Embed Size (px)
Citation preview

No increased trapping of multipotentmesenchymal stromal cells in bone marrow filters
compared with other bone marrow cells
M Sundin1, M Remberger1,2, H Lonnies1, B Sundberg1, O Ringden1,2 and
K Le Blanc1,3
1Division of Clinical Immunology, Department of Laboratory Medicine, Karolinska Institutet, and 2Centre for Allogeneic Stem Cell
Transplantation, and 3Hematology Centre, Karolinska University Hospital Huddinge, Stockholm, Sweden
Background
Multipotent mesenchymal stromal cells (MSC) are candidates for
cellular therapy in regenerative medicine and as treatment of graft-
versus-host-disease (GvHD) after hematopoietic stem cell (HSC)
transplantation. It has been suggested that MSC may be trapped in
bone marrow (BM) filters during the stem cell procurement and lost
from the HSC graft.
Methods
We investigated filtered BM and filters from six HSC donors. MSC
were expanded from the two sources and investigated by flow cytometry,
doubling capacity, differentiation ability and suppression in mixed
lymphocyte cultures.
Results
A range of 0.3�3.4% cells was trapped in the filters. By flow cytometry,
there was no difference in the proportions of different cell types between
the filter-retrieved and filtered BM cells. The phenotype, immuno-
suppressive capacity, differentiation and growth were equal in MSC
expanded from the two cell sources.
Discussion
Given the low number of trapped cells, filters do not appear to be a good
source of MSC. When intended for clinical transplantation, MSC need
to be expanded ex vivo to achieve sufficient doses for a clinical effect.
Keywords
graft-versus-host disease, immunosuppression, stem cell.
IntroductionMultipotent mesenchymal stromal cells (MSC) [1], pre-
viously referred to as mesenchymal stem cells, have been
used to improve the outcome of hematopoietic stem cell
transplantation (HSCT) [2,3]. Reversion of therapy-resis-
tant grade IV acute graft-versus-host-disease (GvHD) of
the skin, gastrointestinal tract and liver and prolonged
survival have been seen after treatment with MSC [4,5].
The cells may have a wider role in regenerative medicine
and treatment of autoimmune disorders because of in vitro
proliferative capacity, ability to differentiate into more
mature lineages and immunosuppressive features [6�12].MSC may be regarded as ‘universal donors’ because they
seem to avoid rejection and reduce alloreactivity, despite
expression of HLA class I and even when induced to
express HLA class II [2,8].
In HSCT the cell dose is known to play an important
role and it has been shown that bone marrow (BM) grafts
rich in cells are associated with a better outcome [13�19].It is unclear whether this results from hematopoietic or
non-hematopoietic cells in the graft. Non-hematopoietic
cells, which support long-term hematopoiesis, commonly
form complexes with hematopoietic cells. These com-
plexes are trapped in marrow filters during the procure-
ment [20,21]. Vicente et al. [22] detached cells trapped in
BM filters and infused them into HSCT recipients
along with the transplant, which resulted in reduced
acute GvHD and transplantation-related mortality. It was
Correspondence to: Dr Mikael Sundin, MD, Division of Clinical Immunology, F79, Karolinska Institutet, Karolinska University Hospital
Huddinge, SE-141 86 Stockholm, Sweden. E-mail: [email protected].
Cytotherapy (2008) Vol. 10, No. 3, 238�242
– 2008 ISCT DOI: 10.1080/14653240801965164

postulated that the retrieved cells became an additional
dose of MSC, resulting in reduced acute GvHD and
enhanced cell counts post-transplantation.
In the present study, we investigated whether the cells
trapped in BM filters display MSC features and therefore
could explain the above findings. Cells were cultured,
induced to differentiate, assayed by flow cytometry and
used in mixed lymphocyte cultures (MLC).
MethodsBM and filter cells
In the operating room, BM was harvested from six healthy
donors for clinical purposes. Donors had given informed
consent and our study was approved by the regional ethics
review board. Aspirated BM was injected into transfusion
bags through a 200-mm filter made of a co-polymer
(Green line B83; CODAN Medizinische Gerate GmbH &
Co. KG, Lensahn, Germany). An aliquot of the filtered
BM was withdrawn for this study and the rest was used in
clinical transplantation. BM filters were collected and
filled with 2 mM EDTA (Invitrogen, Paisley, UK) diluted
in phosphate-buffered saline (PBS) at 88C. Thereafter thefilters were shaken, the fluid aspirated and the filters
refilled three times. After this procedure the filters
appeared clean compared with the yellowish color before.
Filtered BM and filter-retrieved cells were counted in
Turk staining.
MSC isolation and culture
MSC were isolated and cultured from filtered BM and
cells retrieved from filters as previously described in detail
[8]. Briefly, mononuclear cells were separated and cultured
in medium supplemented with 10% fetal calf serum (FCS;
Invitrogen). The serum lot was selected on the basis of
optimal MSC growth, with maximal retention of osteo-
genic, chondrogenic and adipogenic differentiation. The
cell doubling time was calculated using the following
equation: doubling time�t/(log2 (y/x)), where t�time in
culture, y�cell count at confluence and x�cell count at
start.
Flow cytometric characterization of
filtered marrow cells, filter-retrieved cells
and isolated MSC
Filtered BM cells, filter-retrieved cells and isolated
third-passage MSC were assayed by flow cytometry.
The cells were incubated with monoclonal antibodies
against CD3, CD14, CD34, CD45 and CD73 (BD
Biosciences, San Jose, CA, USA), CD29 (Coulter, Miami,
FL, USA), CD105 (DakoCytomation, Carpentaria, CA,
USA) and CD166 (Serotec, Oxford, UK). Non-specific
fluorescence was determined using equal aliquots of cell
preparation incubated with isotype controls (BD Bios-
ciences). Finally, the cells were assayed in a flow
cytometer and analyzed with Cellquest software (BD
Biosciences). Fluorescence signals from 10 000 cells were
counted and recorded.
Differentiation ability of MSC isolated from
filtered BM and filter-retrieved cells
The ability of second-passage MSC, isolated from BM and
filter-retrieved cells, to differentiate into the more mature
cells, i.e. osteogenic and adipogenic cell types, was
investigated as described previously [10]. The MSC
differentiated into osteogenic and adipogenic cells were
stained with alizarin red S staining and oil red O solution
(Sigma-Aldrich, St Louis, MO, USA), respectively, and
morphologically examined by microscopy. Thereafter, the
staining were dissolved in 10% cetylpryidium chloride and
100% isopropanol, and assayed at 562 nm and 500 nm in a
spectrophotometer, respectively.
Inhibition of lymphocyte proliferation by MSC
isolated from filtered BM and filter-retrieved
cells
Peripheral blood lymphocytes (PBL) were isolated from
volunteer donors and used in MLC, to determine the
ability to suppress lymphocyte proliferation by addition of
MSC from filtered BM and filter-retrieved cells. Isolation
of PBL and MLC was performed as described elsewhere
[23,24].
Statistics
All comparisons between the cells, filtered BM or filter-
retrieved, and MLC with and without MSC, were tested
using a t-test in Statistica software (Statsoft Inc., Tulsa, OK,
USA).
Results and discussionSmall amounts of cells were trapped in
the BM filters
A mean of 0.09�109 (range 0.02�0.182) nucleated cells
was retrieved from the BM filters, which averaged 1.1%
(range 0.3�3.4) of the total amount of aspirated nucleated
No increased trapping of MSC in filters 239

cells. The filter-retrieved cells were 68% (median, range
56�82) mononucleated. Two of the pediatric donors
exhibited the highest cell doses retrieved, whereas the
other donors showed comparable results (Table 1). It
was evident that some BM cells were trapped in the
transfusion-set filters during the procurement. More cells
seemed to be retrieved from filters of pediatric donors,
which may be because pediatric BM is richer in cells than
adult BM.
Equal distribution of cell types in filtered
BM and filter-retrieved cells
The cells were analyzed in three regions according to size
and thereafter expression of the above-mentioned surface
markers. A high expression of hematopoietic markers
(CD3, CD14, CD34 and CD45) was shown in comparable
levels for filtered BM and filter-retrieved cells. In both
groups, cells negative for hematopoietic markers were
found in equal amounts. Cells negative for hematopoietic
markers but positive for unspecific MSC markers (CD73,
CD105 and CD166) could be demonstrated at equal and
small numbers in both filtered BM cells and cells retrieved
from filters (data not shown). The recent report by Vicente
et al. [22] suggested that MSC were trapped in BM filters
to a greater extent compared with other BM cells. Our
study could not confirm that MSC were more prone to be
trapped, as the expression of surface markers was compar-
able regardless of the source. MSC size is 30 mm versus
filter pores of 200 mm, so MSC should pass through filters
when they are not in complex formations. However, the
MSC may bind the highly polar plastic in our filters
because the cells are adhesive and consist of proteins
(Professor M. Hedenqvist, personal communication). To
minimize the potential loss of cells, which could be
beneficial in the case of ‘poor marrow’ [14], filters of less
polar materials should be used.
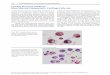
No differences in MSC phenotype
Characterized by flow cytometry, culture-expanded MSC
isolated from filtered BM and filters uniformly expressed
CD73, CD90 and CD105. The cells did not express the
hematopoietic markers CD3, CD14, CD34 and CD45. No
differences between the two groups could be demonstrated
(Figure 1A).
No statistically significant differences in the doubling
time during MSC culture could be demonstrated between
MSC isolated from filtered BM and filter-retrieved cells.
The doubling time was c. 4 and c. 6 days for both groups in
first and second passages, respectively. In the third passage,
the doubling time was c. 4 and c. 8 days for MSC isolated
from filter-retrieved cells and filtered BM, respectively
(data not shown). Doubling time increasing over time is
commonly seen.
The ability of the MSC from the two sources to
differentiate was equal (data not shown). In the absorbance
analysis of osteogenic and adipogenic differentiation, cells
isolated from BM filters exhibited values of 4.6 and 4.4,
compared with 4.3 and 3.3, respectively, for cells isolated
from filtered marrow (p�0.05). MSC from both sources
was immunosuppressive in vitro, as addition to MLC
significantly suppressed lymphocyte proliferation (Figure
1B).
The effect of reduced GvHD after infusion of filter-
retrieved cells, as reported by Vicente et al. [22], may be
explained by trapping of large doses or particularly
immunosuppressive MSC. Our results indicate that MSC
are not trapped to a greater extent than other cells in BM
filters. Therefore, a higher incidence of GvHD in
transplantation of filtered BM cannot be explained by
Table 1. Details of donors and cells
MSC 82 84 86 89 91 93
Donor (sex, male or female/age, in years) M/1 F/19 M/7 M/45 F/55 M/4
Number of filters 1 3 1 4 5 1
Total cell dose aspirated �109 5.1 18.7 7.4 12.6 19 6.8
Total cell dose retrieved �109 0.172 0.182 0.02 0.035 0.084 0.061
Percentage cells trapped in filter 3.4 1 0.3 0.3 0.4 0.9
Mononucleated cell dose retrieved�109 0.1 0.5 0.01 0.02 0.06 0.05
Percentage mononucleated cells retrieved 71 72 57 56 74 82
240 M Sundin et al.
loss of MSC. We have also demonstrated that MSC
isolated from filtered BM and filter-retrieved cells do
not differ in phenotype (expression of surface markers,
doubling time, differentiation capacity). Immunosuppres-
sion in MLC was equally potent using the MSC isolated
from the two sources, indicating that the trapped MSC do
not posses enhanced MSC features.
BM filters are not a good source of MSC
As MSC are present in low numbers in human BM and as
only small amounts of cells are trapped in BM filters, it
appears unlikely that MSC are responsible for the
beneficial effect of giving filter-retrieved cells to HSCT
recipients. To completely rule out possible MSC involve-
ment, unique surface markers that allow prospective
isolation of MSC are needed. However, to reduce GvHD
in HSCT larger numbers of cells seem to be needed and
therefore clinical ex vivo expansion of MSC, from BM
aspirations, should be applied [3,5].
AcknowledgementsThis study was supported by unrestricted grants from the
Swedish Cancer Society (Katarina Le Blanc, 4562-B05-
05XCC; Olle Ringden, 0070-B06-20XBC), the Children’s
Cancer Foundation (Katarina Le Blanc, 05/007; Olle
Ringden, 06/094), the Swedish Research Council (Katarina
Le Blanc, K2006-32X-14716-04-1, K2005-32P-15457-01A;
Olle Ringden, K2007-64X-05971-27-1), the Tobias Foun-
dation (Katarina Le Blanc), the Cancer Society in Stock-
holm (Katarina Le Blanc and Olle Ringden), the Swedish
Society of Medicine (Katarina Le Blanc), the Stockholm
County Council (Katarina Le Blanc), the Sven and Ebba-
Christina Hagbergs Foundation (Katarina Le Blanc),
the Cancer and Allergy Foundation (Olle Ringden), the
Signe och Olof Wallenius Foundation (Mikael Sundin), the
Foundation Sigurd och Elsa Goljes Minne (Mikael
Sundin), the Blodcancerfonden (Mikael Sundin) and the
Claes Hogman’s SAGMAN-scholarship/Fenwal Blood
Technologies Inc. (Mikael Sundin) and Karolinska
Institutet (Mikael Sundin, Katarina Le Blanc and Olle
Ringden).
References
1 Horwitz EM, Le Blanc K, Dominici M et al. Clarification of the
nomenclature for MSC: the International Society for Cellular
Therapy position statement. Cytotherapy 2005;7:393�5.2 Le Blanc K, Pittenger M. Mesenchymal stem cells: progress
toward promise. Cytotherapy 2005;7:36�45.3 Le Blanc K, Ringden O. Immunomodulation by mesenchymal
stem cells and clinical experience. J Int Med 2007;262:509�25.
Figure 1. Flow cytometric analysis of MSC expanded from filtered
and filter-retrieved BM cells and suppression of lymphocyte prolifera-
tion. (A) The cells from the two sources did not differ in expression of
surface markers. Dashed lines indicate isotype control. Representative
histograms are shown. (B) MSC expanded from filtered and filter-
retrieved BM cells significantly (PB0.05) inhibited lymphocyte
proliferation to allogeneic cells in MLC. There was no significant
(NS) difference in suppression between MSC expanded from the two
different sources. The dark gray bar indicates the proliferation in
MLC without MSC. Light gray bars indicate proliferation after
addition of MSC. Mean proliferation9SEM in the different sets.
No increased trapping of MSC in filters 241

4 Le Blanc K, Rasmusson I, Sundberg B et al. Treatment of severe
acute graft-versus-host disease with third party haploidentical
mesenchymal stem cells. Lancet 2004;363:1439�41.5 Ringden O, Uzunel M, Rasmusson I et al. Mesenchymal stem
cells for treatment of therapy-resistant graft-versus-host disease.
Transplantation 2006;81:1390�7.6 Bruder SP, Jaiswal N, Haynesworth SE. Growth kinetics, self-
renewal, and the osteogenic potential of purified human
mesenchymal stem cells during extensive subcultivation and
following cryopreservation. J Cell Biochem 1997;64:278�94.7 Friedenstein AJ, Petrakova KV, Kurolesova AI et al. Heterotopic
of bone marrow. Analysis of precursor cells for osteogenic and
hematopoietic tissues. Transplantation 1968;6:230�47.8 Le Blanc K, Tammik C, Rosendahl K et al. HLA expression and
immunologic properties of differentiated and undifferentiated
mesenchymal stem cells. Exp Hematol 2003;31:890�6.9 Pittenger MF, Mackay AM, Beck SC et al. Multilineage potential
of adult human mesenchymal stem cells. Science 1999;284:143�7.10 Prockop DJ. Marrow stromal cells as stem cells for nonhema-
topoietic tissues. Science 1997;276:71�4.11 Tyndall A, Walker UA, Cope A et al. Immunomodulatory
properties of mesenchymal stem cells: a review based on an
interdisciplinary meeting held at the Kennedy Institute of
Rheumatology Division, London, UK, 31 October 2005. Arth
Res Ther 2005;9:301.
12 Giordano A, Galderisi U, Marino IR. From the laboratory bench
to the patient’s bedside: an update on clinical trials with
mesenchymal stem cells. J Cell Physiol 2007;211:27�35.13 Dominietto A, Raiola AM, van Lint MT et al. Factors influencing
haematological recovery after allogeneic haemopoietic stem cell
transplants: graft-versus-host disease, donor type, cytomegalo-
virus infections and cell dose. Br J Haematol 2001;112:219�27.14 Gorin NC, Labopin M, Rocha V et al. Marrow versus peripheral
blood for geno-identical allogeneic stem cell transplantation in
acute myelocytic leukemia: influence of dose and stem cell
source shows better outcome with rich marrow. Blood
2003;102:3043�51.15 Sierra J, Storer B, Hansen JA et al. Transplantation of marrow
cells from unrelated donors for treatment of high-risk acute
leukemia: the effect of leukemic burden, donor HLA-matching,
and marrow cell dose. Blood 1997;89:4226�35.16 Storb R, Prentice RL, Thomas ED. Marrow transplantation for
treatment of aplastic anemia. An analysis of factors associated
with graft rejection. New Engl J Med 1977;296:61�6.17 Bortin MM, Gale RP, Kay HE et al. Bone marrow transplantation
for acute myelogenous leukemia. Factors associated with early
mortality. JAMA 1983;249:1166�75.18 Ringden O, Nilsson B. Death by graft-versus-host disease
associated with HLA mismatch, high recipient age, low marrow
cell dose, and splenectomy. Transplantation 1985;40:39�44.19 Paulin T. Importance of bone marrow cell dose in bone marrow
transplantation. Clin Transplant 1992;6:48�54.20 Blazsek I, Liu XH, Anjo A et al. The hematon, a morphogenetic
functional complex in mammalian bone marrow, involves
erythroblastic islands and granulocytic cobblestones. Exp Hematol
1995;23:309�19.21 Blazsek I, Misset JL, Benavides M et al. Hematon, a multicellular
functional unit in normal human bone marrow: structural
organization, hemopoietic activity, and its relationship to
myelodysplasia and myeloid leukemias. Exp Hematol
1990;18:259�65.22 Vicente D, Podesta M, Pitto A et al. Progenitor cells trapped in
marrow filters can reduce GvHD and transplant mortality. Bone
Marrow Transplant 2006;38:111�7.23 Moller G. Induction of DNA synthesis in human lymphocytes:
interaction between non-specific mitogens and antigens. Immu-
nology 1970;19:583�98.24 Sundin M, Orvell C, Rasmusson I et al. Mesenchymal stem cells
are susceptible to human herpesviruses, but viral DNA cannot
be detected in the healthy seropositive individual. Bone Marrow
Transplant 2006;37:1051�9.
242 M Sundin et al.