Embed Size (px)
Citation preview

BASIC RESEARCH www.jasn.org
NFkB-Induced Periostin Activates Integrin-b3Signaling to Promote Renal Injury in GN
Niki Prakoura,* Panagiotis Kavvadas,* Raphaёl Kormann,*† Jean-Claude Dussaule,*†‡
Christos E. Chadjichristos,*† and Christos Chatziantoniou*†
*Institut National de la Santé Et de la Recherche Médicale, Unité Mixte de Recherche Scientifique 1155, TenonHospital, Paris, France; †Sorbonne Université, Université Pierre-et-Marie-Curie Paris 6, Paris, France; and‡Department of Physiology, Saint-Antoine Hospital, Assistance Publique-Hôpitaux de Paris, Paris, France
ABSTRACTDe novo expression in the kidney of periostin, a protein involved in odontogenesis and osteogenesis, hasbeen suggested as a biomarker of renal disease. In this study, we investigated the mechanism(s) of inductionand the roleofperiostin in renal disease. Using a combination ofbioinformatics, reporter assay, and chromatinimmunoprecipitation analyses, we found that NFkB and other proinflammatory transcription factors induceperiostin expression in vitro and that binding of these factors on the periostin promoter is enriched in glo-meruli during experimental GN. Mice lacking expression of periostin displayed preserved renal function andstructureduringGN.Furthermore,delayedadministrationofperiostin antisenseoligonucleotides inwild-typeanimals with GN reversed already established proteinuria, diminished tissue inflammation, and improvedrenal structure. Lack of periostin expression also blunted the de novo renal expression of integrin-b3 andphosphorylation of focal adhesion kinase and AKT, known mediators of integrin-b3 signaling that affect cellmotility and survival, observed during GN in wild-type animals. In vitro, recombinant periostin increased theexpression of integrin-b3 and the concomitant phosphorylation of focal adhesion kinase and AKT in podo-cytes. Notably, periostin and integrin-b3 were highly colocalized in biopsy specimens from patients withinflammatoryGN. These results demonstrate that interplay between periostin and renal inflammationorches-trates inflammatory and fibrotic responses, driving podocyte damage through downstream activation ofintegrin-b3 signaling. Targeting periostin may be a novel therapeutic strategy for treating CKD.
J Am Soc Nephrol 28: 1475–1490, 2017. doi: https://doi.org/10.1681/ASN.2016070709
CKD is amajor burdenworldwidewith currently noavailable early prognostic marker or efficient ther-apeutic treatment. Although various stimuli canlead to CKD, there are common mechanisms char-acterized by chronic inflammation and accumulationof extracellular matrix within the kidney, resulting indecline of renal function and finally, ESRD.
Periostin is a matricellular protein abundant inperiosteum, periodontal ligament, bone, skin, and de-veloping heart.1,2 Periostin interacts with extracellularmatrix components, including collagen I, tenascin-C,and fibronectin, promoting collagen fibrillogenesisand increased strength performance of the extracellu-larmatrix.3–5 Interaction of periostinwith cell-surfaceintegrins avb3 and avb5 has been associated with in-creased adhesion and migration of cancer cells andvascular smooth muscle cells (SMCs).6,7 Periostin is
highly induced in vitro by TGF-b1,1,5,8 and it was alsofound to be upregulated by angiotensin II in cardiacfibroblasts.8 IL-4 and IL-13 increased periostin in lungfibroblasts and bronchial epithelial cells, associatingits induction with asthma.9
Periostin has been found to be upregulated inrenal disease models and kidney biopsy specimens.
Received July 1, 2016. Accepted October 31, 2016.
Published online ahead of print. Publication date available atwww.jasn.org.
Correspondence: Dr. Christos Chatziantoniou, Institut Nationalde la Santé et de la Recherche Médicale, Unité Mixte de Re-cherche Scientifique 1155, Tenon Hospital, 4 rue de la Chine,75020 Paris, France. Email: [email protected]
Copyright © 2017 by the American Society of Nephrology
J Am Soc Nephrol 28: 1475–1490, 2017 ISSN : 1046-6673/2805-1475 1475

It was found highly overexpressed in experimental polycystickidney disease promoting cyst growth and fibrosis.10 Periostintissue expression and urinary levels were inversely correlatedwith renal function in patients with lupus nephritis and type 2diabetes, respectively.11,12 Periostin mRNA expression washighly upregulated in both glomeruli and tubulointerstitiumin patients with different nephropathies.13 We previously re-ported that periostin expression is highly correlated with dis-ease progression in a model of hypertensive nephropathy, andwe subsequently showed that mice lacking periostin are pro-tected against the structural alterations induced by unilateralureteral obstruction.14,15
Despite these findings, the mechanisms of induction andfunction of periostin in renal disease are largely unknown. Inthis study we investigated these mechanisms in an establishedchronic renal disease model induced by nephrotoxic serum(NTS) administration. We found that periostin is inducedby key proinflammatory transcription factors, with the majorfactor being NFkB, and showed that periostin colocalizes withintegrin-b3 at sites of renal damage driving activation ofp-FAK– and p-AKT–dependent pathways. Finally, using apharmacogenetic approach of in vivo antisense administrationafter severe proteinuria and the disease onset, we showed thatinhibition of periostin can be used as a therapeutic strategy toalleviate renal disease progression. To our knowledge, we de-scribe for the first time how periostin is induced tomediate theprogression of renal disease and underline periostin targetingas therapeutic treatment against CKD.
RESULTS
Periostin Is Induced by Proinflammatory TranscriptionFactors in the Model of NTS-Induced GNTo find transcription factors that may induce periostin expres-sion, we performed bioinformatics analysis of a 2-kb region ofperiostin promoter using the Genomatix software.We focusedon known proinflammatory and profibrotic transcription fac-tors and mapped their predicted binding sites on the periostinpromoter sequence (Figure 1A).
To examine the efficiency of these transcription factors toaffect periostin expression, we performed luciferase assaysusing 2 and 1 kb constructs of human or mouse periostinpromoter, together with expression plasmids of these tran-scription factors. The large subunit of NFkB, p65, highlyinduced the activity of both human periostin promoter con-structs and, even more impressively, that of mouse promoterconstructs (Figure 1B). Other proinflammatory transcriptionfactors (c-Jun, Stat1a, and Stat6) induced a two-fold increasein the activity of both human and mouse constructs (Figure1C). In contrast, from the group of profibrotic transcriptionfactors, only Smad4 induced a sustained two-fold promoteractivity with Smad3 and Smad5, resulting in a much smallerinduction of promoter activity (Figure 1D). The increase ofperiostin mRNA paralleled the promoter activation after
transfection with the transcription factor plasmids, confirm-ing the results of the luciferase assay (Figure 1E).
We performed chromatin immunoprecipitation (ChIP) ex-periments to verify the in vivo binding of these transcriptionfactors on periostin promoter in a chronic renal diseasemodel, theNTS-induced GN. Indeed, p65was highly enrichedon its respective binding sites on periostin promoter after NTSadministration (Figure 1F). p-c-Jun and p-Stat1 were also en-riched on several of their predicted binding sites on periostinpromoter in the NTS group (Figure 1, G and H). On the con-trary, p-Smad3 was not enriched on periostin promoter afterNTS, in agreement with its low efficiency to induce periostinpromoter activity or mRNA expression (Figure 1I).
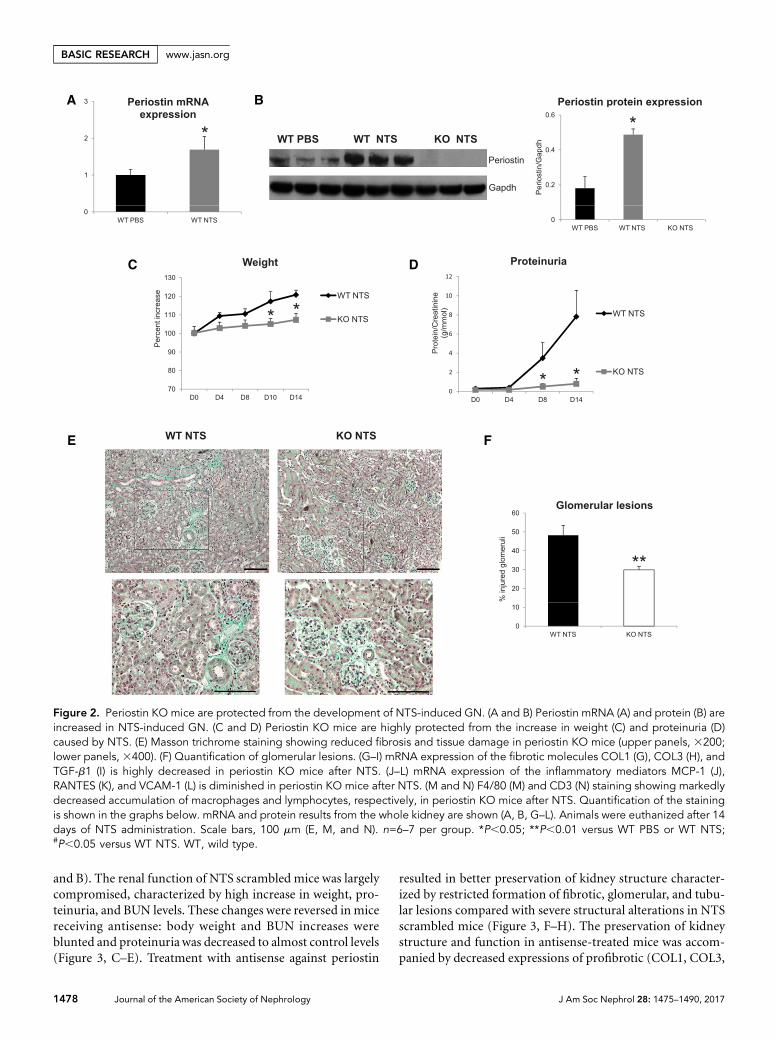
Genetic Deletion of Periostin Preserves RenalStructure and Function in the NTS ModelWe used periostin null mice in the C57BL/6 background andconfirmedwhether periostin expressionwas inducedafterNTSadministration. Transcriptional and protein analysis showedincreased renal expression of periostin in wild-type NTS micecompared with control littermates treated with PBS, whereasknockout (KO) mice did not display any periostin expression(Figure 2, A and B). Next, we analyzed functional and struc-tural alterations indicative of disease progression. The increasein weight caused by edema and the severe increase of protein-uria were markedly attenuated in KO mice, showing that lackof periostin resulted in preservation of kidney function (Fig-ure 2, C and D). Histologic examination of kidney sectionsstained with Masson trichrome showed that mice lacking per-iostin developed less fibrotic lesions and had highly preservedkidney structure together with decreased number of injuredglomeruli (Figure 2, E and F).
We next examined the expression of known proinflamma-tory mediators and fibrotic molecules, as well as the inflam-matory cell infiltration profile of the diseased kidneys. Bothcollagens I and III and profibrotic TGF-b1were upregulated inwild-type NTS mice, but their expression was highly attenu-ated in KO mice (Figure 2, G–I). In parallel, the induction ofproinflammatory mediators, such as MCP-1, Rantes, andVCAM-1, inwild-typemice after NTSwas completely bluntedin mice lacking periostin (Figure 2, J–L). Infiltration of mac-rophages and lymphocytes was markedly reduced in KO NTSmice (Figure 2, M and N), establishing a close connectionbetween periostin and inflammation probably leading to de-velopment of fibrosis and renal damage.
Delayed Inhibition of Periostin Prevents theDevelopment of Renal Disease in the NTS ModelTo assess whether inhibition of periostin can be used as ther-apeutic approach in renal disease, we inhibited periostinexpression via constant administration of antisense oligonu-cleotides (ODN) after induction of GN, and scrambled non-specific ODN was used as control.
Antisense administration in NTS mice efficiently inhibitedthe mRNA and protein upregulation of periostin (Figure 3, A
1476 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org
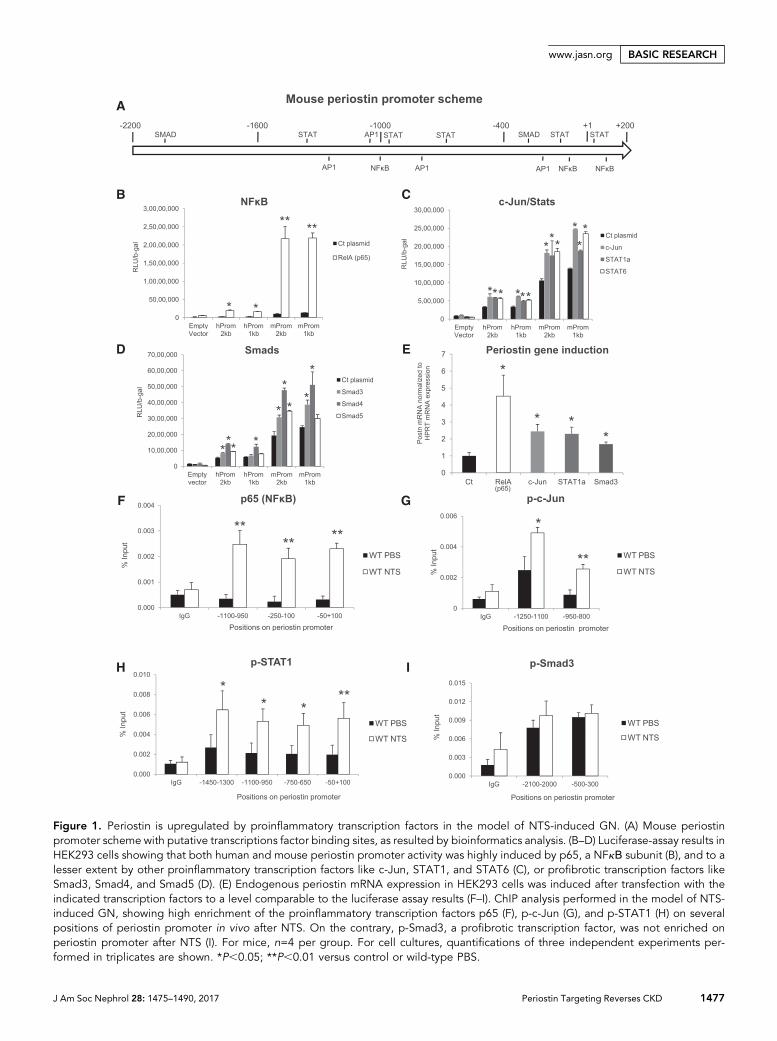
Figure 1. Periostin is upregulated by proinflammatory transcription factors in the model of NTS-induced GN. (A) Mouse periostinpromoter scheme with putative transcriptions factor binding sites, as resulted by bioinformatics analysis. (B–D) Luciferase-assay results inHEK293 cells showing that both human and mouse periostin promoter activity was highly induced by p65, a NFkΒ subunit (B), and to alesser extent by other proinflammatory transcription factors like c-Jun, STAT1, and STAT6 (C), or profibrotic transcription factors likeSmad3, Smad4, and Smad5 (D). (E) Endogenous periostin mRNA expression in HEK293 cells was induced after transfection with theindicated transcription factors to a level comparable to the luciferase assay results (F–I). ChIP analysis performed in the model of NTS-induced GN, showing high enrichment of the proinflammatory transcription factors p65 (F), p-c-Jun (G), and p-STAT1 (H) on severalpositions of periostin promoter in vivo after NTS. On the contrary, p-Smad3, a profibrotic transcription factor, was not enriched onperiostin promoter after NTS (I). For mice, n=4 per group. For cell cultures, quantifications of three independent experiments per-formed in triplicates are shown. *P,0.05; **P,0.01 versus control or wild-type PBS.
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1477
www.jasn.org BASIC RESEARCH
and B). The renal function of NTS scrambled mice was largelycompromised, characterized by high increase in weight, pro-teinuria, and BUN levels. These changes were reversed in micereceiving antisense: body weight and BUN increases wereblunted and proteinuria was decreased to almost control levels(Figure 3, C–E). Treatment with antisense against periostin
resulted in better preservation of kidney structure character-ized by restricted formation of fibrotic, glomerular, and tubu-lar lesions compared with severe structural alterations in NTSscrambled mice (Figure 3, F–H). The preservation of kidneystructure and function in antisense-treated mice was accom-panied by decreased expressions of profibrotic (COL1, COL3,
Figure 2. Periostin KO mice are protected from the development of NTS-induced GN. (A and B) Periostin mRNA (A) and protein (B) areincreased in NTS-induced GN. (C and D) Periostin KO mice are highly protected from the increase in weight (C) and proteinuria (D)caused by NTS. (E) Masson trichrome staining showing reduced fibrosis and tissue damage in periostin KO mice (upper panels, 3200;lower panels, 3400). (F) Quantification of glomerular lesions. (G–I) mRNA expression of the fibrotic molecules COL1 (G), COL3 (H), andTGF-b1 (I) is highly decreased in periostin KO mice after NTS. (J–L) mRNA expression of the inflammatory mediators MCP-1 (J),RANTES (K), and VCAM-1 (L) is diminished in periostin KO mice after NTS. (M and N) F4/80 (M) and CD3 (N) staining showing markedlydecreased accumulation of macrophages and lymphocytes, respectively, in periostin KO mice after NTS. Quantification of the stainingis shown in the graphs below. mRNA and protein results from the whole kidney are shown (A, B, G–L). Animals were euthanized after 14days of NTS administration. Scale bars, 100 mm (E, M, and N). n=6–7 per group. *P,0.05; **P,0.01 versus WT PBS or WT NTS;#P,0.05 versus WT NTS. WT, wild type.
1478 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org
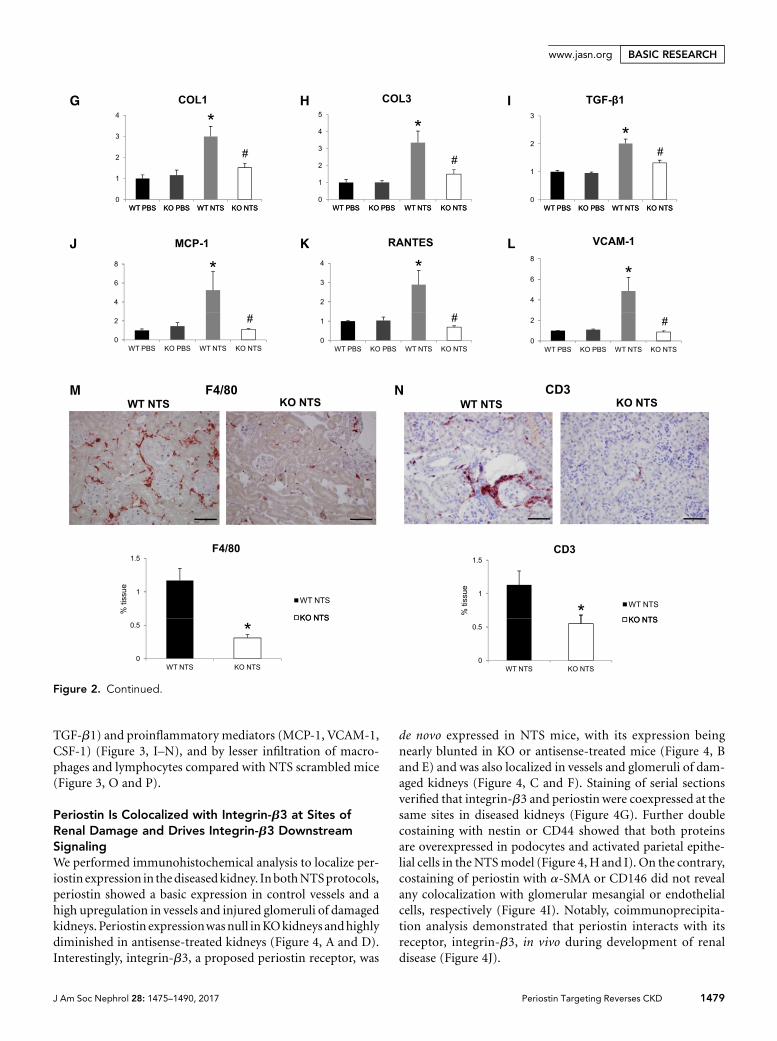
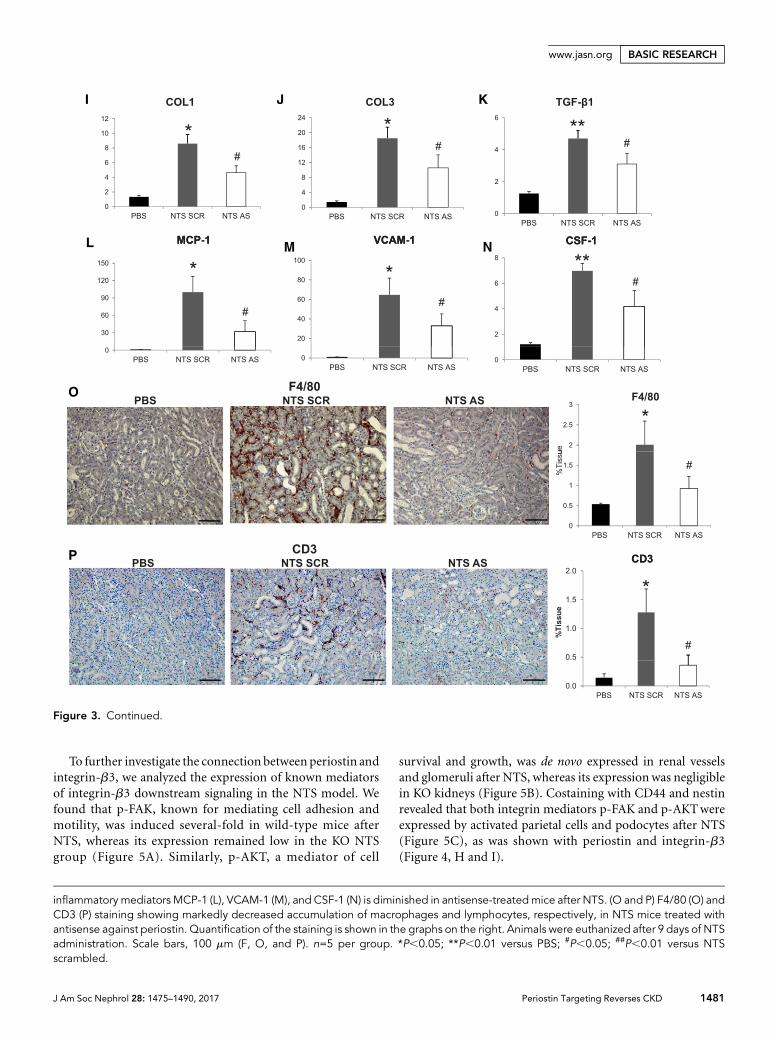
TGF-b1) and proinflammatory mediators (MCP-1, VCAM-1,CSF-1) (Figure 3, I–N), and by lesser infiltration of macro-phages and lymphocytes compared with NTS scrambled mice(Figure 3, O and P).
Periostin Is Colocalized with Integrin-b3 at Sites ofRenal Damage and Drives Integrin-b3 DownstreamSignalingWe performed immunohistochemical analysis to localize per-iostin expression in thediseasedkidney. InbothNTSprotocols,periostin showed a basic expression in control vessels and ahigh upregulation in vessels and injured glomeruli of damagedkidneys. Periostin expressionwasnull inKOkidneys andhighlydiminished in antisense-treated kidneys (Figure 4, A and D).Interestingly, integrin-b3, a proposed periostin receptor, was
de novo expressed in NTS mice, with its expression beingnearly blunted in KO or antisense-treated mice (Figure 4, Band E) and was also localized in vessels and glomeruli of dam-aged kidneys (Figure 4, C and F). Staining of serial sectionsverified that integrin-b3 and periostin were coexpressed at thesame sites in diseased kidneys (Figure 4G). Further doublecostaining with nestin or CD44 showed that both proteinsare overexpressed in podocytes and activated parietal epithe-lial cells in the NTSmodel (Figure 4, H and I). On the contrary,costaining of periostin with a-SMA or CD146 did not revealany colocalization with glomerular mesangial or endothelialcells, respectively (Figure 4I). Notably, coimmunoprecipita-tion analysis demonstrated that periostin interacts with itsreceptor, integrin-b3, in vivo during development of renaldisease (Figure 4J).
Figure 2. Continued.
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1479
www.jasn.org BASIC RESEARCH
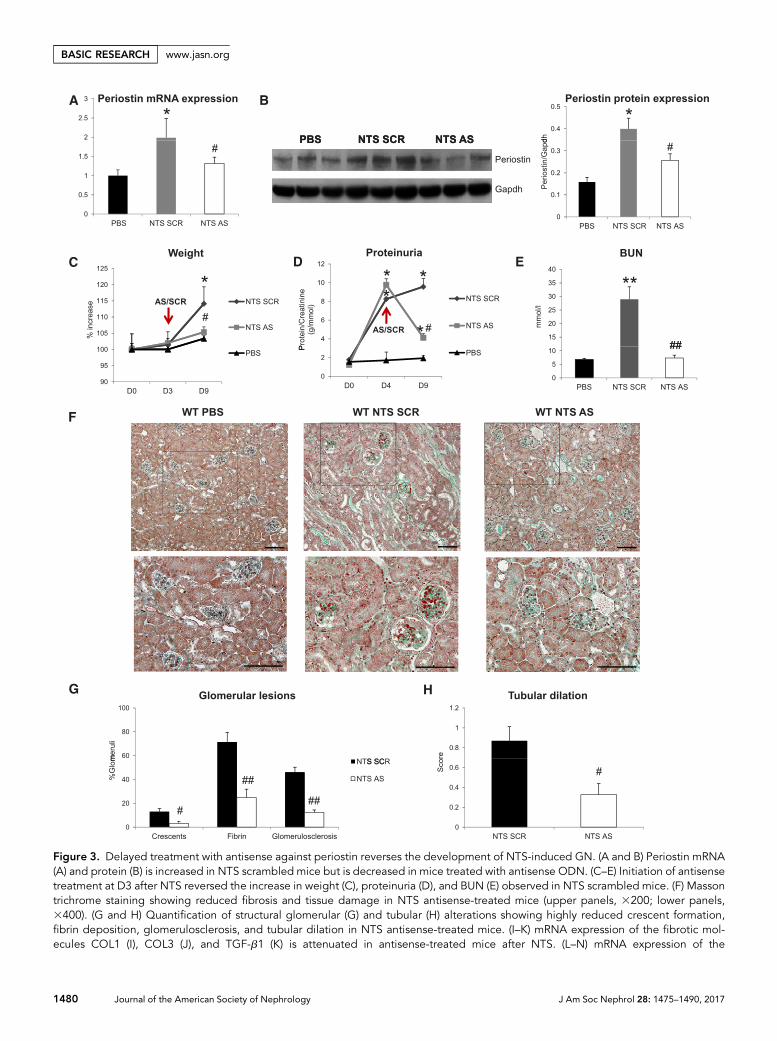
Figure 3. Delayed treatment with antisense against periostin reverses the development of NTS-induced GN. (A and B) Periostin mRNA(A) and protein (B) is increased in NTS scrambled mice but is decreased in mice treated with antisense ODN. (C–E) Initiation of antisensetreatment at D3 after NTS reversed the increase in weight (C), proteinuria (D), and BUN (E) observed in NTS scrambled mice. (F) Massontrichrome staining showing reduced fibrosis and tissue damage in NTS antisense-treated mice (upper panels, 3200; lower panels,3400). (G and H) Quantification of structural glomerular (G) and tubular (H) alterations showing highly reduced crescent formation,fibrin deposition, glomerulosclerosis, and tubular dilation in NTS antisense-treated mice. (I–K) mRNA expression of the fibrotic mol-ecules COL1 (I), COL3 (J), and TGF-b1 (K) is attenuated in antisense-treated mice after NTS. (L–N) mRNA expression of the
1480 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org
To further investigate the connection between periostin andintegrin-b3, we analyzed the expression of known mediatorsof integrin-b3 downstream signaling in the NTS model. Wefound that p-FAK, known for mediating cell adhesion andmotility, was induced several-fold in wild-type mice afterNTS, whereas its expression remained low in the KO NTSgroup (Figure 5A). Similarly, p-AKT, a mediator of cell
survival and growth, was de novo expressed in renal vesselsand glomeruli after NTS, whereas its expressionwas negligiblein KO kidneys (Figure 5B). Costaining with CD44 and nestinrevealed that both integrin mediators p-FAK and p-AKTwereexpressed by activated parietal cells and podocytes after NTS(Figure 5C), as was shown with periostin and integrin-b3(Figure 4, H and I).
inflammatory mediators MCP-1 (L), VCAM-1 (M), and CSF-1 (N) is diminished in antisense-treatedmice after NTS. (O and P) F4/80 (O) andCD3 (P) staining showing markedly decreased accumulation of macrophages and lymphocytes, respectively, in NTS mice treated withantisense against periostin. Quantification of the staining is shown in the graphs on the right. Animals were euthanized after 9 days of NTSadministration. Scale bars, 100 mm (F, O, and P). n=5 per group. *P,0.05; **P,0.01 versus PBS; #P,0.05; ##P,0.01 versus NTSscrambled.
Figure 3. Continued.
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1481
www.jasn.org BASIC RESEARCH
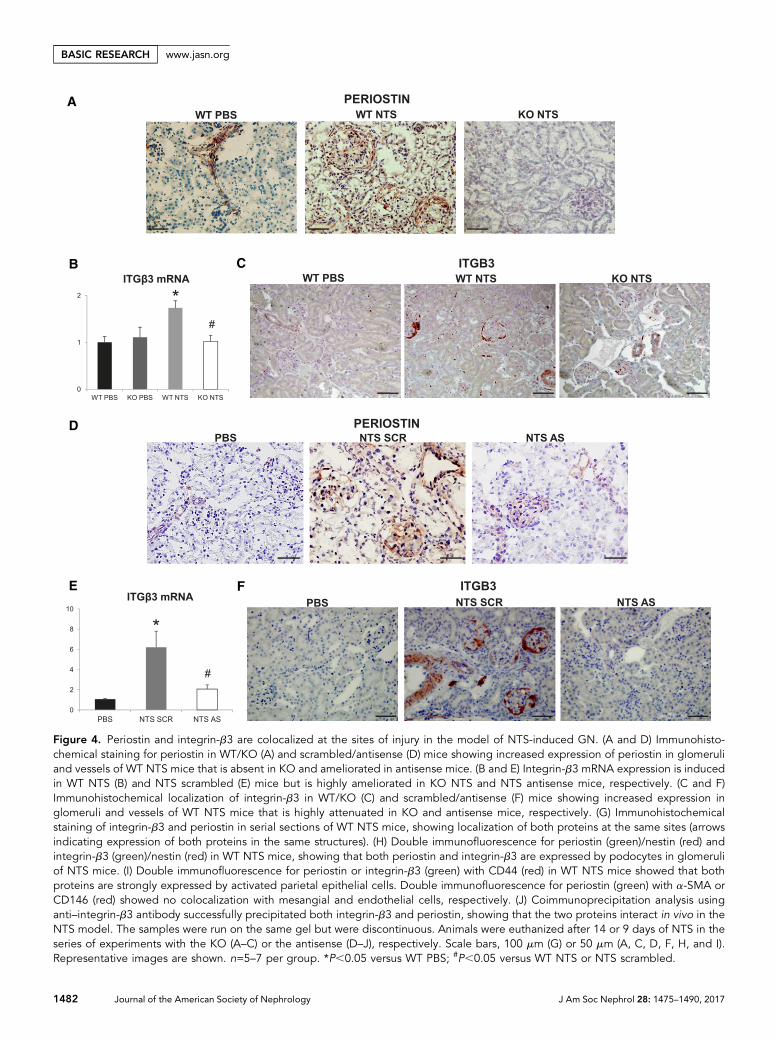
Figure 4. Periostin and integrin-b3 are colocalized at the sites of injury in the model of NTS-induced GN. (A and D) Immunohisto-chemical staining for periostin in WT/KO (A) and scrambled/antisense (D) mice showing increased expression of periostin in glomeruliand vessels of WT NTS mice that is absent in KO and ameliorated in antisense mice. (B and E) Integrin-b3 mRNA expression is inducedin WT NTS (B) and NTS scrambled (E) mice but is highly ameliorated in KO NTS and NTS antisense mice, respectively. (C and F)Immunohistochemical localization of integrin-b3 in WT/KO (C) and scrambled/antisense (F) mice showing increased expression inglomeruli and vessels of WT NTS mice that is highly attenuated in KO and antisense mice, respectively. (G) Immunohistochemicalstaining of integrin-b3 and periostin in serial sections of WT NTS mice, showing localization of both proteins at the same sites (arrowsindicating expression of both proteins in the same structures). (H) Double immunofluorescence for periostin (green)/nestin (red) andintegrin-b3 (green)/nestin (red) in WT NTS mice, showing that both periostin and integrin-b3 are expressed by podocytes in glomeruliof NTS mice. (I) Double immunofluorescence for periostin or integrin-b3 (green) with CD44 (red) in WT NTS mice showed that bothproteins are strongly expressed by activated parietal epithelial cells. Double immunofluorescence for periostin (green) with a-SMA orCD146 (red) showed no colocalization with mesangial and endothelial cells, respectively. (J) Coimmunoprecipitation analysis usinganti–integrin-b3 antibody successfully precipitated both integrin-b3 and periostin, showing that the two proteins interact in vivo in theNTS model. The samples were run on the same gel but were discontinuous. Animals were euthanized after 14 or 9 days of NTS in theseries of experiments with the KO (A–C) or the antisense (D–J), respectively. Scale bars, 100 mm (G) or 50 mm (A, C, D, F, H, and I).Representative images are shown. n=5–7 per group. *P,0.05 versus WT PBS; #P,0.05 versus WT NTS or NTS scrambled.
1482 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org
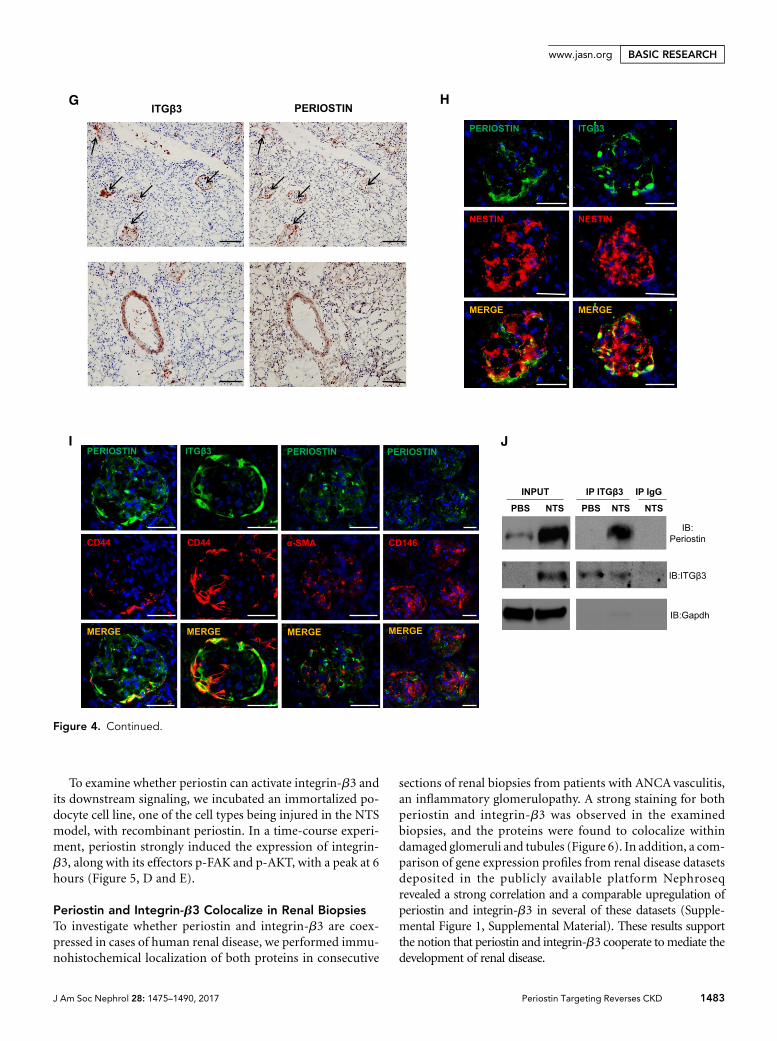
To examine whether periostin can activate integrin-b3 andits downstream signaling, we incubated an immortalized po-docyte cell line, one of the cell types being injured in the NTSmodel, with recombinant periostin. In a time-course experi-ment, periostin strongly induced the expression of integrin-b3, along with its effectors p-FAK and p-AKT, with a peak at 6hours (Figure 5, D and E).
Periostin and Integrin-b3 Colocalize in Renal BiopsiesTo investigate whether periostin and integrin-b3 are coex-pressed in cases of human renal disease, we performed immu-nohistochemical localization of both proteins in consecutive
sections of renal biopsies from patients with ANCA vasculitis,an inflammatory glomerulopathy. A strong staining for bothperiostin and integrin-b3 was observed in the examinedbiopsies, and the proteins were found to colocalize withindamaged glomeruli and tubules (Figure 6). In addition, a com-parison of gene expression profiles from renal disease datasetsdeposited in the publicly available platform Nephroseqrevealed a strong correlation and a comparable upregulation ofperiostin and integrin-b3 in several of these datasets (Supple-mental Figure 1, Supplemental Material). These results supportthe notion that periostin and integrin-b3 cooperate tomediate thedevelopment of renal disease.
Figure 4. Continued.
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1483
www.jasn.org BASIC RESEARCH
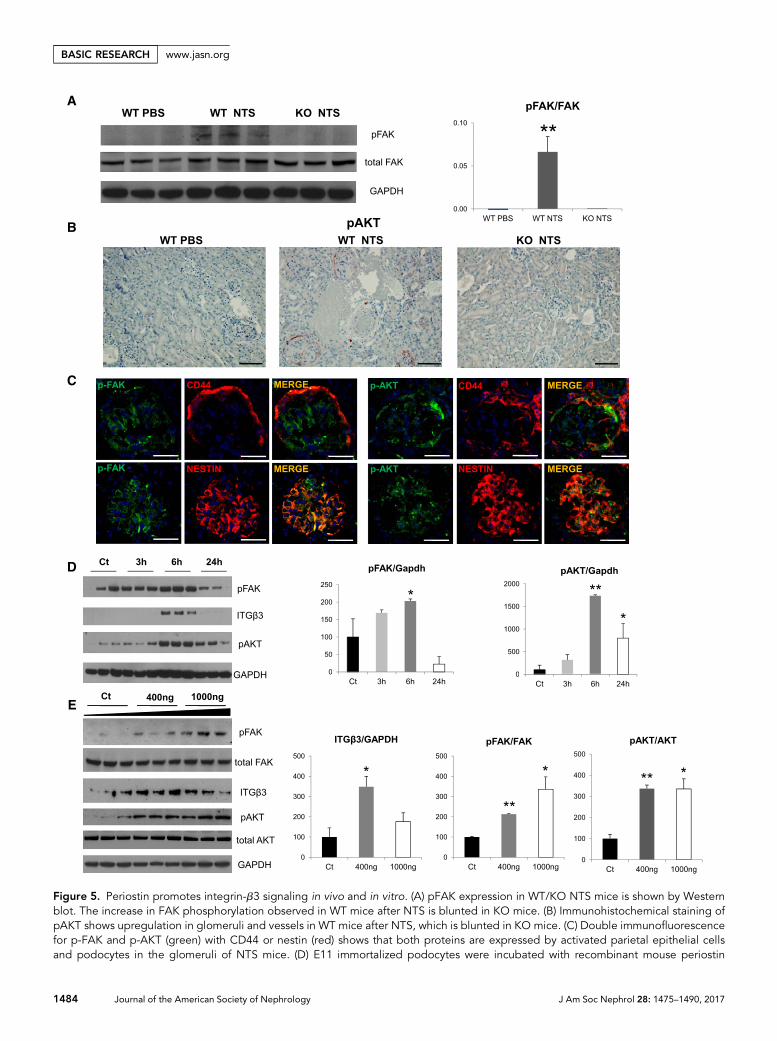
Figure 5. Periostin promotes integrin-b3 signaling in vivo and in vitro. (A) pFAK expression in WT/KO NTS mice is shown by Westernblot. The increase in FAK phosphorylation observed in WT mice after NTS is blunted in KO mice. (B) Immunohistochemical staining ofpAKT shows upregulation in glomeruli and vessels in WT mice after NTS, which is blunted in KO mice. (C) Double immunofluorescencefor p-FAK and p-AKT (green) with CD44 or nestin (red) shows that both proteins are expressed by activated parietal epithelial cellsand podocytes in the glomeruli of NTS mice. (D) E11 immortalized podocytes were incubated with recombinant mouse periostin
1484 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org

DISCUSSION
The increasing prevalence of CKD and the lack of efficient treat-ment necessitate the discovery of novel prognostic markers andtherapeutic targets. Periostin, a matricellular protein associatedwithmesenchymal phenotype and fibrous structures, was foundbyus andother investigators tobehighly induced in thekidney inseveral types of renal disease, both in animal models and patientbiopsies.16,17 Nevertheless, the mechanism(s) of induction andthe actual role of periostin during disease progression have notbeen examined. This study aimed to unravel these mechanismsin an established model of NTS-induced renal disease. Ourresults showed for thefirst time that proinflammatory transcrip-tion factors induce periostin in renal disease. Moreover, perios-tin was found to drive both the expression and downstreamsignaling of integrin-b3, a proposed periostin receptor, causingphenotypical changes associated with disease progression. Fi-nally, we showed that delayed in vivo inhibition of periostin afterthe disease onset can efficiently alleviate the progression of severerenal disease. The proposedmechanisms of induction and func-tion of periostin in renal disease are summarized in Figure 7.
Little is presently knownabout the transcription factors thatcontrol periostin gene expression. Twistwas shown tobind and
induce periostin promoter activity in preosteoblasts, whereasYY1 was found to bind on a conserved enhancer region onperiostin promoter, regulating tissue-specific periostin expres-sion in the cardiac cushion mesenchyme.18,19 Our bioinfor-matics analysis revealed the presence of several putativeproinflammatory transcription factor binding sites on perios-tin promoter, whereas transcription factor binding sites of theclassic TGF-b1 signaling pathway, which has been shown toupregulate periostin in vitro,1,5,8 were rare. In luciferase assaysand at the endogenous gene level, we showed that proinflamma-tory transcription factors such as NFkΒ (p65), c-Jun, and STAT1directly induced periostin promoter activity or gene expression.The active forms of these transcription factors were found to behighly enriched in vivo on periostin promoter in NTS-inducedGN, indicating that periostin was upregulated in response tothese transcription factors in the NTS model. The fact thatp-Smad3 was not enriched on periostin promoter implies thateither periostin is not induced by TGF-b1 in the NTS model orthat TGF-b1 may control periostin expression by an Smad-independent pathway, involving the activation of proinflamma-tory transcription factors like NFkB, Jun, and STAT.20
The induction of periostin by proinflammatory pathwayscorrelates with its observed role to promote inflammatory
(400 ng/ml) for the indicated time points and Western blot analysis was performed for integrin-b3, pFAK, pAKT, and GAPDH, withquantification graphs shown on the right. At 6 hours of periostin incubation integrin-b3 and its downstream effectors pFAK andpAKTwereupregulated. (E) E11 cells were incubated for 6 hours with increasing doses of recombinant mouse periostin (400 and 1000 ng/ml), fol-lowed by Western blot analysis of integrin-b3, pFAK, FAK, pAKT, AKT, and GAPDH. Quantifications are shown on the right. pFAK andpAKT remain elevated after incubation with increasing doses of periostin. Scale bars, 100 mm (B and C). n=6–7 mice per group. Animalswere euthanized after 14 days of NTS administration. For cell cultures, quantifications of three independent experiments performed intriplicates are shown. *P,0.05 versus control; **P,0.01 versus WT PBS or control.
Figure 6. Periostin and integrin-b3 are coexpressed in human renal biopsies. (A) Immunohistochemical staining of integrin-b3 andperiostin in consecutive sections from a biopsy specimen of a patient with ANCA vasculitis, depicting a strong expression and a highdegree of colocalization between periostin and integrin-b3 both in damaged glomeruli and tubules. (B) Magnified images of the panelsdrawn in (A), showing in more detail the coexpression of periostin and integrin-b3 in damaged glomeruli and surrounding tubules.Arrows indicate colocalization sites. Scale bars, 100 mm (A) or 30 mm (B). Representative images are shown.
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1485
www.jasn.org BASIC RESEARCH

reactions in several disease contexts. In allergen-induced skininflammation, IL-4 and IL-13 were shown to induce periostin,which further amplified Th-2 type immune responses.21 Fur-ther, periostin-deficient lung fibroblasts showed lower pro-duction of chemokines in response to TNF-a, accountingfor the lack of inflammatory response in pulmonary fibro-sis.22 In a model of glioblastoma xenografts, periostin secretedby stem cells was found to recruit tumor-associated macro-phages promoting malignant growth.23 These studies are inaccordance with our observations that lack or inhibition ofperiostin reduced the production of proinflammatory cyto-kines and the subsequent macrophage and lymphocyte infil-tration to the injured kidneys. To examine whether thedifference in the inflammatory response was because of straindifference in the activation of inflammatory cells, we com-pared the activated monocytes/macrophages of wild-typeand KO mice after thioglycolate-induced peritonitis. Numer-ation of the infiltrated peritoneal monocytes after thioglyco-late treatment did not show any differences between wild-typeand KO mice (Supplemental Figure 2A). Leukocyte cell anal-ysis before and after thioglycolate treatment did not reveal anyvariations in the leukocyte subtypes (Supplemental Figure2B). In migration assays toward chemoattractant MCP-1,the isolated peritoneal monocytes of wild-type and KO miceshowed the same migratory capacities, also explained by thesimilar expression levels of various receptors and cytokines(Supplemental Figure 2, C and D). Notably, periostin expres-sion was null in the isolated wild-type monocytes. Thus, peri-ostin expression by the damaged tissue is crucial in inducingproduction of chemokines to attract inflammatory cells at thesites of injury. Accordingly, recombinant periostin was foundto directly enhance adhesion of eosinophils and monocytes tofibronectin and function as a chemoattractant to primarymonocytes and macrophages in migration assays.24,25
Chronic inflammation plays a central role in development offibrosis and kidney damage. Concomitantly to the attenuationof inflammation, periostin deficiency diminished the expression
of collagens and TGF-b1, and improved renal structure andfunction. Periostin has been associated with fibrosis in severaltissues. Analysis of periostin expression in human skin pathol-ogies demonstrated that it is overexpressed during scarformation and associated with wound healing and pathologicremodeling.26 Accordingly, periostin null dermal fibroblastsshowed reduced expression of Acta2 and Col1a1 transcriptsupon TGF-b1 treatment,27 whereas mesenchymal cells treatedwith exogenous periostin showed increased collagen productionand wound closure.28 In amodel of acute myocardial infarction,periostin null mice exhibited impaired cardiac healing as a resultof reduced myocardial stiffness and collagen fibril formation,whereas inducible overexpression of periostin protected micefrom cardiac rupture.29,30 The ability of periostin to interactand stabilize fibrillar collagens and other extracellular matrixmolecules is probably a key event by which periostin controlsextracellular matrix deposition and development of fibrosis.3–5
We found de novo expressed periostin to colocalize and in-teract with integrin-b3 in glomeruli and vascular SMCs of NTSmice. Overexpression of periostin in aortic SMCs induced mi-gration together with increased integrin-b3 expression and FAKphosphorylation.7 In addition, periostin expression was in-creased in vascular SMCs subjected to mechanical strain, andthis increase was followed by FAK activation and MCP-1 secre-tion, which were inhibited by a periostin-neutralizing anti-body.31 In agreement with these studies, induction of periostinin the NTSmodel was determinant for the de novo expression ofintegrin-b3 followed by downstream FAK and AKT activation.Thus, periostinmay induce and activate integrin-b3, promotingmotility, permeability, and secretion of chemokines essential forremodeling and inflammatory cell infiltration.32
Activation of integrin avb3 in podocytes has been shown tomediate podocyte dysfunction, motility, and proteinuria inFSGS and LPS-induced renal disease.33,34 In the NTS model,induction of integrin-b3 and its downstream signaling in pa-rietal cells and podocytes was dependent on the expression ofperiostin. Treatment of cultured podocytes with recombinant
Figure 7. Proposed mechanism of how periostin mediates renal damage. Proinflammatory transcription factors, with the major factorbeing NFkΒ, induce expression of periostin, which activates integrin-b3, inducing integrin downstream signaling and leading to cellmotility, survival, and podocyte foot process effacement. In parallel, periostin promotes the tissue expression of several chemokines(MCP-1, RANTES, VCAM-1, and CSF-1) and profibrotic molecules (COL1, COL3, and TGF-b1), inducing inflammatory cell infiltrationand matrix stabilization, and podocyte damage further amplifies these effects, causing a vicious circle.
1486 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org

periostin was sufficient to increase integrin-b3 expression fol-lowed by FAK and AKTactivation. Activation of FAK and AKTin the injured podocytes and activated parietal cells inducescell motility, invasion,35 and crosstalk between the two glo-merular cell types, resulting ultimately in foot process efface-ment, proteinuria, and kidney damage.
We found strong expression and a high degree of colocal-ization between periostin and integrin-b3 in renal biopsiesfrom patients with ANCA vasculitis. In addition to their glo-merular expression, both proteins were also localized in tubu-lar cells in the human biopsies. This difference with the NTSmodel may reflect the distinct stimuli causing the disease,which in the case of the NTS model, target primarily the glo-merulus, whereas they affect the whole kidney in the case ofANCA vasculitis. To further investigate the coexpression pat-tern of periostin and integrin-b3 in patients with renal disease,we performed ameta-analysis of gene expression profiles fromrenal disease datasets deposited in the Nephroseq platform(www.nephroseq.org). We compared the expression levels ofperiostin and integrin-b3 in a similar disease dataset as thebiopsies used in our study (patients with vasculitis versushealthy donors, n=52), and found a strong correlationbetween periostin and integrin-b3 (R=0.54, P,0.001). More-over, both proteins were significantly upregulated to a com-parable level in several other datasets (Supplemental Figure 1).
Presently, only two studies in animal models of pulmonaryfibrosis and ovarian cancer have shown that inhibition of peri-ostin via blocking antibodies could be used as a therapy againstthe disease progression.28,36 We used an alternative strategy ofin vivo antisense administration after establishment of severeproteinuria and found that inhibition of periostin could re-markably diminish the inflammation and reverse the increasein proteinuria. The ability of periostin to play distinctive rolesin crucial events during development of renal disease by promot-ing tissue inflammatory response, accumulation of extracellularmatrix and podocyte damage underline periostin targeting as apotential efficient future treatment against CKD.
CONCISE METHODS
Immunohistochemical Staining of Patient BiopsiesAll procedures and use of human tissue were performed according to
the national ethical guidelines and were in accordance with the Dec-
laration of Helsinki. Written informed consent was given by the pa-
tients for use of part of their biopsy specimen for scientific purposes
before inclusion in the study.
Renal biopsies from patients were retrospectively analyzed. Exper-
iments were performed on biopsies from patients with ANCAvascu-
litis. The biopsies were characterized by diffuse glomerular lesions,
fibrin deposits, weak lesions of acute tubular necrosis, and extraca-
pillary crescentic proliferation. Formalin-fixed, paraffin-embedded
consecutive kidney sections (4-mm thick) were stained with primary
antibodies for periostin (ab14041; Abcam, Inc., Cambridge, MA) or
integrin-b3 (ab75872; Abcam, Inc.). Appropriate secondary
antibodies (N-Histofine; Nichirei Biosciences) and AEC (Dako) as
substrate were used. Slides were counterstained with hematoxylin QS
(Vector Laboratories, Burlingame, CA) and mounted with perma-
nent aqueous mounting medium (ScyTek). Biopsies from four pa-
tients were used for this set of experiments.
Animal ModelsAll procedures regarding animal experimentation were in accordance
with the European Union Guidelines for the Care and Use of Labo-
ratory Animals and approved by the local ethics committee of the
National Institute for Health andMedical Research (Institut National
de la Santé et de la Recherche Médicale). Animals were housed at
constant temperature with free access to water and food.
The strain of periostin KO mice was created at the laboratory of Dr.
Simon Conway and has been previously described.15 For the model of
NTS-induced GN, decomplemented NTS was prepared as previously
described.37,38 Femalewild-type andKO littermatemice of the C57BL/6
background (8–10 weeks old) received intravenous injections of 15 ml
NTS per g body wt over two consecutive days (days 0 and 1) to induce
crescentic GN. Control mice were injected with PBS. Mice were eutha-
nized 14 days after the first injection (n=6–7 per group). Blood, urine,
and renal tissues were collected for subsequent analyses.
For the antisense experiments, SV129malewild-typemice aged 8–
10weeks were used. Themice received intravenous injections of 12ml
NTS per g body wt over two consecutive days (days 0 and 1) to induce
crescentic GN, whereas control mice were injected with PBS. Perios-
tin expression was inhibited with a cocktail of two different ODNs
specifically targeting periostin mRNA, designed using the Integrated
DNATechnologies platform, also previously described.15 Scrambled
nonspecific ODNs were used as control. The ODN sequences were
modified with phosphorothioate to prevent their in vivo hydrolysis by
nucleases (Sigma-Aldrich, St. Louis, MO; Supplemental Table 1). For
administration, the ODNs were diluted in normal saline and placed
in osmotic mini-pumps (model 1002; Alzet, Cupertino, CA), which
were subsequently implanted subcutaneously, constantly releasing a
dose of 0.25 ml/h corresponding to a release of 150 pmol of ODN per
day. The implantation of mini-pumps was performed at day 3 after
administration of NTS, taking 1 day for the pump to start function-
ing. Mice were euthanized 9 days after the first injection (n=5 per
group). Blood, urine, and renal tissues were collected.
Proteinuria and BUN MeasurementsUrine was collected at several time points during the mouse protocols
and blood was collected at the end of each protocol. Proteinuria was
measured using aKonelab analyzer (ThermoFisher Scientific, Vernon
Hills, IL), and was normalized to urine creatinine. BUN levels were
measured with an enzymatic method (Konelab analyzer) and ex-
pressed in millimoles per liter.
Histologic EvaluationFormalin-fixed, paraffin-embedded, 4-mmthick kidney sectionswere
stained with Masson trichrome. Quantifications were performed by
examining at least tenmicrophotographs of random, nonoverlapping
fields at 2003 magnification per mouse. Crescents, fibrin deposits,
and glomerulosclerosis were expressed as percentage of the total
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1487
www.jasn.org BASIC RESEARCH

glomeruli evaluated. Tubular dilation was examined using a zero (no
dilation) to three (high dilation) injury scale per photograph and the
mean value was calculated for each mouse.
The evaluation of glomerular injury on the resistant C57BL/6
background was on the basis of the combined examination of me-
sangial matrix expansion, endothelial wall thickening, capillary rup-
ture, and glomerular infiltration and was expressed as percentage of
injured glomeruli per total glomeruli examined.
Western Blot AnalysisProteins from half kidneys or cell culture were extracted in a com-
mercial RIPA lysis buffer supplemented with PMSF, a protease
inhibitor cocktail and sodium orthovanadate (Santa Cruz Biotech-
nologies, Santa Cruz, CA), and total protein concentration was mea-
suredusing theBradford assay. Equal amounts of proteinswere loaded
on a NuPAGE 4%–12% gradient gel (Invitrogen, Carlsbad, CA) and
transferred to nitrocellulose membrane (Bio-Rad, Hercules, CA).
Immunoblotting was performed for periostin (MOSF20; R&D Sys-
tems, Minneapolis, MN), integrin-b3 (ab75872; Abcam, Inc.), pFAK
(#3283; Cell Signaling Technology, Danvers,MA), FAK (sc-557; Santa
Cruz Biotechnologies), pAKT (#9271; Cell Signaling Technology),
and AKT 1/2/3 (sc-8312; Santa Cruz Biotechnologies). Gapdh
(Sigma-Aldrich) was used as loading control.
Quantitative Real-Time PCRTotal RNAwas extracted from half kidneys using TRI Reagent (MRC)
and from cell cultures using the EZ-10 Spin Column Total RNAMini-
preps Super kit (Bio Basic Inc). RNAqualitywas verified bymeasuring
the OD 260:280 ratio and residual genomic DNA was removed by
DNase I treatment (Thermo Fisher Scientific) for 30minutes at 37°C.
A total of 1 mg RNAwas transcribed to cDNAusing the Maxima First
Strand cDNA Synthesis Kit from Thermo Fisher Scientific per man-
ufacturer’s instructions. Real-time PCR was performed with the Ro-
che Light Cycler 480 detection system using SYBRGreen PCRMaster
Mix (RocheDiagnostics, Indianapolis, IN). Specific primers for target
mRNAs (Supplemental Table 2) were used for amplification under the
following program: 95°C for 5 minutes, 45 cycles at 95°C for 15 seconds
and 60°C for 15 seconds, and 72°C for 15 seconds. For quantitative anal-
ysis, experimental geneswere normalized toGusbmRNAorHprtmRNA
expression using the DDCTmethod. Dissociation curves were analyzed
in order to determine that a single product was amplified.
Immunohistochemistry and DoubleImmunofluorescenceFor immunostaining of macrophages and lymphocytes, 4-mm thick
formalin-fixed, paraffin-embedded kidney sections were stained with
primary antibodies to F4/80 (MCA497R; Abd Serotec) or CD3
(A0452; Dako), respectively. Appropriate secondary antibodies
(N-Histofine; Nichirei Biosciences) and AEC (Dako) as substrate
were used. Slides were counterstained with hematoxylin QS (Vector
Laboratories) and mounted with permanent aqueous mounting me-
dium (ScyTek). The F4/80 and CD3 positive area was quantified in at
least ten photographs at 2003magnification per animal, using pub-
licly available image processing software (ImageJ; Fiji), and expressed
as percentage of the total tissue area.
For periostin immunostaining, 5-mm frozen tissue sections were
fixed in 4% PFA for 15 minutes, blocked in 10% FBS, and incubated
with primary anti-periostin antibody (PA5–34641; Thermo Fisher
Scientific). Integrin-b3 (ab75872; Abcam, Inc.) and p-AKT immu-
nostainings (ab81283; Abcam, Inc.) were performed on 4-mm
formalin-fixed, paraffin-embedded kidney sections. Incubation
with secondary antibodies and signal detection was performed as
described above.
Dual immunolabeling was performed on 5-mm frozen or 4-mm
paraffin tissue sections incubated with primary antibodies to perios-
tin, integrin-b3, nestin (556309; BD Pharmingen), CD44 (11–624-
C100; Exbio), a-SMA (A5228; Sigma-Aldrich), CD146 (kind gift
from Professor Françoise Dignat-George), pAKT and pFAK
(ab55335; Abcam, Inc.). The signal was detected with Alexa Fluor
–488 and –546 secondary antibodies followed by DAPI counterstaining
and mounting in PermaFluor medium (Thermo Fisher Scientific).
E11 Podocyte Cell CultureThe E11 immortalized mouse podocyte cell line was cultured as pre-
viously described.17 Briefly, cells were maintained in RPMI 1640
medium supplemented with 10% FBS, 100 U/ml penicillin streptomy-
cin, and 10 U/ml recombinant mouse INFg (Peprotech) to induce syn-
thesis of the immortalizing T antigen at 33°C and 5% CO2. To initiate
differentiation, cells were trypsinized and thermoshifted to 37°C in me-
dium without INFg for 10–15 days. For treatment with recombinant
mouse periostin, differentiated cells in six-well plates were serum starved
for 24 hours and incubated with recombinant periostin (R&D Systems)
at a concentration of 400 or 1000 ng/ml for the indicated time points.
Cell Culture and TransfectionThe HEK293 cell line was maintained in DMEM 4.5 g/L glucose supple-
mented with 10% FBS, 100 U/ml penicillin streptomycin, and 2 mM
L-glutamine at 37°C/5% CO2. Transfection was performed in 12-well
plates via the CaCl2 method. Briefly, 2 hours before transfection, cells
were replenished with fresh culture medium. Each well was transfected
with a mixture of 0.35mg pGL3 plasmid, 0.35 mg CMV–b-galactosidase
plasmid (used for normalization), and 1.25 mg transcription factor ex-
pression plasmid added to 14ml 2MCaCl2 and 111ml 23HBS solution
(Sigma-Aldrich) in a final volume of 222 ml. The day after transfection,
cell medium was replaced with fresh medium and the cells were incu-
bated for another day before being tested in the luciferase assay or sub-
jected to RNA extraction followed by quantitative real-time PCR.
Plasmid ConstructionThe pGL3-basic luciferase reporter vector (Promega, Madison, WI),
the CMV-b-galactosidase expression plasmid, and the expression
plasmids for Smad3, Smad4, and Smad5 were kindly provided by
Dr. A. Charonis (Biomedical Research Foundation of the Academy
of Athens). Expression plasmids for RelA/p65 (#21984), c-Jun
(#47443), Stat1a (#8690), and Stat6 (#35482) were obtained from
Addgene. PUC19 encoding for an inactive fragment of b-galactosidase
was used as control plasmid for transfection. The human and mouse
periostin promoter regions hProm 2 kb (22003+113), hProm 1 kb
(2888+113), mProm 2 kb (22037+20), and mProm 1 kb (2876+20),
taking +1 as the transcription start site, were amplified by PCR from
1488 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org

human or mouse DNA and cloned into the multiple cloning site se-
quence of pGL3-basic vector. The translation start site (ATG) for the
human and mouse periostin gene is at +119 and +35 after the tran-
scription start site, respectively.
Bioinformatics AnalysisBioinformatics analysis of human andmouseperiostin promoterswas
performed by the Genomatix software (www.genomatix.de). The de-
sired promoter sequence was retrieved by the Genomatix database
using the tool Gene2Promoter followed by multiple alignment to
check for conserved sequences and detection of putative transcrip-
tion factor binding sites per sequence similarity.
Luciferase AssaysFor luciferase activity measurement, the Luciferase Assay System from
Promega (E4030) was used according to manufacturer’s instructions.
Briefly, the cells were lysed in Reporter Lysis Buffer, the extract was
collected after centrifugation and removal of cell debris, and luciferase
activity was measured immediately in a luminometer after mixing 15ml
extract with 70 ml Luciferase Assay Reagent. To normalize luciferase
measurements, the activity of b-galactosidase was measured in a
b-galactosidase assay. For each sample, 30 ml cell extract were mixed
with 3mlmagnesiumsolution (0.1MMgCl2, 4.5Mb-mercaptoethanol),
66 ml ONPG (substrate), and 201 ml sodium phosphate (pH 7.5) and
incubated at 37°C for several minutes until the development of a faint
yellow color. The reactionwas stopped by addition of 1MNa2CO3 and
absorbance wasmeasured at 420 nm. The results were expressed as the
ratio of the relative luciferase units to b-galactosidase measurements.
ChIPChIP was performed in kidneys from wild-type SV129 mice injected
with NTS or PBS (control), using the SimpleChIP Plus Enzymatic
Chromatin IPKit (#9005; Cell Signaling Technology) permanufacturer’s
instructions. Briefly, whole kidneys were finely minced and crosslinked
with 1.5% formaldehyde for 20 minutes under constant rotation, fol-
lowed by addition of 0.1 M glycine to stop the reaction. The tissue was
disaggregated using a Dounce homogenizer, nuclei were prepared, and
chromatin was digested, sonicated, and analyzed by gel electrophoresis
and OD260 measurement to determine DNA concentration. Precipita-
tionwas performedwith the followingChIP-graded antibodies fromCell
Signaling Technology: NF-kB p65 (#8242), p-c-Jun (#3270), p-Stat1
(#7649), and p-Smad3 (#9520). Rabbit IgG was used as control. The
precipitated complexes were purified with the use of Protein GMagnetic
Beads, crosslinks were reversed, and DNA was eluted and purified.
Sequence-specific primers for selected regions of mouse periostin pro-
moter (Supplemental Table 3) were used in subsequent quantitative
real-time PCR analysis to check for enrichment of precipitated DNAs
in theNTS over the PBS group. The results were expressed as percentage
of the input (total DNA used for immunoprecipitation).
Coimmunoprecipitation AnalysisProtein interactions were stabilized by crosslinking minced half kidneys
with DTSSP (#21578; Thermo Fisher Scientific) per manufacturer’s in-
structions. Proteins were extracted in Pierce IP lysis buffer (#87787) sup-
plemented with PMSF, a protease inhibitor cocktail, and sodium
orthovanadate, and total protein concentration was measured using the
Bradford assay. Equal amounts of proteins were precipitated with a rabbit
anti–integrin-b3 antibody (ab119992; Abcam, Inc.) and rabbit IgG was
used as a control. The precipitated complexes were purifiedwith the use of
Protein GMagnetic Beads (#8740; Cell Signaling Technology) and loaded
on a denaturating SDS-PAGE followed by Western blotting. Equal
amounts of input were also loaded on the same gel. Immunoblotting
was performed for periostin (MOSF20; R&DSystems) and themembrane
was strippedusingRestoreWesternBlot StrippingBuffer (#21059;Thermo
Fisher Scientific) and reblotted for integrin-b3 (ab75872; Abcam, Inc.)
and Gapdh (Sigma-Aldrich). Veriblot was used for immunoprecipitation
analysis of secondary antibody (ab131366; Abcam, Inc.), preferentially
detecting the nonreduced form of IgG to avoid nonspecific signals from
coeluted reduced forms of the antibodies used for immunoprecipitation.
Statistical AnalysesData are expressed asmean6SEM. Datawere analyzed using one-way
ANOVA followed by a Fisher test. Values of P,0.05 were considered
significant.
ACKNOWLEDGMENTS
We thank Dr. Aristidis Charonis and Dr. Panagiotis Politis (from
Biomedical Research Foundation of the Academy of Athens) for
providing plasmid expression vectors. We also thank Dr. Nicole
Endlich for providing the podocyte cell line and Caroline Martin for
valuable help with animal breading. The CD146 antibody was a
generous gift from Professor Françoise Dignat-George.
This work was supported by grants from the Institut National de la
Santé et de la Recherche Médicale. N.P. was the recipient of a long-
term fellowship from the Society of European Renal Association-
European Dialysis and Transplantation.
DISCLOSURESNone.
REFERENCES
1. Horiuchi K, Amizuka N, Takeshita S, Takamatsu H, Katsuura M, OzawaH, Toyama Y, Bonewald LF, Kudo A: Identification and characterizationof a novel protein, periostin, with restricted expression to periosteumand periodontal ligament and increased expression by transforminggrowth factor beta. J Bone Miner Res 14: 1239–1249, 1999
2. Kruzynska-Frejtag A, Machnicki M, Rogers R, Markwald RR, Conway SJ:Periostin (an osteoblast-specific factor) is expressed within the embryonicmouse heart during valve formation.Mech Dev 103: 183–188, 2001
3. Norris RA, Damon B, Mironov V, Kasyanov V, Ramamurthi A, Moreno-Rodriguez R, Trusk T, Potts JD, Goodwin RL, Davis J, Hoffman S,Wen X,Sugi Y, Kern CB, Mjaatvedt CH, Turner DK, Oka T, Conway SJ,Molkentin JD, Forgacs G, Markwald RR: Periostin regulates collagenfibrillogenesis and the biomechanical properties of connective tissues.J Cell Biochem 101: 695–711, 2007
4. Kii I, Nishiyama T, Li M, Matsumoto K, Saito M, Amizuka N, Kudo A: In-corporationof tenascin-C into theextracellularmatrix byperiostin underliesan extracellularmeshwork architecture. J Biol Chem 285: 2028–2039, 2010
J Am Soc Nephrol 28: 1475–1490, 2017 Periostin Targeting Reverses CKD 1489
www.jasn.org BASIC RESEARCH

5. Snider P, Hinton RB,Moreno-Rodriguez RA,Wang J, Rogers R, LindsleyA, Li F, Ingram DA, Menick D, Field L, Firulli AB, Molkentin JD,Markwald R, Conway SJ: Periostin is required for maturation and ex-tracellular matrix stabilization of noncardiomyocyte lineages of theheart. Circ Res 102: 752–760, 2008
6. Gillan L, Matei D, Fishman DA, Gerbin CS, Karlan BY, Chang DD: Per-iostin secreted by epithelial ovarian carcinoma is a ligand for alpha(V)beta(3) and alpha(V)beta(5) integrins and promotes cell motility.CancerRes 62: 5358–5364, 2002
7. Li G, Jin R, Norris RA, Zhang L, Yu S, Wu F, Markwald RR, Nanda A,Conway SJ, Smyth SS, Granger DN: Periostinmediates vascular smoothmuscle cell migration through the integrins alphavbeta3 and al-phavbeta5 and focal adhesion kinase (FAK) pathway. Atherosclerosis208: 358–365, 2010
8. Li L, FanD,WangC,WangJY,Cui XB,WuD,ZhouY,WuLL: Angiotensin IIincreasesperiostin expressionviaRas/p38MAPK/CREBandERK1/2/TGF-b1 pathways in cardiac fibroblasts. Cardiovasc Res 91: 80–89, 2011
9. Takayama G, Arima K, Kanaji T, Toda S, Tanaka H, Shoji S, McKenzieAN, Nagai H, Hotokebuchi T, Izuhara K: Periostin: A novel componentof subepithelial fibrosis of bronchial asthma downstream of IL-4 andIL-13 signals. J Allergy Clin Immunol 118: 98–104, 2006
10. Wallace DP, White C, Savinkova L, Nivens E, Reif GA, Pinto CS, RamanA, Parnell SC, Conway SJ, Fields TA: Periostin promotes renal cystgrowth and interstitial fibrosis in polycystic kidney disease. Kidney Int85: 845–854, 2014
11. Wantanasiri P, Satirapoj B, Charoenpitakchai M, Aramwit P: Periostin: Anovel tissue biomarker correlates with chronicity index and renalfunction in lupus nephritis patients. Lupus 24: 835–845, 2015
12. Satirapoj B, Tassanasorn S, Charoenpitakchai M, Supasyndh O: Peri-ostin as a tissue and urinary biomarker of renal injury in type 2 diabetesmellitus. PLoS One 10: e0124055, 2015
13. Sen K, Lindenmeyer MT, Gaspert A, Eichinger F, Neusser MA, Kretzler M,SegererS,CohenCD:Periostin is induced inglomerular injury andexpressedde novo in interstitial renal fibrosis. Am J Pathol 179: 1756–1767, 2011
14. Guerrot D, Dussaule JC, Mael-Ainin M, Xu-Dubois YC, Rondeau E,Chatziantoniou C, Placier S: Identification of periostin as a criticalmarker of progression/reversal of hypertensive nephropathy. PLoSOne 7: e31974, 2012
15. Mael-Ainin M, Abed A, Conway SJ, Dussaule JC, Chatziantoniou C: In-hibition of periostin expression protects against the development of renalinflammation and fibrosis. J Am Soc Nephrol 25: 1724–1736, 2014
16. Kavvadas P, Dussaule JC, Chatziantoniou C: Searching novel diagnosticmarkers and targets for therapy of CKD. Kidney Int Suppl (2011) 4: 53–57, 2014
17. Prakoura N, Kavvadas P, Chatziantoniou C: New targets for end-stagechronic kidney disease therapy. J Crit Care Med 1: 92–95, 2015
18. Oshima A, Tanabe H, Yan T, Lowe GN, Glackin CA, Kudo A: A novelmechanism for the regulation of osteoblast differentiation: Transcrip-tion of periostin, a member of the fasciclin I family, is regulated by thebHLH transcription factor, twist. J Cell Biochem 86: 792–804, 2002
19. Lindsley A, Snider P, Zhou H, Rogers R, Wang J, Olaopa M, Kruzynska-Frejtag A, Koushik SV, Lilly B, Burch JB, Firulli AB, Conway SJ: Identificationand characterization of a novel Schwann and outflow tract endocardialcushion lineage-restrictedperiostin enhancer.DevBiol307: 340–355, 2007
20. Moustakas A, Heldin CH: Non-Smad TGF-beta signals. J Cell Sci 118:3573–3584, 2005
21. MasuokaM,ShiraishiH,OhtaS, Suzuki S,ArimaK,Aoki S, TodaS, InagakiN,Kurihara Y, Hayashida S, Takeuchi S, Koike K, Ono J, Noshiro H, Furue M,Conway SJ, Narisawa Y, Izuhara K: Periostin promotes chronic allergic in-flammation in response toTh2cytokines.JClin Invest122:2590–2600,2012
22. UchidaM, Shiraishi H,Ohta S, Arima K, Taniguchi K, Suzuki S, OkamotoM, Ahlfeld SK, Ohshima K, Kato S, Toda S, Sagara H, AizawaH, HoshinoT, Conway SJ, Hayashi S, Izuhara K: Periostin, a matricellular protein,plays a role in the induction of chemokines in pulmonary fibrosis. Am JRespir Cell Mol Biol 46: 677–686, 2012
23. Zhou W, Ke SQ, Huang Z, Flavahan W, Fang X, Paul J, Wu L, Sloan AE,McLendon RE, Li X, Rich JN, Bao S: Periostin secreted by glioblastomastem cells recruits M2 tumour-associated macrophages and promotesmalignant growth. Nat Cell Biol 17: 170–182, 2015
24. Blanchard C, Mingler MK, McBride M, Putnam PE, Collins MH, ChangG, Stringer K, Abonia JP, Molkentin JD, Rothenberg ME: Periostin fa-cilitates eosinophil tissue infiltration in allergic lung and esophagealresponses. Mucosal Immunol 1: 289–296, 2008
25. Arima M, Yoshida S, Nakama T, Ishikawa K, Nakao S, Yoshimura T, AsatoR, Sassa Y, Kita T, Enaida H, Oshima Y, Matsuda A, Kudo A, Ishibashi T:Involvement of periostin in regression of hyaloidvascular system duringocular development. Invest Ophthalmol Vis Sci 53: 6495–6503, 2012
26. Zhou HM, Wang J, Elliott C, Wen W, Hamilton DW, Conway SJ: Spa-tiotemporal expression of periostin during skin development and in-cisional wound healing: Lessons for human fibrotic scar formation. JCell Commun Signal 4: 99–107, 2010
27. Yang L, Serada S, Fujimoto M, Terao M, Kotobuki Y, Kitaba S, Matsui S,KudoA,Naka T,MurotaH, Katayama I: Periostin facilitates skin sclerosisvia PI3K/Akt dependent mechanism in a mouse model of scleroderma.PLoS One 7: e41994, 2012
28. NaikPK,BozykPD,BentleyJK,PopovaAP,BirchCM,WilkeCA,FryCD,WhiteES, Sisson TH, Tayob N, Carnemolla B, Orecchia P, Flaherty KR, HershensonMB, Murray S, Martinez FJ, Moore BB; COMET Investigators: Periostin pro-motes fibrosis and predicts progression in patients with idiopathic pulmonaryfibrosis. Am J Physiol Lung Cell Mol Physiol 303: L1046–L1056, 2012
29. Shimazaki M, Nakamura K, Kii I, Kashima T, Amizuka N, Li M, Saito M,Fukuda K, Nishiyama T, Kitajima S, Saga Y, Fukayama M, Sata M, KudoA: Periostin is essential for cardiac healing after acute myocardial in-farction. J Exp Med 205: 295–303, 2008
30. Oka T, Xu J, Kaiser RA, Melendez J, Hambleton M, Sargent MA, Lorts A,Brunskill EW,DornGW2nd ,Conway SJ, AronowBJ, Robbins J,MolkentinJD: Genetic manipulation of periostin expression reveals a role in cardiachypertrophy and ventricular remodeling. Circ Res 101: 313–321, 2007
31. Yamashita O, Yoshimura K, Nagasawa A, Ueda K, Morikage N, Ikeda Y,Hamano K: Periostin links mechanical strain to inflammation in ab-dominal aortic aneurysm. PLoS One 8: e79753, 2013
32. LiuAY,ZhengH,OuyangG:Periostin,amultifunctionalmatricellularprotein ininflammatory and tumor microenvironments.Matrix Biol 37: 150–156, 2014
33. Wei C, El Hindi S, Li J, Fornoni A, Goes N, Sageshima J, Maiguel D,Karumanchi SA, Yap HK, Saleem M, Zhang Q, Nikolic B, Chaudhuri A,Daftarian P, Salido E, Torres A, SalifuM, Sarwal MM, Schaefer F,MorathC, Schwenger V, ZeierM,Gupta V, RothD, RastaldiMP, BurkeG, Ruiz P,Reiser J: Circulating urokinase receptor as a cause of focal segmentalglomerulosclerosis. Nat Med 17: 952–960, 2011
34. Wei C, Möller CC, Altintas MM, Li J, Schwarz K, Zacchigna S, Xie L, HengerA, Schmid H, Rastaldi MP, Cowan P, Kretzler M, Parrilla R, Bendayan M,GuptaV,NikolicB,Kalluri R,Carmeliet P,Mundel P, Reiser J:Modificationofkidney barrier function by the urokinase receptor.NatMed 14: 55–63, 2008
35. Guo W, Giancotti FG: Integrin signalling during tumour progression.Nat Rev Mol Cell Biol 5: 816–826, 2004
36. Zhu M, Saxton RE, Ramos L, Chang DD, Karlan BY, Gasson JC, SlamonDJ: Neutralizing monoclonal antibody to periostin inhibits ovarian tu-mor growth and metastasis. Mol Cancer Ther 10: 1500–1508, 2011
37. Mesnard L, Keller AC, Michel ML, Vandermeersch S, Rafat C,Letavernier E, Tillet Y, Rondeau E, Leite-de-Moraes MC: Invariant nat-ural killer T cells and TGF-beta attenuate anti-GBMglomerulonephritis.J Am Soc Nephrol 20: 1282–1292, 2009
38. El Machhour F, Keuylian Z, Kavvadas P, Dussaule JC, Chatziantoniou C:Activation of Notch3 in glomeruli promotes the development of rapidlyprogressive renal disease. J Am Soc Nephrol 26: 1561–1575, 2015
This article contains supplemental material online at http://jasn.asnjournals.org/lookup/suppl/doi:10.1681/ASN.2016070709/-/DCSupplemental.
1490 Journal of the American Society of Nephrology J Am Soc Nephrol 28: 1475–1490, 2017
BASIC RESEARCH www.jasn.org





























