively (Reactions 15 and 16).
5.3. Carbonate Radical (CO3●-)
Although less oxidizing than HO●, CO3
●- (Eo´ =1.78 V) (Table 1) is a very strong one-electron oxidant. Relatively recent data demonstrated that CO3
●- is an electrophilic oxygen-centered radical and a strong acid (pKa <0). Consequently, it will be negatively charged in all physiological environments, including those of acidic pH [13, 28]. The most recognized biological source of CO3
●- is the reaction between peroxynitrite and CO2 (Reaction 8) (Figure 4). CO3
●- is also produced by the peroxidase activity of SOD, by the turnover of XO and by transition metal-ion catalyzed decomposition of peroxymonocarbonate (HCO4
-) [13, 28]. The latter is an oxidant found in equilibrium in aqueous solutions of H2O2 and bicarbonate (HCO3
-
) (Reaction 17), whose potential participation in biological processes has been recently discussed [13, 28].
Most CO3
●- reactions are oxidations by both electron transfer and hydrogen abstraction mechanisms to produce radicals from the oxidized targets. Addition reactions of the carbonate radical to produce stable target adducts are virtually unknown in the literature [13, 28]. As a charged species, CO3
●- is an important oxidizing agent in aqueous environments where intracellular GSH is a particularly relevant target (Table 1). In addition, CO3
●- oxidizes solvent-exposed amino acid residues in proteins, particularly Tyr, Trp and Cys. Guanine moieties in nucleotides, nucleosides and nucleic acids are efficiently oxidized by CO3
●-. Antioxidants, such as ascorbic and uric acid, are also rapidly oxidized by CO3
●-. Because CO3
●- mostly oxidizes biomolecules to radicals, its effects are difficult to discriminate from those of other strong one-electron oxidants. This fact contributes to the limited attention that CO3
●- has received so far, despite bicarbonate being the main physiological buffer [13, 28].
5.4. Peroxyl(ROO●) and Alkoxyl (RO●) Radicals ROO● and RO● are moderately strong oxidants (E’ > 1.0 V) (Table 1). They can be
generated from organic hydroperoxide (ROOH) decomposition induced by heat or radiation
(Reaction 15 and 16)
k= 1 1010 M-1 s-1
k= 4.3 109 M-1 s-1
Cl + HO∙ Cl∙ + HO
NO2 + HO∙ NO2
∙ + HO
H2O2 + HCO3
(Reaction 17)
H2O +HCO4

Ohara Augusto and Sayuri Miyamoto 14
and by ROOH reaction with transition metal ions and other oxidants capable of abstracting hydrogen (Reactions 18 and 19). ROO● are also important intermediates in processes involving carbon-centered radicals, which react rapidly with O2 (k>109 M-1 s-1). In addition, biomolecule-derived ROO● and RO● can be generated from the oxidation of lipids, proteins and nucleic acids.
The reactivity of ROO● and RO● is influenced by the substituents at the -carbon. An
electron-withdrawing group increases the reactivity (for instance, chloroperoxyl radical, CCl3OO●), whereas electron-donating groups decreases it (for instance, phenoxyl radicals). Aromatic ROO● and RO●· tend to be less reactive because of unpaired electron delocalization. The reactions of ROO● and RO● with biomolecules often involve hydrogen-abstraction, which is facilitated in compounds containing weakly bound hydrogens. This is the case for lipids, thiols, and several chain-breaking antioxidants. Lipids are particularly susceptible to hydrogen abstraction and this reaction is the rate-limiting step in the propagation of lipid peroxidation chain reactions (Chapter 7). The second-order rate constant value of ROO●-mediated hydrogen abstraction from lipids is low (k < 102 M-1 s-1) [29, 30] and increases with the number of allylic or double-allylic hydrogens. For unsaturated fatty acids the second-order rate constant value decreases in the sequence 22:6>20:5>20:4>18:2>18:1. Notably, the second-order rate constant values of RO● reactions (k 106-107 M-1 s-1) [29] are about 4-5 orders of magnitude higher than those of ROO●. Both ROO● and RO● can undergo rapid monomolecular rearrangements or fragmentations that compete with hydrogen abstraction reactions. ROO● formed on aromatic rings and those with -carbon linked to hydroxy or amino groups can decompose to liberate O2
●- or HO2●. This type of reaction has been reported
for amino acids, such as lysine. ROO●· can also react with another ROO●· by the Russell mechanism, generating a ketone, an alcohol and singlet molecular oxygen (1O2) (see section 6.3) [31].
6. Chemistry of Biologically Relevant Non-radical Oxygen Species
Biologically relevant two-electron oxidants derived from oxygen include H2O2, HOCl
and related species, 1O2, biomolecule-derived hydroperoxides and ozone (O3). These species are discussed below with regard to main properties and reactions in physiological environments.
(Reaction 18 and 19)
ROOH + Fe2+ RO∙ + HO + Fe3+
ROOH + Fe3+ ROO∙ + H+ + Fe2+

Oxygen Radicals and Related Species 15
6.1. Hydrogen Peroxide (H2O2) Biological production of H2O2 can occur by chemical and photochemical processes and
by enzymatic reactions catalyzed by several oxidases, such as NOX, XO and monoamine oxidase (MAO). In addition, H2O2 is produced continuously from O2
●- dismutation, either spontaneous or catalyzed by SOD (Reaction 2). In physiological environments, H2O2 can be rapidly inactivated to H2O by seleno-, heme- and thiol- peroxidases. For instance, the second-order rate constant value of the reaction of H2O2 with glutathione peroxidases (GPx), catalases and Prx are ~108 M-1 s-1, ~106 M-1s-1 and 105 -107 M-1s-1, respectively. Thus, steady-state concentrations of H2O2 in cells and tissues are low, being estimated to be around 10-7 to 10-8 M.
H2O2 is a powerful two electron-oxidant (E = 1.77 V, pKa 11.6) (Table 1). However, its reactivity toward most of biological molecules is low because of the high activation energy of these oxidations. Most damaging effects of H2O2 in vivo are considered to be mediated either by transition metal ions or enzymes, such as heme-peroxidases. These processes generate secondary species, which are more reactive and include radicals, such as HO● and NO2
●, and non-radical species, such as HOCl and related species. Reaction of H2O2 with reduced transition metal ions, such as copper (I) and iron (II), leads to the generation of HO● (Reaction 4). This reaction has been studied for more than a century (Chapter 5). The second-order rate constant values of Fenton reactions are dependent on the metal-ligand and on the medium pH, ranging from 102 to 104 M-1 s-1 for iron ions.
In addition to being substrate for several heme-peroxidases, H2O2 can be consumed by other hemoproteins, such as hemoglobin, myoglobin and cytochrome c, in processes that oxidize the proteins. In phagocytic cells, myeloperoxidase (MPO) uses H2O2 mainly for the production of HOCl (Reactions 20 and 21). MPO has also been reported to use H2O2 to oxidize NO2
- to NO2● [32] and Tyr to Tyr● [33]. Thus, MPO can be an efficient mediator of
protein nitration, particularly at acidic pH [34]. MPO is released during phagocytosis and is thought to play an important role in microbial killing [35]. However, excessive MPO-mediated production of HOCl, NO2
● and other oxidants can cause host tissue injury, contributing to the development of several diseases [36, 37].
Among the physiological reactions of H2O2, the oxidation of biothiols is receiving
increasing attention in the literature [15, 16]. The second-order rate constant values of the oxidation of low molecular weight thiols (RSH) by H2O2 are relatively low (1-5 x 102 M-1 s-1). The value increases for thiols with low pKa [26], indicating that the effective substrate is the thiolate form (RS-). In fact, it is proposed that a nucleophilic attack of RS- on peroxide oxygen occurs to produce sulfenic acid (RSOH) (Reaction 22), which subsequently reacts with a second thiol to produce disulfide (RSSR) (Reaction 23).
k1
k -1
(Reaction 20 and 21)
MPO + H2O2 MPO-I + H2O
MPO + Cl MPO + HOClk2

Ohara Augusto and Sayuri Miyamoto 16
It is important to emphasize that some classes of proteins possess Cys residues whose
pKa values are in the range of ~4.0 to ~6.5, which are much lower than that of free Cys (pKa=8.4) or GSH (pKa=9.2). Examples of these proteins are the Prx, which react extremely rapid with H2O2 (k~105-107 M-1 s-1). Currently, it is accepted that Cys residues with low pKa are only one of the factors contributing to the high reactivity of Prx towards peroxides [18]. It will be important to unravel Prx catalysis at the molecular level because these enzymes are likely to participate in H2O2-mediated signaling cascades (see also Chapter 22) [16-20].
6.2. Hypochlorous acid (HOCl) and Related Species HOCl and related species (HOX, X= Cl, Br, I and SCN) are moderately strong two-
electron oxidants (E’HOCl/Cl=1.28 V, HOBr/Br=1.13 V, HOSCN/SCN=0.56V) generated during inflammatory processes (Chapter 33). They are primarily produced from the reaction of H2O2 with halide and pseudo-halide ions (Cl, Br, I and SCN ) catalyzed by MPO and eosinophil peroxidase [36, 37]. In terms of plasma concentration, the most abundant halide ion is Cl (100-140 mM) followed by Br (20-100 µM), SCN (20-120 µM) and I (0.1-0.6 µM) [38]. Thus, it is believed that HOCl is the major hypohalous acid produced during phagocytosis [37, 39]. However, the second-order rate constant value for the halogenation reaction is about 10-20 fold higher for SCN and Br compared to Cl and significant amounts of HOSCN and HOBr can also be produced.
At physiological pH, HOCl is in equilibrium with its conjugate base, hypochlorite (OCl-, pKa 7.59 at 25C) [36] (Reaction 24), and both forms appear to be responsible for oxidation and/or halogenation reactions. In acidic conditions, HOCl can be in equilibrium with molecular chlorine (Cl2, pKa 3.3) [40] (Reaction 25). In vitro studies suggest that Cl2 might be the agent that mediates formation of chlorinated products during phagocytosis [40].
HOCl is reactive towards several biomolecules. Amino (RNH2) and thiol (RSH) groups
of amino acids and peptides are among the most important targets (Table 1). Oxidation of these groups by HOCl yields unstable chloramines (RNHCl) and sulfenyl chlorides (RSCl), respectively [41-43]. Both intermediates induce further reactions that lead to an increased oxidative damage to biomolecules [36]. HOCl is also reactive towards aromatic rings in amino acids and nucleobases. Reaction of HOCl with Tyr yields 3-chlorotyrosine and 3,5-
(Reaction 22 and 23)
H2O2 + RS RSOH + H2O
RSOH + RSH RSSR + H2O
pKa = 7.4
(Reaction 24 and 25)
HOCl H+ + OCl
HOCl + Cl + H+ Cl2 + H2OpKa = 3.3

Oxygen Radicals and Related Species 17
dichlorotyrosine [44, 45]. With lipids, HOCl adds across carbon-carbon double bonds in fatty acids and cholesterol yielding chlorohydrins [46]. HOCl also reacts with H2O2/ROOH. The reaction with H2O2 produces stoichiometric amounts of 1O2 (Reaction 26) [47, 48]. Similarly, HOCl can react with lipid hydroperoxides to yield 1O2 through a mechanism involving the generation of ROO● [49].
6.3. Singlet Molecular Oxygen (1O2)
1O2 refers to the excited states of O2, the 1g and 1g+ state. They have energies of 94.3
kJ/mol and 156.9 kJ/mol above the triplet ground state, respectively. The 1g+ state has an
extremely short lifetime in H2O (~10-11 s), decaying rapidly to the 1g state, which is considered the biologically relevant form of 1O2. The lifetime of 1O2 is greatly influenced by the solvent type, being in the range of 1-5 x 10-6 s in H2O and about ten times higher in deuterium oxide (D2O). Thus, D2O is commonly employed in experiments devised to prove 1O2 formation. Photosensitization is the most conventional source of 1O2 in biological systems. Typically, this occurs by photoexcitation (type II reactions) of endogenous photosensitizers (porphyrins, flavins, quinones, etc.) exposed to UVA [50]. These processes are especially important in the skin. Excessive production of 1O2 has been associated with disease states, such as porphyrias that are characterized by porphyrin accumulation in the skin. 1O2 promotes tumor cell death and this property is exploited in photodynamic therapy. 1O2 can also be generated by non-photochemical reactions, usually involving inorganic peroxides (such as H2O2 and peroxynitrite) and organic hydroperoxides (ROOH). For instance, the generation of 1O2 has been evidenced during phagocytosis [35, 51], lipid peroxidation [52] and peroxidase turnover [53, 54]. The reaction between HOCl and H2O2 (Reaction 26) is the likely source of 1O2 during phagocytosis. The generation of 1O2 during lipid peroxidation has been attributed to the combination of ROO● radicals by the Russell mechanism [31, 55]. It is important to note that tertiary peroxyl radicals are unable to generate singlet oxygen because the hydrogen- on one of the ROO● is required for the elimination of O2 in the singlet-excited state [56]. The yield of 1O2 by this mechanism has been estimated to be approximately 3.9 to 14% [57].
1O2 is a strong two-electron oxidant that displays considerable reactivity towards electron-rich organic molecules, including nucleic acids, proteins and lipids [58]. Typically, 1O2 adds to -bonds by three common mechanisms: addition to alkenes containing allylic hydrogen by a ene-type reaction yielding hydroperoxides; 1,4-cycloaddition to 1,3-dienes (Diels-Alder reaction) forming endoperoxides; and 1,2- cycloaddition to electron-rich alkenes producing dioxetanes [59]. Chemical reactions of 1O2 occur with second-order rate constant values that are usually lower than 107 M-1 s-1. For instance, the values obtained for the chemical reaction of 1O2 with unsaturated fatty acids are in the order of 104 M-1 s-1. In contrast, physical quenching of 1O2 occurs at much faster rates and values near the diffusion-controlled limit were observed for carotenoids, among which lycopene exhibited the highest
(Reaction 26)
H2O2 + HOCl 1O2 + Cl + H2O + H+

Ohara Augusto and Sayuri Miyamoto 18
rate [60]. Reactions of 1O2 with thiols and ascorbate have been also reported. In the case of thiols, 1O2 reacts preferentially with the thiolate anion yielding several oxidation products [61]. Apparent second-order rate constant values calculated for low molecular weight thiols, such as Cys, N-acetylCys and GSH, were in the order of 106 M-1 s-1 [61]. Ascorbate reacts with 1O2 (k = 3 × 108 M-1 s-1) producing H2O2 and dehydroascorbic acid [62].
6.4. Organic Hydroperoxides from Biomolecules Several classes of organic hydroperoxides (ROOH) are produced upon oxidation of
biomolecules, including lipids, proteins and DNA [63]. Lipid hydroperoxides can be formed enzymatically during lipoxygenase, cyclooxygenase, cytochrome P450 and heme-peroxidase turnover [64, 65]. Moreover, a great number of ROOH are generated non-enzymatically by the oxidation of biomolecules mediated by radicals and 1O2 [64]. ROOH are relatively stable; however, they can participate in reactions that decrease or increase their toxicity. Normally, cells contain enzymes that reduce ROOH to their corresponding alcohols, decreasing their reactivity and toxicity. The enzymatic reduction of lipid hydroperoxides has been extensively studied. Three classes of enzymes are known to mediate their two-electron reduction: GPx, glutathione S-transferases (GST) [64, 66] and Prx [20]. Toxicity is increased when ROOH are converted to RO or ROO [3]. This can occur especially in the reaction of ROOH with transition metal ions, hemoproteins and other one-electron oxidants [3].
ROOH can react with reduced and oxidized states of free transition metal ions to form oxyl radicals. In the first case, RO● is produced whereas in the second case, ROO● is produced [3, 64]. Oxyl radicals are responsible for propagating the oxidation process as well as for the generation of other highly reactive products capable of causing modifications in proteins and nucleic acids. These include electrophilic aldehydes, epoxides, ketones, and excited species, such as 1O2 and electronically excited carbonyl species [63, 67].
6.5. Ozone (O3) O3 is present in polluted atmospheres, and inhalation of this toxic triatomic gas can
induce lung injury and inflammation. In biological systems, O3 is thought to be generated during antibody catalyzed oxidation of H2O to H2O2 [68]. In the proposed mechanism, antibodies use H2O as an electron source, facilitating its addition to 1O2 to form hydrogen trioxide (H2O3) as the first intermediate in a cascade of reactions that eventually leads to O3. The generation of O3 in human tissues has been postulated based on the detection of chemical signature products, including isatin sulfonic acid and cholesterol secoaldehyde [69]. However, these markers are not specific for O3, casting doubts on whether O3 can be generated endogenously by cells [70-72].
Reactions of O3 with fatty acids, cholesterol, amino acids and DNA have been characterized. O3 adds directly to the double bonds in fatty acids, giving rise to Criegee ozonide (10%) and a hydroxyhydroperoxide intermediate (90%) that decomposes to form aldehydes and H2O2 (Reaction 27) [70]. With saturated organic targets and inorganic compounds, O3 reacts with nucleophiles, especially those containing nitrogen or sulfur atoms

Oxygen Radicals and Related Species 19
by oxygen-transfer mechanism to generate a trioxide intermediate. Decomposition of this intermediate is reported to generate 1O2 and the corresponding oxidized product [73]. In aqueous solutions, one electron-transfer reactions can also occur, leading to the formation of ozonide (O3
), HO● and O2●- [73].
7. Conclusions In its initial four decades, the field of “free radical research in biology” has focused on
understanding the mechanisms by which radicals and oxidants are produced in vivo and how proteins, lipids and DNA are oxidized by them resulting in cell damage and, eventually, disease. The current picture is more complex, though. An increasing body of evidence suggests that apart from being potentially toxic, radicals and oxidants also exert important (patho)physiological signaling functions (Figure 3). The diverse biological activities of oxygen radicals and related species are likely rooted in the oxygen-dependent evolution of complex life forms. Although the chemical properties and reactivities of relevant biological oxidants are becoming well understood, it remains less clear how these properties translate into cellular and tissue responses. Further advances in the field will require interdisciplinary approaches combining chemical reasoning with biological insights.
Acknowledgments Our laboratories are supported by grants from Fundação de Amparo à Pesquisa do Estado
de São Paulo (FAPESP), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). The authors are members of the INCT de Processos Redox em Biomedicina-Redoxoma (CNPq/FAPESP/CAPES).
References
[1] Falkowski PG. Evolution: tracing oxygen's imprint on earth's metabolic evolution. Science 2006;311:1724-5.
[2] Lane N. The molecule that made the World. Oxford: Oxford Univ. Press.; 2004.
+ + H2O2
(Reaction 27)
O3
H2O

Ohara Augusto and Sayuri Miyamoto 20
[3] Halliwell B, Gutteridge JMC. Free radicals in biology and medicine. 4th ed. Oxford: Oxford University Press; 2007.
[4] Augusto O, Vaz SM. EPR spin-trapping of protein radicals to investigate biological oxidative mechanisms. Amino Acids 2007;32:535-42.
[5] Koppenol WH. Names for inorganic radicals (IUPAC recommendations 2000). Pure Appl Chem 2000;72:437-46.
[6] Augusto O. Free radicals: good, bad and natural (in Portuguese). São Paulo, Brazil: Oficina de Textos; 2006.
[7] Gerschman R, Gilbert DL, Nye SW, Dwyer P, Fenn WO. Oxygen poisoning and X-irradiation - a mechanism in common. Science 1954;119:623-6.
[8] McCord JM, Fridovich I. Superoxide dismutase an ezymic function for erythrocuprein (hemocuprein). J Biol Chem 1969;244:6049-55.
[9] Sies H. Biochemistry of oxidative stress. Angew Chem Int Ed 1986;25:1058-71. [10] Ignarro LJ. Biosynthesis and metabolism of endothelium-derived nitric oxide. Annu
Rev Pharmacol Toxicol 1990;30:535-60. [11] Moncada S, Palmer RMJ, Higgs EA. Nitric-oxide - physiology, pathophysiology, and
pharmacology. Pharmacol Rev 1991;43:109-42. [12] Beckman JS, Beckman TW, Chen J, Marshall PA, Freeman BA. Apparent hydroxyl
radical production by peroxynitrite - implications for endothelial injury from nitric-oxide and superoxide. Proc Natl Acad Sci U S A 1990;87:1620-4.
[13] Augusto O, Bonini MG, Amanso AM, Linares E, Santos CCX, De Menezes SL. Nitrogen dioxide and carbonate radical anion: two emerging radicals in biology. Free Radic Biol Med 2002;32:841-59.
[14] Szabo C, Ischiropoulos H, Radi R. Peroxynitrite: biochemistry, pathophysiology and development of therapeutics. Nat Rev Drug Disc 2007;6:662-80.
[15] Finkel T. Oxidant signals and oxidative stress. Curr Opin Cell Biol 2003;15:247-54. [16] Winterbourn CC, Hampton MB. Thiol chemistry and specificity in redox signaling.
Free Radic Biol Med 2008;45:549-61. [17] Winterbourn CC. Reconciling the chemistry and biology of reactive oxygen species.
Nat Chem Biol 2008;4:278. [18] Trujillo M, Ferrer-Sueta G, Radi R. Kinetic studies on peroxynitrite reduction by
peroxiredoxins. Methods Enzymol 2008;441:173-96. [19] Ogusucu R, Rettori D, Munhoz DC, Netto LES, Augusto O. Reactions of yeast
thioredoxin peroxidases I and II with hydrogen peroxide and peroxynitrite: rate constants by competitive kinetics. Free Radic Biol Med 2007;42:326-34.
[20] Rhee SG, Chae HZ, Kim K. Peroxiredoxins: A historical overview and speculative preview of novel mechanisms and emerging concepts in cell signaling. Free Radic Biol Med 2005;38:1543-52.
[21] Lancaster JR. Diffusion of free nitric oxide. Methods Enzymol 1996;268:31-50. [22] NDRL/NIST Solution Kinetics Database on the Web. NIST Standard Reference
Database 40. A compilation of kinetics data on solution-phase reactions. In: http://kineticsnistgov/solution/; 2002.
[23] Bielski BHJ, Richter HW. Study of superoxide radical chemistry by stopped-flow radiolysis and radiation-induced oxygen-consumption. J Am Chem Soc 1977;99:3019-23.

Oxygen Radicals and Related Species 21
[24] D'Autreaux B, Toledano MB. ROS as signalling molecules: mechanisms that generate specificity in ROS homeostasis. Nat Rev Mol Cell Biol 2007;8:813.
[25] Winterbourn CC, Kettle AJ. Radical-radical reactions of superoxide: a potential route to toxicity. Biochem Biophys Res Commun 2003;305:729-36.
[26] Winterbourn CC, Metodiewa D. Reactivity of biologically important thiol compounds with superoxide and hydrogen peroxide. Free Radic Biol Med 1999;27:322-8.
[27] Winterbourn CC, Metodiewa D. The reaction of superoxide with reduced glutathione. Arch Biochem Biophys 1994;314:284.
[28] Medinas DB, Cerchiaro G, Trindade DF, Augusto O. The carbonate radical and related oxidants derived from bicarbonate buffer. IUBMB Life 2007;59:255-62.
[29] Antunes F, Salvador A, Marinho HS, Alves R, Pinto RE. Lipid peroxidation in mitochondrial inner membranes.1. An integrative kinetic model. Free Radic Biol Med 1996;21:917-43.
[30] Xu L, Davis TA, Porter NA. Rate constants for peroxidation of polyunsaturated fatty acids and sterols in solution and in liposomes. J Am Chem Soc 2009;131:13037-44.
[31] Russell GA. Deuterium-isotope effects in the autoxidation of aralkyl hydrocarbons - mechanism of the interaction of peroxy radicals. J Am Chem Soc 1957;79:3871-7.
[32] van der Vliet A, Eiserich JP, Halliwell B, Cross CE. Formation of reactive nitrogen species during peroxidase-catalyzed oxidation of nitrite. J Biol Chem 1997;272:7617-25.
[33] Heinecke JW, Li W, Francis GA, Goldstein JA. Tyrosyl radical generated by myeloperoxidase catalyzes the oxidative cross-linking of proteins. J Clin Invest 1993;91:2866-72.
[34] Vaz SM, Prado FM, Di Mascio P, Augusto O. Oxidation and nitration of ribonuclease and lysozyme by peroxynitrite and myeloperoxidase. Arch Biochem Biophys 2009;484:127-33.
[35] Klebanoff SJ. Oxygen metabolites from phagocytes. In: Gallin JI, Snyderman R, eds. Inflammation: basic principles and clinical correlates. Philadelphia: Lippincott Williams & Wilkins; 1999:721-68.
[36] Davies MJ, Hawkins CL, Pattison DI, Rees MD. Mammalian heme peroxidases: from molecular mechanisms to health implications. Antioxid Redox Signal 2008;10:1199-234.
[37] Klebanoff SJ. Myeloperoxidase: friend and foe. J Leukoc Biol 2005;77:598-625. [38] Furtmuller PG, Burner U, Obinger C. Reaction of myeloperoxidase compound I with
chloride, bromide, iodide, and thiocyanate. Biochemistry 1998;37:17923-30. [39] Hampton M, Kettle A, Winterbourn C. Inside the neutrophil phagosome: oxidants,
myeloperoxidase, and bacterial killing. Blood 1998;92:3007-17. [40] Hazen SL, Hsu FF, Mueller DM, Crowley JR, Heinecke JW. Human neutrophils
employ chlorine gas as an oxidant during phagocytosis. J Clin Invest 1996;98:1283-9. [41] Prutz WA. Hypochlorous acid interactions with thiols, nucleotides, DNA, and other
biological substrates. Arch Biochem Biophys 1996;332:110-20. [42] Drozdz R, Naskalski JW, Sznajd J. Oxidation of amino acids and peptides in reaction
with myeloperoxidase, chloride and hydrogen peroxide. Biochim Biophys Acta 1988;957:47-52.
[43] Hawkins CL, Pattison DI, Davies MJ. Hypochlorite-induced oxidation of amino acids, peptides and proteins. Amino Acids 2003;25:259-74.

Ohara Augusto and Sayuri Miyamoto 22
[44] Winterbourn CC, Kettle AJ. Biomarkers of myeloperoxidase-derived hypochlorous acid. Free Radic Biol Med 2000;29:403-9.
[45] Hazen SL, Heinecke JW. 3-Chlorotyrosine, a specific marker of myeloperoxidase-catalyzed oxidation is markedly elevated in low density lipoprotein isolated from human atherosclerotic intima. J Clin Invest 1997;99:2075-81.
[46] Winterbourn CC, Kettle AJ. Biomarkers of myeloperoxidase-derived hypochlorous acid. Free Radic Biol Med 2000;29:403-9.
[47] Khan AU, Kasha M. Chemiluminescence arising from simultaneous transitions in pairs of singlet oxygen molecules. J Am Chem Soc 1970;92:3293-300.
[48] Held AM, Halko DJ, Hurst JK. Mechanism of chlorine oxidation of hydrogen peroxide. J Am Chem Soc 1978;100:5732-40.
[49] Miyamoto S, Martinez GR, Rettori D, Augusto O, Medeiros MHG, Di Mascio P. Linoleic acid hydroperoxide reacts with hypochlorous acid, generating peroxyl radical intermediates and singlet molecular oxygen. Proc Natl Acad Sci U S A 2006;103:293-8.
[50] Baier J, Maisch T, Maier M, Engel E, Landthaler M, Baumler W. Singlet oxygen generation by UVA light exposure of endogenous photosensitizers. Biophys J 2006;91:1452-9.
[51] Hampton MB, Kettle AJ, Winterbourn CC. Inside the neutrophil phagosome: oxidants, myeloperoxidase, and bacterial killing. Blood 1998;92:3007-17.
[52] Cadenas E, Sies H. Low-level chemi-luminescence as an indicator of singlet molecular-oxygen in biological-systems. Methods Enzymol 1984;105:221-31.
[53] Kiryu C, Makiuchi M, Miyazaki J, Fujinaga T, Kakinuma K. Physiological production of singlet molecular oxygen in the myeloperoxidase-H2O2-chloride system. FEBS Lett 1999;443:154-8.
[54] Kanofsky JR. Singlet oxygen production by lactoperoxidase. J Biol Chem 1983;258:5991-3.
[55] Miyamoto S, Martinez GR, Medeiros MHG, Di Mascio P. Singlet molecular oxygen generated from lipid hydroperoxides by the Russell mechanism: studies using 18O-labeled linoleic acid hydroperoxide and monomol light emission measurements. J Am Chem Soc 2003;125:6172-9.
[56] Howard JA, Ingold KU. Self-reaction of sec-butylperoxy radicals. Confirmation of Russell mechanism. J Am Chem Soc 1968;90:1056-8.
[57] Mendenhall GD, Sheng XC, Wilson T. Yields of excited carbonyl species from alkoxyl and from alkylperoxyl radical dismutations. J Am Chem Soc 1991;113:8976-7.
[58] Cadet J, Di Mascio P. Peroxides in biological system. In: Rappoport Z, ed. The chemistry of peroxides. West Sussex: John Wiley & Sons Ltd; 2006:915-99.
[59] Frimer A. Singlet O2. Boca Raton, FL: CRC; 1985. [60] Di Mascio P, Kaiser S, Sies H. Lycopene as the most efficient biological carotenoid
singlet oxygen quencher. Arch Biochem Biophys 1989;274:532-8. [61] Devasagayam TPA, Sundquist AR, Di Mascio P, Kaiser S, Sies H. Activity of thiols as
singlet molecular oxygen quenchers. J Photochem PhotobiolB 1991;9:105-16. [62] Kramarenko GG, Hummel SG, Martin SM, Buettner GR. Ascorbate reacts with singlet
oxygen to produce hydrogen peroxide. Photochem Photobiol 2006;82:1634-7. [63] Miyamoto S, Ronsein GE, Prado FM, et al. Biological hydroperoxides and singlet
molecular oxygen generation. IUBMB Life 2007;59:322.

Oxygen Radicals and Related Species 23
[64] Girotti AW. Lipid hydroperoxide generation, turnover, and effector action in biological systems. J Lipid Res 1998;39:1529-42.
[65] Caro AA, Cederbaum AI. Role of cytochrome P450 in phospholipase A2- and arachidonic acid-mediated cytotoxicity. Free Radic Biol Med 2006;40:364-75.
[66] Brigelius-Flohé R. Tissue-specific functions of individual glutathione peroxidases. Free Radic Biol Med 1999;27:951-65.
[67] Esterbauer H, Schaur RJ, Zollner H. Chemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic Biol Med 1991;11:81-128.
[68] Wentworth P, Jr., McDunn JE, Wentworth AD, et al. Evidence for antibody-catalyzed ozone formation in bacterial killing and inflammation. Science 2002;298:2195-9.
[69] Wentworth P, Jr., Nieva J, Takeuchi C, et al. Evidence for ozone formation in human atherosclerotic arteries. Science 2003;302:1053-6.
[70] Pryor WA, Houk KN, Foote CS, et al. Free radical biology and medicine: it's a gas, man! Am J Physiol Regul Integr Comp Physiol 2006;291:R491-511.
[71] Brinkhorst J, Nara SJ, Pratt DA. Hock ceavage of cholesterol 5α-hydroperoxide: an ozone-free pathway to the cholesterol ozonolysis products identified in arterial plaque and brain tissue. J Am Chem Soc 2008;130:12224-5.
[72] Uemi M, Ronsein GE, Miyamoto S, Medeiros MHG, Di Mascio P. Generation of cholesterol carboxyaldehyde by the raction of singlet molecular oxygen [O2 (
1Δg)] as well as ozone with cholesterol. Chem Res Toxicol 2009;22:875-84.
[73] Munoz F, Mvula E, Braslavsky SE, von Sonntag C. Singlet dioxygen formation in ozone reactions in aqueous solution. J Chem Soc-Perkin Trans 2 2001:1109-16.
[74] Buettner GR. The pecking order of free radicals and antioxidants: lipid peroxidation, -tocopherol and ascorbate. Arch Biochem Biophys 1993;300:535-43.



![[Doi 10.1016_j.cbi.2014.10.016] v. I. Lushchak -- Free Radicals, Reactive Oxygen Species, Oxidative Stress and Its Classification](https://img.dokumen.tips/doc/110x75/577cc15e1a28aba71192c95f/doi-101016jcbi201410016-v-i-lushchak-free-radicals-reactive-oxygen.jpg)

























