Embed Size (px)
Citation preview

411
Arch. Biol. Sci., Belgrade, 67(2), 411-426, 2015 DOI:10.2298/ABS140417007K
MORPHOLOGICAL IDENTIFICATION OF VESICULAR-ARBUSCULAR MYCORRHIZA ON BULBOUS PLANTS (TAURUS MOUNTAIN IN TURKEY)
Emel Karaarslan1,*, Refik Uyanöz1 and Süleyman Doğu2
1 Selcuk University, Agriculture Faculty, Soil Science and Plant Nutrition Department, Kampus, Konya, Turkey
3 Necmettin Erbakan University, Ahmet Kelesoglu Faculty of Education, Science Teacher Training Department, Meram, Konya, Turkey
*Corresponding author: [email protected]
Abstract: This study was conducted to investigate the morphological identification of vesicular-arbuscular mycorrhiza (VAM) on bulbous plants in the Taurus Mountains in Turkey. Thirteen soil samples and bulbous roots were taken from the rhizosphere of bulbous plants. The soils were analyzed for the number of VAM spores and chemical and physical properties. In addition, the roots were examined for infection levels, and morphologi-cal identification of VAM spores was made. All tested plants are considered mycorrhizal plants. We determined three spore species (Glomus mosseae, Glomus hoi and Scutellospora calospora) from the surveyed soils. The spore distribution rates were as follows: G. Mossea 61.54 %, G. Hoi 23.07 % and Scutellospora calospora 15.38 %. Results suggest that VAM fungal spores and root colonization display variation in rhizosphere under bulbous plants in different ecological conditions.
Key words: Taurus Mountains; bulbous plants; mycorrhiza; spore; morphological identification.
Received April 17, 2014; Accepted October 20, 2014
INTRODUCTION
The mutualistic association observed between vesicular-arbuscular mycorrhiza (VAM) and host plants is important in various natural and agricultural ecosystems (Sylvia and Williams, 1992). This association plays an essential role in enhancing plant growth in semi-arid agro-ecosystems (McGee, 1989), especially for plants grown in degraded soils.
The presence of arbuscular mycorrhizal fungi (AMF) may be essential for ecosystem sustain-ability, establishment of plants and maintenance of biological diversity. The participation of AMF in the biodiversity and ecosystem functioning is now being recognized, particularly due to their effect on plant diversity and productivity (van der Heijden et al., 1998). Several authors have report-ed positive relationships between plant diversity and AMF colonization (Grime et al., 1987; van der Heijden et al., 1998).

412 Karaarslan et al.
Mycorrhizal fungi provide inorganic nutrients, mainly phosphorus and other complexed com-pounds, to the plant through the extensive network of their hyphae that forage for soil nutrients more effectively than plant roots (Van der Heijden et al., 1998). For this association to occur, there must be a host plant (the phytobiont), an ecological habitat (the soil) and a suitable fungus (the mycobiont).
Interactions among AMF, host plants and en-vironmental factors (particularly the availability of nutrients in the soil and other soil organisms) are complex. Mycorrhizal associations of plant
roots with fungi are ubiquitous in nature and therefore, this symbiosis is of enormous ecologi-cal importance (Sykorová, 2007). In other words, soils, plants and management factors mainly af-fect the mycorrhizal fungi and their development in an ecosystem. There are some indications that the diversity of VAM fungal species declines from natural ecosystems to high-input agricultural sys-tems (Sharif and Moawad, 2006).
These fungi apparently have a limited toler-ance range to environmental conditions (Stahl et al., 1988) and possess specific adaptations to the
Table 1. Localization of the searched plants.
Number Name Localization Height References1 Crocus biflorus Mill, sub sp. isauricus
(Sieche ex Bowles) MathewKaraman; Sarıveliler, Asar Hill, rocky slopes
1750 m 25.04.2009, S. Doğu 1992 ve Bağcı
2 Eranthis hyemalis (L.) Salisb Karaman; Sarıveliler, Turanşah Mountain, Küllüin Locality, rocky localities
1750 m 25.04.2009, S. Doğu 1995 ve Bağcı.
3 Tulipa armena Boiss. var. lycica (Baker) Marais
Karaman; Sarıveliler, Kabalaktaş Hill, rocky sites
1750 m 25.04.2009, S. Doğu 2004 ve Bağcı.
4 Galanthus elwesii Hook. f. Karaman; Sarıveliler, Boyalı Y, rocky slopes
1600 m 25.04.2009, S. Doğu 2005 ve Bağcı.
5 Ornithogalum lanceolatum Labill. Karaman; Sarıveliler, Kabalaktaş Hill, rocky localities,
1750 m 25.04.2009, S. Doğu 2006 ve Bağcı.
6 Gagea villosa (Bieb.) Duby var. villosa Karaman; Sarıveliler, Turanşah Dağ, Küllüin Mevkii, rocky localities,
1750 m 25.04.2009, S. Doğu 2014 ve Bağcı.
7 Gagea granatellii (Parl.) Parl. Karaman; Ermenek, Elmay-urdu Köyü, Dulavrat T., rocky localities,
1550 m 25.04.2007, S. Doğu 1995 ve Bağcı.
8 Hyacinthella lazulina K.M. Press.&Jim. Press.
Karaman; Ermenek, Elmayurdu Village, Dulavrat T., Degraded Pinus opening
1760 m 25.04.2009, S. Doğu 2009 ve Bağcı.
9 Muscari muscarimi Medikus Karaman; Sarıveliler, Esentepe Village, İskeleç Locality, rocky localities,
1250 m 25.04.2009, S. Doğu 2012 ve Bağcı.
10 Muscari bourgaei Baker. Karaman; Sarıveliler, At Meydanı Site, rocky localities,
1200 m 25.04.2009, S. Doğu 2011 ve Bağcı.
11 Muscari armeniacum Leichtlin ex Baker Karaman; Sarıveliler, Çorak Mountain, rocky localities,
1520 m 25.04.2009, S. Doğu 2015 ve Bağcı.
12 Gagea peduncularis (J.&C.Presl) Pascher Karaman; Sarıveliler, Turanşah Mountain, Küllüin Site, rocky localities,
1750 m 25.04.2009, S. Doğu 2010.
13 Muscari neglectum GUSS. Karaman; Sarıveliler, Çorak Mountain, rocky localities,
1600 m 25.04.2009, S. Doğu 2016 ve Bağcı.

VAM IDENTIFICATION ON BULBOUS PLANTS 413
soil in which they occur (Lambert et al., 1980). These adaptations apparently can influence the outcome of competition between VAM fungi (Gerschefske Kitt et al., 1987). For example, high or low soil P levels (Boerner, 1990; Davis et al., 1984; Henkel et al., 1989; Haas and Krikun, 1985; Johnson, 1976; Krikun, 1983; Thomson et al., 1986), soil micronutrient levels (Killham, 1985), aridity (Bethlenfalvay et al., 1989; Sieverding and Toro, 1988; Simpson and Daft, 1990a; Stahl and Smith, 1984), salinity (Pond et al., (1984), low soil pH (Adelman and Morton, 1986; Hayman and Tavares, 1985; Howeler et al., 1987; Koomen et al., 1987; Porter et al., 1987b), toxic levels of met-als (Dueck et al., 1986; Gildon and Tinker, 1983; Koslowsky and Boerner, 1989), low or high tem-peratures (Dodd and Jeffries, 1986; Raju et al., 1990; Schenck and Smith 1982; Sieverding, 1988).
Indeed, mycorrhizas, not roots, are the main organs of nutrient uptake by most terrestrial plants. Mycorrhiza can exist in many forms; its morphology is determined by the characteris-tics of each partner involved and by the specific plant-fungus combination. This research was conducted to investigate morphological proper-ties of VAM in some bulbous plants grown in the Taurus Mountains.
MATERIALS AND METHODS
Description of the study sites
The vegetation of Anti-Taurus Mountains (Aladaglar), which are part of the middle Tau-rus Mountain system, includes vegetation types
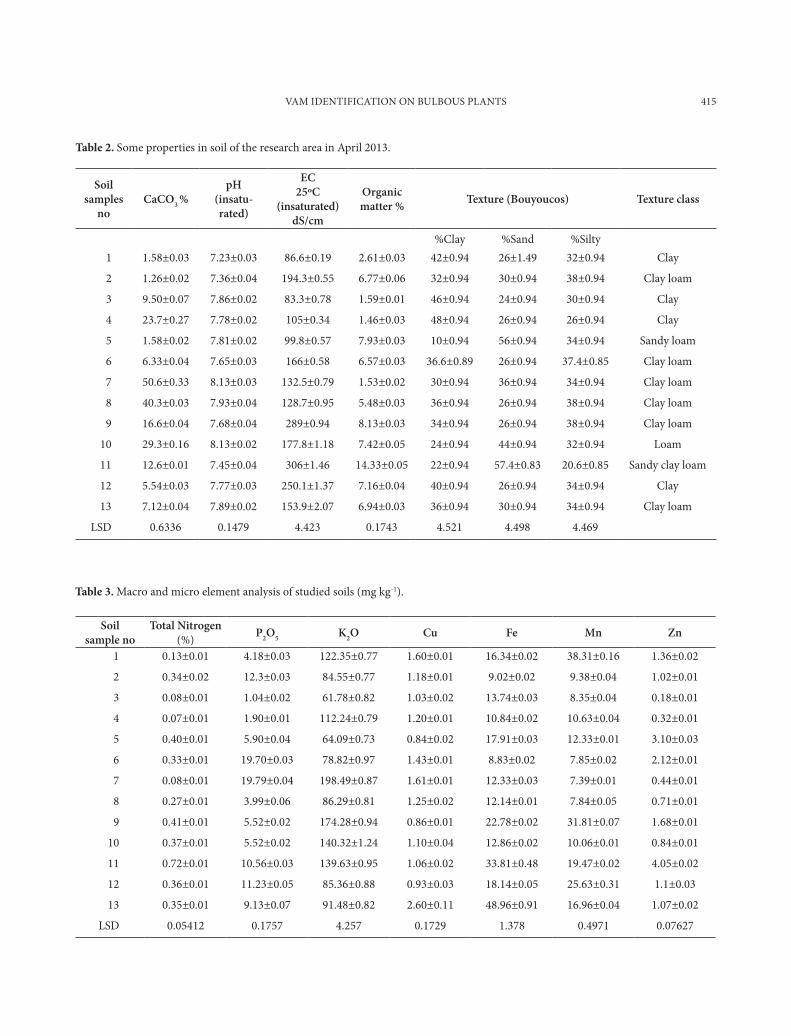
Fig. 1. Glomus mosseae

414 Karaarslan et al.
of Mediterranean, Iranian-Turan and European-Siberian floristic regions such as forests, shrubs and grassland formations. Several environmen-tal conditions such as climate, relief (topogra-phy), soil, hydrographic and biotic factors have a significant impact on vegetation development. These factors have led to rich and strong vegeta-tion formations. However, climate is the main factor that determines the rich biodiversity in the study area. Topography makes a difference by helping climate conditions; on the other hand, soil and hydrography have a local impact on veg-etation growth. Biotic factors have a negative im-pact on vegetation. In particular, human beings have been destroying the vegetation formations for several hundred years. Because of degrada-tion caused by humans, erosion has become one of the main problems in the study area (Toroğlu and Ünaldı, 2008).
Thirteen species of bulbous plants were sampled in the Taurus Mountains during the spring of 2013. The study was conducted in two localities. One of them is Karaman/Sarıveliler and the other is Karaman/Ermenek. These two areas are located in the central part of the Tau-rus Mountains. Two soil and plant materials were collected from the Ermenek location and ten samples were sampled from Sarıveliler. The sample areas’ height ranges from 1 200 to 1 760 m (Table 1.).
Soil and plant samples
Soil samples were taken from soil layers at 0 to 20 cm below the surface using a spade to collect about 1 kg of soil. The number of soil samples was based on the number of dominant native plant. Plant names, localities and heights are pre-sented in Table 1. The samples were placed in a plastic bag and stored in a refrigerator on the day
of collection; they were immediately refrigerated at 4°C when they arrived in the laboratory. Root fragments in these samples were examined for VAM colonization (Phillips and Hayman, 1970). The soil samples were then air-dried in the shade at laboratory temperature for spore counting and identification. The total vegetation cover is quite different in terms of density and endemic species. (The plants were identified by Biologist Dr. Yavuz Bağci according to the procedures de-scribed by Davis, 1965-1985).
Physical and chemical properties of the studied site soils
The soil samples taken from 0 to 20 cm depth were air dried and passed through a 2-mm sieve before laboratory analysis. The analysis was con-ducted to determine soil characteristics such as electrical conductivity (EC), pH, available P, K, total nitrogen, CaCO3, organic matter, texture, Fe, Zn, Cu, Mn and soil moisture. The electri-cal conductivity (EC) and pH were determined by using an electric and pH meter, respectively (Richards, 1954). The soluble potassium was de-termined with a flame photometer according to Knudsen et al. (1982). The available P was de-termined using a spectrophotometer according to Olsen et al. (1954). Total nitrogen was deter-mined according to Bremner (1965) using the Kjeldahl method while the CaCO3 percentage was determined according to Hızalan and Ünal (1965); organic matter was determined accord-ing to Jackson (1973) and texture according to Bouyoucos (1995) using the hydrometer meth-od. In addition, Zn, Fe, Mn and Cu contents of the soil samples were analyzed using ICP-AES (Soltanpour and Workman, 1981). All the soil analyses and measurements were triplicated and average values were used in the statistical analysis.
VAM IDENTIFICATION ON BULBOUS PLANTS 415
Table 2. Some properties in soil of the research area in April 2013.
Soil samples
noCaCO3 %
pH (insatu-rated)
EC 25ºC
(insaturated) dS/cm
Organic matter % Texture (Bouyoucos) Texture class
%Clay %Sand %Silty1 1.58±0.03 7.23±0.03 86.6±0.19 2.61±0.03 42±0.94 26±1.49 32±0.94 Clay
2 1.26±0.02 7.36±0.04 194.3±0.55 6.77±0.06 32±0.94 30±0.94 38±0.94 Clay loam
3 9.50±0.07 7.86±0.02 83.3±0.78 1.59±0.01 46±0.94 24±0.94 30±0.94 Clay
4 23.7±0.27 7.78±0.02 105±0.34 1.46±0.03 48±0.94 26±0.94 26±0.94 Clay
5 1.58±0.02 7.81±0.02 99.8±0.57 7.93±0.03 10±0.94 56±0.94 34±0.94 Sandy loam
6 6.33±0.04 7.65±0.03 166±0.58 6.57±0.03 36.6±0.89 26±0.94 37.4±0.85 Clay loam
7 50.6±0.33 8.13±0.03 132.5±0.79 1.53±0.02 30±0.94 36±0.94 34±0.94 Clay loam
8 40.3±0.03 7.93±0.04 128.7±0.95 5.48±0.03 36±0.94 26±0.94 38±0.94 Clay loam
9 16.6±0.04 7.68±0.04 289±0.94 8.13±0.03 34±0.94 26±0.94 38±0.94 Clay loam
10 29.3±0.16 8.13±0.02 177.8±1.18 7.42±0.05 24±0.94 44±0.94 32±0.94 Loam
11 12.6±0.01 7.45±0.04 306±1.46 14.33±0.05 22±0.94 57.4±0.83 20.6±0.85 Sandy clay loam
12 5.54±0.03 7.77±0.03 250.1±1.37 7.16±0.04 40±0.94 26±0.94 34±0.94 Clay
13 7.12±0.04 7.89±0.02 153.9±2.07 6.94±0.03 36±0.94 30±0.94 34±0.94 Clay loam
LSD 0.6336 0.1479 4.423 0.1743 4.521 4.498 4.469
Table 3. Macro and micro element analysis of studied soils (mg kg-1).
Soil sample no
Total Nitrogen (%) P2O5 K2O Cu Fe Mn Zn
1 0.13±0.01 4.18±0.03 122.35±0.77 1.60±0.01 16.34±0.02 38.31±0.16 1.36±0.02
2 0.34±0.02 12.3±0.03 84.55±0.77 1.18±0.01 9.02±0.02 9.38±0.04 1.02±0.01
3 0.08±0.01 1.04±0.02 61.78±0.82 1.03±0.02 13.74±0.03 8.35±0.04 0.18±0.01
4 0.07±0.01 1.90±0.01 112.24±0.79 1.20±0.01 10.84±0.02 10.63±0.04 0.32±0.01
5 0.40±0.01 5.90±0.04 64.09±0.73 0.84±0.02 17.91±0.03 12.33±0.01 3.10±0.03
6 0.33±0.01 19.70±0.03 78.82±0.97 1.43±0.01 8.83±0.02 7.85±0.02 2.12±0.01
7 0.08±0.01 19.79±0.04 198.49±0.87 1.61±0.01 12.33±0.03 7.39±0.01 0.44±0.01
8 0.27±0.01 3.99±0.06 86.29±0.81 1.25±0.02 12.14±0.01 7.84±0.05 0.71±0.01
9 0.41±0.01 5.52±0.02 174.28±0.94 0.86±0.01 22.78±0.02 31.81±0.07 1.68±0.01
10 0.37±0.01 5.52±0.02 140.32±1.24 1.10±0.04 12.86±0.02 10.06±0.01 0.84±0.01
11 0.72±0.01 10.56±0.03 139.63±0.95 1.06±0.02 33.81±0.48 19.47±0.02 4.05±0.02
12 0.36±0.01 11.23±0.05 85.36±0.88 0.93±0.03 18.14±0.05 25.63±0.31 1.1±0.03
13 0.35±0.01 9.13±0.07 91.48±0.82 2.60±0.11 48.96±0.91 16.96±0.04 1.07±0.02
LSD 0.05412 0.1757 4.257 0.1729 1.378 0.4971 0.07627

416 Karaarslan et al.
Assessment of arbuscular mycorrhizal fungi colonization and spores
The root samples were washed carefully with deionized water and the roots were cleared. The staining procedure was performed according to the method described by Koske and Gemma (1989). Roots were treated with KOH (10%) for 15-20 min at 121ºC in an autoclave. Clear pieces of roots were rinsed with tap water to remove KOH. Dark roots were further bleached with 10% H2O2 for 3 min at room temperature. Then the roots were acidified with 2% HCl (v/v) for 1 h at room temperature and stained with Try-pan blue (0.05%) (Phillips and Hayman, 1970), with the lactophenol being changed to lacto-glycerol (Lactic acid:glycerol:deionized water, in a 5:1:1 ratio). Excess stain was removed using 50% glycerol for 1-2 h. Samples were then stored for microscopic analysis. Three slides, each with 10 randomly selected stained roots (1 cm long) were prepared from every individual plant sam-ple. A total of 30 roots per species were examined to determine structures characteristic of VAM.
The percentages of root colonization were examined under a microscope (Novew b-series biological trinocular microscope) at 10X-20 X and 40 X magnification and were calculated by the gridline intersect method (Giovanetti and Mosse, 1980).
Each soil sample (10 g fresh mass) was sieved according to the sieving and decanting procedure of Gerdeman and Nicolson (1963) and INVAM (2004). The sieved samples were centrifuged at 3500 rpm (10 g) for 10 min. The pellet was resuspended in 50% sucrose and centrifuged again at 3500 rpm (10 g) for 1 min. After centrifuging, the superna-tants were poured on to 38-50, 50-100, 100-250 and >250μm sieve team, washed thoroughly with de-ionized water and then placed in 9-cm Petri dishes for examination under a stereomicroscope (Olym-
pus SX 60 trinocular microscope) at 40 X magni-fication. Turgid spores (suggesting viability) were grouped under the dissecting microscope accord-ing to morphological characteristics. Diagnostic permanent slides were prepared for each different spore morphotype using either polyvinyl alcohol alone or mixed with Melzer’s solution (1:1).
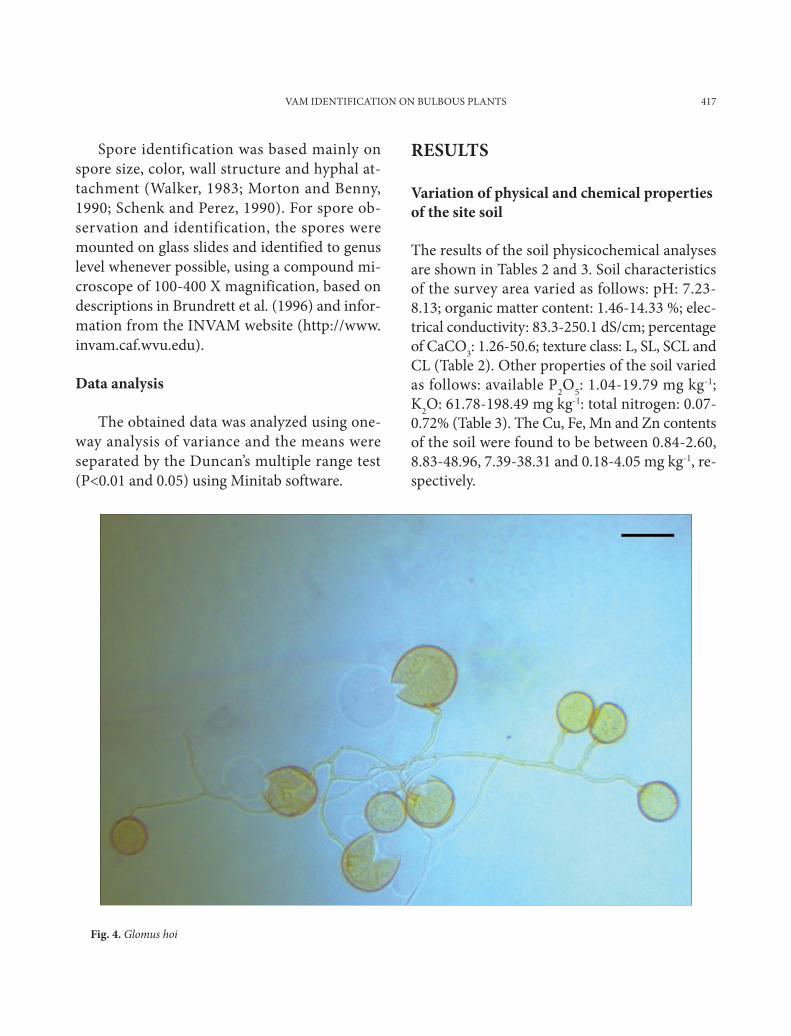
Fig. 2. Sporocarps in the soil-water suspension
Fig. 3. Mycorrhizal infected in sampled plant roots (Crocus biflorus Mill, subsp. isauricus (Siehe ex Bowles) Mathew
VAM IDENTIFICATION ON BULBOUS PLANTS 417
Spore identification was based mainly on spore size, color, wall structure and hyphal at-tachment (Walker, 1983; Morton and Benny, 1990; Schenk and Perez, 1990). For spore ob-servation and identification, the spores were mounted on glass slides and identified to genus level whenever possible, using a compound mi-croscope of 100-400 X magnification, based on descriptions in Brundrett et al. (1996) and infor-mation from the INVAM website (http://www.invam.caf.wvu.edu).
Data analysis
The obtained data was analyzed using one-way analysis of variance and the means were separated by the Duncan’s multiple range test (P<0.01 and 0.05) using Minitab software.
RESULTS
Variation of physical and chemical properties of the site soil
The results of the soil physicochemical analyses are shown in Tables 2 and 3. Soil characteristics of the survey area varied as follows: pH: 7.23-8.13; organic matter content: 1.46-14.33 %; elec-trical conductivity: 83.3-250.1 dS/cm; percentage of CaCO3: 1.26-50.6; texture class: L, SL, SCL and CL (Table 2). Other properties of the soil varied as follows: available P2O5: 1.04-19.79 mg kg-1; K2O: 61.78-198.49 mg kg-1: total nitrogen: 0.07-0.72% (Table 3). The Cu, Fe, Mn and Zn contents of the soil were found to be between 0.84-2.60, 8.83-48.96, 7.39-38.31 and 0.18-4.05 mg kg-1, re-spectively.
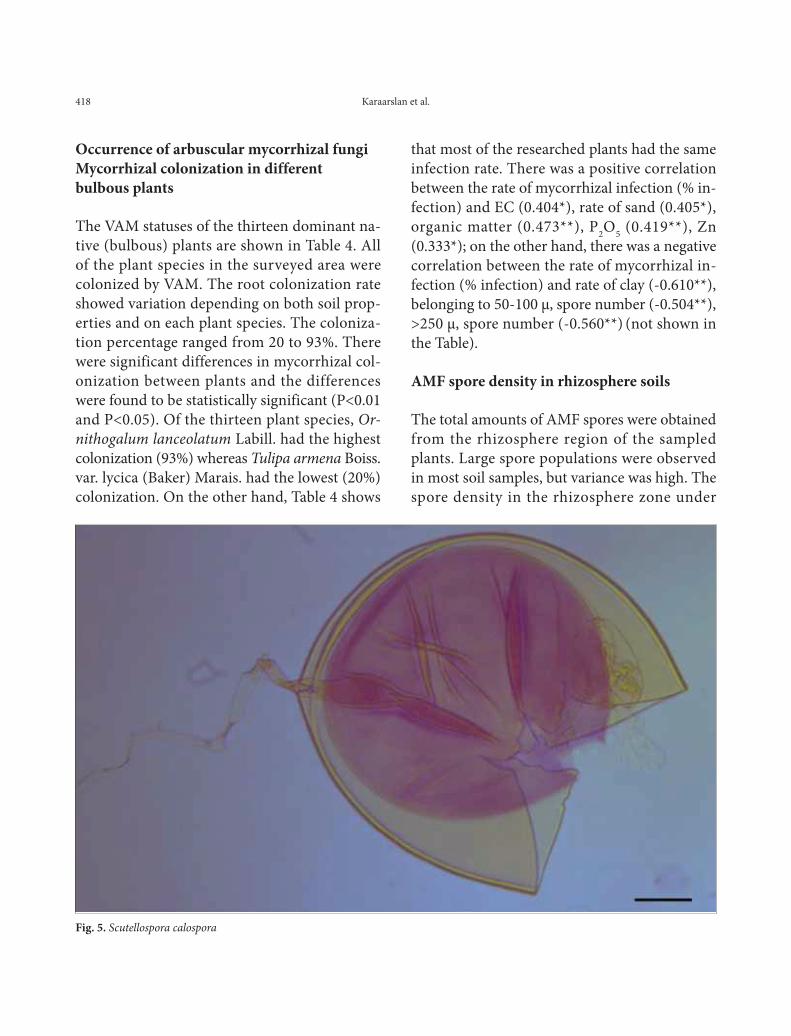
Fig. 4. Glomus hoi
418 Karaarslan et al.
Occurrence of arbuscular mycorrhizal fungi Mycorrhizal colonization in different bulbous plants
The VAM statuses of the thirteen dominant na-tive (bulbous) plants are shown in Table 4. All of the plant species in the surveyed area were colonized by VAM. The root colonization rate showed variation depending on both soil prop-erties and on each plant species. The coloniza-tion percentage ranged from 20 to 93%. There were significant differences in mycorrhizal col-onization between plants and the differences were found to be statistically significant (P<0.01 and P<0.05). Of the thirteen plant species, Or-nithogalum lanceolatum Labill. had the highest colonization (93%) whereas Tulipa armena Boiss. var. lycica (Baker) Marais. had the lowest (20%) colonization. On the other hand, Table 4 shows
that most of the researched plants had the same infection rate. There was a positive correlation between the rate of mycorrhizal infection (% in-fection) and EC (0.404*), rate of sand (0.405*), organic matter (0.473**), P2O5 (0.419**), Zn (0.333*); on the other hand, there was a negative correlation between the rate of mycorrhizal in-fection (% infection) and rate of clay (-0.610**), belonging to 50-100 µ, spore number (-0.504**), >250 µ, spore number (-0.560**) (not shown in the Table).
AMF spore density in rhizosphere soils
The total amounts of AMF spores were obtained from the rhizosphere region of the sampled plants. Large spore populations were observed in most soil samples, but variance was high. The spore density in the rhizosphere zone under
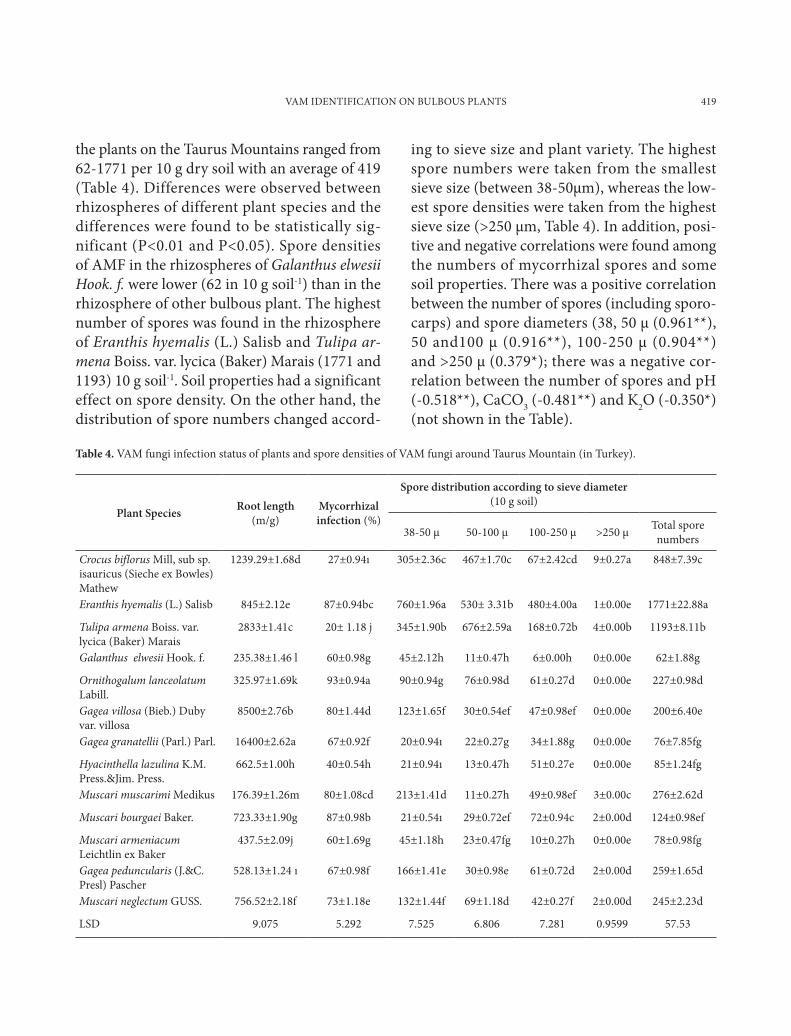
Fig. 5. Scutellospora calospora
VAM IDENTIFICATION ON BULBOUS PLANTS 419
the plants on the Taurus Mountains ranged from 62-1771 per 10 g dry soil with an average of 419 (Table 4). Differences were observed between rhizospheres of different plant species and the differences were found to be statistically sig-nificant (P<0.01 and P<0.05). Spore densities of AMF in the rhizospheres of Galanthus elwesii Hook. f. were lower (62 in 10 g soil-1) than in the rhizosphere of other bulbous plant. The highest number of spores was found in the rhizosphere of Eranthis hyemalis (L.) Salisb and Tulipa ar-mena Boiss. var. lycica (Baker) Marais (1771 and 1193) 10 g soil-1. Soil properties had a significant effect on spore density. On the other hand, the distribution of spore numbers changed accord-
ing to sieve size and plant variety. The highest spore numbers were taken from the smallest sieve size (between 38-50µm), whereas the low-est spore densities were taken from the highest sieve size (>250 µm, Table 4). In addition, posi-tive and negative correlations were found among the numbers of mycorrhizal spores and some soil properties. There was a positive correlation between the number of spores (including sporo-carps) and spore diameters (38, 50 µ (0.961**), 50 and100 µ (0.916**), 100-250 µ (0.904**) and >250 µ (0.379*); there was a negative cor-relation between the number of spores and pH (-0.518**), CaCO3 (-0.481**) and K2O (-0.350*) (not shown in the Table).
Table 4. VAM fungi infection status of plants and spore densities of VAM fungi around Taurus Mountain (in Turkey).
Plant Species Root length (m/g)
Mycorrhizal infection (%)
Spore distribution according to sieve diameter (10 g soil)
38-50 μ 50-100 μ 100-250 μ >250 μ Total spore numbers
Crocus biflorus Mill, sub sp. isauricus (Sieche ex Bowles) Mathew
1239.29±1.68d 27±0.94ı 305±2.36c 467±1.70c 67±2.42cd 9±0.27a 848±7.39c
Eranthis hyemalis (L.) Salisb 845±2.12e 87±0.94bc 760±1.96a 530± 3.31b 480±4.00a 1±0.00e 1771±22.88a
Tulipa armena Boiss. var. lycica (Baker) Marais
2833±1.41c 20± 1.18 j 345±1.90b 676±2.59a 168±0.72b 4±0.00b 1193±8.11b
Galanthus elwesii Hook. f. 235.38±1.46 l 60±0.98g 45±2.12h 11±0.47h 6±0.00h 0±0.00e 62±1.88g
Ornithogalum lanceolatum Labill.
325.97±1.69k 93±0.94a 90±0.94g 76±0.98d 61±0.27d 0±0.00e 227±0.98d
Gagea villosa (Bieb.) Duby var. villosa
8500±2.76b 80±1.44d 123±1.65f 30±0.54ef 47±0.98ef 0±0.00e 200±6.40e
Gagea granatellii (Parl.) Parl. 16400±2.62a 67±0.92f 20±0.94ı 22±0.27g 34±1.88g 0±0.00e 76±7.85fg
Hyacinthella lazulina K.M. Press.&Jim. Press.
662.5±1.00h 40±0.54h 21±0.94ı 13±0.47h 51±0.27e 0±0.00e 85±1.24fg
Muscari muscarimi Medikus 176.39±1.26m 80±1.08cd 213±1.41d 11±0.27h 49±0.98ef 3±0.00c 276±2.62d
Muscari bourgaei Baker. 723.33±1.90g 87±0.98b 21±0.54ı 29±0.72ef 72±0.94c 2±0.00d 124±0.98ef
Muscari armeniacum Leichtlin ex Baker
437.5±2.09j 60±1.69g 45±1.18h 23±0.47fg 10±0.27h 0±0.00e 78±0.98fg
Gagea peduncularis (J.&C.Presl) Pascher
528.13±1.24 ı 67±0.98f 166±1.41e 30±0.98e 61±0.72d 2±0.00d 259±1.65d
Muscari neglectum GUSS. 756.52±2.18f 73±1.18e 132±1.44f 69±1.18d 42±0.27f 2±0.00d 245±2.23d
LSD 9.075 5.292 7.525 6.806 7.281 0.9599 57.53
420 Karaarslan et al.Ta
ble
5. M
orph
olog
ical
char
acte
r of s
pore
s of V
AM
fung
i in
stud
ied
plan
ts.
(http
://w
ww.
zor.z
ut.ed
u.pl
/Glo
mer
omyc
ota/
Spec
ies%
20de
scrip
tions
%20
of%
20A
MF.h
tml a
nd (h
ttp://
inva
m.w
vu.ed
u/th
efun
gi/c
lass
ifica
tion/
glom
acea
e/gl
omus
)
Mor
pho-
lo
gica
l ch
arac
ters
fo
r ide
nti-
ficat
ion
Plan
ts a
nd S
oils
of N
umbe
r
1.2.
3.4.
5.6.
7.8.
9.10
.11
.12
.13
.
Shap
e:
Glo
bose
to
ovoi
dG
lobo
se,
subg
lobo
se,
ellip
soid
al o
r irr
egul
ar
Glo
bose
to
ovoi
dG
lobo
se to
ov
oid
Glo
bose
, su
bglo
bose
, el
lipso
idal
or
irreg
ular
Glo
bose
to
ovoi
dG
lobo
se to
ov
oid
Glo
bose
to
subg
lobo
seG
lobo
se to
ov
oid
Glo
bose
to
ovoi
dgl
obos
e, su
bglo
bose
, el
lipso
idal
or
irreg
ular
Glo
bose
to
subg
lobo
seG
lobo
se to
ov
oid
Size
:10
5-31
0x
110-
305µ
20-3
0(-5
0)
µ di
am
(50-
) 80-
120
(-15
5) x
(45-
) 75
-120
(-14
0)
µm d
iam
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
(50-
) 80-
120
(-15
5) x
(45-
) 75
-120
(-14
0)
µm d
iam
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
(160
-)21
4
(-26
0) o
r 15
0-19
0x
210-
260
µ
diam
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
(50-
) 80-
120
(-15
5) x
(45-
) 75
-120
(-14
0)
µm d
iam
(160
-)21
4
(-26
0) o
r 15
0-19
0x
210-
260
µ
diam
105-
310x
11
0-30
5µ
20-3
0(-5
0)
µ di
am
Col
or:
Pale
ye
llow
(2
A2)
to
gol
den
ye
llow
(5
B8)
light
yel
low
to
oran
ge-y
ello
wPa
le
yello
w
(2A
2)
to g
olde
n
yello
w
(5B8
)
Pale
ye
llow
(2
A2)
to
gol
den
ye
llow
(5
B8)
light
yel
low
to
oran
ge-y
ello
wPa
le y
ello
w
(2A
2) to
go
lden
ye
llow
(5
B8)
Pale
yel
low
(2
A2)
to
gold
en
yello
w
(5B8
)
Past
el
yello
w
(2A
4)
to d
ark
or
ange
(5
A8)
Pale
ye
llow
(2
A2)
to
gol
den
ye
llow
(5
B8)
Pale
ye
llow
(2
A2)
to
gol
den
ye
llow
(5
B8)
light
yel
low
to
oran
ge-y
ello
wPa
stel
ye
llow
(2
A4)
to
dark
or
ange
(5
A8)
Pale
ye
llow
(2
A2)
to
gol
den
ye
llow
(5
B8)
Hyp
hal
atta
chm
ent:
Sept
ate
hy
phae
(2
-12µ
)
Subt
endi
ng
hyph
a sin
gle
Sept
ate
hy
phae
(2
-12µ
)
Sept
ate
hy
phae
(2
-12µ
)
Subt
endi
ng
hyph
a sin
gle
Sept
ate
hy
phae
(2
-12µ
)
Sept
ate
hy
phae
(2
-12µ
)
Bulb
ous
susp
enso
rSe
ptat
e
hyph
ae
(2-1
2µ)
Sept
ate
hy
phae
(2
-12µ
)
Subt
endi
ng
hyph
a sin
gle
Bulb
ous
susp
enso
rSe
ptat
e
hyph
ae
(2-1
2µ)
Auxi
liary
ce
ll:no
neno
neno
neno
neno
neno
neno
neno
neno
neno
neno
neno
neno
ne
Spor
ocar
p:pr
esen
cepr
esen
cepr
esen
cepr
esen
cepr
esen
cepr
esen
cepr
esen
cepr
esen
ceno
nepr
esen
cepr
esen
ceno
nepr
esen
ce
Ger
min
atio
n sh
ield
:no
neno
neno
neno
neno
neno
neno
nepr
esen
ceno
neno
neno
nepr
esen
ceno
ne
Surf
ace
or
nam
enta
-tio
n:
No,
but
O
uter
laye
rs
of th
e sp
ore
wal
l ofte
n slo
ugh
as
the
spor
e ag
es (i
n so
il or
in p
ot
cultu
re
stor
age)
.
No,
but
Out
er
laye
rs o
f the
sp
ore
wal
l of-
ten
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
No,
but
O
uter
laye
rs
of th
e sp
ore
wal
l ofte
n slo
ugh
as
the
spor
e ag
es (i
n so
il or
in p
ot
cultu
re
stor
age)
.
No,
but
O
uter
laye
rs
of th
e sp
ore
wal
l ofte
n slo
ugh
as
the
spor
e ag
es (i
n so
il or
in p
ot
cultu
re
stor
age)
.
No,
but
Out
er
laye
rs o
f the
sp
ore
wal
l of-
ten
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
No,
but
O
uter
la
yers
of
the
spor
e w
all o
ften
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
No,
but
O
uter
la
yers
of
the
spor
e w
all o
ften
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
with
or
with
out
orna
men
-ta
tions
, ou
ter o
rna-
men
tatio
ns
deve
lop
(if
pre
sent
in
the
sp
ecie
s)
No,
but
O
uter
la
yers
of
the
spor
e w
all o
ften
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
No,
but
O
uter
la
yers
of
the
spor
e w
all o
ften
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
No,
but
Out
er
laye
rs o
f the
sp
ore
wal
l of-
ten
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
with
or
with
out
orna
men
-ta
tions
oute
r orn
a-m
enta
tions
de
velo
p
(if p
rese
nt
in th
e
spec
ies)
No,
but
O
uter
la
yers
of
the
spor
e w
all o
ften
sloug
h as
th
e sp
ore
ages
(in
soil
or in
po
t cul
ture
st
orag
e).
Vesic
le:
pres
ence
pres
ence
pres
ence
pres
ence
pres
ence
pres
ence
pres
ence
none
pres
ence
pres
ence
pres
ence
none
pres
ence

VAM IDENTIFICATION ON BULBOUS PLANTS 421
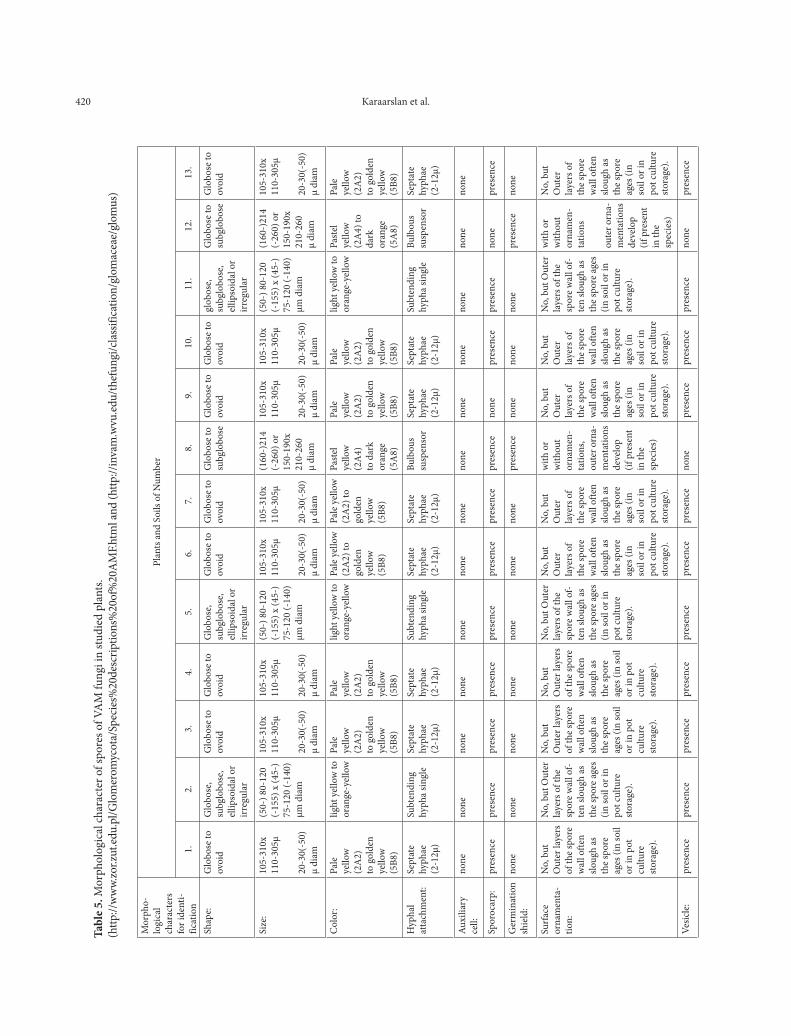
Morphological identification of VAM spore
Spores were isolated from the soil of the research area’s soil and then examined for some proper-ties determined by INVAM (Table 5.) .The spore colors were also determined according to INVAM colors chart. G. mosseae was the most common species among the samples, being found in 85% of the soil samples collected (Table 6.). These species are typical of arid and semi-arid environments. It has been reported that G. mosseae has a vari-able response to different environmental condi-tions due to the variable phenotypic plasticity of this species (Heslop-Harrison, 1964). The Glomus genus was reported to be dominant in arid and semi-arid climates due to its resistance to high soil temperature (Al-Raddad, 1993; Mohammad et al., 2003). Spores of Glomus mosseae (Fig. 1) were found in abundance in almost every soil sample, and spores of Glomus hoi (Fig. 4) were also identified in the soil samples. Another species determined was Scutellospora calospora (Fig. 5).
Indigenous VAM fungal communities gener-ally contain several fungal species. Normally 5-15 species may be found in agro-ecosystems. The spatial distribution of VAM fungal species can vary and even when the number is the same at two different soil sites, the species composition of the fungal population can be completely different (Sieverding, 1989). The spores of VAM fungi are very distinctive. Generally, spore density in the soil samples seemed to be dominated by the species of Glomus mosseae, Glomus hoi and S. calospora.
The properties of the identified spore species are as follows:
Glomus mosseae (Nicol. & Gerd.) Gerd. & Trappe. Sporocarps have 1-10 spores, are more or less spherical, their shapes vary from globose to ellipsoid, and up to 1 mm in diameter. Peridium
is variable, consisting of irregularly-branched septate hyphae frequently anastomosing to form a thin network, enclosing the chlamydospores entirely, or incomplete with spores extruding; sporocarps containing chlamydospores, some-times only one, usually 2-6, but up to 32 in num-ber; chlamydospores yellow, spherical to oval, occasionally irregularly-shaped, variable in size 60-320 µ, generally with only one but occasional-ly two distinct funnel-shaped attachments. Walls of spores have two layers, a thin outer layer and a thick inner layer. The inner wall is 2-7 µ; ectocar-pic spores are similar to endocarpic spores, abun-dant in some collections, rare in others, formed in soil or sometimes in root tissue.
This species is named in honor of Dr Bar-bara Mosse, who was the first to demonstrate ex-perimentally that a sporocarpic Endogone could produce endotrophic mycorrhizae (Mosse, 1953, 1956). Spores are single in the soil, in loose ag-gregates or compact sporocarps; pale yellow (2A2) to golden yellow (5B8) (Nicolson and Ger-deman 1968) (Fig. 1) (http://www.zor.zut.edu.pl/Glomeromycota/Glomus%20mossea.html).
Glomus hoi. This description is a combina-tion of information obtained from the protologue (Berch and Trappe, 1985), type specimens, and universal patterns of morphological organization and structure in Glomaceae. A living culture of this species has never been obtained by INVAM.
Spores are borne singly in the soil, globose, subglobose, ellipsoidal or irregular in shape, (50-) 80-120 (-155) x (45-) 75-120 (-140) µm in diam-eter and light brown in color. The spore wall is composed of two distinct, separable layers (L1 and L2). The outer layer (L1) is light yellow to orange-yellow in color under transmitted light; (2-) 4-6 (-8) µm thick, with an outer surface that fractures and sloughs. The inner layer (L2)

422 Karaarslan et al.
is hyaline to light yellow, very thin, 1 um thick. The subtending hypha is single, cylindrical or slightly flared toward the point of attachment, (5-) 8-11(-13) µm wide at the spore base. The hyphal wall consists of a single layer, 2.5-5 µm thick, sometimes bearing fine, thin-walled, sep-tate, lateral branches. The occlusion appears to be a thin, curved septum in the hypha lumen at or somewhat below its point of attachment to the spore (Berch and Trappe, 1985) (Fig. 4).
Scutellospora calospora (Nicol. & Gerd.) Walker & Sanders. Spores formed singly in the soil are terminally on a bulbous suspensor-like cell, translucent, hyaline to pale greenish-yellow; globose, ellipsoidal or cylindrical, occasionally broader than long; 114-285(-511) x 110-412(-511) µm. There are four walls in the spore struc-ture and they are divided into two groups (groups A and B). Group A: i.e. the inner wall, is brittle, hyaline to pale yellow, very finely laminated wall (wall 2) 3-5 µm thick that may be surrounded by a thin, very closely appressed hyaline unit wall (wall 1), 0.5-1 µm thick. Group B has two hyaline membranous walls (walls 3 and 4). Wall 3 is 0.5-1 µm thick, often wrinkling in crushed spores. Wall 4 is 1-1.5 µm thick, staining red in Melzer’s re-agent. Germination shield is oval, 35-70 x 50-90 µm, often with invaginations along the margin. Suspensor-like cell is borne terminally on a sep-tate subtending hypha, 33-48 µm wide; walls of the spore are concolorous with each other; walls are 1 µm thick distally, thickening somewhat near the spore base (Koske and Walker, 198
DISCUSSION
This is the first study reporting the morphologi-cal identification of VAM associated with bul-bous plants of the Taurus Mountains in Turkey.
We have determined rates of VAM colonization in plant roots and mycorrhizal spore number in rhizosphere soils belonging to bulbous plants. The results showed that soils in two locations of the Taurus Mountains contained the highest num-ber of mycorrhiza spores and the highest mycor-rhizal infection rate in plant roots. The number of spores examined (62-1771 spores/10g soil) is much higher than the number isolated from agricultural soils (0.1 to 5 spores/g soil, Mosse 1979). However, there was much variability in the number of spores from rhizosphere to rhi-zosphere. We found that the numbers of VAM spores vary with soil texture. The clay-loamy and clay soils were generally richer in VAM spores (with high variation in frequency), while sandy soil contained lower numbers of endogonaceous spores, the range being from 78-124 spores/l0 g soil. The present survey indicated strong depen-dence of VAM spores on the pH and CaCO3, K2O content of the soil, but this dependence was found as negative. Unlike findings by other researchers, we found a very high percentage of organic matter present in the topsoil since the topsoil was cov-ered with plant residue. Sheikh et al. (1975) did not find high numbers of spores with respect to organic matter levels (Table 2). However, the re-sults of some other studies show that organic mat-ter content of the soil was positively related to the endogonaceous spore population. Our findings about the number of spores have shown fluctua-tion depending on organic matter content. The results obtained from this survey further showed that the rate of mycorrhizal colonization showed variation among bulbous plants and the root colo-nization rate varied depending on both soil prop-erties and plant species. The variation could be due to environmental factors and physicochemi-cal characteristics of the soil, which can have an effect on colonization. Also, we found significant and positive effects on plant root colonization of organic matter, total nitrogen, phosphorus, EC,

VAM IDENTIFICATION ON BULBOUS PLANTS 423
Zn and soil texture (r=0.473, 0.473, 0.419, 0.404, 0.333, 0.405, respectively). We can say that the mentioned parameters increased root infection rate in such a way that P2O5 is one of the impor-tant factors that affects both mycorrhizal infec-tion rate and root length. It affected both mycor-rhizal infection rate and root length positively. Generally, a high density of P2O5 had a negative effect on mycorrhizal colonization but the level of P2O5 in the studied soil was not very high for the colonization rate. Furthermore, the impor-tance of soil parameters for the diversity of the VAM fungus species is not well known, but over-all VAM fungus production in the form of spore propagules may increase with soil pH and organic carbon, and decrease with increasing amounts of soil phosphorous. The level of phosphorus in the plant has also been shown to influence the es-tablishment of VAM with high levels inhibiting colonization by mycorrhizae (Menge et al., 1978).
Table 6. Occurrence of genera and species of VAM fungi and their hosts Taurus Mountains of two locations (Karaman/Sarıveliler, Karaman/Ermenek). Percent occurrence is based on the number of plant species associated with a specific genus and species of VAM fungi divided by the total (13) major plant spe-cies present.
Genus Species % Occurrence
Hosts
Glomus mosseae 61.54 1, 3, 4, 6, 7, 9, 10, 13
hoi 23.07 2, 5, 11
Scutellospora calospora 15.38 8, 12
In the present study, the population dynamics of AM fungi were determined by collecting the resting spores under rhizosphere soils of differ-ent bulbous plants. As VAM fungi are widespread, they occurred in almost all soils, but with a varia-tion in both number and type of spores and sporo-carps. The surveyed soils gave very high numbers of VAM propagules and they changed from plant to plant. This is due to low nutrient status (1.04-19.79 mg/kg of P2O5 and 61.78-198.49 mg/kg
of K), zinc (0.18-4.05 mg/kg). Altogether, three VAM fungal species were isolated from thirteen different bulbous plants under rhizosphere soils. The VAM fungal species belonging to the genera of Glomus, and Scutellospora were isolated.
In conclusion, there is no direct relation be-tween the number of spores and the number of species. It is a well-established fact that these two factors need not be directly proportional to one another. In the present study, Glomus dominated the rhizosphere soils of bulbous plants. This was similar to earlier reports of VAM association on other vegetable crops (Bagyaraj et al., 1979; Subha, 2001). Similar reference was observed in the case of chili (Bagyaraj and Manjunath, 1980; Bagyaraj and Sreeramulu, 1982; Manoharachary and Su-lochana, 1989; Janakirani, 1991; Vani, 1993; Hin-dumathi, 1999; Subha, 2001) in studies of the AM fungal association in onion, chili, okra, sesame, safflower, castor, sorghum and sweet potato. Also, Karaarslan and Uyanöz (2010; 2011) reported that Glomus dominated the rhizosphere soils of natu-ral plants. We can say that early researchers ob-tained results similar to the present study (Kara-arslan and Uyanöz, 2008; Uyanöz and Karaarslan, 2012). Spores, subtending hyphae, and sporocarp morphology are used as taxonomical features in VAM fungal identification. However, fungal anat-omy in roots is not generally used in taxonomi-cal descriptions to separate taxa below the generic level. Morton (1985) and Hall (1977) reached the conclusion that the morphological characteristics (spore size, color and structure) and ontogenetic characteristics (production of arbuscules, vesicles and number of spores) were not influenced sig-nificantly by the host.
All major genera (Acaulospora, Entrophospora, Gigaspora, Scutellospora, Sclerocystis and Glomus) of VAM fungi have been found in arid, semi-ar-id, and sand dune soils around the globe (Khan,

424 Karaarslan et al.
1974; El-Giahmi et al., 1976; Redhead, 1977; Diem et al., 1981; Schwab and Reeves, 1981; Bloss, 1985; Halvorson and, Koske, 1988; McGee, 1989). Of the 148 species of VAM fungi, 31 species were first re-ported from sand dunes (Schenck and Pérez, 1990).
The 31 species are in three genera: Glomus (14 species), Scutellospora (12 species), and Acau-lospora (5 species). VAM inoculum density varies from site to site and decreases with soil depth (Sutton and Barron, 1972; Miller, 1979; Reeves et al., 1979; Buchholz and Motto, 1981; Schwab and Reeves, 1981; Allen et al., 1984; Zajicek et al., 1986; Koide and Mooney, 1987). The distribu-tion of VAM fungal species appears to be more closely related to host plant, soil structure and environmental conditions than to the competi-tion by other VAM fungal species (Koske, 1981).
REFERENCES
Adelman, M.J. and J.B. Morton (1986). Infectivity of vesicular-arbuscular mycorrhizal fungi: influence of host-soil diluent combinations on MPN estimates and percentage coloniza-tion. Soil Biol. Biochem. 18, 77-83.
Allen, M.F., MacMahon, J.A. and D.C. Andersen (1984). Re-estab-lishment of Endogonaceae on Mount St. Helens: survival of residuals. Mycologia 76: 1031-1038.
Bagyaraj, D.J. (1979). Plant Soil 51, 397-403.Bagyaraj, D.J and A. Manjunath (1980). Selection of a suitable
host for mass production of VA mycorrhizal inoculum. Plant Soil 55: 495-498.
Bagyaraj, D.J. and K.R. Sreeramulu (1982). Preinoculation with V.A. mycorrhiza improves growth and yield of chili trans-planted şn the field and saves phosphatic fertilizer. Plant Soil 69: 375-381.
Berch, S.M. and J.M. Trappe (1985). A new species of Endogona-ceae, Glomus hoi. Mycologia 77: 654-657.
Bethlenfalvay, G.J., Franson, R.L., Brown, M.S. and K.L. Mihara (1989). The Glycine-Glomus-Rhizobium symbiosis. XI. Nutritional, morphological and physiological responses of nodulated soybean to geographic isolates of the mycor-rhizal fungus Glomus mosseae. Physiol. Plant. 76, 226-232.
Blaskowski, J. (2003). Species Descriptions and Illustrations. From http://www.zor.zut.edu.pl/Glomeromycota/Species%20descriptions%20of%20AMF.html
Bloss, H.E. (1985). Studies of symbiotic microflora and their role in the ecology of desert plants. Desert Plants 7: 119-127.
Boerner, R.E.J. (1990). Role of mycorrhizal fungus origin in growth and nutrient uptake by Geranium robertianum. Am. J. Bot. 77, 483-489.
Bouyoucos, G.J. (1995). A recalibration of the hydrometer method for making mechanical analysis of the soils. Agronomy J. 4(9): 434.
Bremner, J.M. (1965). Methods of Soil Analysis Part 2. Chemical and Microbiological Properties. Ed. A. C. A. Black Amer. Soc. of Agron Inc. Pub. Agron. Series No: 9 Madison USA.
Brundrett, M., Bougher, N., Dell, B., Grove, T. and N. Malajczuk (1996). Working with mycorrhizas in forestry and agricul-ture. ACIAR Monograph 32, 374.
Buchholz, K. and H. Motto (1981). Abundance and vertical dis-tributions of mycorrhizae in Plains and Barren forest soils from the New Jersey Pine Barrens. Bull. Torrey Bot. Club 108: 268-270.
Davis, P.H. (1965-1985). Flora of Turkey and the East Aegean Islands. Vol. 1-9. Edinburgh: Edinburgh University Press.
Davis, E.A., Young, J.L. and S.L. Rose (1984). Detection of high-phosphorus tolerant VAM-fungi colonizing hops and pep-permint. Plant Soil 81, 29-36.
Diem, H.G., Gueye, I., Gianinazzi-Pearson, V., Fortin, J.A. and Y.R. Dommergues (1981). Ecology of VA mycorrhizae in the tropics: the semi-arid zone of Senegal. Acta Oecologica-Oecologia Plantarum 2: 53-62.
Dodd, J.C. and J.C. Jeffries (1986). Early development of vesicular-arbuscular mycorrhizas in autumn-sown cereals. Soil Biol. Biochem. 18, 149-154.
Dueck Th, A, Visser, P., Ernst, W.H.O. and H. Schat (1986). Vesicular-arbuscular mycorrhizae decrease zinc toxicity to grasses growing in zinc polluted soil. Soil Biol. Biochem. 18, 331-333.
El-Giahmi, A.A., Nicolson, T.H. and M.J. Daft (1976). Endomy-corrhizal fungi from Libyan soils. Trans. Brit. Mycol. Soc. 67: 164-169.
Gerdemann, J.W. and T.W. Nicolson (1963). Spores of mycorrhizal endogone species extracted from soil by wet-sieving and decanting method. Transact. Brit. Mycol. Soc. 46: 235-245.
Gerschefske Kitt, D., Daniels Hetrick, B.A. and G.W. Thompson Wilson (1987). Sporulation of two vesicular-arbuscular mycorrhizal fungi in nonsterile soil. Mycologia 79, 896-899.
Gildon, A. and P.B. Tinker (1983). Interactions of vesicular-arbus-cular mycorrhizal infection and heavy metals in plants. The effects of heavy metals on the development of vesicular-arbuscular mycorrhizas. New Phytol. 95, 247-261.
Giovanetti, M. and B. Mosse (1980). An evaluation of techniques for measuring vesicular-arbuscular mycorrhizal infection in roots. New Phytol. 84: 489-500.
Grime, J.P., Mackey, J.M., Hillier, S.H. and D.J. Read (1987). Floris-tic diversity in a model system using experimental micro-cosms. Nature, 328, 420-422.
Haas, J.H. and J. Krikun (1985). Efficacy of endomycorrhizal-fun-gus isolates and inoculum quantities required for growth response. New Phytol. 100, 613-621.
Hall, I.R. (1977). Species and mycorrhizal infections of New Zea-land Endogonaceae. Trans. Brit. Mycol. Soc. 68: 341-356.

VAM IDENTIFICATION ON BULBOUS PLANTS 425
Halvorson, W.L. and R.E. Koske (1988). Coastal dune communi-ties. Pp. 80-90. In: Freshwater and marine plants of Rhode Island. Eds., G. S. Robert and M. M. Harlin. Kendall/Hunt-Press, Dubuque, Iowa.
Hayman, D.S. and M. Tavares (1985). Plant growth responses to vesicular-arbuscular mycorrhiza XV. Influence of soil pH on the symbiotic efficiency of different endophytes. New Phytol. 100, 367-379.
Henkel, T.W., Smith, W.K. and M. Christensen (1989). Infectivity and effectivity of indigenous vesicular-arbuscular mycor-rhizal fungi from contiguous soils in southwestern Wyo-ming. USA. New Phytol. 112, 205-214.
Hızalan, E. and H. Ünal (1965). Toprakta Kimyasal Analizler. Ankara Üniv. Ziraat Fak. Yayınları, 273, Ankara.
Hindumathi, A. (1999). Role of arbuscular mycorrhizae in plant growth and biocontrol of charcoal rot in sorghum. [Ph. D. thesis]. Hyderabad, India: Osmania University.
Howeler, R.H., Sieverding, E. and S. Saif (1987). Practical aspects of mycorrhizal technology in some tropical crops and pas-tures. Plant Soil 100, 249-283.
Jackson, M.L. (1973). Soil Chemistry Analysis, Prentice, Hall of India Private Ltd. New Delhi, p. 498.
Janakirani, S. (1991). Studies on Vesicular arbuscular mycorrhizal fungi and plant growth in safflower. [Ph. D. thesis] Hyder-abad, India: Osmania University.
Johnson, P. N. (1976). Effects of soil phosphate level and shade on plant growth and mycorrhizas. N. Z. J. Bot. 14, 333-340.
Karaarslan, E. and R. Uyanöz (2008). Konya Ovası’ndaki Büyük Toprak Gruplarından İzole Edilen Arbusküler Mikoriza (A.M.) Sporlarının Büyüklüklerine Göre Dağılımı, İnfek-siyon Etkinlikleri ve Toprağın Bazı Özellikleri İle Ara-sındaki İlişkiler. S. Ü. Ziraat Fakültesi Dergisi. 22 (45): (2008) 21-28.
Karaarslan, E. and R. Uyanöz (2010). Morphologic Properties and Numbers of Vesicular-Arbuscular Mycorrhizal (V.A.M) Spores Fungi in Major Soil Groups of Konya Basin/Tur-key. International Scientific Conference Devoted To The 65-Th Anniversary Of The U.U. Uspanov Institute of Soil Science and Agrichemistry. pp.455-462. 15-16 September Almaty/ Kazakhstan.
Karaarslan, E. and R. Uyanoz (2011). Occurrence of arbuscular mycorrhizal fungi in some native plants grown on saline soils around the lake Tuz in Turkey and its relations with some physical and chemical properties of soil. Scientific Research and Essays Vol. 6 (20), pp. 4238-4245, 19 Septem-ber, 2011 (Available online at http://www.academicjour-nals.org/SRE. ISSN 1992-2248 ©2011 Academic Journals.)
Khan, A.G. (1974). Vesicular-arbuscular mycorrhizae in plants colonizing black wastes from bituminous coal mining in the Illawarra region of New South Wales. New Phytol. 81: 53-63.
Killham, K. (1985). Vesicular-arbuscular mycorrhizal mediation of trace and minor element uptake in perennial grasses: relation to livestock herbage. In: Ecological Interactions in Soils: Plants, Microbes and Animals (Ed. by A. H. Fitter, D. Atkinson, D. J. Read and M. B. Usher), pp. 225-232. Blackwell, Oxford.
Knudsen, D., Peterson, G.A. and P.F. Pratt (1982). Lithium, sodium and potassium. Pages 225–246 in A. L. Page, R. H. Miller and D. R. Keeney, editors. Methods of soil analysis, part 2. Chemical and microbiological properties. American Soci-ety of Agronomy, Madison, Wisconsin, USA.
Koide, R.T. and H.A. Mooney (1987). Spatial variation in inocu-lum potential of vesicular-arbuscular mycorrhizal fungi caused by formation of gopher mounds. New Phytol. 107: 173-182.
Koomen, I., Grace, C. and D.S. Hayman (1987). Effectiveness of single and multiple mycorrhizal inocula on growth of clo-ver and strawberry plants at two soil pHs. Soil Biol. Bio-chem. 19, 539-544.
Koske, R.E. (1981). A preliminary study of interactions between species of vesicular arbuscular fungi in a sand dune. Trans. Br. Mycol. Soc. 76, 411-416.
Koske, R.E. and C. Walker (1986). Species of Scutellospora (Endo-gonaceae) with Smooth-Walled Spores from Maritime Sand Dunes: Two New Species and A Redescription of the Spores of Scutellospora pellucidia and Scutellospora calos-pora. Mycotaxon 27: 219-235.
Koske, R.E. and J.N. Gemma (1989). A Modified Procedure for Staining Roots to Detect VAM-Mycological Research 92: 486-505.
Koslowsky, S.D. and R.E.J. Boerner (1989). Interactive effects of aluminum, phosphorus and mycorrhizae on growth and nutrient uptake of Panicum virgatum L. (Poaceae). Envir. Pollut. 61, 107-125.
Kriku, J. (1983). Edaphic and climatic factors in the evolution of vesicular-arbuscular mycorrhizal fungi. In: Developments in Ecological and Environmental Quality pp. 31-36. Eco-logical Society, Hebrew University of Jerusalem, Jerusalem.
Lambert, D.H., Cole, Jr. H. and D.E. Baker (1980). Adaptation of vesicular-arbuscular mycorrhizae to edaphic factors. New Phytol., 85:513-520.
Manoharachary, C. and T. Sulochana (1989). Importance of vesicu-lar arbuscular mycorrhiza in sesame seed production. In: Proc. National Seminar on advance in seed science and tech-nology. Shekar Shetty H and Prakash HS (eds.). p. 170-174.
McGee, P.A. (1989). Variation in propagule numbers of vesicular arbuscular mycorrhizal fungi in a semi-arid soil. Mycol. Res. 92: 28-33.
Menge, J.A., Johnson, E.L.V. and R.G. Platt (1978). Mycorrhizal dependency of several citrus cultivars under three nutrient regimes. New Phytologist, 81, 553-559.
Miller, R.M. (1979). Some occurrences of vesicular-arbuscular mycorrhizae in natural and disturbed ecosystems of the Red Desert. Canad. J. Bot. 57: 619-623.
Morton, J.B. (1985). Variation in mycorrhizal and spore mor-phology of Glomus occultum and Glomus diaphanum as influenced by plant host and soil environment. Mycologia 77: 192-204.
Morton, J. (2013). International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. Vest Virginia University. Last modified from http://invam.wvu.edu/the-fungi/clas-sification/glomaceae/glomus

426 Karaarslan et al.
Mosse, B. (1979). Vesicular-arbuscular mycorrhiza research for tropical agriculture. College of Tropical Agric., Univ. of Hawaii, Misc. Publ.
Nicolson, T.H. and J.W. Gerdeman (1968). Mycorrhizal Endogone species. Mycologia 60: 313-325.
Olsen, S.R., Cole, C.V., Watanebe, F.S. and L.A. Dean (1954). Esti-mation of available phosphorus in soils by extraction with sodium bicarbonate. US. Dept. Of Agric. Cric. 939.
Phillips, J.M. and D.S. Hayman (1970). Improved procedures for clearing roots and staining parasitic and vesicular-arbus-cular mycorrhizal fungi for rapid assessment of infection. Trans. Brit. Mycol. Soc., 55: 157-160.
Pond, E.C., Menge, J.A. and W.M. Jarrell (1984). Improved growth of tomato in salinized soil by vesicular-arbuscular mycor-rhizal fungi collected from saline soils. Mycologia 76, 74-84.
Porter, W.M., Robson, A.D. and L.K. Abbott (1987b). Field sur-vey of the distribution of vesicular-arbuscular mycorrhizal fungi in relation to soil pH. J. App. Eco1. 24, 659-662.
Raju, P.S., Clark, R.B., Ellis, J.R. and J.W. Maramille (1990). Effects of species of VA-mycorrhizal fungi on growth and mineral uptake of sorghum at different temperatures. Plant Soil 121, 165-170.
Redhead, J.F. (1977). Endotrophic mycorrhizas in Nigeria: Spe-cies of the Endogonaceae and their distribution. Trans. Brit. Mycol. Soc. 69: 275-280.
Reeves, F.B., Wagner, D., Moorman, T. and J. Kiel (1979). The role of endomycorrhizae in revegetation practices in the semi-arid West. I. A comparison of incidence of mycorrhizae in severely disturbed vs. natural environments. Amer. J. Bot. 66: 6-13.
Richards, L.A. (1954). Diagnosis and improvements saline and alkali soils. U.S. Dep. Agr. Handbook 60, Stroudsburg, U.S.A.
Schenck, N.C. and G.S. Smith (1982). Responses of six species of vesicular-arbuscular mycorrhizal fungi and their effects on soybean at four soil temperatures. New Phyto1. 92, 193-201.
Schenck, N.C. and Y. Perez (1990). A Manual for identification of vesicular arbuscular mycorrhizal fungi in VAM, 3rd Edi-tion. University of Florida, Gainesville, Florida. 286 pp.
Schwab, S. and F.B. Reeves (1981). The role of endomycorrhizae in revegetation practices in the semi-arid West. III. Verti-cal distribution of vesicular-arbuscular (VA) mycorrhiza inoculum potential. Amer. J. Bot. 68: 1293-1297.
Sharif, M. and A.M. Moawad (2006). Arbuscular Mycorrhizal Incidence and Infectivity of Crops in North West Frontier Province of Pakistan World Journal of Agricultural Sciences 2: 123-132.
Sheikh, N.A., Saif, S.E. and Khan, A.G. (1975). Ecology of Endo-gone II. Relationship of Endogone spore population with chemical soil factors. Islamabad J. Sci. 2: 6-9.
Sieverding, E. (1988). Effect of soil temperature on performance of different VA mycorrhizal isolates with cassava. Angew. Botanik 62, 295-300.
Sieverding, E. and T.T. Toro (1988). Influence of soil water regimes on VA mycorrhiza. V. performance of different VAM fungal species with cassava. J. Agron. Crop Sci. 161, 322-332.
Simpson, D. and M.J. Daft (1990a). Interactions between water-stress and different mycorrhizal inocula on plant growth and mycorrhizal development in maize and sorghum. Plant Soil 121, 179-186.
Soltanpour, P.N. and S.M. Workman (1981). Use of inductively-coupled plasma spectroscopy for the simultaneous deter-mination of macro and micro nutrients in NH
4HCO
3-
DTPA extracts of soils. In Barnes R.M. (ed). Developments in Atomic Plasma Analysis, pp. 673-680, USA.
Stahl, P.D. and W.K. Smith (1984). Effects of different geographic isolates of Glomus on the water relations of Agropyron smithii. Mycologia 76, 261-26Subha, P. (2001). The role of AM fungi and PGPR on growth and metabolism of Ipomoea batatus Lam. [Ph. D. thesis]. Hyderabad, India: Osmania University.
Sutton, J.C. and G.L. Barron, (1972). Population dynamics of Endogone spores in soil. Canad. J. Bot. 50: 1909-1914.
Sykorová, Z. (2007). Molecular ecological analyses of specific interactions between symbionts in the arbuscular mycor-rhizal symbiosis. Inauguraldissertation Erlangung der Würde eines Doktors der Philosophie vorgelegt der Philos-ophisch-Naturwissenschaftlichen Fakultät der Universität Basel aus Pardubice, Tschechische Republik. pp. 18. Basel.
Sylvia, D.M. and S.E. Williams (1992). Vesicular-Arbuscular Mycorrhizae and Environmental Stresses. Pages 101-123. In: Mycorrhizae in Sustainable Agriculture. Edited by G.J. Bothlenfalvay and R.G. Linderman. American Society of Agronomy, Inc. Madison, Wisconsin, USA.
Thomson, B.D., Robson, A.D. and L.K. Abbott (1986). Effects of phosphorus on the formation of mycorrhizas by Gigaspora calospora and Glomus fasciculatum in relation to root car-bohydrates. New Phytol. 103, 751-765.
Toroğlu, E. and Ü.E. Ünaldı (2008). Ecologic Conditions of Vege-tation on the Aladaglar (Taurus Mountain) Fırat University Journal of Social Science 18 (2): 23-48 ELAZIĞ.
Uyanöz, R. and E. Karaarslan (2012). Arbuscular Mycorrhizal Fungi for Some Wild Plants in Saline Soils of Central Anatolian Region-Turkey. International Conference of Ecosystems (ICE) Proceeding Book. pp.515-521. Essays on Ecosystem and Environmental Research. 1-6, June, Tirana, Albania.
Van Der Heijden, M., Klironomos, J., Ursic, M., Moutoglis, P., Streitwolf-Engel, R., Boller, T., Wiemken, A. and I. Sand-ers (1998). Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity. Nature, 396: 69-72.
Vani, P. (1993). Studies on vesicular arbuscular mycorrhizal fungi in relation to the growth and biomass of castor. [Ph.D the-sis]. Hyderabad, India: Osmania University.
Zajicek, J.M., Daniels-Hetrick, B.A. and C.E. Owensby (1986). The influence of soil depth on mycorrhizal colonization of forbs in the tallgrass prairie. Mycologia 78: 316-320.





























