Embed Size (px)
Citation preview

Human Cancer Biology
Neutrophil Degranulation and Immunosuppression inPatients with GBM: Restoration of Cellular Immune Functionby Targeting Arginase I
Trisha R. Sippel1,2, Jason White1, Kamalika Nag, Vadim Tsvankin1, Marci Klaassen1,B.K. Kleinschmidt-DeMasters1,3, and Allen Waziri1,2
AbstractPurpose: The source of glioblastoma (GBM)-associated immunosuppression remainsmultifactorial.We
sought to clarify and therapeutically targetmyeloid cell–derived peripheral immunosuppression in patients
with GBM.
Experimental Design: Direct ex vivo T-cell function, serum Arginase I (ArgI) levels, and circulating
myeloid lineage populations were compared between patients with GBM and normal donors or patients
with other intracranial tumors. Immunofunctional assays were conducted using bulk and sorted cell
populations to explore the potential transfer ofmyeloid cell–mediated immunosuppression and to identify
a potential mechanism for these effects. ArgI-mediated immunosuppression was therapeutically targeted in
vitro through pharmacologic inhibition or arginine supplementation.
Results: We identified a significantly expanded population of circulating, degranulated neutrophils
associated with elevated levels of serum ArgI and decreased T-cell CD3z expression within peripheral blood
from patients with GBM. Sorted CD11bþ cells from patients with GBM were found to markedly suppress
normal donor T-cell function in coculture, and media harvested frommitogen-stimulated GBM peripheral
bloodmononuclear cell (PBMC)orGBM-associatedmixed lymphoid reactions showedArgI levels thatwere
significantly higher than controls. Critically, T-cell suppression inboth settings couldbe completely reversed
through pharmacologic ArgI inhibition or with arginine supplementation.
Conclusions: These data indicate that peripheral cellular immunosuppression in patients with GBM is
associatedwith neutrophil degranulation and elevated levels of circulating ArgI, and that T-cell function can
be restored in these individuals by targeting ArgI. These data identify a novel pathway of GBM-mediated
suppression of cellular immunity and offer a potential therapeutic window for improving antitumor
immunity in affected patients. Clin Cancer Res; 17(22); 6992–7002. �2011 AACR.
Introduction
Treatment of patients diagnosed with glioblastoma(GBM) is one of the lasting challenges ofmodernmedicine.Responses to standard external beam radiotherapy andchemotherapy remain dismal, providing only limitedimprovement in survival. Most experimental therapies haveproven ineffective. The overall clinical strategy for affectedindividuals has not changed significantly in basic design for
several decades. In spite of all best attempts, tumor recur-rence is a nearly uniform phenomenon and the significantmajority of patients succumb to progressive brain disease injust over 1 year from diagnosis (1). It is clear that newapproaches for developing effective and targeted treatmentoptions are needed for patients with GBM.
To this end, immunotherapy has become a focus of recentresearch in GBM due to the potential for combined targetspecificity and sensitivity. Numerous groups have testedimmunotherapeutic strategies in patientswithGBM.Unfor-tunately, as has been seen with similar approaches in mostother cancers, these efforts have been in large part unsuc-cessful (2). A major potential pitfall for immunotherapy inGBM is the known suppression of cellular immunity seen inaffected patients, which has been well described over thepast few decades. Many groups have reported on the varietyof functional defects seen in the circulating pool of T cellsfrom these individuals (3). We, and others, have documen-ted the exceedingly rare and ultimately ineffectual T-cellinfiltrates found within GBM (4). In spite of these appar-ently local as well as global aberrations in cellular
Authors' Affiliations: Departments of 1Neurosurgery, 2Cancer Biology,and 3Pathology,University ofColoradoAnschutzMedical Campus, Aurora,Colorado
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
Corresponding Author: Allen Waziri, Department of Neurosurgery, Univer-sity of Colorado Anschutz Medical Campus, Academic Office Building 1,Room 5001, 12631 E. 17th Ave., Aurora, CO 80045. Phone: 303-724-4134;Fax: 303-724-6012; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-11-1107
�2011 American Association for Cancer Research.
ClinicalCancer
Research
Clin Cancer Res; 17(22) November 15, 20116992
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107

immunity, patients withGBMare generally not systemicallyimmunocompromised prior to the growth of their tumor.This fact, combined with the potential for recovery ofcellular immune function following surgical resection(5), has implicated a tumor-derived factor in the suppres-sion of cell-mediated immune responses. It is thereforelikely that tumor-associated immunosuppressive factorswill similarly affect clinical attempts to augment antitumorresponses. Therefore, targeting tumor-associated immuno-suppression in patients with GBM will be critical for thedevelopment of meaningful immunotherapeutic strategies.Cells of myeloid lineage have been increasingly associ-
ated with immunosuppression in a number of systems,including various forms of cancer. Myeloid-derived cells atdifferent states of maturation have been studied as potentinactivators of both CD4þ and CD8þ T cells (6). Popula-tions of immature myeloid cells as well as more mature,differentiated monocytes and granulocytes have been pre-viously shown to possess immunosuppressive abilities(7–9). Given prior observations of T-cell dysfunction inpatients with GBM and the documentation of myeloid cellswith immunosuppressive characteristics in patients withother cancers, the current study is attempted to identify amyeloid-derived source of peripheral immunosuppressionin patients with GBM.
Materials and Methods
Patient and sample collectionPeripheral blood was collected from patients undergoing
neurosurgical resection of intracranial tumors (GBM, ana-plastic glioma, meningioma, and pituitary tumor) at the
University of Colorado Hospital with appropriate Institu-tional Review Board approval. Patient age and gender didnot vary significantly between groups. Preoperative steroidtreatment was taken into consideration; however, no sta-tistically significant differences were found between preop-erative steroid use and the presence of activated neutrophils(reviewed in Supplementary Table S1). Normal donor(ND) blood was collected from anonymous donors fromthe blood bank at the University of Colorado.
Within 1 hour from harvest, plasma was removed fromperipheral blood samples and stored at �70�C. Peripheralblood mononuclear cells (PBMC) were purified by centri-fugation over a Ficoll Histopaque density gradient (Sigma)according to themanufacturer’s protocol. PBMCswere usedimmediately, without freezing, for T-cell functional assaysor staining by flow cytometry. ND granulocytes used forstaining by flow cytometry were collected within the flow-through fraction of the Ficoll prep; red blood cells werelysed via brief incubation in 0.84% ammonium chloride.
T-cell functional assaysMitogenic stimulations were conducted using bulk
PBMCs or isolated T cells from ND or patients cultured inRPMI-1640 media with 10% FBS and 1% penicillin–strep-tomycin. T cells were isolated using CD3þ selection mag-netic beads per manufacturer’s protocol (Miltenyi Biotec).Cells were plated at 1 � 105 cells per well in a 48-well platewith 500 mL of media. The cells were stimulated with either1 � 105 Dynabeads (Invitrogen) or 5 mg/mL phytohemag-glutinin (PHA; Sigma) per well and incubated for 48 and 72hours poststimulation. Media IFN-g levels were assayed byELISA (Thermo Scientific) according to the manufacturer’sprotocol. In functional assays where proliferation was mea-sured by carboxyfluorescein succinimidyl ester (CFSE)staining, isolated PBMCs at a concentration of 6 � 105
cells/mL were mixed with 5 mmol/L CFSE (BD Pharmin-gen) for 5 minutes and washed with media. Cells were thenstimulated with 5 mg/mL PHA and incubated for 72 hoursprior to flow cytometry. For flow cytometric analysis, sam-pleswere stainedwith anti-CD3-APC andCD3þ T cells weregated upon for subsequent evaluation of CFSE fluorescence.For comparative measurement of T-cell proliferation asmeasured by CFSE dilution, the proliferation index (PI)was calculated on the basis of proportion of proliferatingcells over total T cells.
Mixed lymphoid reactions (MLR) were carried out usingbulk PBMCs collected from patients and ND. "Modified"MLR used purified CD11bþ myeloid cells and CD3þ T cellsfrom patients and ND, again isolated using positive beadselection. Cells from 2 different ND or a ND and a tumorpatient were mixed at 1.0 � 105 cells per well of each celltype in 200 mLmedia in 96-well plates. MLRwere incubatedfor 48 and 72 hours. Media IFN-g levels were assayed byELISA as described above.
Flow cytometryDirectly after isolation, 1� 106 cells were resuspended in
200 mL FACS buffer (PBSþ 20% FBS). Cells were incubated
Translational Relevance
Glioblastoma (GBM) remains one of the most lethaltumors known to modern medicine, and new therapeu-tic options are desperately needed for affected patients.Immunotherapeutic strategies have been for the mostpart unsuccessful in GBM, likely due to tumor-mediatedsuppression of cellular immune responses. The currentstudy explored the role and functional mechanisms ofperipheral myeloid cells in the suppression of cellularimmunity in patients with GBM. We provide the firstdocumentation of transferable immunosuppressionassociated with neutrophilic degranulation andincreased circulating levels of Arginase I (ArgI) withinperipheral blood from patients with GBM. In addition,we show that cellular immune function in patients withGBM can be restored through pharmacologic inhibitionof ArgI or by supplementation with exogenous arginine.These data identify a novel pathway of GBM-mediatedsuppression of cellular immunity and offer a potentialtherapeutic window for improving antitumor immunityin affected patients.
Neutrophils and Immunosuppression in GBM
www.aacrjournals.org Clin Cancer Res; 17(22) November 15, 2011 6993
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107

with antibodies against CD11b, CD33, CD14, HLA-DR,CD15, or CD66 (BD Biosciences) for 45 minutes at 4�Cprior to measuring expression on a FACSCalibur flow cyt-ometer. Intracellular staining for CD3z was carried out byresuspending CD3-stained PBMCs in 100 mL of Cytofix/Cytoperm solution (BD Biosciences) for 20 minutes at 4�C,washed with Perm/Wash Buffer (BD Biosciences), andstainedwith anti-CD3z (CD247; Santa Cruz Biotechnology)for 45 minutes at 4�C prior to measurement. Flow data wereanalyzed using the FlowJo Software program (Treestar).
Flow-sorting, cytospin, and GBM histopathologicanalysis
CD11bþCD33lo and CD11bþCD33hi populations with-in PBMCs from patients with GBM were sorted using aFACSAria flow sorter. In parallel experiments, putative gran-ulocytes within PBMCs were isolated using magnetic beadseparation by positive CD66 selection (Miltenyi Biotec). Theflow-through fraction from the CD66þ selection was collect-ed and incubated with CD11bþ selection beads to collectCD11bþCD66� monocytes. Sorted populations were spunontoSuperfrostmicroscope slides (Fischer)bycentrifugingat750 rpm for 2 minutes. Slides were stained with Wright–Giemsa stain and visualized at high power. For evaluation ofactively necrotic GBM samples, formalin-fixed specimenswere prepared as per standard procedures and stained withhematoxylin and eosin. Representative sections were alsosubject to immunohistochemical analysis for CD15 (Ven-tana Medical Systems) and myeloperoxidase (Dako) usingstandard techniques. Pathologic slides were reviewed by theneuropathologist on the study (B.K. DeMasters).
Arginase I measurementPlasma samples and media from T-cell functional assays
described above were subject to Arginase I (ArgI) ELISA(Hycult Biotechnology) according to the manufacturer’sprotocol. Samples were diluted 1:1 with kit dilution buffer.For evaluation of ArgI levels within necrotic material fromGBM, necrotic tissue was weighed and diluted in unsup-plemented RPMI media to a ratio of 60 mL/mg. The tissuewas then disbanded and vortexed to suspend extracellularcontents into the media. The resulting samples were cen-trifuged at 5,000 rpm for 10minutes to remove excess tissueand supernatants were collected. Supernatant fromnecroticGBM samples were diluted over a range of 1:1 to 1:100 andused for ArgI ELISA as above.
Induction of degranulation using fMLPFormyl-methionyl-leucyl-phenylalanine (fMLP; Sigma)
was added to whole blood at a concentration of 1 mmol/Land incubatedat roomtemperature for1hour.Wholebloodswithout fMLP were used as controls. Following incubation,PBMCs were collected using a Ficoll density gradient asdescribed above. Bulk PBMCs were used to assess T-cellfunction through PHA stimulation and stained for flowcytometry as described previously.
For detection of dose-dependent neutrophilic suppres-sion of T-cell proliferation, neutrophils were sorted from
ND whole blood using density centrifugation purificationover a 42%/51%Percoll gradient, followed by CD66þ beadseparation. Purified neutrophils were activated with 1mmol/L fMLP. T cells were purified using CD3þ separation.Cultures were then prepared with varying T-cell per neu-trophil ratios as outlined in Supplementary Fig. S2 andstimulatedwith 5mg/mLPHA for 72hours. BrdUwas addedto cultures for the final 20 hours and cells were thenharvested for flow cytometric quantification of prolifera-tion. Media ArgI levels from these cultures were analyzed byELISA as described above.
Assays to overcome arginase activity in vitroT-cell functional assays (PHA stimulation and MLR)
using bulk PBMCs were conducted as above. Groups ofsamples were treated with 7.81 mg/mL L-arginine (Sigma)or 40 mmol/L nor-NOHA (Cayman Chemicals) at the timeof plating and PHA addition or cell mixing. Dose–responseprofiles for each compound were developed prior to testingon patient samples to identify the highest possible dose thatdid not affect baseline T-cell functional response (i.e.,toxicity or augmented functional response) in ND samples(data not shown). Cells were incubated for 48 and 72 hoursand media IFN-g levels were tested by ELISA as describedabove.
Statistical analysisData are represented asmean� SEM.Multigroup analysis
was conducted using ANOVA. Differences between 2 vari-ables were determined using the Student t test. P values lessthan 0.05 were considered significant.
Results
Direct ex vivo T cells from patients with GBM arefunctionally suppressed in vitro
To confirm prior reports describing decreased prolifer-ative responses of T cells from patients with GBM andPBMCs were purified, stained with CFSE, and stimulatedwith PHA directly ex vivo. Flow cytometric analysis ofstimulated T cells from patients with GBM indeed showedsignificantly lower levels of proliferation than seen fromNDor patients with other intracranial tumors (Fig. 1A). Toevaluate stimulation-induced cytokine production, levelsof IFN-g withinmedia fromPHA-stimulated PBMC cultureswere assayed by ELISA. Cultures from patients with GBMgenerated significantly less IFN-g at both 48 and 72 hoursthan did matched samples from ND or patients with otherintracranial tumors (Fig. 1B). Taken together, these resultscorroborate prior experimental data documenting thehyporesponsive nature of T cells in patients with GBM.
PBMCs and purified CD11bþ cells from patients withGBM suppress normal donor T-cell function
To confirm the presumptive cellular source of peripheralT-cell suppression in patients with GBM, we explored thepossibility that GBM-associated immunosuppression couldbe transferred to ND T cells. MLR were prepared using
Sippel et al.
Clin Cancer Res; 17(22) November 15, 2011 Clinical Cancer Research6994
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107
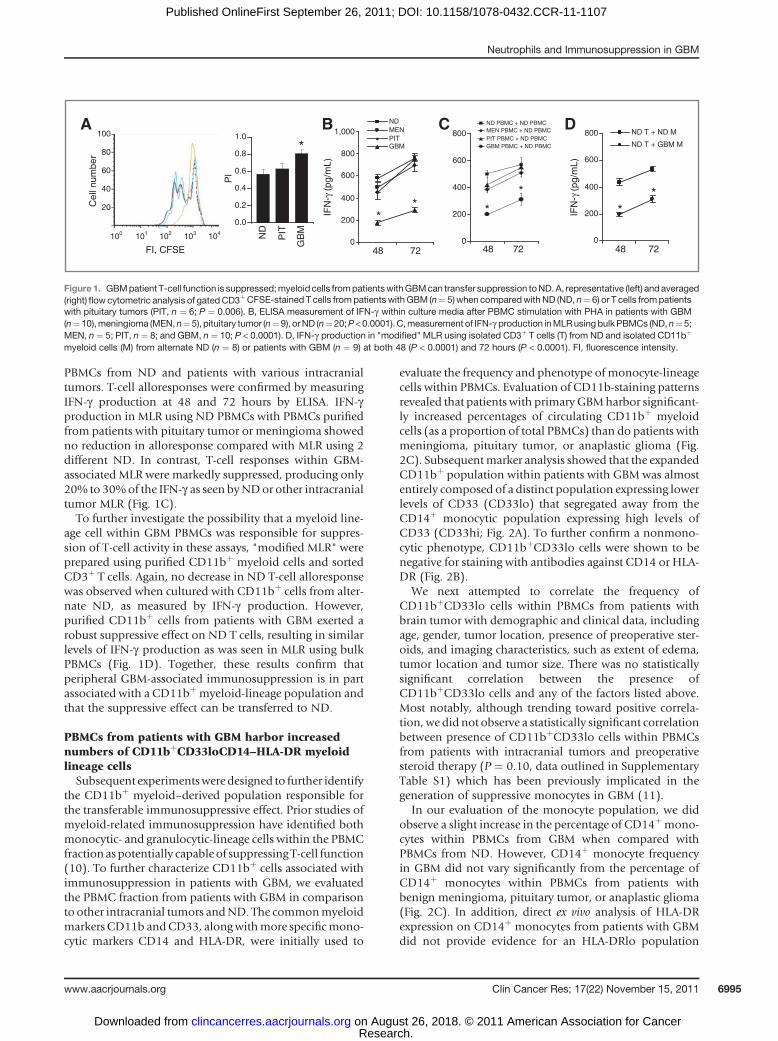
PBMCs from ND and patients with various intracranialtumors. T-cell alloresponses were confirmed by measuringIFN-g production at 48 and 72 hours by ELISA. IFN-gproduction in MLR using ND PBMCs with PBMCs purifiedfrom patients with pituitary tumor or meningioma showedno reduction in alloresponse compared with MLR using 2different ND. In contrast, T-cell responses within GBM-associated MLR were markedly suppressed, producing only20% to 30%of the IFN-g as seen byNDor other intracranialtumor MLR (Fig. 1C).To further investigate the possibility that a myeloid line-
age cell within GBM PBMCs was responsible for suppres-sion of T-cell activity in these assays, "modified MLR" wereprepared using purified CD11bþ myeloid cells and sortedCD3þ T cells. Again, no decrease in ND T-cell alloresponsewas observed when cultured with CD11bþ cells from alter-nate ND, as measured by IFN-g production. However,purified CD11bþ cells from patients with GBM exerted arobust suppressive effect on ND T cells, resulting in similarlevels of IFN-g production as was seen in MLR using bulkPBMCs (Fig. 1D). Together, these results confirm thatperipheral GBM-associated immunosuppression is in partassociated with a CD11bþmyeloid-lineage population andthat the suppressive effect can be transferred to ND.
PBMCs from patients with GBM harbor increasednumbers of CD11bþCD33loCD14–HLA-DR myeloidlineage cellsSubsequent experimentswere designed to further identify
the CD11bþ myeloid–derived population responsible forthe transferable immunosuppressive effect. Prior studies ofmyeloid-related immunosuppression have identified bothmonocytic- and granulocytic-lineage cells within the PBMCfractionaspotentially capableof suppressing T-cell function(10). To further characterize CD11bþ cells associated withimmunosuppression in patients with GBM, we evaluatedthe PBMC fraction from patients with GBM in comparisonto other intracranial tumors andND. The commonmyeloidmarkers CD11b andCD33, alongwithmore specificmono-cytic markers CD14 and HLA-DR, were initially used to
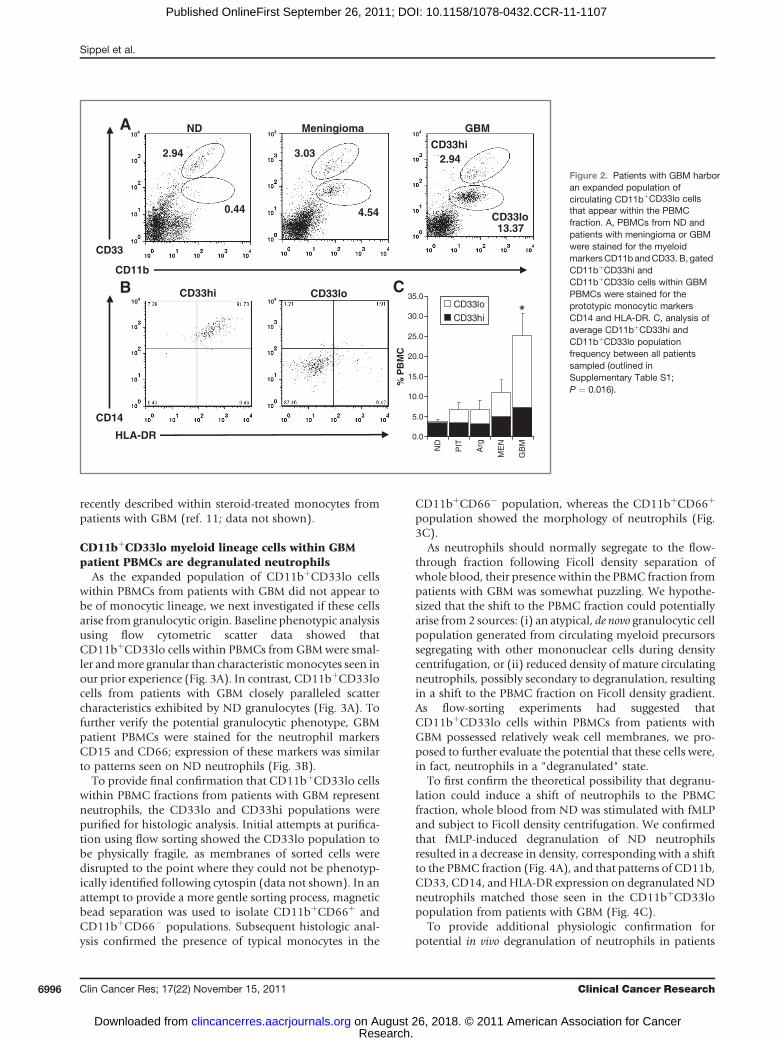
evaluate the frequency and phenotype of monocyte-lineagecells within PBMCs. Evaluation of CD11b-staining patternsrevealed that patients with primaryGBMharbor significant-ly increased percentages of circulating CD11bþ myeloidcells (as a proportion of total PBMCs) than do patients withmeningioma, pituitary tumor, or anaplastic glioma (Fig.2C). Subsequentmarker analysis showed that the expandedCD11bþ population within patients with GBM was almostentirely composed of a distinct population expressing lowerlevels of CD33 (CD33lo) that segregated away from theCD14þ monocytic population expressing high levels ofCD33 (CD33hi; Fig. 2A). To further confirm a nonmono-cytic phenotype, CD11bþCD33lo cells were shown to benegative for staining with antibodies against CD14 or HLA-DR (Fig. 2B).
We next attempted to correlate the frequency ofCD11bþCD33lo cells within PBMCs from patients withbrain tumor with demographic and clinical data, includingage, gender, tumor location, presence of preoperative ster-oids, and imaging characteristics, such as extent of edema,tumor location and tumor size. There was no statisticallysignificant correlation between the presence ofCD11bþCD33lo cells and any of the factors listed above.Most notably, although trending toward positive correla-tion, we did not observe a statistically significant correlationbetween presence of CD11bþCD33lo cells within PBMCsfrom patients with intracranial tumors and preoperativesteroid therapy (P ¼ 0.10, data outlined in SupplementaryTable S1) which has been previously implicated in thegeneration of suppressive monocytes in GBM (11).
In our evaluation of the monocyte population, we didobserve a slight increase in the percentage of CD14þmono-cytes within PBMCs from GBM when compared withPBMCs from ND. However, CD14þ monocyte frequencyin GBM did not vary significantly from the percentage ofCD14þ monocytes within PBMCs from patients withbenign meningioma, pituitary tumor, or anaplastic glioma(Fig. 2C). In addition, direct ex vivo analysis of HLA-DRexpression on CD14þ monocytes from patients with GBMdid not provide evidence for an HLA-DRlo population
0
200
400
600
800
7248
ND PBMC + ND PBMCMEN PBMC + ND PBMCPIT PBMC + ND PBMCGBM PBMC + ND PBMC
0
200
400
600
800
7248
ND T + ND M
ND T + GBM M
0
200
400
600
800
1,000
7248
NDMENPITGBM
IFN
-γ (p
g/m
L)
A
0.0
0.2
0.4
0.6
0.8
1.0
PI
PIT
GB
M
ND
B
FI, CFSE
Cel
l num
ber
C*
***
*
D
** IF
N-γ
(pg/
mL)
100
80
60
40
20
100 101 102 103 104
Figure 1. GBMpatient T-cell function is suppressed;myeloid cells frompatientswithGBMcan transfer suppression toND. A, representative (left) andaveraged(right) flow cytometric analysis of gatedCD3þCFSE-stained T cells frompatientswith GBM (n¼ 5) when comparedwith ND (ND, n¼ 6) or T cells frompatientswith pituitary tumors (PIT, n ¼ 6; P ¼ 0.006). B, ELISA measurement of IFN-g within culture media after PBMC stimulation with PHA in patients with GBM(n¼10),meningioma (MEN,n¼5), pituitary tumor (n¼9), orND (n¼20;P<0.0001).C,measurement of IFN-g production inMLRusingbulkPBMCs (ND,n¼5;MEN, n¼ 5; PIT, n ¼ 8; and GBM, n ¼ 10; P < 0.0001). D, IFN-g production in "modified" MLR using isolated CD3þ T cells (T) from ND and isolated CD11bþ
myeloid cells (M) from alternate ND (n ¼ 8) or patients with GBM (n ¼ 9) at both 48 (P < 0.0001) and 72 hours (P < 0.0001). FI, fluorescence intensity.
Neutrophils and Immunosuppression in GBM
www.aacrjournals.org Clin Cancer Res; 17(22) November 15, 2011 6995
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107
recently described within steroid-treated monocytes frompatients with GBM (ref. 11; data not shown).
CD11bþCD33lo myeloid lineage cells within GBMpatient PBMCs are degranulated neutrophils
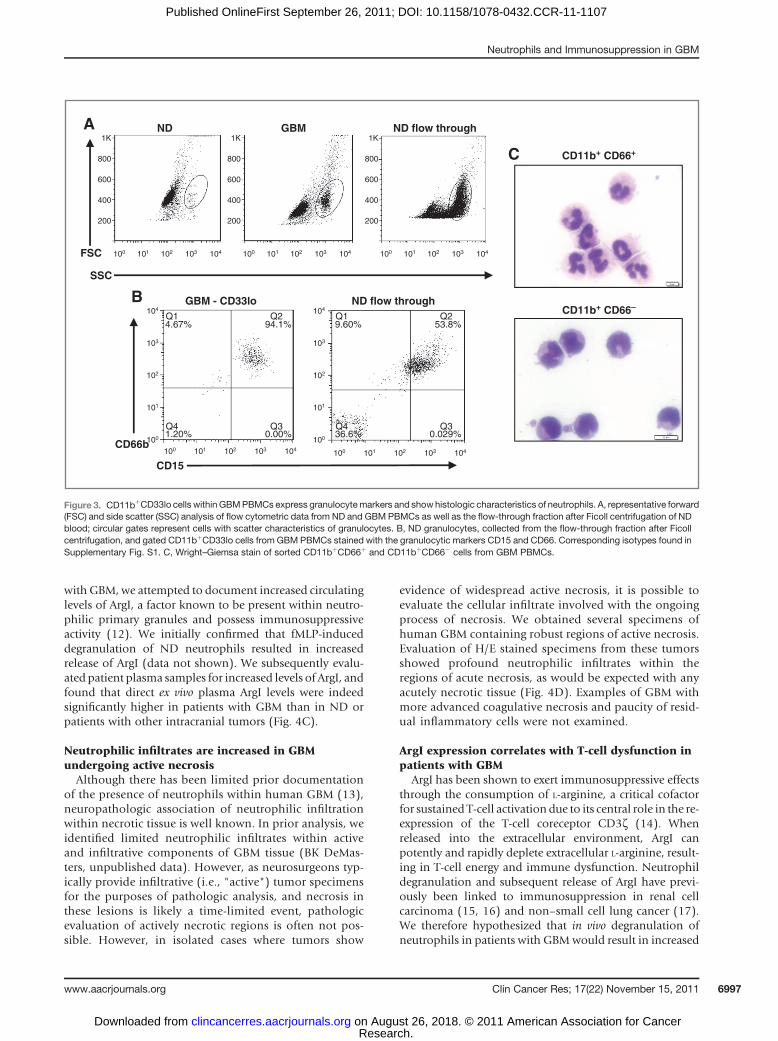
As the expanded population of CD11bþCD33lo cellswithin PBMCs from patients with GBM did not appear tobe of monocytic lineage, we next investigated if these cellsarise from granulocytic origin. Baseline phenotypic analysisusing flow cytometric scatter data showed thatCD11bþCD33lo cells within PBMCs fromGBMwere smal-ler andmore granular than characteristic monocytes seen inour prior experience (Fig. 3A). In contrast, CD11bþCD33locells from patients with GBM closely paralleled scattercharacteristics exhibited by ND granulocytes (Fig. 3A). Tofurther verify the potential granulocytic phenotype, GBMpatient PBMCs were stained for the neutrophil markersCD15 and CD66; expression of these markers was similarto patterns seen on ND neutrophils (Fig. 3B).
To provide final confirmation that CD11bþCD33lo cellswithin PBMC fractions from patients with GBM representneutrophils, the CD33lo and CD33hi populations werepurified for histologic analysis. Initial attempts at purifica-tion using flow sorting showed the CD33lo population tobe physically fragile, as membranes of sorted cells weredisrupted to the point where they could not be phenotyp-ically identified following cytospin (data not shown). In anattempt to provide a more gentle sorting process, magneticbead separation was used to isolate CD11bþCD66þ andCD11bþCD66� populations. Subsequent histologic anal-ysis confirmed the presence of typical monocytes in the
CD11bþCD66� population, whereas the CD11bþCD66þ
population showed the morphology of neutrophils (Fig.3C).
As neutrophils should normally segregate to the flow-through fraction following Ficoll density separation ofwhole blood, their presence within the PBMC fraction frompatients with GBM was somewhat puzzling. We hypothe-sized that the shift to the PBMC fraction could potentiallyarise from2 sources: (i) an atypical, de novo granulocytic cellpopulation generated from circulating myeloid precursorssegregating with other mononuclear cells during densitycentrifugation, or (ii) reduced density of mature circulatingneutrophils, possibly secondary to degranulation, resultingin a shift to the PBMC fraction on Ficoll density gradient.As flow-sorting experiments had suggested thatCD11bþCD33lo cells within PBMCs from patients withGBM possessed relatively weak cell membranes, we pro-posed to further evaluate the potential that these cells were,in fact, neutrophils in a "degranulated" state.
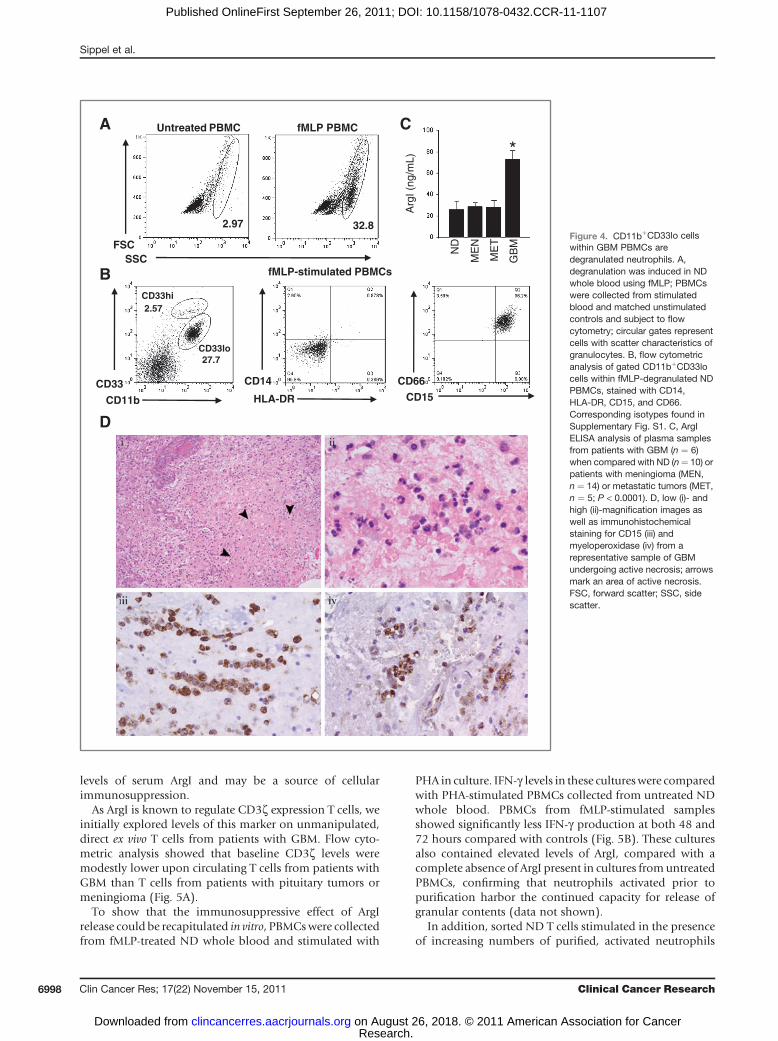
To first confirm the theoretical possibility that degranu-lation could induce a shift of neutrophils to the PBMCfraction, whole blood from ND was stimulated with fMLPand subject to Ficoll density centrifugation. We confirmedthat fMLP-induced degranulation of ND neutrophilsresulted in a decrease in density, corresponding with a shiftto the PBMC fraction (Fig. 4A), and that patterns of CD11b,CD33, CD14, andHLA-DR expression on degranulated NDneutrophils matched those seen in the CD11bþCD33lopopulation from patients with GBM (Fig. 4C).
To provide additional physiologic confirmation forpotential in vivo degranulation of neutrophils in patients
104 104 104
104 104
0.0
5.0
10.0
15.0
20.0
25.0
30.0
35.0
ND
PIT
Arg
ME
N
GB
M
% P
BM
C
CD33lo
CD33hi
CD33
CD11b
2.94
0.44 4.54 CD33lo13.37
CD33hi2.943.03
A GBMMeningiomaND
CD14
HLA-DR
B CD33hi CD33lo C
*
Figure 2. Patients with GBM harboran expanded population ofcirculating CD11bþCD33lo cellsthat appear within the PBMCfraction. A, PBMCs from ND andpatients with meningioma or GBMwere stained for the myeloidmarkersCD11b andCD33. B, gatedCD11bþCD33hi andCD11bþCD33lo cells within GBMPBMCs were stained for theprototypic monocytic markersCD14 and HLA-DR. C, analysis ofaverage CD11bþCD33hi andCD11bþCD33lo populationfrequency between all patientssampled (outlined inSupplementary Table S1;P ¼ 0.016).
Sippel et al.
Clin Cancer Res; 17(22) November 15, 2011 Clinical Cancer Research6996
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107
with GBM, we attempted to document increased circulatinglevels of ArgI, a factor known to be present within neutro-philic primary granules and possess immunosuppressiveactivity (12). We initially confirmed that fMLP-induceddegranulation of ND neutrophils resulted in increasedrelease of ArgI (data not shown). We subsequently evalu-ated patient plasma samples for increased levels of ArgI, andfound that direct ex vivo plasma ArgI levels were indeedsignificantly higher in patients with GBM than in ND orpatients with other intracranial tumors (Fig. 4C).
Neutrophilic infiltrates are increased in GBMundergoing active necrosisAlthough there has been limited prior documentation
of the presence of neutrophils within human GBM (13),neuropathologic association of neutrophilic infiltrationwithin necrotic tissue is well known. In prior analysis, weidentified limited neutrophilic infiltrates within activeand infiltrative components of GBM tissue (BK DeMas-ters, unpublished data). However, as neurosurgeons typ-ically provide infiltrative (i.e., "active") tumor specimensfor the purposes of pathologic analysis, and necrosis inthese lesions is likely a time-limited event, pathologicevaluation of actively necrotic regions is often not pos-sible. However, in isolated cases where tumors show
evidence of widespread active necrosis, it is possible toevaluate the cellular infiltrate involved with the ongoingprocess of necrosis. We obtained several specimens ofhuman GBM containing robust regions of active necrosis.Evaluation of H/E stained specimens from these tumorsshowed profound neutrophilic infiltrates within theregions of acute necrosis, as would be expected with anyacutely necrotic tissue (Fig. 4D). Examples of GBM withmore advanced coagulative necrosis and paucity of resid-ual inflammatory cells were not examined.
ArgI expression correlates with T-cell dysfunction inpatients with GBM
ArgI has been shown to exert immunosuppressive effectsthrough the consumption of L-arginine, a critical cofactorfor sustained T-cell activation due to its central role in the re-expression of the T-cell coreceptor CD3z (14). Whenreleased into the extracellular environment, ArgI canpotently and rapidly deplete extracellular L-arginine, result-ing in T-cell energy and immune dysfunction. Neutrophildegranulation and subsequent release of ArgI have previ-ously been linked to immunosuppression in renal cellcarcinoma (15, 16) and non–small cell lung cancer (17).We therefore hypothesized that in vivo degranulation ofneutrophils in patients with GBMwould result in increased
ND1K
800
600
400
200
1K
800
600
400
200
GBM
FSC
SSC
A
CD15
CD66b
B GBM - CD33lo
CD11b+ CD66+
CD11b+ CD66–
C
ND flow through
ND flow through
1K
800
600
400
200
100 101 102 103 104 100 101 102 103 104 100 101 102 103 104
100 101 102 103 104100 101 102 103 104
104
103
102
101
100
104
103
102
101
100
Q1 Q2
Q4 Q3
4.67% 94.1%
1.20% 0.00%
Q1 Q2
Q4 Q3
9.60% 53.8%
36.6% 0.029%
Figure 3. CD11bþCD33lo cells within GBMPBMCs express granulocytemarkers and showhistologic characteristics of neutrophils. A, representative forward(FSC) and side scatter (SSC) analysis of flow cytometric data from ND and GBM PBMCs as well as the flow-through fraction after Ficoll centrifugation of NDblood; circular gates represent cells with scatter characteristics of granulocytes. B, ND granulocytes, collected from the flow-through fraction after Ficollcentrifugation, and gated CD11bþCD33lo cells from GBM PBMCs stained with the granulocytic markers CD15 and CD66. Corresponding isotypes found inSupplementary Fig. S1. C, Wright–Giemsa stain of sorted CD11bþCD66þ and CD11bþCD66� cells from GBM PBMCs.
Neutrophils and Immunosuppression in GBM
www.aacrjournals.org Clin Cancer Res; 17(22) November 15, 2011 6997
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107
levels of serum ArgI and may be a source of cellularimmunosuppression.
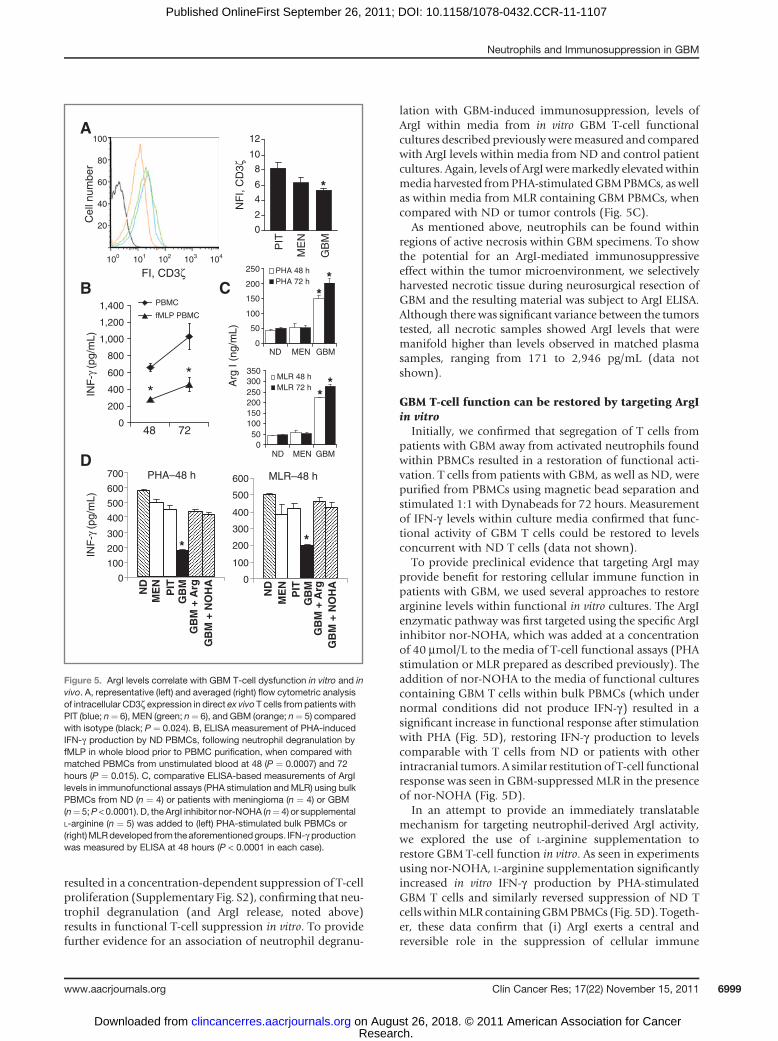
As ArgI is known to regulate CD3z expression T cells, weinitially explored levels of this marker on unmanipulated,direct ex vivo T cells from patients with GBM. Flow cyto-metric analysis showed that baseline CD3z levels weremodestly lower upon circulating T cells from patients withGBM than T cells from patients with pituitary tumors ormeningioma (Fig. 5A).
To show that the immunosuppressive effect of ArgIrelease could be recapitulated in vitro, PBMCswere collectedfrom fMLP-treated ND whole blood and stimulated with
PHA in culture. IFN-g levels in these cultureswere comparedwith PHA-stimulated PBMCs collected from untreated NDwhole blood. PBMCs from fMLP-stimulated samplesshowed significantly less IFN-g production at both 48 and72 hours compared with controls (Fig. 5B). These culturesalso contained elevated levels of ArgI, compared with acomplete absence of ArgI present in cultures fromuntreatedPBMCs, confirming that neutrophils activated prior topurification harbor the continued capacity for release ofgranular contents (data not shown).
In addition, sorted ND T cells stimulated in the presenceof increasing numbers of purified, activated neutrophils
Untreated PBMC fMLP PBMC
FSCSSC
CD33
CD11b HLA-DR
A
CD66CD14
CD15
B
2.97 32.8
27.7
2.57
ND
ME
N
GB
M
Arg
I (ng
/mL)
C
ME
T
*
D
fMLP-stimulated PBMCs
CD33hi
CD33lo
Figure 4. CD11bþCD33lo cellswithin GBM PBMCs aredegranulated neutrophils. A,degranulation was induced in NDwhole blood using fMLP; PBMCswere collected from stimulatedblood and matched unstimulatedcontrols and subject to flowcytometry; circular gates representcells with scatter characteristics ofgranulocytes. B, flow cytometricanalysis of gated CD11bþCD33locells within fMLP-degranulated NDPBMCs, stained with CD14,HLA-DR, CD15, and CD66.Corresponding isotypes found inSupplementary Fig. S1. C, ArgIELISA analysis of plasma samplesfrom patients with GBM (n ¼ 6)when compared with ND (n¼ 10) orpatients with meningioma (MEN,n ¼ 14) or metastatic tumors (MET,n ¼ 5; P < 0.0001). D, low (i)- andhigh (ii)-magnification images aswell as immunohistochemicalstaining for CD15 (iii) andmyeloperoxidase (iv) from arepresentative sample of GBMundergoing active necrosis; arrowsmark an area of active necrosis.FSC, forward scatter; SSC, sidescatter.
Sippel et al.
Clin Cancer Res; 17(22) November 15, 2011 Clinical Cancer Research6998
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107
resulted in a concentration-dependent suppression of T-cellproliferation (Supplementary Fig. S2), confirming that neu-trophil degranulation (and ArgI release, noted above)results in functional T-cell suppression in vitro. To providefurther evidence for an association of neutrophil degranu-
lation with GBM-induced immunosuppression, levels ofArgI within media from in vitro GBM T-cell functionalcultures described previously weremeasured and comparedwith ArgI levels within media from ND and control patientcultures. Again, levels of ArgIweremarkedly elevatedwithinmedia harvested fromPHA-stimulatedGBMPBMCs, aswellas within media from MLR containing GBM PBMCs, whencompared with ND or tumor controls (Fig. 5C).
As mentioned above, neutrophils can be found withinregions of active necrosis within GBM specimens. To showthe potential for an ArgI-mediated immunosuppressiveeffect within the tumor microenvironment, we selectivelyharvested necrotic tissue during neurosurgical resection ofGBM and the resulting material was subject to ArgI ELISA.Although there was significant variance between the tumorstested, all necrotic samples showed ArgI levels that weremanifold higher than levels observed in matched plasmasamples, ranging from 171 to 2,946 pg/mL (data notshown).
GBM T-cell function can be restored by targeting ArgIin vitro
Initially, we confirmed that segregation of T cells frompatients with GBM away from activated neutrophils foundwithin PBMCs resulted in a restoration of functional acti-vation. T cells from patients with GBM, as well as ND, werepurified from PBMCs using magnetic bead separation andstimulated 1:1 with Dynabeads for 72 hours. Measurementof IFN-g levels within culture media confirmed that func-tional activity of GBM T cells could be restored to levelsconcurrent with ND T cells (data not shown).
To provide preclinical evidence that targeting ArgI mayprovide benefit for restoring cellular immune function inpatients with GBM, we used several approaches to restorearginine levels within functional in vitro cultures. The ArgIenzymatic pathway was first targeted using the specific ArgIinhibitor nor-NOHA, which was added at a concentrationof 40 mmol/L to the media of T-cell functional assays (PHAstimulation or MLR prepared as described previously). Theaddition of nor-NOHA to the media of functional culturescontaining GBM T cells within bulk PBMCs (which undernormal conditions did not produce IFN-g) resulted in asignificant increase in functional response after stimulationwith PHA (Fig. 5D), restoring IFN-g production to levelscomparable with T cells from ND or patients with otherintracranial tumors. A similar restitution of T-cell functionalresponse was seen in GBM-suppressed MLR in the presenceof nor-NOHA (Fig. 5D).
In an attempt to provide an immediately translatablemechanism for targeting neutrophil-derived ArgI activity,we explored the use of L-arginine supplementation torestore GBM T-cell function in vitro. As seen in experimentsusing nor-NOHA, L-arginine supplementation significantlyincreased in vitro IFN-g production by PHA-stimulatedGBM T cells and similarly reversed suppression of ND TcellswithinMLR containingGBMPBMCs (Fig. 5D). Togeth-er, these data confirm that (i) ArgI exerts a central andreversible role in the suppression of cellular immune
0
50
100
150
200
250
GBM MENND
PHA 48 hPHA 72 h
050
100150200250300350
GBM MENND
MLR 48 hMLR 72 h
0
200
400
600
800
1,000
1,200
1,400
7248
PBMC
fMLP PBMC
INF
-γ (p
g/m
L)
FI, CD3ζ
A
Cel
l num
ber
0
2
4
6
8
10
12
ME
N
GB
M
PIT
NF
I, C
D3ζ
*
B
Arg
I (n
g/m
L)
C
**
**
**
0
100
200
300
400
500
600
700
0
100
200
300
400
500
600
ND
ME
NP
ITG
BM
GB
M +
Arg
GB
M +
NO
HA
ND
ME
NP
ITG
BM
GB
M +
Arg
GB
M +
NO
HA
INF
-γ (p
g/m
L)
DPHA–48 h MLR–48 h
* *
100
80
60
40
20
100 101 102 103 104
Figure 5. ArgI levels correlate with GBM T-cell dysfunction in vitro and invivo. A, representative (left) and averaged (right) flow cytometric analysisof intracellular CD3z expression in direct ex vivo T cells from patients withPIT (blue; n¼ 6), MEN (green; n¼ 6), and GBM (orange; n¼ 5) comparedwith isotype (black; P ¼ 0.024). B, ELISA measurement of PHA-inducedIFN-g production by ND PBMCs, following neutrophil degranulation byfMLP in whole blood prior to PBMC purification, when compared withmatched PBMCs from unstimulated blood at 48 (P ¼ 0.0007) and 72hours (P ¼ 0.015). C, comparative ELISA-based measurements of ArgIlevels in immunofunctional assays (PHA stimulation andMLR) using bulkPBMCs from ND (n ¼ 4) or patients with meningioma (n ¼ 4) or GBM(n¼ 5;P < 0.0001). D, the ArgI inhibitor nor-NOHA (n¼ 4) or supplementalL-arginine (n ¼ 5) was added to (left) PHA-stimulated bulk PBMCs or(right)MLRdeveloped from theaforementionedgroups. IFN-g productionwas measured by ELISA at 48 hours (P < 0.0001 in each case).
Neutrophils and Immunosuppression in GBM
www.aacrjournals.org Clin Cancer Res; 17(22) November 15, 2011 6999
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107

function in patients withGBM, and (ii) that reversal of ArgI-mediated effects through either pharmacologic inhibitionor addition of exogenous L-arginine can restore GBM T-cellfunction to levels equivalent with normal controls.
Discussion
In spite of a long history of study outlining GBM-asso-ciated effects on cellular immunity, there has been littleunderstanding of the underlying factors responsible for theobserved suppression. Our analysis has confirmed that Tcells from patients with GBM show minimal proliferationand IFN-g production upon activation directly ex vivo.However, to our knowledge, the current study representsthe first documentation of increased numbers of degranu-lated neutrophils within the peripheral circulation ofpatients with GBM. This phenomenon has been previouslydescribed in several other human cancers (16, 17, 18),perhaps first outlined by Rodriguez and colleagues in theiranalysis of patients with renal cell cancer. We observedsimilar expression patterns of myeloid-lineage markerswithin the cell population of interest and confirmed theneutrophilic phenotype through histopathologic analysis.As in the renal cell cancer cohort,weobserved increasedArgIin plasma samples frompatientswithGBM.Wehave furtherexpanded upon the hypothetical effect of increased ArgIrelease in vivo by confirming a concomitant decrease insurface CD3z expression on T cells from patients with GBManalyzed directly ex vivo. Importantly, we provide herein thefirst evidence that neutrophil-mediated suppression of T-cell function in patients with cancer can be reversed in vitrousing either selective pharmacologic inhibition of ArgI or,more simply, through the addition of exogenous L-arginine.In addition, the identification of increased frequency ofneutrophils andmassively elevatedArgI levelswithin active-ly necrotic GBM specimens offers not only potential insightinto the ontologic source of degranulated neutrophils inthese patients, but also a potential mechanism throughwhich cellular immunity may be disrupted within thetumor microenvironment.
Myeloid lineage cells with immunosuppressive proper-ties, recently categorized as myeloid-derived suppressorcells (MDSC), have been previously associated with cellularimmunosuppression in a number of disease states (7). Inanimals, phenotypic classification of MDSC has been fairlystraightforward and is well correlated with functional sup-pression of T-cell activity. More recently, increasing data inhumans has confirmed the presence of cells with functionalcharacteristics of MDSC, although the phenotypic nomen-clature remains somewhat less clear (6–9). For the mostpart, MDSC can be subdivided into 2 phenotypic popula-tions having either monocytic or granulocytic characteris-tics, identified in humans as CD14þHLA-DRneg orCD14negHLA-DRneg, respectively. Both subsets can sup-press T-cell function although multiple lineage-specificmechanisms for this effect have been proposed(10, 19, 20). Prior studies have explored immunosuppres-sive qualities of monocytic populations within patients
with GBM (11, 21). Gustafson and colleagues recentlyidentified an expanded population of MDSC within ste-roid-treated patients with GBM, phenotypically defined asCD14þHLA-DRlo/neg (11).Wewere unable to document asimilar monocytic population within our cohort and couldnot identify a difference in monocytic expression patternsassociated with steroid treatment. A potential explanationfor this discrepancy could derive fromdifferingmethods fortissue handling;most notably, all analyses in our studywereconductedwith fresh (1–4hours postresection) PBMCs andfrozen samples were never used. It has been our experiencethat monocytic expression patterns can change rapidly withfreeze–thaw cycles, and neutrophils frompatients or NDdonot survive the freeze–thaw process.
Though it is likely that monocytic populations withinpatients with GBMmay contribute to the observed immu-nosuppressive effect, our studies have shown a strongcorrelation between the presence of degranulated neu-trophils and T-cell dysfunction. The ability of normalgranulocytes to suppress T-cell function has been previ-ously described (22) and has also been linked to immu-nosuppression in non–small cell lung, pancreatic, colonand breast cancer (17, 18). Populations of granulocyticMDSC have also been described in renal cell carcinoma(15) and non–small cell lung cancer (23). Whetherdescribed as MDSC or normal neutrophils, the mecha-nism by which granulocytic cells induce immunosuppres-sion is commonly linked to the release of ArgI into theextracellular environment. While monocytic MDSC–derived immunosuppression has also been linked to ArgIexpression, human monocytic cells tend to deplete extra-cellular L-arginine via increased CAT2B expression andintracellular transport. Transported L-arginine is subse-quently metabolized by intracellular ArgI (a biologiccharacteristic shown by alternatively activated (M2)macrophages; ref. 24). The fact that ArgI is found atincreased extracellular levels within the plasma ofpatients with GBM suggests that the enzyme is releasedfrom the expressing cell, consistent with a granulocyticmechanism of ArgI-mediated immunoregulation.
Neutrophils have been well characterized in their role forpromoting inflammation and combating infections aftertissue damage has occurred (25). With these biologic char-acteristics inmind, it is perhaps no surprise that neutrophilswould be attracted to the tumor microenvironment. Thisphenomenon is particularly likely within GBM, as this fast-growing tumor harbors necrosis as one of its definingpathologic characteristics. In regards to potential sourcesof active neutrophilic recruitment in GBM, previous studieshave shown that pseudopalisading cells surroundingregions of necrosis within GBM release the cytokine inter-leukin (IL)-8, a factor with strong trophic effects uponneutrophils (26, 27). However, given complex cellular andbiologic characteristics of the tumor microenvironment, alarge range of additional candidate mechanisms that mayinduce neutrophilic recruitment and induction of degran-ulation mandate significant further experimentation.Ongoing studies in our group are exploring potential
Sippel et al.
Clin Cancer Res; 17(22) November 15, 2011 Clinical Cancer Research7000
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107

GBM-specific factors that would explain the observed neu-trophilic changes in affected patients.In the current study, we have identified 2 possible
mechanisms through which ArgI-mediated T-cell dysfunc-tion may be reversed in vitro. Perhaps most importantly, wehave confirmed that the simple supplementation of extra-cellular L-arginine to T-cell functional assays can reverse theimmunosuppressive phenotype. Ironically, L-arginine sup-plementation has been previously used within noncancerclinical settings. On an initially empiric basis, oral argininesupplementation was explored and found to show efficacyfor improving immune function in patients sufferingmajortrauma or undergoing extensive surgical procedures (28). Itwas subsequently confirmed that ArgI is transiently found atincreased levels in these patients (29, 30), supporting theclinical utility for dietary L-arginine supplementation in thereversal of immunosuppression. These clinical resultsencourage parallel translation to patients with cancer. OralL-arginine supplementation is clinically attractive due tolow cost, ease of delivery, and negligible toxicity. On thebasis of our recent data,wehave initiated a pilot clinical trialexploring the utility of oral L-arginine supplementation forrestoring endogenous cellular immunity that is, in part,
suppressed by activated neutrophils in newly diagnosedpatients with GBM. Although augmentation of T-cell func-tion by targeting ArgI in vivo may not, in isolation, confersignificant clinical benefit in regards to tumor clearance, wepredict that reversal of ArgI-mediated suppression of cellu-lar immunity may offer a critical therapeutic adjuvant forthe development of effective immunotherapy in patientswith GBM.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Grant Support
This work was supported by grants from the American Cancer Society, theCancer League of Colorado, and the Neurosurgery Research and EducationFoundation.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
ReceivedApril 26, 2011; revised September 1, 2011; accepted September 9,2011; published OnlineFirst September 26, 2011.
References1. Wen PY, Kesari S. Malignant gliomas in adults. N Engl J Med
2008;359:492–507.2. Rolle CE, Sengupta S, Lesniak MS. Challenges in clinical design of
immunotherapy trials for malignant glioma. Neurosurg Clin N Am2010;21:201–14.
3. Waziri A. Glioblastoma-derivedmechanisms of systemic immunosup-pression. Neurosurg Clin N Am 2010;21:31–42.
4. Waziri A, Killory B, Ogden AT III, Cannoll P, Anderson RC, Kent SC,et al. Preferential in situ CD4þCD56þ T cell activation and expansionwithin human glioblastoma. J Immunol 2008;180:7673–80.
5. BrooksWH, Latta RB, Mahaley MS, Roszman TL, Dudka L, Skaggs C.Immunobiology of primary intracranial tumors. Part 5: correlation of alymphocyte index and clinical status. J Neurosurg 1981;54:331–7.
6. Marigo I, Dolcetti L, Serafini P, Zanovello P, Bronte V. Tumor-inducedtolerance and immune suppression by myeloid derived suppressorcells. Immunol Rev 2008;222:162–79.
7. Gabrilovich DI, Nagaraj S. Myeloid-derived suppressor cells as reg-ulators of the immune system. Nat Rev Immunol 2009;9:162–74.
8. Peranzoni E, Zilio S, Marigo I, Dolcetti L, Zanovello P, Mandruzzato S,et al. Myeloid-derived suppressor cell heterogeneity and subset def-inition. Curr Opin Immunol 2010;22:238–44.
9. Serfani P, Borrello I, Bronte V. Myeloid suppressor cells in cancer:recruitment, phenotype, properties and mechanisms of immune sup-pression. Semin Cancer Biol 2006;16:53–65.
10. Mandruzzato S, Solito S, Falisi E, Francescato S, Chiarion-Sileni V,Mocellin S, et al. IL4Raþmyeloid-derived suppressor cell expansion incancer patients. J Immunol 2009;182:6562–8.
11. Gustafson MP, Lin Y, New KC, Bulur PA, O'Neill BP, Gastineau DA,et al. Systemic immune suppression in glioblastoma: the interplaybetween CD14þHLA-DRlo/neg monocytes, tumor factors, and dexa-methasone. Neuro Oncol 2010;12:631–44.
12. Munder M, Mollinedo F, Calafat J, Canchado J, Gil-Lamaignere C,Fuentes JM, et al. Arginase I is constitutively expressed in humangranulocytes and participates in fungicidal activity. Blood 2005;105:2549–56.
13. Fossati G, Ricevuti G, Edwards SW, Walker C, Dalton A, Rossi ML.Neutrophil infiltration into human gliomas. Acta Neuropathol 1999;98:349–54.
14. Rodriguez PC, Zea AH, DeSalvo J, Culotta KS, Zabaleta J, QuicenoDG, et al. L-arginine consumption by macrophages modulates theexpression of CD3 zeta chain in T lymphocytes. J Immunol 2003;171:1232–9.
15. ZeaAH,RodriguezPC,AtkinsMB,HernandezC,Signoretti S, ZabaletaJ, et al. Arginase-producing myeloid suppressor cells in renal cellcarcinoma patients: a mechanism of tumor evasion. Cancer Res2005;65:3044–8.
16. Rodriguez PC, Ernstoff MS, Hernandez C, Atkins M, Zabaleta J, SierraR, et al. ArginaseI-producingmyeloid-derived suppressor cells in renalcell carcinoma are a subpopulation of activated granulocytes. CancerRes 2009;69:1553–60.
17. Rotondo R, Barisione G, Mastracci L, Grossi F, Orengo AM, CostaR, et al. IL-8 induces exocytosis of arginase 1 by neutrophil poly-morphonuclears in nonsmall cell lung cancer. Int J Cancer 2009;125:887–93.
18. Schmielau J, FinnOJ. Activatedgranulocytes andgranulocyte-derivedhydrogen peroxide are the underlying mechanisms of suppression ofT-cell function in advanced cancer patients. Cancer Res 2001;61:4756–60.
19. Youn JI, Srinivas N, Collazo M, Gabrilovich DI. Subsets of myeloid-derived suppressor cells in tumor-bearing mice. J Immunol 2008;181:5791–802.
20. Movahedi K, Guilliams M, Van den Bossche J, Van den Bergh R,Gysemans C, Beschin A, et al. Identification of discrete tumor-induced myeloid-derived suppressor cell subpopulationswith distinct T cell-suppressive activity. Blood 2008;111:4233–44.
21. Rodrigues JC, Gonzalez GC, Zhang L, Ibrahim G, Kelly JJ, GustafsonMP, et al. Normal human monocytes exposed to glioma cells acquiremyeloid-derived suppressor cell-like properties. Neuro Oncol 2010;12:351–65.
22. Munder M, Schneider H, Luckner C, Giese T, Langhans CD, FuentesJM, et al. Suppression of T-cell functions by human granulocytearginase. Blood 2006;108:1627–34.
23. Liu CY,Wang YM,WangCL, Feng PH, KoHW, Liu YH, et al. Populationalterations of L-arginase- and inducible nitric oxide synthase-expressedCD11bþ/CD14�/CD15þ/CD33þmyeloid-derived suppressor cells and
Neutrophils and Immunosuppression in GBM
www.aacrjournals.org Clin Cancer Res; 17(22) November 15, 2011 7001
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107

CD8þ T lymphocytes in patients with advanced-stage non–small celllung cancer. J Cancer Res Clin Oncol 2010;136:35–45.
24. Rodriquez PC, Ochoa AC. Arginine regulation by myeloid derivedsuppressor cells and tolerance in cancer:mechanismsand therapeuticperspectives. Immunol Rev 2008;222:180–91.
25. Nathan C. Neutrophils and immunity: challenges and opportunities.Nat Rev Immunol 2006;6:173–82.
26. Rong Y, Durden D, Van Meir EG, Brat DJ. 'Psuedopalisading' necrosisin glioblastoma: a familiar morphologic feature that links vascularpathology, hypoxia and angiogenesis.J Neuropathol Exp Neurol2006;65:529–39.
27. Waugh DJ,Wilson C. The Interleukin-8 pathway in cancer. Clin CancerRes 2008;14:6735–41.
28. Neilly PJD, Kirk SJ, Gardiner KR, Rowlands BJ. The L-arginine/nitricoxide pathway—biological properties and therapeutic applications.Ulster Med J 1994;63:193–200.
29. Ochoa JB, Bernard AD, O'Brien WE, Griffen MM, Maley ME, RockichAK, et al. Arginase I expression and activity in human mononuclearcells after injury. Ann Surg 2001;233:393–9.
30. TsueiBJ,BernardAC,ShaneMD,Shirley LA,MaleyME,BoulangerBR,et al. Surgery induces human mononuclear cell arginase I expression.J Trauma 2001;51:497–502.
Sippel et al.
Clin Cancer Res; 17(22) November 15, 2011 Clinical Cancer Research7002
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107

2011;17:6992-7002. Published OnlineFirst September 26, 2011.Clin Cancer Res Trisha R. Sippel, Jason White, Kamalika Nag, et al. by Targeting Arginase IPatients with GBM: Restoration of Cellular Immune Function Neutrophil Degranulation and Immunosuppression in
Updated version
10.1158/1078-0432.CCR-11-1107doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2011/09/26/1078-0432.CCR-11-1107.DC1Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/17/22/6992.full#ref-list-1
This article cites 30 articles, 11 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/17/22/6992.full#related-urls
This article has been cited by 8 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
SubscriptionsReprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://clincancerres.aacrjournals.org/content/17/22/6992To request permission to re-use all or part of this article, use this link
Research. on August 26, 2018. © 2011 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 26, 2011; DOI: 10.1158/1078-0432.CCR-11-1107





























