Embed Size (px)
Citation preview

Vol. 38. No. 2INFECTION AND IMMUNITY, Nov. 1982, p. 745-7540019-9567/82/110745-10$02.00/0Copyright © 1982, American Society for Microbiology
Neutralization of Chlamydia trachomatis Infectivity withAntibodies to the Major Outer Membrane Protein
HARLAN D. CALDWELL* AND LINDA J. PERRY
Department of Health and Human Services, Public Health Service, National Institutes of Health, NationalInstitute ofAllergy and Infectious Diseases, Laboratory of Microbial Structure and Function, Rocky
Mountain Laboratories, Hamilton, Montana 59840-2999
Received 17 May 1982/Accepted 6 July 1982
Rabbit immunoglobulin G (IgG) antibodies raised against the major outermembrane protein of the Chlamydia trachomatis lymphogranuloma venereumstrain 434 neutralized the infectivity of the parasite for HeLa 229 cells. Themechanism by which anti-major outer membrane protein IgG prevented C.trachomatis from establishing infection was studied by using intrinsically "4C-radiolabeled elementary bodies. Neutralized elementary bodies were filterablethrough a polycarbonate filter (pore diameter, 600 nm), demonstrating thatreduction in infectivity was not due to the aggregation of elementary bodies bycross-linking IgG. Antibody-neutralized elementary bodies attached to and pene-trated HeLa cells at rates nearly identical to those for infectious organismsexposed to nonneutralizing control IgG. These results suggest that antibodyinterferes with the infectious process of the parasite after its internalization. Anti-major outer membrane protein Fab fragments could not be substituted forneutralizing IgG antibodies. The requirement for intact IgG implies that cross-
linking of antibodies to the major outer membrane protein on the surfaces of theorganisms may be instrumental in neutralization.
Organisms of the genus Chlamydia are obli-gate intracellular bacteria characterized by acomplex, poorly understood growth cycle. Theyexist as two distinct particle types which differfrom each other morphologically, physiological-ly, and biochemically (33). The elementary body(EB) is the infectious form, whereas the reticu-late body is the noninfectious, metabolicallyactive intracellular form of the parasite. The EBinitiates the infectious process by attaching tothe host cell. Once attached, the EB enters thecell by endocytosis. This endocytosis is highlyefficient and is preferential since chlamydiae areingested by nonprofessional phagocytic cellsmore quickly than are other bacteria or inertparticles (6). After endocytosis, the chlamydia-laden phagosome provides the parasite with aprotected intracellular environment for itsgrowth by preventing fusion with the lysosomesof the host cell (9, 36). The ability both to inducephagocytosis and to prevent phagolysosomalfusion is directed by chlamydial cell surfaceantigens (33). The surface structures that medi-ate these interesting biological responses areunknown. Only recently have investigators be-gun to probe the chlamydial cell surface. Prepa-ratory methods for the isolation of chlamydialouter membranes have recently been described,and preliminary analysis has shown that a single
protein species predominates in the chlamydialouter membrane (7). This protein is exposed onthe surface of the organism (7, 15, 32) and isestimated to constitute as much as 60% of thetotal outer membrane protein (7). Because of itsabundance, it has been termed the major outermembrane protein (MOMP). Nothing is knownabout the biological function of this protein withrespect to chlamydia-host relationships. For ex-ample, is the MOMP important in the attach-ment of chlamydiae to host cells? Is it a viru-lence factor that induces phagocytosis orinhibits phagolysosomal fusion?
In previous investigations, we have focusedmuch of our attention on defining the antigenic(8) and structural (6a) properties of the MOMPof chlamydiae. In the course of these studies,hyperimmune antisera have been made to puri-fied sodium dodecyl sulfate (SDS)-denaturedMOMPs isolated from several Chlamydia tra-chomatis strains. These sera were immunologi-cally specific for the MOMP and were highlyreactive with intact chlamydiae as determinedby microimmunofluorescence. We thought thatantibodies against the MOMP might be valuableprobes to explore the biological function of thisprotein. In this study, we show that anti-MOMPimmunoglobulin G (IgG) neutralizes chlamydialinfectivity for HeLa 229 cells. Our data reveal
745

746 CALDWELL AND PERRY
that neutralization mediated by anti-MOMP IgGdoes not affect either chlamydial attachment orpenetration but inhibits infectivity of the orga-nism after its internalization. The possible bio-logical mechanisms that may be involved in thisneutralization process are discussed.
MATERIALS AND METHODSOrganisms and growth conditions. The C. trachoma-
tis lymphogranuloma venereum (LGV-434) serotypeL2 was used throughout this study. Organisms weregrown in HeLa 229 cells as previously described (7).EBs were purified (7) and quantitated by calculatinginclusion-forming units (IFU) as described by Furnesset al. (10). HeLa cells were periodically tested formycoplasma with standard culture methods and werenegative.
Radiolabeling of organisms. HeLa 229 cell monolay-ers of 2 x 107 cells grown in 150-cm2 plastic tissueculture flasks were infected for 1 h at 37°C with 1 ml ofserotype L2 inoculum containing approximately 107IFU. The inoculum was removed, and monolayerswere fed with 50 ml of minimal essential mediumsupplemented with 10% fetal bovine serum (MEM-10).After 8 to 10 h of incubation at 37°C, the medium wasremoved and replaced with 50 ml of MEM-10 contain-ing 1 ,ug of emetine hydrochloride (Sigma ChemicalCo., St. Louis, Mo.) per ml. The cultures were incu-bated at 37°C for 2 h, at which time the medium wasagain removed and replaced with 50 ml of MEM-10containing one-tenth the normal amino acid concentra-tion, 1 p.g of emetine per ml and 1 ,uCi of "4C-aminoacids (specific activity, 1.88 mCi/mg; ICN, Chemicaland Radioisotope Division, Irvine, Calif.) per ml. Thecultures were then incubated for an additional 30 h, atwhich time chlamydiae were harvested, and '4C-la-beled EBs were purified by centrifugation throughdiscontinuous Renografin density gradients (7). Puri-fied "4C-labeled EBs were suspended in 0.25 M su-crose-10 mM sodium phosphate-5 mM L-glutamicacid, pH 7.2 (SPG). The suspension was divided into1-ml portions and kept frozen at -74°C until used. In10% cold trichloroacetic acid, 99% of the radioactivitywas precipitable.IgG and Fab preparation. Hyperimmune rabbit anti-
sera to the serotype L2 39,500-molecular-weightMOMP and to ovalbumin (Sigma) were prepared aspreviously described (8). The IgG fraction of theseantisera was isolated by adsorption to Protein A-Sepharose CL-4B (Sigma) as described by Goding(11). Briefly, 10 ml of serum was passed through acolumn (0.9 by 10 cm) of Protein A-Sepharose CL-4Bpreviously equilibrated with 50 mM NaPO4-0.15 MNaCl-0.02% NaN3, pH 7.2 (PBS-Az). The columnwas then washed with 10 volumes of PBS-Az. BoundIgG was eluted with 30 ml of 0.25 M acetic acid, pH3.2, and dialyzed against three changes of 100 volumeseach of cold phosphate-buffered saline without NaN3.The protein concentration of isolated IgG was deter-mined by the method of Lowry et al. (24), and workingsolutions were adjusted to 10 mg/ml. IgG prepared bythis procedure yielded a single immunoprecipitin lineby immunodiffusion with guinea pig antisera preparedagainst whole rabbit serum and was homogeneousafter SDS-polyacrylamide gel electrophoresis (7) un-
der nonreducing conditions. Fab fragments were pre-pared by papain hydrolysis of purified rabbit IgG asdescribed by Porter (30) and modified by McLaughlinand Pitot (25), except that fraction 1 obtained aftercarboxymethyl cellulose column fractionation (25) wasdialyzed against distilled water and then was rechro-matographed on a Protein-A Sepharose CL-4B col-umn. This step was necessary to remove smallamounts of undigested IgG that cofractioned with theFab fragments by carboxymethyl cellulose columnchromatography. The purity of isolated Fabs wasdetermined by SDS-polyacrylamide gel electrophore-sis under reducing and nonreducing conditions andwas in agreement with that observed by other investi-gators (28). The biological activity and the specificityof isolated Fab fragments prepared from anti-MOMPIgG were determined by the ability of Fabs to inhibitprecipitation of purified MOMP antigen (7) by doubleimmunodiffusion and by the binding of 125I-radiola-beled Fabs to purified EBs.
Ultrafiltration. Suspensions (2 ml) of '4C-radiola-beled EBs (8.9 x 105 IFU, 5.8 x 104 cpm) in SPGbuffer were filtered through 13-mm polycarbonatefilters (pore diameters, 200, 400, 600, 800, and 1,000nm; Nuclepore Corp., Pleasanton, Calif.) followingthe instructions of the manufacturer. The filterabilityof EBs for each membrane pore size was determinedby calculating the percentages of both radioactivityand IFU found in each of the polycarbonate membraneultrafiltrates. These experiments were done to selectthe appropriate filter for antibody-mediated neutraliza-tion experiments. When antibody-pretreated 14C-la-beled EBs were used in filtration experiments, only600-nm polycarbonate filters were used. For theseexperiments, an equal volume of IgG solution dilutedin SPG was added to 14C-labeled EBs to give a finalconcentration of 6.8 x 105 IFU (4 x 104 cpm) per ml.The suspension was incubated at 37°C for 30 min andthen was immediately filtered. The percentages ofradioactivity and IFU found in the ultrafiltrates weredetermined, and these values were used to evaluatethe role of aggregation as a mechanism in chlamydialneutralization.
Neutralization assays. All assays were done in tissueculture chamber slides (eight chambers per slide, 100mm per chamber; Lab-Tek Products, Miles Labora-tories, Naperville, Ill.) containing 105 HeLa 229 cellsper chamber. To perform the assay, 14C-labeled EBswere diluted in SPG buffer to give a final concentrationof 1.36 x 106 IFU/ml. This was then mixed with anequal volume of either IgG or Fab fragments in phos-phate-buffered saline to give a final concentration of6.8 x 105 IFU/ml. In experiments involving comple-ment, guinea pig serum was added to the antibody-organism reaction mixture to give a final dilution of1:32. The 14C-labeled EB-antibody mixtures wereimmediately incubated for 30 min at 37°C in a shakingwater bath. After this incubation, 100 ,I of the suspen-sion (6.8 x 104 IFU) was placed directly onto HeLacell monolayers and was incubated at 37°C with 5%CO2 for various lengths of time depending on theparticular experimental protocol. After incubation, theinoculum was removed, and the monolayers werewashed twice with 200 ,ul of Hanks balanced saltsolution (HBSS) and then were refed with 200 ,ul ofMEM-10. Cells were incubated for 42 h at 37°C with
INFECT. IMMUN.

NEUTRALIZATION OF C. TRACHOMATIS INFECTIVITY 747
FIG. 1. Anti-MOMP neutralization of C. trachomatis infectivity. (A) HeLa 229 cells inoculated with 6.8 x iO'IFU of C. trachomatis serotype L2 pretreated with a 1:32 dilution of rabbit anti-ovalbumin IgG. Numerousintracellular cytoplasmic inclusions are evident. Two inclusions (i) are identified. (B) HeLa 229 cells inoculatedwith 6.8 x 104 IFU of C. trachomatis serotype L2 pretreated with a 1:32 dilution of anti-MOMP IgG. Note theabsence of cytoplasmic inclusions. The concentration of anti-ovalbumin and anti-MOMP IgG before dilution was10 mg/ml. Monolayers were Giemsa stained 42 h after inoculation of IgG-treated chlamydiae.
VOL. 38, 1982

748 CALDWELL AND PERRY
5% C02, fixed in absolute methanol, and then wereGiemsa stained. IFU values were calculated by count-ing at least 30 microscopic fields (450x) for eachstained preparation. This procedure was followed forexperiments involving chlamydial attachment and en-try with the following modifications:
(i) For attachment assays, 100 ,ul of 14C-labeled EBinoculum was adsorbed to HeLa cell monolayers for 2,15, 30, 45, 60, and 90 min. Six monolayers wereinoculated for each time period. After adsorption theinoculum was removed, and the monolayers werewashed three times with 200 ,ul of HBSS. Three of themonolayers were refed with MEM-10 and were incu-bated at 37°C for 42 h. These were then fixed, Giemsastained, and used to determine IFU values. The otherthree monolayers were lysed with 200 pL1 of 2% (wt/vol) SDS in distilled water. The solubilized cells weretransferred to a 10-ml glass scintillation vial. The wellswere then washed twice with 200 ,u1 of the SDSsolution, and this material was pooled with the originalSDS-cell lysate. To this suspension 10 ml of Aquasol(New England Nuclear Corp., Boston, Mass.) wasadded, and the radioactivity in the sample was deter-mined with a Beckman LS8100 liquid scintillationcounter (Beckman Instruments, Inc., Fullerton, Cal-if.).
(ii) For assays to determine the effect of antibodytreatment on chlamydial penetration, the washedmonolayers were incubated for 90 min with inoculumand were refed with 200 p.1 of MEM-10. At 2, 4, 6, 10,24, and 48 h after the addition of the inoculum, themedium was removed and discarded, and the mono-layers were assayed for infectivity and intracellularradioactivity. For infectivity titrations, monolayerswere refed with MEM-10 and were reincubated for atotal of 42 h. At this time they were fixed and stained,and IFU values were determined. To assay intracellu-lar radioactivity, monolayers were washed twice withHBSS and then twice with 200 p.l of trypsin (100 ,ug/ml; Sigma) in phosphate-buffered saline 10 mM sodiumphosphate, 0.15 M NaCl; pH 7.2). A 200-,ul sample oftrypsin solution was then added to each monolayer,and the cells were incubated for 10 to 15 min at roomtemperature until they had detached from the glass.The cell suspensions were refluxed several times andtransferred to 2-ml conical glass centrifuge tubes. Thetrypsinized glass surface was washed once with 200 p.lof HBSS, and this material was pooled with theoriginal trypsinized cell suspensions. The cell suspen-sions were centrifuged at 500 x g for 5 min at 4°C. Thesupernatants were removed; and 200 p.l of a 2% SDSsolution was added to each cell pellet to lyse the cells.The cell lysates were transferred to scintillation vials.The centrifuge tubes were then washed twice with 200pl. of the SDS solution, and this was pooled with eachof the original SDS cell lysates. A 10-ml portion ofAquasol was added to each of the cell lysates, and theradioactivity was determined. The radioactivity recov-ered in these cells was taken to represent internalizedEBs.
RESULTSAnti-MOMP-mediated neutralization. Neutral-
ization was determined by the ability of IgGantibodies to prevent the inclusion-forming ca-
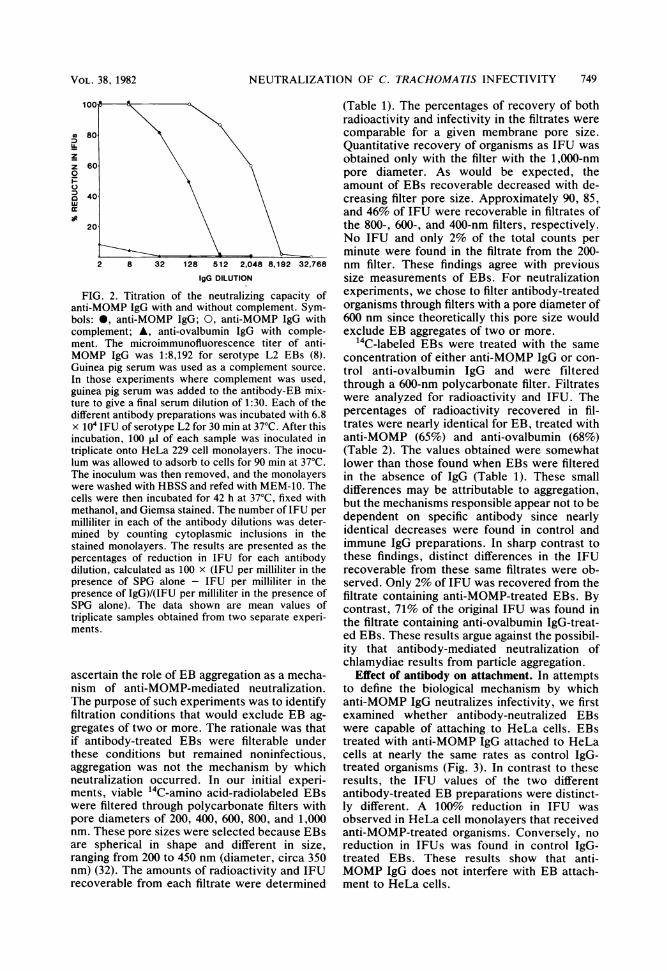
pability of the organism. Differences observed inintracytoplasmic inclusions found 42 h afterHeLa cell monolayers were inoculated with EBstreated with either control IgG or anti-MOMPIgG are shown in Fig. 1. Numerous cytoplasmicinclusions were evident in HeLa cells inoculatedwith 6.8 x 104 serotype L2 IFU that werepretreated with anti-ovalbumin IgG (Fig. 1A). Incontrast, no inclusions were found in HeLa cellmonolayers inoculated with the same number ofEBs pretreated with anti-MOMP IgG (Fig. 1B).The neutralizing titer of anti-MOMP IgG wasdetermined by limiting dilution both with andwithout guinea pig complement (Fig. 2). Signifi-cant chlamydial-neutralizing activity (approxi-mately 50% reduction in IFU) was obtained at a1:128 dilution of anti-MOMP IgG. The neutraliz-ing titer was substantially increased when com-plement was added to the antibody-organismreaction mixture. With complement present, a60% reduction in IFU was obtained at a 1:2,048dilution of anti-MOMP IgG. Anti-ovalbumin IgGeither with or without complement did not resultin significant neutralization, demonstrating thespecificity of the neutralization reaction ob-served with anti-MOMP IgG. The mechanism bywhich complement enhanced the neutralizingactivity of antibody was not studied further.Instead, we focused our attention on the mecha-nism involved in antibody-mediated neutraliza-tion. It was thought that studying the nature ofthis neutralization might yield information aboutthe biological function of the MOMP.
Determination of the mechanism of anti-MOMP-mediated neutralization. The mechanismby which anti-MOMP IgG neutralized chlamydi-al infectivity in the experiments described abovemight have two alternate or cooperative expla-nations. The first would involve a physical re-duction in the number of EBs by cross-linkingchlamydial particles with bivalent IgG. Reduc-tion in infectivity would then simply be a resultof the aggregation of many infectious particlesinto a few large infectious units. The secondexplanation would involve the binding of anti-body to the MOMP, and this MOMP-boundantibody would then affect some step or steps inthe infectious process of the organism.
Effect of antibody on aggregation. In our initialstudies, experimental conditions were designedwhich minimized the complex secondary anti-gen-antibody interactions required for aggrega-tion. For example, the concentration of EBantigen was small (6.8 x 105 IFU/ml of reactionmixture), and the 30-min incubation time ofantibody and antigen was minimal. Althoughthese conditions were unfavorable for particleaggregation, they could not justify discardingaggregation as a factor in neutralization. Filtra-tion experiments were therefore designed to
INFECT. IMMUN.
NEUTRALIZATION OF C. TRACHOMATIS INFECTIVITY 749
,80\
z
z 60X0
40w
20
2 8 32 128 512 2,048 8,192 32,768
IgG DILUTION
FIG. 2. Titration of the neutralizing capacity ofanti-MOMP IgG with and without complement. Sym-bols: 0, anti-MOMP IgG; 0, anti-MOMP IgG withcomplement; A, anti-ovalbumin IgG with comple-ment. The microimmunofluorescence titer of anti-MOMP IgG was 1:8,192 for serotype L2 EBs (8).Guinea pig serum was used as a complement source.In those experiments where complement was used,guinea pig serum was added to the antibody-EB mix-ture to give a final serum dilution of 1:30. Each of thedifferent antibody preparations was incubated with 6.8x 104 IFU of serotype L2 for 30 min at 370C. After thisincubation, 100 ,ul of each sample was inoculated intriplicate onto HeLa 229 cell monolayers. The inocu-lum was allowed to adsorb to cells for 90 min at 37°C.The inoculum was then removed, and the monolayerswere washed with HBSS and refed with MEM-10. Thecells were then incubated for 42 h at 37°C, fixed withmethanol, and Giemsa stained. The number of IFU permilliliter in each of the antibody dilutions was deter-mined by counting cytoplasmic inclusions in thestained monolayers. The results are presented as thepercentages of reduction in IFU for each antibodydilution, calculated as 100 x (IFU per milliliter in thepresence of SPG alone - IFU per milliliter in thepresence of IgG)/(IFU per milliliter in the presence ofSPG alone). The data shown are mean values oftriplicate samples obtained from two separate experi-ments.
ascertain the role of EB aggregation as a mecha-nism of anti-MOMP-mediated neutralization.The purpose of such experiments was to identifyfiltration conditions that would exclude EB ag-gregates of two or more. The rationale was thatif antibody-treated EBs were filterable underthese conditions but remained noninfectious,aggregation was not the mechanism by whichneutralization occurred. In our initial experi-ments, viable 14C-amino acid-radiolabeled EBswere filtered through polycarbonate filters withpore diameters of 200, 400, 600, 800, and 1,000nm. These pore sizes were selected because EBsare spherical in shape and different in size,ranging from 200 to 450 nm (diameter, circa 350nm) (32). The amounts of radioactivity and IFUrecoverable from each filtrate were determined
(Table 1). The percentages of recovery of bothradioactivity and infectivity in the filtrates werecomparable for a given membrane pore size.Quantitative recovery of organisms as IFU wasobtained only with the filter with the 1,000-nmpore diameter. As would be expected, theamount of EBs recoverable decreased with de-creasing filter pore size. Approximately 90, 85,and 46% of IFU were recoverable in filtrates ofthe 800-, 600-, and 400-nm filters, respectively.No IFU and only 2% of the total counts perminute were found in the filtrate from the 200-nm filter. These findings agree with previoussize measurements of EBs. For neutralizationexperiments, we chose to filter antibody-treatedorganisms through filters with a pore diameter of600 nm since theoretically this pore size wouldexclude EB aggregates of two or more.
14C-labeled EBs were treated with the sameconcentration of either anti-MOMP IgG or con-trol anti-ovalbumin IgG and were filteredthrough a 600-nm polycarbonate filter. Filtrateswere analyzed for radioactivity and IFU. Thepercentages of radioactivity recovered in fil-trates were nearly identical for EB, treated withanti-MOMP (65%) and anti-ovalbumin (68%)(Table 2). The values obtained were somewhatlower than those found when EBs were filteredin the absence of IgG (Table 1). These smalldifferences may be attributable to aggregation,but the mechanisms responsible appear not to bedependent on specific antibody since nearlyidentical decreases were found in control andimmune IgG preparations. In sharp contrast tothese findings, distinct differences in the IFUrecoverable from these same filtrates were ob-served. Only 2% of IFU was recovered from thefiltrate containing anti-MOMP-treated EBs. Bycontrast, 71% of the original IFU was found inthe filtrate containing anti-ovalbumin IgG-treat-ed EBs. These results argue against the possibil-ity that antibody-mediated neutralization ofchlamydiae results from particle aggregation.
Effect of antibody on attachment. In attemptsto define the biological mechanism by whichanti-MOMP IgG neutralizes infectivity, we firstexamined whether antibody-neutralized EBswere capable of attaching to HeLa cells. EBstreated with anti-MOMP IgG attached to HeLacells at nearly the same rates as control IgG-treated organisms (Fig. 3). In contrast to theseresults, the IFU values of the two differentantibody-treated EB preparations were distinct-ly different. A 100% reduction in IFU wasobserved in HeLa cell monolayers that receivedanti-MOMP-treated organisms. Conversely, noreduction in IFUs was found in control IgG-treated EBs. These results show that anti-MOMP IgG does not interfere with EB attach-ment to HeLa cells.
VOL. 38, 1982

750 CALDWELL AND PERRY
TABLE 1. Filterability of "C-labeled EBs through polycarbonate filters with various pore sizes"
Pore size '4C-labeled EBs found in filtrateb Infectivity found in filtrate"(nm) Amt (cpm/ml) % of total Amt (IFU/ml) % of total
200 1.3 x 103 2 0 0400 2.2 x 104 38 4.1 x 105 46600 4.4 x 104 76 7.6 x 105 85800 4.7 x 104 81 8.0 x 105 90
1,000 4.8 x 104 83 8.6 x 105 97a A 2-ml portion of 14C-radiolabeled EBs was filtered for each pore size. Samples of each filtrate were then
assayed for radioactivity and infectivity (IFU) as described in the text.b Total filtered, 5.8 x 104 cpm/ml for each pore size.Total filtered, 8.9 x 105 IFU/ml for each pore size.
Effect of antibody on entry. The above resultsindicate that neutralization occurs at a stage ofthe infectious process of the parasite after itsattachment to the host cell membrane. Becausechlamydiae induce phagocytosis (6), we askedwhether anti-MOMP IgG might be inhibiting thisrather unusual biological function. HeLa cellmonolayers inoculated with 14C-labeled EBstreated with anti-MOMP or control IgG weretested for the amount of intracellular radioactiv-ity at various times during a 48-h period. Heat-killed EBs were included as a positive controlsince they enter cells at reduced rates comparedwith viable EBs (4). To ensure that only intracel-lular EBs were being measured, monolayerswere treated with trypsin to remove EBs thatwere attached but had failed to enter HeLa cells.Trypsin not only removes chlamydial host cellreceptors but also disassociates chlamydiaebound to host cell membranes (4, 5; unpublisheddata). Trypsin-treated cells were then lysed withSDS, and the radioactivity was determined. Theradioactivity associated with the lysed cell sus-pensions was taken to represent only internal-ized organisms. Infectivity determinations werealso performed at each time period. As expect-ed, heat-treated EBs were noninfectious andwere clearly less efficient in entering HeLa cellsthan were unheated control organisms (Fig. 4).
Organisms treated with anti-MOMP IgG werenot unlike heat-killed chlamydiae in that theywere noninfectious for HeLa cells. They dif-fered, however, from killed EBs in that theyentered cells at a rate similar to that of infectiousorganisms which were treated with control IgG.These data suggest that the effect of anti-MOMPIgG on infectivity occurs after internalization ofthe parasite. In attempts to define more clearlythe role of antibody in chlamydial neutralization,we prepared monovalent Fab fragments fromanti-MOMP IgG and tested their capacity toneutralize. Anti-MOMP Fab fragments did notneutralize infectivity. The inability of Fabs toneutralize was not attributed to a lack of immu-noreactivity since these fragments inhibited im-munoprecipitation of purified MOMP by an-ti-MOMP sera in gel immunodiffusion and125I-labeled Fabs were found to specifically bindviable EBs (data not shown). The data indicatethat cross-linking of IgG to the MOMP is re-quired for neutralization. The biological mecha-nism involved in the process is unknown.
DISCUSSION
Antibody-mediated neutralization of chlamyd-ial infectivity has been described previously invarious host cell systems. In those studies,
TABLE 2. Filterability of antibody-treated EBs'
"4C-labeled EBs Infectivity
Amt (cpm/ml) Amt (IFU/ml)Antibody %o in %S in
filtrationb In filtrate filtrationh In filtrate filtrate"Anti-ovalbumin 3.4 x 104 2.3 x 104 68 2.4 x 105 1.7 x 105 71Anti-MOMP 3.7 x 104 2.4 x 104 65 6.6 x 103 4.4 x 103 2
a Each value is the mean of triplicate samples for two separate experiments. A 1:32 dilution of anti-MOMP oranti-ovalbumin IgG in SPG buffer was incubated with 14C-radiolabeled EBs (4 x 104 cpm, 6.8 x 105 IFU) at 370Cfor 30 min in a shaking water bath.
b Values obtained after 30-min incubation with lgG but without filtration.Percentage of IFU was calculated by dividing IFU found in the filtrates by IFU found in the anti-ovalbumin
IgG control before filtration (2.4 x 105 IFU/ml).
INFECT. IMMUN.

NEUTRALIZATION OF C. TRACHOMATIS INFECTIVITY 751
p. - - -- - - - --_ - -- .- - - -4
III
I
II
iI
0 15 30 45 60MINUTES
FIG. 3. Effect of anti-MOMP IgG on the attachment of "4C-amino acid-labeled L2 EBs to HeLa 229 cells(solid lines) and percentage of reduction in IFU (dashed line). In these experiments, 6.8 x 105 IFU of 14C-radiolabeled EBs (4 x 104 cpm) were incubated with either a 1:32 dilution of anti-MOMP IgG (0) or anti-ovalbumin IgG (0) at 37°C for 30 min. A 100-,ul portion (4 x 103 cpm) of this antibody-EB mixture was inoculatedonto 105 HeLa cells. Six HeLa cell monolayers were inoculated for each sampling time shown for both antibodypreparations. At the times designated, the inoculum was removed, and the monolayers were washed three timeswith HBSS. Three of the monolayers were refed with MEM-10 and incubated at 37°C for 42 h. They were thenfixed, Giemsa stained, and used to determine IFU values. The remaining three monolayers were lysed with 200,ul of 2% SDS. The lysates were each mixed with 10 ml of scintillation cocktail, and the radioactivity associatedwith the cell lysates was determined. For a given experiment, the values obtained from triplicate samples at eachtime period were used to calculate the mean. The data shown are mean values from three separate experiments.
neutralization was described by using eitherantisera against whole organisms (1, 13, 20, 31)or secretions from chlamydia-infected tissues (2,12, 27). Neither the antibody specificity in-volved in neutralization nor the mechanism bywhich neutralization occurs was studied bythose investigators. The studies presented hereare the first to describe in vitro neutralization ofC. trachomatis infectivity with antibody againsta characterized surface antigen of the parasite.Our data clearly demonstrate that IgG antibod-ies against the MOMP of C. trachomatis neutral-ize the infectivity of the organism for HeLa 229cells. In addition, we have attempted to ascer-tain the means by which MOMP-specific anti-body inactivates chlamydial infectivity.On the basis of our findings, the following
points can be made about the mechanisms ofanti-MOMP-mediated neutralization. (i) Neu-tralization was independent of complement anddid not occur extracellulary as a result of EBaggregation (Table 2). (ii) Neither the attach-ment (Fig. 3) nor entry (Fig. 4) phase of theinfectious cycle of the parasite was blocked byneutralizing MOMP antibody, demonstratingthat inactivation of infectivity occurred after theinternalization of the parasite. (iii) MonovalentFab antibody fragments could not substitute fordivalent IgG, suggesting that the intact IgGmolecule is functionally important in the neutral-ization process. We considered the possibilitythat the requirement for intact IgG for neutral-
ization is due to the Fc portion of the moleculeand that HeLa 229 cells, being highly undifferen-tiated, may express Fc-like receptors. If thisshould be the case, binding of IgG to thesereceptors might result in host-mediated ratherthan chlamydia-mediated endocytosis, similar tothe uptake of opsonized organisms via Fc-medi-ated endocytosis by professional phagocyticcells. This type of endocytosis could result inphagolysosome fusion and destruction of chla-mydiae. We tested for the presence of Fc recep-tors on HeLa 229 cells by using rabbit antibody-sensitized sheep erythrocytes. We could notdemonstrate rosetting of sensitized erythrocytesonto HeLa 229 cell monolayers, suggesting thatthese cells lack Fc receptors. These data argueagainst HeLa cell-mediated endocytosis of IgG-coated EBs. In addition, the rates of attachmentand uptake of neutralized and nonneutralizedEBs were nearly identical (Fig. 3 and 4). Thesefindings would be unexpected if chlamydiaewere attaching and entering HeLa cells by dif-ferent host cell receptor-mediated events.Although clearly open to speculation at this
point, the presence of intact or cross-linked IgGon the EB surface could (i) interfere with chla-mydia-directed inhibition of phagolysosome fu-sion, (ii) provide more structural rigidity to thechlamydial outer membrane, which prevents thereorganization of EBs to reticulate bodies, or
(iii) inhibit the transport or metabolism of host-derived nutrients required for further reorgani-
-J-Jw
.. 14-J
I lo 12
- X 10
muJ
100
80 u,U-
z60 -z
0
40 D
ae
.20
090
VOL. 38, 1982

752 CALDWELL AND PERRY
c)
wcmJc:oLJ E4I-CC o_
wgC
_z
--0-<-- - -O - - -
100
80 =U-
z60 z
0
40 awcc:
20
HOURS
FIG. 4. Effect of anti-MOMP IgG on the entry of '4C-amino acid-radiolabeled serotype L2 EBs into HeLa 229cells. Symbols: 0, anti-MOMP IgG; 0, anti-ovalbumin IgG; A, EBs heated at 60°C for 10 min; solid lines,intracellular EBs; dashed lines, percentage of reduction in IFU. In these experiments, 6.8 x 105 IFU of 14C-radiolabeled EBs (4 x 104 cpm) were incubated with either anti-MOMP IgG or anti-ovalbumin IgG at 370C for 30min. The same number of labeled EBs was also heated at 60°C for 10 min. A 100-,ul portion of each suspensionwas then inoculated onto an HeLa 229 cell monolayer (105 cells). After a 90-min incubation period the inoculumwas removed, and the cell monolayers were washed three times with 200 ,ul of HBSS. The cells were then refedwith MEM-10, and at the times shown triplicate cultures were assayed for intracellular chlamydiae as describedin the text.
zation activities of the parasite. There is evi-dence to support some of these proposedmechanisms. Both Friis (9) and Wyrick et al.(36) have shown by transmission electron mi-croscopy that when EBs of Chlamydia psittaciare treated with homologous antiserum raisedagainst whole organisms, they lose their abilityto prevent phagolysosomal fusion. If the chla-mydial MOMP functions in preventing phagoly-sosome fusion, the binding of IgG to this proteincould be inhibitory. If this is the case, divalentantibody appears to be necessary to block thisfunction since monovalent Fab antibody frag-ments failed to neutralize infectivity. Alterna-tively, cross-linking the MOMP with IgG mayrestrict the fluidity of the outer membrane,which would mean that outer membrane compo-nents other than the MOMP may be preventedfrom interacting with host receptors. If thesemembrane components are virulence macromol-ecules, their function may be inhibited becauseof decreased membrane fluidity. There is evi-dence to substantiate this idea. Nicolson andPainter (28) have shown that aggregation of thehuman erythrocyte cytoskeleton protein spec-trin with antispectrin IgG (but not antispectrinFab fragments) controls the lateral distributionof membrane glycoproteins. Conceptually, asimilar mechanism may function in neutralizinginfectivity by interfering with the transition ofthe infectious EBs into the noninfectious meta-bolically active reticulate bodies. During this
poorly understood transition, the EBs increasein size approximately fivefold without undergo-ing cell division. The structural and physiologi-cal characteristics of the outer membrane arealmost certainly changed during this period toallow for the transport of host cell macromol-ecules that chlamydiae require for growth (14,26). Cross-linking of anti-MOMP IgG on thesurfaces of EBs might prevent these transforma-tional and physiological membrane changesfrom occurring. Obviously, to understand theprecise mechanism by which anti-MOMP IgGneutralizes infectivity, detailed ultrastructuraland cytochemical studies will be required. Nev-ertheless, the findings reported here provide forexciting investigation of chlamydia-host rela-tionships.The effect of antibody on the intracellular
multiplication of other intracellular parasites hasbeen studied. Antibody treatment of Legionellapneumophila, a facultative intracellular bacteri-um, promotes its binding to human monocytesbut does not inhibit the intracellular multiplica-tion of the organism (18, 19). Similarly, Hinrichsand Jerells (17) reported that antiserum againstthe obligate intracellular rickettsial organismCoxiella burnetii fails to inhibit intracellular rep-lication of this parasite in macrophages. Thesefindings differ from those reported here for chla-mydiae and those reported by Jones (21) andmore recently by Hauser and Remington (16) forthe obligate intracellular protozoan parasite
INFECT. IMMUN.

NEUTRALIZATION OF C. TRACHOMATIS INFECTIVITY 753
Toxoplasma gondii. The latter investigatorshave shown that monoclonal antibodies specificfor membrane-associated antigenic determinantsof T. gondii facilitate uptake of the organisms bymacrophages and prepare them for intracellulardestruction. Considering the differences withinthis diverse group of intracellular parasites withrespect to their host interactions, differences intheir susceptibilities to antibody are not undulybothersome. For example, C. burnetii multipliesin phagolysosomes (3). Coating the surfaces ofthese parasites with antibody may actually bebeneficial since it could facilitate their intracellu-lar compartmentalization into lysosomes via Fcor other host cell receptor-mediated endocytosisevents. In contrast, both chlamydiae and toxo-plasmata (22, 23) multiply in phagosomes, andunlike the aforementioned organisms, each ofthese parasites has evolved highly specializedcell surface structures both for entering cells andfor preventing phagolysosome fusion. Conse-quently, antibody bound to their surfaces maybe damaging since it could interfere with thefunction of these specialized macromolecules.
Considering the findings of this study, itwould be most interesting to determine if anti-bodies against the MOMP of chlamydiae areprotective in vivo. Recently, Williams et al. (34)have described a murine model of chlamydialpneumonia by using the C. trachomatis mousepneumonitis strain. In studying the immune re-sponse of the host to the mouse pneumonitisagent, they found that serum antibody plays asignificant role in the resistance of mice tochlamydial infection (35). The murine pneumo-nia model provides an excellent opportunity forstudying the immune response of the host to theMOMP of the mouse pneumonitis agent. Prepar-ative biochemical fractionation procedures areavailable for the purification of large quantitiesof homogeneous MOMP from chlamydiae (7);this MOMP could be used as an immunogen insuch studies. It is hoped that the findings report-ed here will prompt such investigations.
ACKNOWLEDGMENTSWe thank Jim Simmons for expert technical assistance,
Susan Smaus for preparation of the manuscript, Chuck Taylorand Bob Evans for illustration and photography, Alan Liss ofthe Laboratory of Microbial Structure and Function for testingHeLa cells for mycoplasma, and the staff of the Laboratory ofMicrobial Structure and Function for helpful discussions andmanuscript review.
LITERATURE CITED
1. Banks, J., B. Eddie, M. Suny, N. Sugg, J. Schachter, andK. F. Meyer. 1970. Plaque reduction technique for demon-strating neutralizing antibodies for Chlamydia. Infect.Immun. 2:443-447.
2. Barenfanger, J., and A. B. MacDonald. 1974. The role ofimmunoglobulin in the neutralization of trachoma infectiv-ity. J. Immunol. 113:1607-1617.
3. Burton, P. R., N. Kordova, and D. Paretsky. 1971. Elec-
tron microscope studies of the rickettsia Coxiella burnetii:entry, lysosomal response, and fate of rickettsial DNA inL cells. Can. J. Microbiol. 17:143-150.
4. Byrne, G. I. 1976. Requirements for ingestion of Chla-mydia psittaci by mouse fibroblasts (L cells). Infect.Immun. 14:645-651.
5. Byrne, G. I. 1978. Kinetics of phagocytosis of Chlamydiapsittaci by mouse fibroblasts (L cells): separation of theattachment and ingestion stages. Infect. Immun. 19:607-612.
6. Byrne, G. I., and J. W. Moulder. 1978. Parasite-specifiedphagocytosis of Chlamydia psittaci and Chlamydia tra-chomatis by L and HeLa cells. Infect. Immun. 19:598-606.
6a.Caldwell, H. D. 1982. Structural analysis of the majorouter membrane protein of Chlamydia spp., p. 45-50. InP.-A. Mardh et al. (ed.), Chlamydial infections. Elsevier-North Holland, Amsterdam.
7. Caldwell, H. D., J. Kromhout, and J. Schachter. 1981.Purification and partial characterization of the major outermembrane protein of Chiamydia trachomatis. Infect. Im-mun. 31:1161-1176.
8. Caldwell, H. D., and J. Schachter. 1982. Antigenic analy-sis of the major outer membrane protein of Chiamydiaspp. Infect. Immun. 35:1024-1031.
9. Friis, R. R. 1972. Interaction of L cells and Chlamydiapsittaci: entry of the parasite and host responses to itsdevelopment. J. Bacteriol. 110:706-721.
10. Furness, G., D. M. Graham, and P. Reeve. 1960. Thetitration of trachoma and inclusion blennorrhoea virusesin cell cultures. J. Gen. Microbiol. 23:613-619.
11. Goding, S. W. 1976. Conjugation of antibodies with fluo-rochromes: modification to the standard methods. J.Immunol. Methods 13:215-226.
12. Graham, D. M. 1965. Growth and neutralization of thetrachoma agent in mouse lungs. Nature (London)207:1379-1380.
13. Graham, D. M., and J. E. Layton. 1971. The induction ofChlamydia group antibody in rabbit inoculated with tra-choma agents and demonstration of strain-specific neu-tralizing antibody in sera, p. 145-156. In R. L. Nichols(ed.), Trachoma and related disorders caused by chlamyd-ial agents. Excerpta Medica Foundation, New York.
14. Hatch, T. P. 1975. Competition between Chlamydia psit-taci and L cells for host isoleucine pools: a limiting factorin chlamydial multiplication. Infect. Immun. 12:211-220.
15. Hatch, T. P., D. W. Vance, Jr., and E. Al-Hossainy. 1981.Identification of a major envelope protein in Chlamydiaspp. J. Bacteriol. 146:426-429.
16. Hauser, W. E., Jr., and J. S. Remington. 1981. Effect ofmonoclonal antibodies on phagocytosis and killing ofToxoplasma gondii by normal macrophages. Infect. Im-mun. 32:637-640.
17. Hinrichs, D. S., and T. R. Jerrells. 1976. In vitro evalua-tion of immunity to Coxiella burnetii. J. Immunol.117:996-1003.
18. Horwitz, M. A., and S. C. Silverstein. 1981. Interaction ofthe Legionnaires' disease bacterium (Legionella pneumo-phila) with human phagocytes. I. L. pneumophila resistskilling by polymorphonuclear leukocytes, antibody, andcomplement. J. Exp. Med. 153:386-397.
19. Horwitz, M. A., and S. C. Silverstein. 1981. Interaction ofthe Legionnaires' disease bacterium (Legionella pneumo-phila) with human phagocytes. II. Antibody promotesbinding of L. pneumophila to monocytes but does notinhibit intracellular multiplication. J. Exp. Med. 153:398-406.
20. Howard, L. V. 1975. Neutralization of Chlamydia tracho-matis in cell culture. Infect. Immun. 11:698-703.
21. Jones, T. C. 1974. Macrophages and intracellular parasit-ism. RES J. Reticuloendothel. Soc. 15:439-450.
22. Jones, T. C., and J. G. Hirsch. 1972. The interactionbetween Toxoplasma gondii and mammalian cells. II. Theabsence of lysosomal fusion with phagocytic vacuolescontaining living parasites. J. Exp. Med. 136:1173-1194.
VOL. 38, 1982

754 CALDWELL AND PERRY
23. Jones, T. C., S. Yen, and J. G. Hirsch. 1972. The interac-tion between Toxoplasma gondii and mammalian cells. I.Mechanism of entry and intracellular fate of the parasite.J. Exp. Med. 136:1157-1172.
24. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folin phenolreagent. J. Biol. Chem. 193:265-275.
25. McLaughlin, C. A., and H. C. Pitot. 1976. The effect ofvarious treatments in vitro and in vivo on the binding of125I-labeled anti-rat serum albumin Fab' to rat tissuepolyribosomes. Biochemistry 15:3541-3550.
26. Moulder, J. W. 1966. The relation of the psittacois group(Chlamydiae) to bacteria and viruses. Annu. Rev. Micro-biol. 20:107-130.
27. Nichols, R. L., R. E. Oertley, C. E. Fraser, A. B. Mac-Donald, and D. E. McComb. 1973. Immunity to chlamydi-al infections of the eye. VI. Homologous neutralization ofTrachoma infectivity for the owl monkey conjunctivae byeye secretions from humans with trachoma. J. Infect. Dis.127:429-432.
28. Nicolson, G. L., and R. G. Painter. 1973. Anionic sites ofhuman erythrocyte membranes. II. Antispectrin-inducedtransmembrane aggregation of the binding sites for posi-tively changed colloidal particles. J. Cell Biol. 59:395-406.
INFECT. IMMUN.
29. Pain, R. H. 1963. The molecular weights of the peptidechains of y-globulin. Biochem. J. 88:234-239.
30. Porter, R. R. 1959. The hydrolysis of rabbit y-globulin andantibodies with crystalline papain. Biochem. J. 73:119-126.
31. Reeve, P., and D. M. Graham. 1962. A neutralization testfor trachoma and inclusion blennorrhoea viruses grown inHeLa cell cultures. J. Gen. Microbiol. 27:177-180.
32. Salari, S. H., and M. E. Ward. 1981. Polypeptide compo-sition of Chlamydia trachomatis. J. Gen. Microbiol.123:197-207.
33. Schachter, J., and H. D. Caldwell. 1980. Chlamydiae.Annu. Rev. Microbiol. 34:285-309.
34. Williams, D. W., J. Schachter, D. J. Drutz, and C. V.Sumaya. 1981. Pneumonia due to Chlamydia trachomatisin the immuno-compromised (nude) mouse. J. Infect. Dis.143:238-241.
35. Williams, D. W., J. Schachter, B. Grubbs, and C. V.Sumaya. 1982. The role of antibody in host defense againstthe agent of mouse pneumonitis. J. Infect. Dis. 145:200-205.
36. Wyrick, P. B., and E. A. Brownridge. 1978. Growth ofChlamydia psittaci in macrophages. Infect. Immun.19:1054-1060.





























