Embed Size (px)
Citation preview

Research in Veterinary Science 93 (2012) 836–842
Contents lists available at SciVerse ScienceDirect
Research in Veterinary Science
journal homepage: www.elsevier .com/locate / rvsc
Multilocus genotyping of Giardia duodenalis in lambs from Spain reveals ahigh heterogeneity
María Teresa Gómez-Muñoz a,b,⇑, Carmen Cámara-Badenes a, María del Carmen Martínez-Herrero a,María Auxiliadora Dea-Ayuela a, María Teresa Pérez-Gracia a, Salceda Fernández-Barredo c,Mónica Santín d, Ronald Fayer d
a Departamento de Producción Animal, Sanidad Animal y Ciencia y Tecnología de los Alimentos, Facultad de Veterinaria, Universidad CEU Cardenal-Herrera,Avenida Seminario S/N, 46113 Moncada, Valencia, Spainb Departamento de Sanidad Animal, Facultad de Veterinaria, Universidad Complutense de Madrid, Avenida Puerta de Hierro S/N, 28040 Madrid, Spainc CEDIVET (Centro de Diagnóstico Veterinario), C/Marqués de Montortal 37, bajo izquierda, 46019 Valencia, Spaind Environmental Microbial and Food Safety Laboratory, Animal and Natural Resources Institute, Agricultural Research Service, United States Department of Agriculture,Building 173, BARC-East, 10300 Baltimore Avenue, Beltsville, MD 20705, USA
a r t i c l e i n f o
Article history:Received 29 August 2011Accepted 16 December 2011
Keywords:GiardiaSheepAssemblage AZoonoticMultilocus genotyping
0034-5288/$ - see front matter � 2011 Elsevier Ltd. Adoi:10.1016/j.rvsc.2011.12.012
⇑ Corresponding author at: Departamento de SaVeterinaria, Universidad Complutense de Madrid, Av28040 Madrid, Spain.
E-mail address: [email protected]
a b s t r a c t
Fecal specimens from 120 lambs in Valencia (Spain) were analyzed for Giardia duodenalis by IFA andnested-PCR using the beta giardin (bg), glutamate dehydrogenase (gdh), triose phosphate isomerase (tpi)and small subunit ribosomal RNA (ssurRNA) genes. The highest prevalence was obtained using the ssurRNAgene (89.2%), whereas values from other techniques ranged from 64.1% to 69.2%. Sequences of thessurRNA showed a high proportion of assemblage A or mixed assemblage A/E samples (55.1% and25.2%, respectively). When the other 3 loci were analyzed, between 6.5% and 15.4% were found to beassemblage A or A/E, respectively. Nested PCR for the tpi gene was the most variable of the targetsemployed. Twelve new sequences of gdh and tpi for G. duodenalis from sheep were found. Multilocusgenotyping resulted in 63 patterns from the 71 samples sequenced at the four loci. This high variabilityamong isolates possibly reflects the high frequency of mixed infections.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
Giardia duodenalis is a ubiquitous parasite that infects humansand many other mammals. Because the parasite shows little mor-phologic variation, microscopically based investigation of out-breaks to determine zoonotic potential is impossible. Therefore,molecular techniques have been developed and used for identifica-tion. Seven assemblages (assemblages A–G) have been described inmammals mainly based on allozymic and genetic analysis (Moniset al., 2003). However, two new genotypes have been recently re-ported in mammals, genotype H and the quenda genotype (Adamset al., 2004; Lasek-Nesselquist et al., 2010; Ng et al., 2011).Most assemblages are found in closely related host species, forexample, assemblage C and D are found in canids, assemblage Fin felids, assemblage E in hoofed mammals and assemblage G inrodents). However, assemblages A and B, have been found in awide variety of mammals including humans.
ll rights reserved.
nidad Animal, Facultad deenida Puerta de Hierro S/N,
m.es (M.T. Gómez-Muñoz).
Three assemblages have been found in sheep, assemblage Ewhich predominates in all the genotyping studies, and assem-blages A and B which have been found mostly sporadically butare potentially zoonotic. A recent study showed that assemblageA was more prevalent in older lambs than in younger animals(Sweeny et al., 2011). In some studies sheep have not appearedto be an important potential source of infection for humans (Ryanet al., 2005; Santín et al., 2007; Yang et al., 2009; Robertson et al.,2010), whereas in other studies lambs were found be frequently in-fected with zoonotic assemblages. Although the percentage ofsheep infected with potentially zoonotic assemblages variesgreatly among studies, values approximating 40–50% have beenobtained (Geurden et al., 2008; Lebbad et al., 2010) depending onthe gene examined in molecular studies. Such data indicate thatsheep could be an important source of infection for humans(reviewed by Robertson, 2009).
Most molecular studies involving sheep have used a single locusto assign the isolates to assemblages (Aloisio et al., 2006; Santínet al., 2007; Giangaspero et al., 2005; Castro-Hermida et al., 2006,2010; Di Giovanni et al., 2006; Ryan et al., 2005; Gómez-Muñoz etal., 2009; Nolan et al., 2010). Only a few studies have used two ormore gene loci but these have used a small number of samples

M.T. Gómez-Muñoz et al. / Research in Veterinary Science 93 (2012) 836–842 837
(Castro-Hermida et al., 2007; Yang et al., 2009; Robertson et al.,2010; Lebbad et al., 2010, Geurden et al., 2008). The use of onlyone locus in animal studies has recently raised concern regardingassemblage assignment of isolates, but a similar situation existsin studies involving humans (Cacciò et al., 2008). Therefore, ithas been proposed that multilocus genotyping (MLG) using at leastfour genes could improve the assignment of each isolate to a spe-cific assemblage and thereby better clarify the epidemiology ofgiardiasis (Cacciò et al., 2008).
The present study was designed to compare the utility of thefour most frequently used loci for genotyping Giardia, beta giardin(bg), glutamate dehydrogenase (gdh), triose phosphate isomerase (tpi)and small subunit ribosomal RNA (ssurRNA) genes, on a large num-ber of samples from sheep to identify the most sensitive loci fordetecting the presence of Giardia while determining if multilocuspatterns provide helpful epidemiological information.
2. Material and methods
2.1. Faecal samples
Individual faeces were collected directly from the rectum of 1-to 3-month-old lambs on 4 farms near Valencia (Spain). On eachfarm, 30 lambs were randomly selected resulting in 120 samples.Three grams of faeces from each lamb were suspended in 30 mlof distilled water, filtered through a 50 lm pore size mesh sieveand centrifuged for 10 min at 1000�g. Between samples the sieveswere placed in a 5% aqueous solution of sodium hypochlorite for10 min. Each pellet was suspended in 7.5 ml of distilled water,mixed with 7.5 ml of cesium chloride (d = 1.4) in 15 ml tubes,and centrifuged at 300�g for 20 min. After centrifugation, 2 mlfrom the top of each tube was collected and washed twice in15 ml of distilled water by centrifugation at 2000�g for 10 min.The pellet was resuspended in 500 ll of distilled water and storedat 4 �C for DNA isolation and immunofluorescence analysis (IFA)for no longer than one week.
2.2. IFA
Twenty microliters of each pellet was analyzed using the MerI-Fluor kit (Meridian Diagnostics, Cincinatti, OH, USA) for detectionof Giardia cysts following the manufacturer’s instructions. Cystswere counted at 200 magnification and classified as follows: 0cysts per well (cpw): 0; 1–10 cpw: 1; 11–50 cpw: 2; 51–500cpw: 3; >500 cpw: 4.
2.3. DNA isolation and PCR
One hundred microliters of each pellet was used for DNAextraction. DNA was obtained using a commercial DNA extractionkit (DNeasy Tissue Kit, QIAGEN, Valencia, CA, USA) following man-ufacturer’s recommendations. The elution step was carried out in100 ll instead of 200 ll to render a higher DNA concentration.The DNA obtained was stored at �20 �C until use. Four distinct pro-tocols of nested PCR were used depending on the target to beamplified.
A fragment of 292 bp of the ssurRNA was obtained using theexternal primers Gia 2029 and Gia 2150c and internal primersRH4 and RH11 (Appelbee et al., 2003; Hopkins et al., 1997).
A semi-nested PCR was used to amplify a fragment of approxi-mately 432 bp of the gdh. The primers used were GDHeF, GDHiFand GDHiR (Read et al., 2004).
A fragment of approximately 500 bp of the tpi gene was ob-tained using the external primers AL3543 and AL3546, and internalprimers AL3544 and AL3545 (Sulaiman et al., 2003).
The amplification of the bg gene (511 bp) was achieved usingthe external primers G7 and G759 (Cacciò et al., 2002) and theinternal primers GBF and GBR (Lalle et al., 2005).
PCR reactions were performed in a Mastercycler thermocycler(Eppendorf, Germany) following the protocols described by theprevious authors. PCR reactions were visualized by electrophoresisin 1% agarose gels stained with ethidium bromide. For each set ofPCR reactions, positive and negative controls were included. DNAisolation, PCR preparation and addition of sample DNA were car-ried out in separated areas. Filter tips were used for DNA isolationand PCR preparation. PCR and sequencing were repeated in sam-ples positive only to one PCR target.
2.4. DNA sequencing and alignment
PCR products were cleaned using the MinElute PCR Purificationkit (QIAGEN, Valencia, CA, USA) and sequenced in both directionsin an ABI PRISM DNA 377 sequencer (PE, Applied Biosystems, Fos-ter City, CA, USA) using the same primers used in the second PCRs.
Sequences were assembled and corrected using the SeqMan™software (DNAstar Inc, Madison, WI, USA). Nucleotide sequencesobtained in this study were aligned by Clustal W using the Meg-Align™ module (DNAstar Inc, Madison, WI, USA). For each locus,reference sequences published in GenBank were also aligned withthe sequences obtained in this study: AY297957 (Assemblage E,Bos taurus), EU562195 (Assemblage A, Vicugna pacos), DQ385547(Assemblage B, Canis latrans) at the ssurRNA; sequencesAY072729 (Assemblage E-1, Sus scrofa domesticus), AY545650(Assemblage E-2, Bos taurus), AY653159 (Assemblage E-3, Bos tau-rus), AY655702 (Assemblage AI, Bos taurus), AY072723 (Assem-blage AII, Homo sapiens), AY072724 (Assemblage AIII, Homosapiens) at the bg locus; sequences EU769231 (Assemblage E,sheep, Ovis aries), EU769232 (Assemblage E, Dama dama),GQ168944 (genotype AII, Homo sapiens), EF685701 (AssemblageAI, Homo sapiens) at the gdh locus and sequences EU189331(Assemblage E, Capra hircus hircus), AY228645 (Assemblage E,genotype S-1, Bos taurus), AY228646 (Assemblage E, genotype S-2, Bos taurus), EU518565 (Assemblage A genotype 1, Larus argenta-rus), EU518562 (Assemblage A, genotype 2, Larus argentarus),DQ650648 (Assemblage A, Cervus dama) at the tpi locus.
2.5. GenBank submission
The sequences obtained in this study were subjected to BLASTsearches in GenBank. New sequences and sequences previously de-scribed in a different host and/or country were deposited in Gen-Bank with accession numbers JF792400-JF792423.
2.6. Statistic analysis
A Spearman coefficient was calculated to measure the correla-tion between the number of cysts per well and the percentage ofpositive samples found in each analysis (microscopy, IFD, nestedPCR of four targets). Agreement between diagnostic techniqueswas obtained by kappa coefficient determination. Kappa valueswere interpreted according to Altman (1991). SPSS 15.0 softwarepackage was used for statistical analysis.
3. Results
3.1. Comparison of diagnostic methods
The number of samples detected by IFA as well as each of thefour nested-PCR techniques used in this study is shown in Table 1.Above all, the nested-PCR for the ssurRNA was the most sensitive of

Table 1Number of positive (P) and negative (N) samples detected using different diagnostic methods (nested-PCR for the small subunit ribosomal RNA, beta giardin, triose phosphateisomerase and glutamate dehydrogenase genes). Number of cysts detected by IFA is used as a reference. Correlation between the number of cysts detected by IFA and the number ofsamples detected by each method was calculated by Spearman coefficient.
IFA* Nested-PCR ssurRNA Nested-PCR bg Nested-PCR tpi Nested-PCR gdh
N P N P N P N P
a 13 30 34 9 36 7 31 12b 0 17 9 8 7 10 6 11c 0 8 0 8 0 8 0 8d 0 19 0 19 0 19 0 19e 0 33 0 33 0 33 0 33
TOTAL 13 107 43 77 43 77 37 83Spearman coefficient 0.403 0.722 0.754 0.668
* a: No cyst in 20 ll; b: 1–10 cyst in 20 ll; c: 11–50 cysts in 20 ll; d: 51–500 cysts in 20 ll; e: >500 cysts in 20 ll.
838 M.T. Gómez-Muñoz et al. / Research in Veterinary Science 93 (2012) 836–842
the methods employed. Of the 120 samples examined 107 werepositive using this technique (89.2%). Thirty samples negative byIFA were positive by nested-PCR at the ssurRNA locus while only7–12 samples negative by IFA were positive using nested-PCR forthe gdh, tpi or bg loci. A good correlation was observed betweenthe number of cysts detected by IFA and the nested-PCR of gdh(0.668), tpi (0.754) and bg (0.722). However, the correlation be-tween IFA and nested-PCR of the ssurRNA was poor (0.403).
The agreement between the employed techniques was ana-lysed. The best coefficient was found between the nested-PCR forbg and tpi (kappa = 0.855, very good agreement). Good agreementwas observed between nested-PCRs for bg and gdh (kappa = 0.701)and nested-PCRs for tpi and gdh (kappa = 0.796). Also, the kappacoefficient calculated between the IFA and each of the nested-PCRsfor bg, tpi and gdh displayed good agreement (0.663–0.746). Only afair or moderate correlation was obtained between the nested-PCRfor the ssurRNA and the other techniques (IFA and nested-PCR fortpi, gdh and bg, kappa values between 0.357 and 0.428). Theselow values were due to the large amount of samples detected so-lely by the nested-PCR for the ssurRNA.
Specificity was determined by sequencing in both directions allthe positive samples to each of the nested-PCRs. In all cases thenucleotide sequences corresponded to G. duodenalis.
3.2. Sequence analysis
A total of 344 sequences were obtained and analyzed in thisstudy, 107 of the ssurRNA gene, 77 of the bg gene, 77 of the tpi geneand 83 of the gdh gene. Of the 120 individual faecal samples 71were successfully analyzed at the four genes. Only one to threegene nucleotide sequences were obtained from the rest of the po-sitive samples.
Tables 2 and 3 show the intra-sub-assemblage substitution ofgenotypes E and A at the bg, gdh and tpi genes found in this study.Only sequences that were identified as non-previously described ordescribed in different hosts are shown.
3.2.1. ssurRNA sequencesThe most common sequence obtained in this study corre-
sponded to Assemblage A (n = 59, 55.1%). Twenty-seven sequenceswere mixed A/E assemblages (n = 27, 25.2%) and all other se-quences were assemblage E (n = 21, 19.6%).
The values were variable depending on the farm. For farm I, 2(6.9%) and 9 (31%) of 29 positive samples were Assemblage E ormixed A/E, respectively. For farm II, 5 (16.7%) and 3 (10%) of 30 po-sitive samples were assemblages E and A/E, respectively. For farmIII, 5 (17.2%) and 15 (51.7) of 29 positive samples were assem-blages E and A/E, respectively. For farm IV, 9 (47.4%) and 0 (0%)of 19 positive samples were E and A/E, respectively. Assemblage
A was found in 18 (62.1%), 22 (73.3%), 9 (31%) and 10 (52.6%) fromfarms I, II, III and IV respectively.
3.2.2. bg SequencesOf the 77 sequences obtained for the bg gene, 51 were Assem-
blage E. Two more sequences were Assemblage A and the rest ofthe sequences were identified as mixed assemblages: 1 as a mixedA/A genotype, 2 were mixed A/E and 21 were mixed E/E. In total, 14different sequences were identified in this study, 5 of them mixed(Table 4).
The most common sequence (46/77) was identical to a se-quence previously found in sheep (EU726980; Gómez-Muñozet al., 2009) and identical to other bg sequences also obtained fromsheep (DQ116604 and 6 more, Di Giovanni et al., 2006) and fromgoats (EU189361, Ruiz et al., 2008). The second most common se-quence (10/77) was a mix of two sequences, EU726980 andEU726984 (Gómez-Muñoz et al., 2009).
One of the sequences from Assemblage A was identical to se-quence AY655702 (Assemblage AI from cattle, Trout et al., 2004)and to sequence EU726988 (Assemblage AI from sheep, Gómez-Muñoz et al., 2009). The other sequence from Assemblage Ashowed 100% homology to sequence DQ904426 (Assemblage Afrom a fox, Hamnes et al., 2007). A mixed sequence of sequencesDQ904426 and AY072723 (Assemblage AII from cattle, Cacciòet al., 2002) was also obtained from one animal.
3.2.3. gdh SequencesOf the 83 sequences obtained at the gdh gene 65 were Assem-
blage E and 5 were Assemblage A. Four sequences were mixedAssemblages A/E and 9 sequences were a mixture of AssemblageE sequences (Table 4). In total, 18 different sequences were ob-served at the gdh locus. Eight new sequences were described fromsheep in Spain for the first time and were deposited at GenBank(FJ792414-FJ792422) (Tables 2 and 3). Six of those have never beenreported previously.
The most common sequence, (FJ792415) (34/83),was previouslyreported from sheep (GQ337965, Robertson et al., 2010) and frompigs (DQ182610, Langkjaer et al., 2007). The second more frequentsequence obtained in this study (FJ792422) (20/83), showed 100%homology to sequences DQ182605 obtained from cattle (Langkjaeret al., 2007) and AY826198 obtained from goat (van der Giessenet al., 2006).
Sequences of Assemblage A were identical to sequenceEF685701, among others, obtained from human (Assemblage AI,Lasek-Nesselquist et al., 2009) (see Table 3).
3.2.4. tpi SequencesThe tpi gene was the most variable of the targets used in this
study. A total of 28 different sequences were obtained (Table 4).Sixty-five of the 77 sequences belonged to Assemblage E (31 of
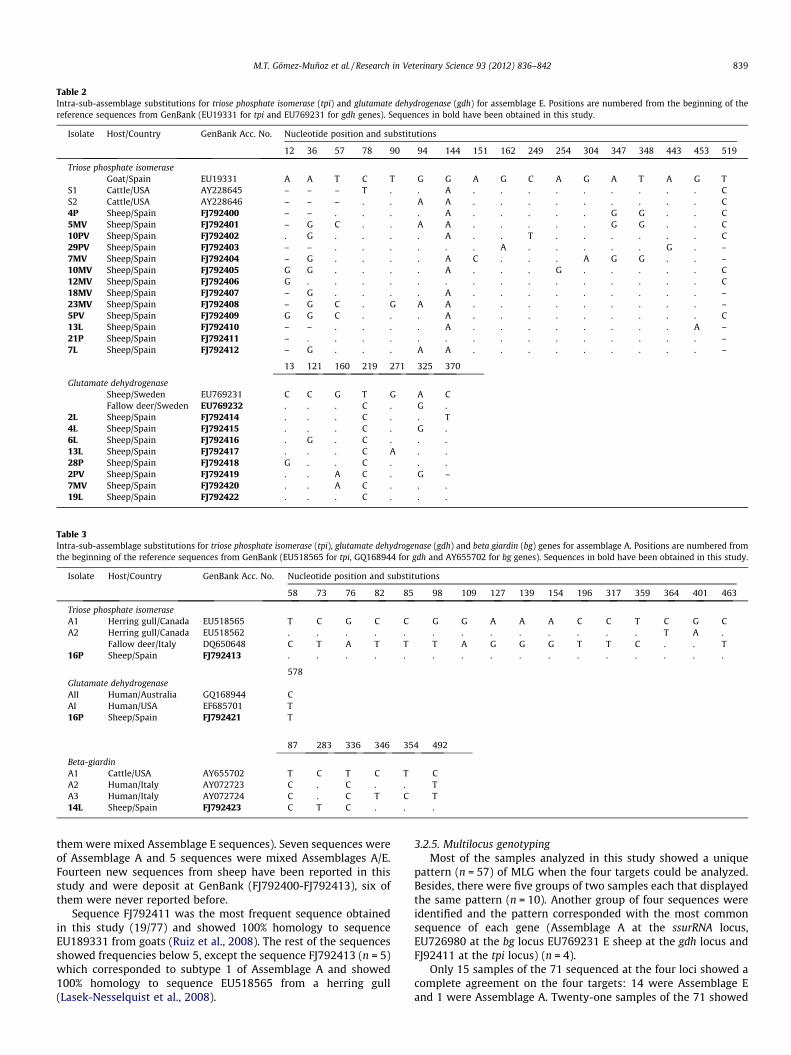
Table 2Intra-sub-assemblage substitutions for triose phosphate isomerase (tpi) and glutamate dehydrogenase (gdh) for assemblage E. Positions are numbered from the beginning of thereference sequences from GenBank (EU19331 for tpi and EU769231 for gdh genes). Sequences in bold have been obtained in this study.
Isolate Host/Country GenBank Acc. No. Nucleotide position and substitutions
12 36 57 78 90 94 144 151 162 249 254 304 347 348 443 453 519
Triose phosphate isomeraseGoat/Spain EU19331 A A T C T G G A G C A G A T A G T
S1 Cattle/USA AY228645 – – – T . . A . . . . . . . . . CS2 Cattle/USA AY228646 – – – . . A A . . . . . . . . . C4P Sheep/Spain FJ792400 – – . . . . A . . . . . G G . . C5MV Sheep/Spain FJ792401 – G C . . A A . . . . . G G . . C10PV Sheep/Spain FJ792402 . G . . . . A . . T . . . . . . C29PV Sheep/Spain FJ792403 – – . . . . . . A . . . . . G . –7MV Sheep/Spain FJ792404 – G . . . . A C . . . A G G . . –10MV Sheep/Spain FJ792405 G G . . . . A . . . G . . . . . C12MV Sheep/Spain FJ792406 G . . . . . . . . . . . . . . . C18MV Sheep/Spain FJ792407 – G . . . . A . . . . . . . . . –23MV Sheep/Spain FJ792408 – G C . G A A . . . . . . . . . –5PV Sheep/Spain FJ792409 G G C . . . A . . . . . . . . . C13L Sheep/Spain FJ792410 – – . . . . A . . . . . . . . A –21P Sheep/Spain FJ792411 – . . . . . . . . . . . . . . . –7L Sheep/Spain FJ792412 – G . . . A A . . . . . . . . . –
13 121 160 219 271 325 370
Glutamate dehydrogenaseSheep/Sweden EU769231 C C G T G A CFallow deer/Sweden EU769232 . . . C . G .
2L Sheep/Spain FJ792414 . . . C . . T4L Sheep/Spain FJ792415 . . . C . G .6L Sheep/Spain FJ792416 . G . C . . .13L Sheep/Spain FJ792417 . . . C A . .28P Sheep/Spain FJ792418 G . . C . . .2PV Sheep/Spain FJ792419 . . A C . G –7MV Sheep/Spain FJ792420 . . A C . . .19L Sheep/Spain FJ792422 . . . C . . .
Table 3Intra-sub-assemblage substitutions for triose phosphate isomerase (tpi), glutamate dehydrogenase (gdh) and beta giardin (bg) genes for assemblage A. Positions are numbered fromthe beginning of the reference sequences from GenBank (EU518565 for tpi, GQ168944 for gdh and AY655702 for bg genes). Sequences in bold have been obtained in this study.
Isolate Host/Country GenBank Acc. No. Nucleotide position and substitutions
58 73 76 82 85 98 109 127 139 154 196 317 359 364 401 463
Triose phosphate isomeraseA1 Herring gull/Canada EU518565 T C G C C G G A A A C C T C G CA2 Herring gull/Canada EU518562 . . . . . . . . . . . . . T A .
Fallow deer/Italy DQ650648 C T A T T T A G G G T T C . . T16P Sheep/Spain FJ792413 . . . . . . . . . . . . . . . .
578Glutamate dehydrogenaseAII Human/Australia GQ168944 CAI Human/USA EF685701 T16P Sheep/Spain FJ792421 T
87 283 336 346 354 492
Beta-giardinA1 Cattle/USA AY655702 T C T C T CA2 Human/Italy AY072723 C . C . . TA3 Human/Italy AY072724 C . C T C T14L Sheep/Spain FJ792423 C T C . . .
M.T. Gómez-Muñoz et al. / Research in Veterinary Science 93 (2012) 836–842 839
them were mixed Assemblage E sequences). Seven sequences wereof Assemblage A and 5 sequences were mixed Assemblages A/E.Fourteen new sequences from sheep have been reported in thisstudy and were deposit at GenBank (FJ792400-FJ792413), six ofthem were never reported before.
Sequence FJ792411 was the most frequent sequence obtainedin this study (19/77) and showed 100% homology to sequenceEU189331 from goats (Ruiz et al., 2008). The rest of the sequencesshowed frequencies below 5, except the sequence FJ792413 (n = 5)which corresponded to subtype 1 of Assemblage A and showed100% homology to sequence EU518565 from a herring gull(Lasek-Nesselquist et al., 2008).
3.2.5. Multilocus genotypingMost of the samples analyzed in this study showed a unique
pattern (n = 57) of MLG when the four targets could be analyzed.Besides, there were five groups of two samples each that displayedthe same pattern (n = 10). Another group of four sequences wereidentified and the pattern corresponded with the most commonsequence of each gene (Assemblage A at the ssurRNA locus,EU726980 at the bg locus EU769231 E sheep at the gdh locus andFJ92411 at the tpi locus) (n = 4).
Only 15 samples of the 71 sequenced at the four loci showed acomplete agreement on the four targets: 14 were Assemblage Eand 1 were Assemblage A. Twenty-one samples of the 71 showed

Table 4Sequence analysis of the nested-PCRs positive samples obtained in this study using bg, gdh and tpi genes. Sequences in bold have been obtained in this study.
bg Sequences n gdh Sequences n tpi Sequences n
AY655702 1 EF685701 5 FJ792400 1FJ792423 1 EF685701/EU769231 3 FJ792400/FJ792423 1FJ792423/AY072723 1 EU769231 20 FJ792400/FJ792407 2EU726980 46 EU769231/FJ792415 1 FJ792400/FJ792411 3EU726980/AY072723 1 EU769231/FJ792417 1 FJ792401 2EU726980/AY545650 1 FJ792414 2 FJ792401/FJ792423 1EU726980/EU726984 10 FJ792415 34 FJ792402 2EU726980/EU726985 4 FJ792415/FJ792416/FJ792417 2 FJ792402/FJ792411 1EU726980/EU726986 1 FJ792415/FJ792418 1 FJ792403 1EU726980/EU726987 3 FJ792415/FJ792419 1 FJ792404 1EU726984 3 FJ792416 1 FJ792404/FJ792407 5EU726984/AY655702 1 FJ792416/FJ792417 1 FJ792404/FJ792411 1EU726985 2 FJ792417 2 FJ792404/FJ792423 1EU726985/EU726987 2 FJ792417/EF685701 1 FJ792405 1
FJ792418 1 FJ792406 3FJ792419 3 FJ792407 1FJ792420 2 FJ792407/FJ792408 3FJ792420/EU769231 2 FJ792407/FJ792411 3
FJ792407/FJ792413 4FJ792408 1FJ792408/FJ792411 2FJ792409 1FJ792410 1FJ792410/FJ792411 3FJ792411 19FJ792411/FJ792423 5FJ792413 7FJ792423 1
Table 5Genotype distribution of ssurRNA, bg, gdh and tpi sequences. na: Sequences were notamplified.
ssurRNA bg gdh tpi No. sequences
A na na na 17A A A 1
A/E A/E 1na E 1
A/E E E 1E A na 1
A/E A 1E A 2E E 21na A 1na na 2
na A E 1A na 1A A 1A/E na 2E E 1E na 4
A/E na na na 1E E E 24
E A/E 2
E na na na 1E E E 14
na na 1E A 1E A/E 2
na E E 1A/E E E 1
TOTAL 107
840 M.T. Gómez-Muñoz et al. / Research in Veterinary Science 93 (2012) 836–842
Assemblage A at the ssurRNA whereas the other 3 loci revealedAssemblage E. Twenty-four samples showed Assemblage E at threeloci and Assemblage A/E only at the ssurRNA. The rest of the sam-ples (n = 47) were negative to some of the targets or showed amixed pattern with a combination of assemblages, suggesting ahigh level of mixed assemblage infection in the sampled lambs(Table 5).
4. Discussion
The sensitivity of the loci used for identifying assemblages of G.duodenalis isolates varies. Also, some loci represent more con-served genes, such as ssurRNA, whereas other loci such bg, gdhand tpi are much less conserved (reviewed in Cacciò and Ryan,2008). In this study the ssurRNA locus was the most sensitive,and therefore well suited for prevalence studies. This finding is inagreement with other studies (Cacciò and Ryan, 2008). The mainreason is most likely the multicopy nature of this gene, withapproximately 60 copies of the rDNA tandem repeat detected inthe parasite genome (Adam, 2000). The variability found in theother three loci used to genotype the parasite (bg, gdh, tpi) mayproduce excessive mismatches in the binding region resulting inlower sensitivity of the PCRs (Cacciò and Ryan, 2008).
In this study, we have seen 89.2% of the animals infected withthe parasite, which is clearly higher than the data obtained by oth-ers or in our previous study (42%, Gómez-Muñoz et al., 2009). Thereasons to explain this fact might be several. First, this value hasbeen obtained with the most sensitive of the techniques (nested-PCR of the ssurRNA gene), whereas in other surveys different geneswere used (bg in Castro-Hermida et al., 2006 and 2010, bg and gdhin Castro-Hermida et al., 2007, bg in Gómez-Muñoz et al., 2009, bgand gdh in Robertson et al., 2010, bg and tpi in Geurden et al.,2008). However, other studies conducted in sheep have also usedthe same PCR technique (Ryan et al., 2005; Yang et al., 2009; Santínet al., 2007). In some of these studies the amount of faecal materialused from each lamb was lower (Yang et al., 2009) probably due tothe scarce material that can be obtained from very young lambs.Also, the age of the animals is an important fact: pre-weaned lambstend to shed a smaller amount of cysts than lambs between 1–3 months of age (Taylor et al., 1993). According to Sweeny et al.(2011), older lambs tend to have a higher proportion of Assem-blage A than younger lambs. In other studies, probably adult sheep,which tend to have much lower infection rates and number of shedcysts (Buret et al., 1990), were the source of samples (Ryan et al.,2005; Santín et al., 2007; Castro-Hermida et al., 2007; Lebbadet al., 2010). In some studies only samples with a relatively high

M.T. Gómez-Muñoz et al. / Research in Veterinary Science 93 (2012) 836–842 841
amount of cysts were genotyped (Robertson et al., 2010). Finally,the variability in percentage of infection found among farms(Gómez-Muñoz et al., 2009; Robertson et al., 2010), can explainthe difference in prevalence values obtained in this and other stud-ies carried out in only 1 farm (Santín et al., 2007).
The assignment to Assemblage A or E was done according to thesequences obtained at the ssurRNA locus. In total, eighty-six sam-ples showed Assemblage A (55.1%) or mixed Assemblage A/E(25.2%) and only 20% showed pure Assemblage E. In other studiesthe percentage of assemblage A or mixed A/E was very low (e.g.,3.1% in Santín et al., 2007) or lower (17% in Sprong et al., 2009,24% in Ryan et al., 2005, 30% in Yang et al., 2009, 38.5% in Lebbadet al., 2010, 50% in Geurden et al., 2008). Also, the percentage ofsamples belonging to each assemblage is variable depending onthe farm. In our study, the 4 farms showed values between 6.9%and 47.4% of Assemblage E sequences, between 0% and 31% ofmixed Assemblage A/E, and between 31 and 73.3% of AssemblageA.
Since ssurRNA gene is strongly conserved among Giardia assem-blages, our sequences for both assemblages differ in only one baseat the amplified region and for that reason we think it is necessaryto take into consideration other genes. When other loci were inves-tigated, the percentage of sequences belonging to genotype A ormixed A/E was much lower (6.5% at bg, 10.8% at the gdh and15.4% at the tpi), although the values vary depending on the targetused. Many of the samples (n = 42) classified as assemblage A ormixed A/E were classified as 0 (no cysts) or 1 (1–10 cysts) byIFA, whereas only 5 samples classified as 0 or 1 at IFA were classi-fied as assemblage E by ssurRNA sequence. Only 11 of these se-quences could be genotyped at the four loci and we found thesame genotype at the four loci only in five of them. The restshowed disagreement in some of the loci. We have to consider aswell that some of the studies investigate the assemblage of Giardiaonly when the number of cysts detected by IFA is sufficient to ob-tain a sequence (Robertson et al., 2010). According to this criterion,many of the samples we had considered as positive only by nestedPCR of the ssurRNA could have been classified as negative.
The most common potentially zoonotic sub-genotype found inthis study was AI. Sprong et al. (2009) reported 78% of the assem-blage A sequences obtained from sheep and goats as genotype AI.This sub-genotype was also frequently found in other studies car-ried out in sheep (Giangaspero et al., 2005; Lebbad et al., 2010;Gómez-Muñoz et al., 2009).
We have found 14 different sequences at the bg locus, 18 at thegdh and 28 at the tpi. The tpi gene showed the greatest variability,followed by gdh and then by bg. This is in agreement with otherauthors (Wielinga and Thompson, 2007; Cacciò and Ryan, 2008).For this reason, we also think that tpi gene is the target to be inves-tigated for strain identification, and it would also be very useful inoutbreak investigations.
Double peaks in the chromatograms might be explained bymixed infections and/or by allelic sequence heterozygosity (re-viewed in Cacciò and Ryan, 2008). In the present study we havefound a considerable amount of mixed templates in the sequencesobtained, although the percentage of mixed sequences varies con-siderably among the genes employed. Forty-four percent of the tpisequences showed double peaks in the chromatograms, whereasonly 15.7% of the gdh sequences were mixed templates. Bg geneshowed an intermediate value of mixed templates (31.1%). Lebbadet al. (2010) and Geurden et al. (2008) also found a high percentageof double peaks in the chromatograms in assemblage E. In our case,and taken into account the high endemicity found in the sampledfarms, we think that most of the mixed templates could be attrib-uted to real mixed infections.
In the present study, we have found 63 different patterns ofMLG in 71 samples analyzed at the four loci. Cacciò et al. (2008)
proposed the use of a nomenclature for the MLG model. Thisnomenclature is of usefulness in humans, but taken into accountthe great variability found in this and other studies carried out insheep, the scenario turns out to be more complicated in this animalspecies (Lebbad et al., 2010).
We have found many samples classified as one assemblage withone target and to another (or mixed) in a different target. Whendiscrepancies like this occur, assemblage A is present in most cases(see Sprong et al., 2009). The same can be found in our study,where only Assemblages A and E have been obtained.
This is the largest study carried out in sheep using four of thegenes usually employed for genotyping of G. duodenalis. Our studysuggests that Giardia infections in sheep are highly common andprobably mixed infections are the rule.
Acknowledgements
This work has been supported by a grant from the Spanish Min-isterio de Ciencia y Tecnología (AGL2007/62435GAN).
References
Adam, R.D., 2000. The Giardia lamblia genome. International Journal for Parasitology30, 475–484.
Adams, P.J., Monis, P.T., Elliot, A.D., Thompson, R.C., 2004. Cyst morphology andsequence analysis of the small subunit rDNA and ef1 alpha identifies a nobelGiardia genotype in a quenda (Isoodon obesulus) from western Ausralia.Infection, Genetics and Evolution 4, 365–370.
Aloisio, F., Filippini, G., Antenucci, P., Lepra, E., Pezzotti, G., Cacciò, S.M., Pozio, E.,2006. Severe weight loss in lambs infected with Giardia duodenalis assemblageB. Veterinary Parasitology 142, 154–158.
Altman, D.G., 1991. Practical Statistics for Medical Research. Chapman and Hall,London.
Appelbee, A.J., Frederick, L.M., Heitman, T.L., Olson, M.E., 2003. Prevalence andgenotyping of Giardia duodenalis from beef calves in Alberta, Canada. VeterinaryParasitology 112, 289–294.
Buret, A., den Hollander, N., Wallis, P.M., Befus, D., Olson, M.E., 1990. Zoonoticpotential in giardiasis in domestic ruminants. Journal of Infectious Diseases 162,231–237.
Cacciò, S.M., Ryan, U., 2008. Molecular epidemiology of giardiasis. Molecular andBiochemical Parasitology 160, 75–80.
Cacciò, S.M., de Giacomo, M., Pozio, E., 2002. Sequence analysis of the b-giardin geneand development of a PCR-RFLP assay to genotype Giardia duodenalis cysts fromhuman fecal samples. International Journal for Parasitology 32, 1023–1030.
Cacciò, S.M., Beck, R., Lalle, M., Marinculic, A., Pozio, E., 2008. Multilocus genotypingof Giardia duodenalis reveals striking differences between assemblages A and B.International Journal for Parasitology 38, 1523–1531.
Castro-Hermida, J.A., Almeida, A., González-Warleta, M., Correia da Costa, J.M.,Mezo, M., 2006. Prevalence and preliminary genetic analysis of Giardia isolatedfrom adult sheep in Galicia (northwest Spain). The Journal of EukaryoticMicrobiology 53, S172–173.
Castro-Hermida, J.A., Almeida, A., González-Warleta, M., Correia da Costa, J.M.,Rumbo-Lorenzo, C., Mezo, M., 2007. Occurrence of Cryptosporidium parvum andGiardia duodenalis in healthy adult domestic ruminants. Parasitology Research101, 1443–1448.
Castro-Hermida, J.A., García-Presedo, I., Almeida, A., González-Warleta, M., Correiada Costa, J.M., Mezo, M., 2010. Cryptosporidium spp. and Giardia duodenalis intwo areas of Galicia (NW Spain). Science of the Total Environment 409, 2451–2459.
Di Giovanni, G.D., Betancourt, W.Q., Hernandez, J., Assadian, N.W., Flores Marquez,J.P., Lopez, E.J., 2006. Investigation of potential zooanthroponotic transmissionof cryptosporidiosis and giardiasis through agricultural use of reclaimedwastewater. International Journal of Environmental Health Research 16, 405–418.
Geurden, T., Thomas, P., Casaert, S., Vercruysse, J., Claerebout, E., 2008. Prevalenceand molecular characterization of Cryptosporidium and Giardia in lambs andgoat kids in Belgium. Veterinary Parasitology 155, 142–155.
Giangaspero, A., Paoletti, B., Lorio, R., Traversa, D., 2005. Prevalence and molecularcharacterization of Giardia duodenalis from sheep in central Italy. ParasitologyResearch 96, 32–37.
Gómez-Muñoz, M.T., Navarro, C., Garijo-Toledo, M.M., Dea-Ayuela, M.A., Fernández-Barredo, S., Pérez-Gracia, M.T., Domínguez-Márquez, M.V., Borrás, R., 2009.Occurrence and genotypes of Giardia isolated from lambs in Spain. ParasitologyInternational 58, 297–299.
Hamnes, I.S., Gjerde, B.K., Forberg, T., Robertson, L.J., 2007. Occurence of Giardia andCryptosporidium in Norwegian red foxes (Vulpes vulpes). Veterinary Parasitology143, 347–353.
Hopkins, R.M., Meloni, B.P., Groth, D.M., Wetheral, J.D., Reynoldson, J.A., Thompson,R.C., 1997. Ribosomal RNA sequencing reveals differences between the

842 M.T. Gómez-Muñoz et al. / Research in Veterinary Science 93 (2012) 836–842
genotypes of Giardia isolates recovered from humans and dogs living in thesame locality. The Journal of Parasitology 83, 44–51.
Lalle, M., Pozio, E., Capelli, G., Bruschi, F., Crotti, D., Cacció, S.M., 2005. Geneticheterogeneity at the b-giardin locus among human and animal isolates ofGiardia duodenalis and identification of potentially zoonotic subgenotypes.International Journal for Parasitology 35, 207–213.
Langkjaer, R.B., Vigre, H., Enemark, H.L., Maddox-Hyttel, C., 2007. Molecular andphylogenetic characterization of Cryptosporidium and Giardia from pigs andcattle in Denmark. Parasitology 134, 339–350.
Lasek-Nesselquist, E., Bogomolni, A.L., Gast, R.J., Welch, D.M., Ellis, J.C., Sogin, M.L.,Moore, M.J., 2008. Molecular characterization of Giardia intestinalis haplotypesin marine animals: variation and zoonotic potential. Diseases of AquaticOrganisms 81, 39–51.
Lasek-Nesselquist, E., Welch, D.M., Thompson, R.C.A., Steuart, R.F., Sogin, M.L., 2009.Genetic exchange within and between Assemblages of Giardia duodenalis.Journal Eukaryotic Microbiology 56, 504–518.
Lasek-Nesselquist, E., Welch, D.M., Sogin, M.L., 2010. The identification of a newGiardia duodenalis assemblage in marine vertebrates and a preliminary analysisof G. duodenalis population biology in marine systems. International Journal forParasitology 40, 1063–1074.
Lebbad, M., Mattsson, J.G., Christensson, B., Ljungström, B., Backhans, A., Anderson,J.O., Svärd, S., 2010. From mouse to moose: Multilocus genotyping of Giardiaisolates from various animal species. Veterinary Parasitology 168, 231–239.
Monis, P.T., Andrews, R.H., Mayrhofer, G., Ey, P.L., 2003. Genetic diversity within themorphological species Giardia intestinalis and its relationship to host origin.Infection, Genetics and Evolution 3, 29–38.
Ng, J., Yang, R., McCarthy, S., Gordon, C., Hijawi, N., Ryan, U., 2011. Molecularcharacterization of Cryptosporidium and Giardia in pre-weaned calves inWestern Australia and New South Wales. Veterinary Parasitology 176, 145–150.
Nolan, M.J., Jex, A.R., Pangasa, A., Young, N.D., Campbell, A.J., Stevens, M., Gasser,R.B., 2010. Analysis of nucleotide variation within the triose-phosphateisomerase gene of Giardia duodenalis from sheep and its zoonoticimplications. Electrophoresis 31, 287–298.
Read, C.M., Monis, P.T., Thompson, R.C.A., 2004. Discrimination of all genotypes ofGiardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP.Infection, Genetics and Evolution 4, 125–130.
Robertson, L.J., 2009. Giardia and Cryptosporidium infection in sheep and goats: areview of the potential for transmission to humans via environmentalcontamination. Epidemiology and Infection 137, 913–921.
Robertson, L.J., Gjerde, B.K., Furuseth Hansen, E., 2010. The zoonotic potential ofGiardia and Cryptosporidium in Norwegian sheep: a longitudinal investigation of6 flocks of lambs. Veterinary Parasitology 171, 140–145.
Ruiz, A., Foronda, P., González, J.F., Guedes, A., Abreu-Acosta, N., Molina, J.M.,Valladares, B., 2008. Ocurrence and genetic characterization of Giardiaduodenalis in goat kids from the Canary Islands, Spain. Veterinary Parasitology154, 137–141.
Ryan, U.M., Bath, C., Robertson, I., Read, C., Elliot, A., McInnes, L., Traub, R., Besier, B.,2005. Sheep may not be an important zoonotic reservoir for Cryptosporidiumand Giardia parasites. Applied and Environmental Microbiology 71, 4992–4997.
Santín, M., Trout, J.M., Fayer, R., 2007. Prevalence and molecular characterization ofCryptosporidium and Giardia species and genotypes in sheep in Maryland.Veterinary Parasitology 146, 17–24.
Sprong, H., Cacciò, S.M., Van der Giesen, J.W.B., 2009. Identification of zoonoticgenotypes of Giardia duodenalis. PLOS Neglected Tropical Disease 3, e558 (1–12).
Sulaiman, I.M., Fayer, R., Bern, C., Gilman, R.H., Trout, J.M., Schantz, P.M., Das, P., Lal,A.A., Xiao, L., 2003. Triosephosphate isomerase gene characterization andpotential zoonotic transmission of Giardia duodenalis. Emerging InfectiousDiseases 9, 1444–1452.
Sweeny, J.P., Ryan, U.M., Robertson, I.D., Yang, R., Bell, K., Jacobson, C., 2011.Longitudinal investigation of protozoan parasites in meat lamb farms insouthern Western Australia. Preventive Veterinary Medicine 101, 192–203.
Taylor, M.A., Catchpole, J., Marshall, R.N., Green, J., 1993. Giardiasis in lambs atpasture. The Veterinary Record 133, 131–133.
Trout, J.M., Santin, M., Greiner, E., Fayer, R., 2004. Prevalence of Giardia duodenalisgenotypes in pre-weaned dairy calves. Veterinary Parasitology 124, 179–186.
Van der Giessen, J.W., de Vries, A., Roos, M., Wielinga, P., Kortbeek, L.M., Mank, T.G.,2006. Genotyping of Giardia in dutch patiens and animals: a phylogeneticanalysis of human and animal isolates. International Journal for Parasitology 36,849–858.
Wielinga, C.M., Thompson, R.C.A., 2007. Comparative evaluation of Giardiaduodenalis sequence data. Parasitology 134, 1795–1821.
Yang, R., Jacobson, C., Gordon, C., Ryan, U., 2009. Prevalence and molecularcharacterization of Cryptosporidium and Giardia species in pre-weaned sheep inAustralia. Veterinary Parasitology 161, 19–24.





























