Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, June 1968, p. 2171-2176Copyright @ 1968 American Society for Microbiology
Vol. 95, No. 6Printed in U.S.A.
Inhibition of Histoplasma capsulatum by Candidaalbicans and Other Yeasts on Sabouraud's
Agar MediaL. KAPICA, C. E. SHAW, AND G. W. BARTLETT
Department ofMicrobiology and Immunology, McGill University, Montreal, P.Q., Canada
Received for publication 8 April 1968
The inhibition of growth of Histoplasma capsulatum by Candida albicans andother yeasts on Sabouraud's agar was investigated. Histoplasma (yeast-phaseinoculum) was grown alone and in mixtures with yeasts at 25 C for 4-week periods.As few as 10 colonies of C. albicans completely inhibited the growth of approxi-mately 50,000 potential colonies of Histoplasma. The pH was determined in cul-tures of 36 colonies of Candida on media containing 1, 2, and 4% glucose by spottingthe agar with pH indicators. A drop in the pH became noticeable in all three mediaabout the 3rd day of incubation, and a pH of 3.5 was reached in about 7 days.Subsequently, the pH remained almost stationary in the 4% glucose-agar, roseslowly in the 2% glucose-agar, and rose sharply in the 1 % glucose-agar. The growthof Histoplasma was inhibited completely at pH 4 and below. When the pH wascontrolled in mixed cultures, some growth of Histoplasma was obtained. Substitu-tion of maltose for glucose delayed the development of acidity and allowed theappearance of numerous mycelial colonies in the presence of Candida. This growthwas arrested as soon as the medium became acid. Four other species which alsoacidified the Sabouraud's medium effected similar inhibition. It was thus shownthat severe and prolonged acidity produced by some yeasts in the sugar-rich Sabou-raud's media is alone sufficient to completely inhibit Histoplasma during the stand-ard 4-week incubation of specimens such as sputum.
In diagnostic mycology, Candida albicans isfrequently encountered in cultures of sputum(1, 2, 6, 13) and it has been found that specimenswhich are only barely positive on direct examina-tion may contain as many as 103 yeast cells perml (8). The failure to isolate Histoplasma capsu-latum from sputum of patients has been com-mented upon in several publications (3, 5, 12).In a survey of 130 publications dealing with theprimary isolation of H. capsulatum between 1934and 1966, it was found that in 57% of theseSabouraud's agar was used as a medium ofprimary isolation. The inhibition of H. capsu-latum by C. albicans on Sabouraud's agar wasreported previously (J. Burns and H. W. Larsh,Bacteriol. Proc., p. 66, 1962) and has also beenobserved repeatedly in our laboratory.
Since marked acidity is produced by yeasts inglucose-rich media (7, 17, 18), the developmentof acidity by yeasts in Sabouraud Dextrose Agarwas investigated in the present study as a factor
1 Present address: American Hospital SupplyCorp., Evanston, Ill.
in the inhibition of Histoplasma, a fungus whichrequires an initial pH of 6.3 to 8.1 for optimalgrowth from yeast-phase inoculum (16). Pre-liminary results of this investigation were re-ported previously (L. Kapica, C. E. Shaw, andG. W. Bartlett, Bacteriol. Proc., p. 68, 1967).
MATERIALS AND METHODS
Media. Commercial media used includedSabouraud Dextrose Agar (Difco), Sabouraud'sagar modified (Difco), Mycosel Agar (BBL), BrainHeart Infusion Agar (Difco) to which 5% (v:v)sheep blood was added, and Brain Heart Infusion-blood-agar, as above, to which 1% (w/v) glucose wasadded. Ingredients per liter of other media preparedwere as follows: beef extract-agar (beef extract, 3 g;Neopeptone, 10 g; cerelose, 10 g; sodium chloride, 5 g;agar, 20 g); McGill nutrient blood-agar (beef heart,defatted, infusion from 454 g; proteose peptone, 10 g;sodium chloride, 2.5 g; potassium chloride, 0.2 g;calcium chloride, 0.1 g; agar, 15 g; human blood, 50ml); Neopeptone (Difco)-maltose- or -glucose-agar(Neopeptone, 10 g; maltose or glucose, 40 g; agar,20 g; autoclaved 50%o maltose solution or filter-
2171
on April 8, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

KAPICA, SHAW, AND BARTLETT
sterilized 50% glucose solution added to a separatelyautoclaved Neopeptone-agar solution).The final pH of all media except Sabouraud
Dextrose Agar (pH 5.6) was pH 7.0 i 0.5. Sterilemedia were cooled to 50 C and dispensed in 20-mivolumes into 250-ml Erlenmeyer flasks; they wereused within 24 to 48 hr. All flasks were plugged withcotton wool.
Buffered Sabouraud Dextrose Agar and Sabou-raud's agar modified were prepared by using 0.1 Mphosphate buffer as a solvent or by adding a separatelyautoclaved suspension of CaCO3 to the media justbefore distribution. Final concentration of the CaCO3was 0.5% and thepH was 7.3. The pH of the Neopep-tone-glucose-agar was adjusted to several acid pHlevels by adding precalculated volumes of 3.5 N HClto sterile samples of the autoclaved medium cooled to50 C. The medium was then stirred thoroughly anddispensed at once.
Strains tested. A strain of H. capsulatum, no. 649,isolated from a case of pulmonary histoplasmosis atthe Royal Victoria Hospital in Montreal in 1965, wasused throughout the experiments. Four strains isolatedfrom recent cases of histoplasmosis at different hospi-tals in Montreal, one strain obtained from the Ameri-can Type Culture Collection, and a mixture of sixstrains were used for comparative studies. Stockcultures were maintained in yeast phase at 37 C onsealed slopes of McGill nutrient blood-agar. Thestrain of C. albicans used throughout the study wasisolated from a culture of a specimen of bronchialaspirations in which it was observed to be inhibitoryto H. capsulatum. For comparative studies, six otherstrains of C. albicans isolated from clinical specimensand seven other species of yeast including C. parapsilo-sis, C. tropicalis, C. curvata, C. brwnptii, C. catenulata,Cryptococcus neoformans, and Saccharomyces cere-visiae were used. Stock cultures were maintained onSabouraud's agar modified at 4 C.
Inoculum and inoculating procedure. Yeast-phaseH. capsulatum was grown on sealed slopes of McGillnutrient blood-agar at 37 C for 2 days, harvested witha loop into 10 ml of saline, and filtered through glasswool to remove large aggregates of cells. The densityof this stock suspension was read on a model 7 Cole-man photo-nephelometer (Standard 21) and wasadjusted to give a reading between 25 and 50. One mlof a 10-3 dilution of stock suspension was mixed with1 ml of 0.7% agar at 45 to 50 C; 1 ml of this agarsuspension was used to inoculate one flask. A thinlyconfluent growth was obtained in which individualcolonies could no longer be discerned. These dilutionswere estimated by serial platings on Sabouraud's agarmodified to produce 5,000 to 7,000 colonies per flask.Microscopic examinations of the stock suspensionsshowed that the majority of the infective units were inthe form of small clumps of two to three cells with agood proportion of single cells; larger clumps wereoccasionally encountered. C. albicans and other yeastswere grown on Sabouraud's agar modified for 2 days;inoculum was prepared in the same manner as that ofH. capsulatum. A 0.5-ml amount of a 10-5 dilutionproduced the desired number of colonies, about 35per flask. Careful readjustment in the preparation of
the dilutions resulted on most oxcasions in closelyreproducible numbers; however, extra units wereusually prepared to allow for elimination of thosecarrying unsuitable numbers of colonies. Microscopicexaminations of the stock suspensions showed thatsingle and budding cells only were present.
Mixed populations were prepared by combiningequal volumes of double-strength saline suspensionsof the two organisms just prior to mixing with theagar. All tests were set up in triplicate. After 1 to 2hr on the bench, the flasks were placed in a 25-Cincubator for a standard 4-week incubation. Thecultures were examined once a week.
In some experiments, mycelial-phase inoculum ofH. capsulatum was required. This was prepared byplacing sterile analytical filter paper rings (outsidediameter, 2 cm; inside diameter, 1 cm), manufacturedby Carl Schleicher & Schuell Co., Keene, N.H., onplates of Neopeptone-glucose-agar. The rings adheredwell to the surface moisture of the agar. The plateswere then flooded with a double volume of standardyeast-phase inoculum and were incubated for 3 daysat 25 C. The rings were then transferred with sterileforceps, without damage, to the test media and placedwith the surface bearing inoculum in an upright posi-tion.
Determination ofpH in agar media. Since agar pHcannot be measured with electrodes, bromocresolpurple (pH 6.8 to 5.2), bromocresol green (pH 5.4 to3.8), and bromophenol blue (pH 4.5 to 3.0) were used.All three were nontoxic for Candida, but only bromo-cresol purple was nontoxic for Histoplasma. pH deter-minations were made by incorporating indicator inthe medium at a final concentration of 0.002% or byspotting the agar with drops of 0.04% solutions ofindicator. The color of the spots was read againstsimilarly made spots on standards. (Standards wereprepared by acidifying lots of Neopeptone-glucose-agar to 12 pH levels between pH 3.0 and 6.5 anddistributing the agar in 20-ml portions to flasks in theroutine manner.) Standards were used because colorsobtained at a given pH did not correspond preciselywith those quoted by the Merck Index, probablybecause of the organic nature of the medium. Shadesof colors read consistently by the same person differedsufficiently to allow, in most cases, the determinationofpH differences of as little as 0.25 of a unit.
RESULTSLower limits of inhibition. Sabouraud Dex-
trose Agar was inoculated with mixed inoculacontaining the standard density of Histoplasmaand decreasing densities of Candida (Table 1).As few as 10 colonies of Candida scattered overthe agar surface completely inhibited the growthof Histoplasma. Fewer colonies allowed irregularpockets of atypical colonies to appear where thespace between the yeast colonies was somewhatlarger than that on the rest of the agar. It wasalso observed that the fewer the yeast coloniesper flask the larger they grew, and it was calcu-lated that scattered colonies of C. albicans were
2172 J. BACTERIOL.
on April 8, 2019 by guest
http://jb.asm.org/
Dow
nloaded from
INHIBITION OF H. CAPSULATUM BY C. ALBICANS
TABLE 1. Lower limits of inhibition ofHistoplasmacapsulatum by Candida albicansa
C. albicans
No. of Growth of H. capsulatumcolonies/ Colony Area
flask diameter occupied
mm %45 5.5 24 No growth26 No growth10 8.5 16 No growth6 Poor, nonaerial3 16 14 Poor, nonaerial0 Abundant, typical
a Sabouraud Dextrose28 days, 25 C.
in the pH was observed in the 4% glucose me-dium (Sabouraud Dextrose Agar). By the 5thday of incubation, the pH had dropped to 3.75to 4.0 and, by the 14th day, to 3.0 to 3.25. Afterthat, a very slow rise occurred to pH 3.5. Thismedium thus remained at a pH below 4 for about24 days of the 4-week period. In the 2% glucosemedium (Sabouraud's agar modified), the pHcurve dropped to its lowest point (pH 3.5 to3.75) about the 12th day. It then began to riseand reached pH 4.75 at the end of the 4th week.In the 1% glucose nmedium, the first pH readingvaried considerably according to the distance
Agar, initial pH, 5.6;
completely inhibitory to Histoplasma when theyoccupied as little as 15 to 20% of the agar surface.When increasingly heavy inocula of Histo-
plasma were tested against the lowest inhibitorynumbers of C. albicans on Sabouraud DextroseAgar, it was found that 10 scattered colonies ofthe yeast were completely inhibitory to about50,000 (heaviest inoculum tested) potentialcolonies of Histoplasma.
It was observed that the nature of the inhibi-tion was fungistatic rather than fungicidal. Whenagar blocks, apparently free of growth, werelifted after 2 weeks of incubation from among theCandida colonies and squashed on fresh slopesof Sabouraud Dextrose Agar, growth of Histo-plasma appeared within a few days.pH changes in cultures of C. albicans on Neo-
peptone-glucose-agar. In preliminary tests, Sabou-raud Dextrose Agar, in which bromocresol purpleindicator had been incorporated, was inoculatedwith pure and mixed populations of the twoorganisms. Since the pH of this medium was 5.6,the initial color of the medium was green. In allCandida-carrying units, yellow zones appearedaround each colony of yeast 2 to 3 days afterinoculation. These zones enlarged visibly at 5days, and at 7 days the whole medium was yellow.In pure cultures of Histoplasma, the mediumturned purple when growth appeared in about10 to 14 days, indicating a rise in the pH.Acid production by the yeasts was demon-
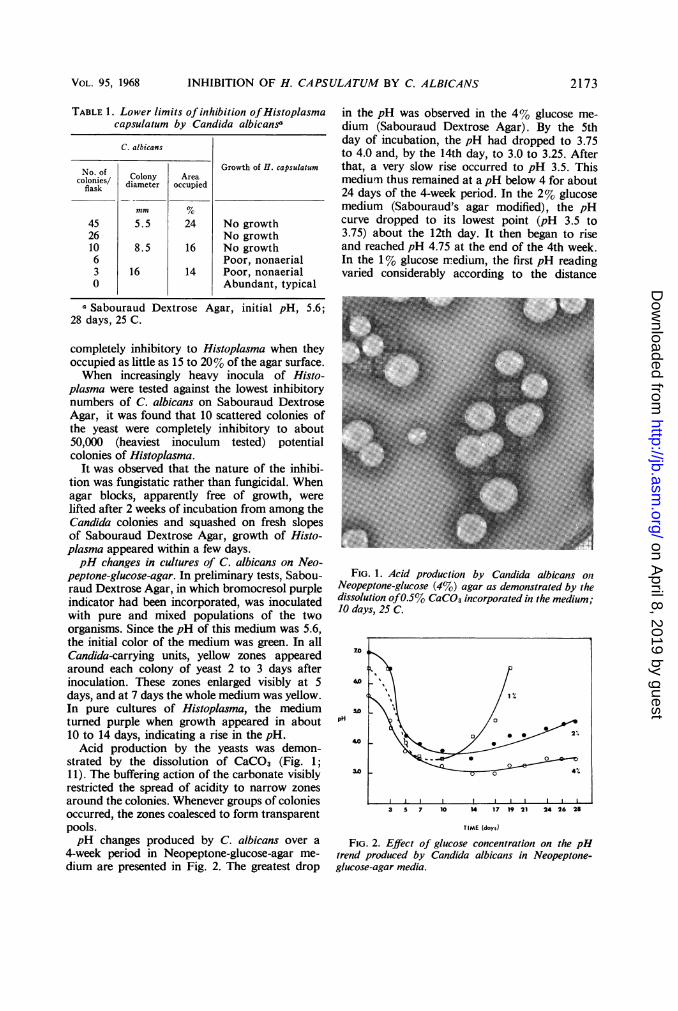
strated by the dissolution of CaCO3 (Fig. 1;11). The buffering action of the carbonate visiblyrestricted the spread of acidity to narrow zonesaround the colonies. Whenever groups of coloniesoccurred, the zones coalesced to form transparentpools.pH changes produced by C. albicans over a
4-week period in Neopeptone-glucose-agar me-dium are presented in Fig. 2. The greatest drop
FIG. 1. Acid production by Candida albicans on?Neopeptone-glucose (4%) agar as demonstrated by thedissolution of0.5% CaCO3 incorporated in the medium;10 days, 25 C.
7.0
pH
40
3.0
TIME (doys)
FIG. 2. Effect of glucose concentration on the pHtrend produced by Candida albicans in Neopeptone-glucose-agar media.
VOL. 95, 1968 2173
on April 8, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

KAPICA, SHAW, ANDBARTLETJ
from the nearest colony (Fig. 2, broken line).The lowest point of this curve (pH 3.5) occurredat about the 10th day of incubation. After that,the curve rose sharply to the original level ofpH 6.5.
Growth of Histoplasma on Neopeptone-glucose-agar at acid pH. Since the drop of pH in Neo-peptone-glucose-agar had a lag phase of about3 days or a period sufficiently long to allow areversion of the yeast phase to mycelial phase,the effect of pH on a 3-day-old mycelial phasewas determined. Flasks of Neopeptone-glucose-agar, adjusted to seven pH levels from 3.0 to6.5, were inoculated with rings of 3-day-oldmycelial-phase inoculum in one series and withthe standard yeast-phase inoculum in anotherseries. Table 2 shows the detrimental effect ofpH 4.5 on growth from yeast-phase inoculum;at pH 4 and below, no growth occurred. The3-day-old mycelial-phase inoculum exhibitedsome tolerance to acid pH. The growth in thisseries appeared first on the rings in the form ofaerial tufts of mycelium (about 50 to 60 coloniesper ring were counted under best conditions)which, when the pH was suitable, spread rapidlyto the agar medium. AtpH 4.5, growth, althoughstill fiirly vigorous on the rings, spread slowlyto the agar. AtpH 4, only a few tufts of myceliumappeared on the rings late during the incubation,and the hyphae did not grow onto the acid agar.
Effect of glucose concentration on inhibition.Flasks of Neopeptone-glucose-agar were pre-pared with five glucose concentrations between0.1 and 4% and were inoculated with pure andmixed populations (Table 3). Whereas bothorganisms in pure population grew better athigher glucose concentrations, the degree ofinhibition of Histoplasma in mixed populationswas directly related to the glucose concentration.As the glucose concentration went down to 1%and less, colonies of Histoplasma began to appeardepending on the distance from the nearestCandida colonies. In the last two series, growthof Histoplasma, although meager, appeared allover, in spite of the presence of Candida; tuftsof aerial mycelium were actually observed on topof Candida colonies. Comparison of growth ofH. capsulatum in pure and mixed cultures in theabsence of glucose showed that the quantity ofgrowth did not appear to differ in the two situa-tions, but the growth in mixed populations wassomewhat stunted as compared to that in purepopulations.
Effect of buffering on inhibition. SabouraudDextrose Agar and Sabouraud's agar modifiedwere buffered with 0.1 M phosphate buffer atpH 7 and calcium carbonate at pH 7.3. Thecultures were inoculated with the standard mixed
TABLE 2. Growth ofHistoplasma capsulatum onNeopeptone-glucose (4%)-agarG
Initial pH of mediumInoculum
6.5 6.0 5.0 4.5 4.0 3.5 3.0
Yeastphase... XXXXb XXXX XXX X 0 0 0
3-day-oldmycelialphase... XXXX XXXXXXxx xxX (X) 0 0
a Adjusted to seven pH levels; 28 days, 25 C.b Symbols: X, growth aerial and typical; the greatest number
of symbols denotes best growth; 0, no growth.
TABLE 3. Effect of glucose concentration oninhibition of Histoplasma capsulatumby Candida albicans on Neopep-
tone-glucose-agara
Glucose Growth of H. capsulatumin mixed populationsb
4.0 No growth2.0 No growth1.0 Pockets of poor, nonaerial colonies0.5 Wide areas of small, tufted colonies0.1 Confluent, thin, stunted aerial growth0.0 Confluent, thin, stunted aerial growth
a Initial pH, 7.0; 28 days, 25 C.b In pure cultures of H. capsulatum, typical
aerial confluent growth occurred at all glucoseconcentrations. Heaviest growth occurred with4% glucose and thinnest growth with 0.1% glucoseor with no glucose.
and pure populations of the two species. (Arather high population of 77 colonies per flaskof Candida was obtained in this experiment.)Bromocresol purple was incorporated in all units.The buffers reduced the duration of the acid
phase in all cultures carrying the yeast, i.e., themedia turned yellow later than under unbufferedconditions and became alkaline before the endof the experimental 4-week incubation, soonerin Sabouraud's agar modified than in SabouraudDextrose Agar. The carbonate buffer controlledthe acidity more effectively than the phosphates;the opacity in these media disappeared almostcompletely in Sabouraud Dextrose Agar andin about 25% in Sabouraud's agar modified.
In mixed cultures, the buffers were only par-tially successful in counteracting the inhibition,the effect varying with the buffers and the mediaused. The phosphates failed completely in Sabou-raud Dextrose Agar, i.e., no growth of Histo-plasma appeared; in Sabouraud's agar modified,some growth which was atypical and nonaerial
2174 J. BACTERIOL.
on April 8, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

INHIBITION OF H. CAPSULATUM BY C. ALBICANS
appeared at the end of the 3rd week of incubation.Both carbonate-buffered media produced growthof Histoplasma, a little more abundantly inSabouraud's agar modified than in SabouraudDextrose Agar; this growth was also atypical. Inpure cultures, both buffers affected adversely thegrowth of Histoplasma in quantity, quality, andtime of appearance. The carbonate buffer did notaffect Candida, but the phosphates did; thecolonies appeared late and remained very smallfor about one week. This growth improved duringthe 3rd week and was normal at the end of the4th week.
Inhibition on Neopeptone-maltose-agar. In purepopulations, typical growth of Histoplasmaappeared 3 days earlier and was more vigorousthan on Sabouraud Dextrose Agar. Colonies ofCandida were slow to appear both in pure andmixed populations. In mixed populations, pin-point filamentous colonies of Histoplasma com-parable to those in pure populations appeared inas little as 6 to 7 days over the entire agar, re-gardless of the presence of Candida. However,this growth failed to develop further and, at 28days, close examination was still required todetect its presence on the medium. A furtherincubation period of 3 weeks showed no change.When a pH curve analogous to those in Fig. 2
was constructed, it was found that, in this me-dium, C. albicans produced a drop in pH whichwas delayed by 2 to 3 days, as compared to thatproduced in Sabouraud Dextrose Agar. After7 days of incubation, however, the maltosemedium became and remained as acid as the 4%glucose medium.
Inhibition by other strains of C. albicans andother species of yeasts. Six other strains of C.albicans were paired against six other strains ofH. capsulatum on Sabouraud Dextrose Agar, andtotal inhibition was observed in every case.Bromocresol purple included in the medium indi-cated a pH below 5.2 over the 4-week' incubationperiod. Four other species of yeasts, C. tropicalis,C. parapsilosis, S. cerevisiae, and Cryptococcusneoformans, tested against H. capsulatum 649 onSabouraud Dextrose Agar, also produced totalinhibition. The yeast population varied from 21to 62 colonies per flask. Bromocresol purple,included in the medium, indicated a pH below5.2 in all cases except in cultures of C. parapsilosis,which became alkaline in the last days of incuba-tion. Some yeasts were found (C. curvata, C.brumptli, and C. catenulata) which allowedstunted and atypical growth of Histoplasma toappear on Neopeptone-glucose (2%)-agar. Bro-mocresol purple indicator, incorporated in themedium, showed that the pH drop produced bythese species was less severe than that produced
by species which caused complete inhibition.Although the agar turned yellow in cultures ofC. brumptii and C. catenulata, purple patcheswere left at the end of the 4th week; in the caseof C. curvata, the agar did not turn yellow butremained green to yellow-green.
Growth of Histoplasma in the presence of C.albicans on other media. Sabouraud DextroseAgar and Sabouraud's agar modified served ascontrol media and exhibited the usual inhibition.(These cultures were kept for another 4 weeksat 25 C. Toward the end of that period, i.e., after8 weeks of incubation, a small amount of growthof Histoplasma appeared among the Candidacolonies on Sabouraud's agar modified but noton Sabouraud Dextrose Agar.) Mycosel Agarand beef extract-agar produced partially inhibitedgrowth of Histoplasma which appeared late andwas obviously reduced in quantity. On about the10th day, the three blood-containing mediaproduced a heavy but atypical growth of Histo-plasma with very poorly developed aerial my-celium. Characteristically, the colonies appearedover the entire agar surface without any evidenceof inhibition, and even grew underneath Candidacolonies.
DIscussioNThe difficulty in obtaining growth of H. capsu-
latum in the presence of C. albicans and otheryeasts on the two commercial brands of Sabou-raud's agar appears to be centered around theproduction by the yeast of an acid pH in themedium. The acid pH alone is strongly inhibitoryto Histoplasma. This would agree with results byGarrison (4), who found that the oxidation ofglucose by the yeast phase of H. capsulatum had adefinite pH optimum of 7.0, whereas hydrogenion concentrations below pH 4 were inhibitoryfor both endogenous respiration and glucoseoxidation. Since the severity of the inhibition isdirectly related to the glucose concentration ofthe medium, detoxification of the medium pre-sumably takes place once the sugar supply of themedium becomes exhausted, and subsequentutilization of organic acids begins. Thus, mediacontaining low concentrations of glucose (0.1 to1.0%) allowed some growth of Histoplasmawithin the 4-week incubation period, whereasmedia containing higher concentrations of glucoseallowed growth only after 8 weeks of incubation(Sabouraud's agar modified) or allowed nogrowth at all (Sabouraud Dextrose Agar).Although undoubtedly there are other factorswhich play a role in inhibition, it is conceivablethat the prevailing low pH inactivates proteolyticenzymes of Histoplasma and, consequently, novisible growth occurs under conditions of nitrogen
2175VOL. 95, 1968
on April 8, 2019 by guest
http://jb.asm.org/
Dow
nloaded from

KAPICA, SHAW, AND BARTLETT
starvation. That this supposition is probablycorrect was shown by the fact that the inhibitorystate of the medium is fungistatic rather thanfungicidal.
Since control of pH in Sabouraud's mediawas only partially successful in reducing theinhibition, and since clinical specimens wouldprobably contain fewer infective units of Histo-plasma than the inoculum used in this study, itcould be expected that chances of isolation ofHistoplasma from yeast-carrying specimens wouldbe small even if alterations were made in thesemedia. Although prolonged incubation of cul-tures for 2 to 3 months may result in positivecultures, this technique is both unreliable andimpractical. Existing naturally buffered media,such as brain heart infusion-glucose-blood-agar(9, 10, 14, 15), grow Histoplasma in the presenceof yeasts without apparent reduction in thenumber of colonies; therefore, it is stronglyrecommended that these media be used, ratherthan Sabouraud's, for the primary isolation ofHistoplasma from specimens from the upperrespiratory tract. The growth of Histoplasmaon such media, although atypical, will becomeeasily recognizable with experience. However,since other fungi may assume similar atypicalmorphology on these media, the suspected fungusmust be transferred for positive identification toa medium on which Histoplasma, in pure culture,will exhibit typical growth and sporulation; forthis purpose, the Sabouraud's media are mostappropriate. In cases where colonies of Histo-plasma are growing in close proximity to yeasts,purification of the fungus of yeasts will be neces-
sary before transferring to Sabouraud's agar.This may be accomplished by intraperitonealpassage through mice or, in case of C. albicans,by the use of the biotin-free medium devised byBurns and Larsh (Bacteriol. Proc., p. 66, 1962).
ACKNOWLEDGMENTSAppreciation is hereby expressed to J. W. Stevenson
for his important hint which helped to set this work onthe right course, to H. Coletta for the photographs,and to the following for furnishing strains of H.capsulatum used in comparative studies: J. DeVries(The Montreal General Hospital), A. Taussig (TheJewish General Hospital, Montreal), M. Domaradzka(St. Joseph Hospital of Rosemount), and E. Mankie-wicz (The Royal Edward Chest Hospital, Montreal).
This work was supported by a grant from theMedical Research Council, Ottawa, Canada.
LrERATupx CIrED1. Armstrong, E. C., and J. A. Hall. 1956. The
incidence of Candida species in routine speci-
mens of sputum. Monthly Bull. Min. HealthLab. Serv. 15:220-224.
2. Blank, H. 1957. Candida albicans, frequent com-panion, occasional foe. Monographs Therapy2:1-6.
3. Carski, T. R., G. C. Cozad, and H. W. Larsh.1962. Detection of Histoplasma capsulatum insputum by means of fluorescent antibody stain-ing. Am. J. Clin. Pathol. 37:465-469.
4. Garrison, R. G. 1961. Studies of the respiratoryactivity of Histoplasma capsulatwn. I. Aspectsof the aerobic metabolism of the yeast phase.J. Infect. Diseases 108:120-124.
5. Hamil, B. M., W. R. Eyker, J. W. Rebuck, andG. A. Logrippo. 1959. Bronchopulmonarymycosis: a 20 year study of siblings. HenryFord Hosp. Med. Bull. 7:295-308.
6. Helms, P. 1956. The occurrence of Candidaalbicans in sputum in Denmark. J. Clin. Pathol.9:372-373.
7. Johnson, S. A. M., M. G. Guzman, and C. T.Aguilera. 1954. Candida (Monilia) albicans.Effect of amino acids, glucose, pH, chlortetra-cycine, dibasic sodium and calcium phosphates,and anaerobic and aerobic conditions on itsgrowth. Arch. Dermatol. Syphilol. 70:49-60.
8. Kapica, L., and A. Clifford. 1968. Quantitativedetermination of yeasts in sputum. Mycopathol.Mycol. Appl. 34:27-32.
9. Kurung, J. M. 1952. The isolation of Histoplasmacapsulatum from sputum. Am. Rev. Tuberc.Pulmonary Diseases 66:578-587.
10. Littman, M. L. 1955. Liver-spleen glucose bloodagar for Histoplasma capsulatum and otherfungi. Am. J. Clin. Pathol. 25:1148-1159.
11. Lodder, J., and N. J. W. Kreger-van Rij. 1952.The yeasts. A taxonomic study, p. 33. NorthHolland Publishing Co., Amsterdam.
12. Miller, A. A., F. Ramsden, and M. R. Geake.1961. Acute disseminated histoplasmosis ofpulmonary origin probably contracted inBritain. Thorax 16:388-394.
13. Orie, N. G. M. 1952. Candida(Monilia) infectionof the respiratory tract. Diseases Chest 22:107-117.
14. Pine, L. 1955. Studies on the growth of Histo-plasma capsulatum. II. Growth of the yeastphase on agar media. J. Bacteriol. 70:375-381.
15. Raftery, A., and F. W. Hartman. 1951. Adjunctsto mycological diagnosis. Am. J. Clin. Pathol.21:133-138.
16. Salvin, S. B. 1947. Cultural studies of the yeast-like phase of Histoplasma capsulatum Darling.J. Bacteriol. 54:655-660.
17. Skinner, C. E., and D. W. Fletcher. 1960. Areview of the genus Candida. Bacteriol. Rev.24:397-416.
18. Thjotta, T., and B. J. Torheim. 1955. Studies onthe fermentation of sugars in the Candidagroup. Acta Pathol. Microbiol. Scand. 36:237-249.
2176 J. BACTERIOL.
on April 8, 2019 by guest
http://jb.asm.org/
Dow
nloaded from





























