Embed Size (px)
Citation preview

Modern foraminifera from coastal settings, northern Argentina 45
ModernforaminiferafromcoastalsettingsinnorthernArgentina:implicationsforthepaleoenvironmentalinterpretationof
MidHolocenelittoraldeposits
Cecilia Laprida1,*, Damián Diego Enrique Chandler1,2, Josefina Ramón Mercau1, Rubén Alvaro López1 y Silvia Marcomini1
1 Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Argentina.
3 Consejo Nacional de Investigaciones Científicas y Técnicas, Rivadavia 1917, Ciudad Autónoma de Buenos Aires, Argentina
ABSTRACT
In Buenos Aires coasts, sedimentary processes were particularly active during the Quaternary owing to eustatic fluctuations in sea level. As a result, during the late Quaternary transgressions, marine and marginal marine sediments were deposited in the coastal plain. In order to interpret these Holocene littoral sequences, we analyzed the distribution, diversity, species composition and taphonomic features of total benthic foraminifera assemblages from modern littoral settings, from the top of the dune to the lower shoreface, in two close but geomorphologically different transects located in the Atlantic coast of Northern Buenos Aires Province (Argentina, South America). Total benthic foraminiferal assemblages from subtidal and supratidal environments are distinguishable in terms of composition, diversity and taphonomic features. In upper shoreface, foreshore, backshore and foredune environments, assemblages are clearly dominated by three species: Buccella peruviana, Ammonia beccarii and Elphidium discoidale (the BAE group). This feature is the result of taphonomic processes that favor the selective preservation of such species. The study of taphonomic modifications of shells in modern assemblages allows a better discrimination between subenvironments than the analysis of taxonomic composition. Although Holocene assemblages have no strict counterparts between total modern assemblages, taxonomic composition and taphonomic modification of shells allow us to infer that the Holocene sequence was deposited between the upper shoreface and the backshore.
Key words: benthic foraminifera, modern beaches, paleoenvironmental reconstruction, Holocene, Argentina.
RESUMEN
En las costas de la provincia de Buenos Aires, los procesos sedimentarios fueron especialmente activos durante el Cuaternario, debido a las fluctuaciones eustáticas en el nivel del mar. Como resultado, durante las transgresiones del Cuaternario tardío se depositaron en la planicie costera sedimentos marinos y marino-marginales. Con el objetivo de interpretar estas secuencias litorales holocenas, se analizó la
Revista Mexicana de Ciencias Geológicas, v. 28, núm. 1, 2011, p. 45-64
Laprida,C.,Chandler,D.D.E.,RamónMercau,J.,López,R.A.,Marcomini,S.,2011,ModernforaminiferafromcoastalsettingsinnorthernArgentina:implicationsforthepaleoenvironmentalinterpretationofMidHolocenelittoraldeposits:RevistaMexicanadeCienciasGeológicas,v.28,núm.1,p.45-64

Laprida et al.46
distribución, diversidad, composición específica y características tafonómicas de las asociaciones totales de foraminíferos bentónicos en ambientes litorales actuales, desde el tope de la duna hasta la playa, en dos transectas cercanas pero geomorfológicamente diferentes, ubicadas en las costas atlánticas del norte de la Provincia de Buenos Aires (Argentina, Sudamérica). Las asociaciones totales de ambientes submareales y supramareales son diferenciables en términos de composición específica, diversidad y características tafonómicas. En los ambientes de playa distal, playa frontal, cara de playa y duna, las asociaciones están dominadas por tres especies: Buccella peruviana, Ammonia beccarii y Elphidium discoidale (grupo BAE). Esta dominancia es el resultado de procesos tafonómicos que favorecen la preservación selectiva de dichas especies. Las características tafonómicas de las asociaciones modernas permiten una mejor discriminación de los subambientes que la composición taxonómica. Si bien las asociaciones holocenas no poseen homólogos estrictos entre las asociaciones totales de ambientes actuales, la composición taxonómica y los rasgos tafonómicos de las asociaciones permiten inferir que la sucesión holocena se depositó entre la parte superior de la anteplaya y la playa distal.
Palabras clave: foraminíferos bentónicos, playas actuales, reconstrucción paleoambiental, Holoceno, Argentina.
INTRODUCTION
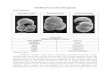
In the low-relief northernBuenosAires coasts(Argentina,southwesternSouthAtlantic),sedimentaryprocesseswereparticularlyactiveduringtheQuaternaryowingtoeustaticfluctuationsinsealevel.Asaresult,marineandmarginalmarinesedimentsweredepositedinthecoastalplainespeciallyduringthelateQuaternarytransgressions.Inthecoastalplainofthelow,gentlyslopingSamborombónBayarea,theHolocenetransgressioniswellpreserved.There,thepostglacialseareacheditsmaximumlevelat+4–6mabovesealevel(asl)ataround6000yearBP,andmiddletolateHolocenecoastalsedimentationwasparticularlyeffective.Theareacomprisesprogradingandregressivemarinesequencesdepositedinshallowen-vironmentscorrespondingtobarrierislands,saltmarshesandbeaches(CodignottoandAguirre,1993;AguirreandWhatley,1995).IncentralSamborombónBay,Holocenebarrierislandcomplexesarerepresentedmainlybysandyandshellydepositsupto+4.5–5masl.Thesebarrierislandcomplexesincludebarrierislandsandbeachridges,andconstitutetheCerrodelaGloriaMemberoftheLasEscobasFormation.DetailedsedimentologicalandpaleoecologicalanalyseshaveledsomeauthorstopostulatethattheCerrodelaGloriaMemberwasdepositedinashallowinfralittoraltointertidalhigh-energy,soft-bottomenvironment(Spallettiet al.,1987;CodignottoandAguirre,1993).Evenwhenitisclearthatthesedepositswereoriginatedunderacom-binationofwaves,coastalcurrentsandtides(CodignottoandAguirre,1993),theidentityofthedrivingprocessanditspreciselocationwithinthebeacharestilldoubtful.Didthedepositsformasshoalsorlongshorebars?Didtheyformasbeachbermsoriginatedbyswashduringmoderate(non-storm)waveconditionsinintertidalsettings?Weretheyoriginatedintheemergedbeachduringextraordinarystormyconditions?
Theobjectiveofthisstudyistoprovidefurtherin-sightintotheoriginoftheHoloceneshellybeachridgespresentincentralSamborombónBay,northernBuenos
Airescoastalmargin,byconsideringmicropaleontologi-calandtaphonomicdata,namelythespeciescompositionandpreservationstateofbenthicforaminiferfossilassem-blages.TheforaminiferahavethepotentialtobeusedasproxiesofenvironmentalfactorsandthereforeareusefulinpaleoenvironmentalreconstructionsincludingHolocenecoastalsandbodies(Rineet al.,1991;Murray-Wallaceet al.,1999;HippensteelandMartin,1999).However,theirproxyvaluedependsontheextenttowhichtheirmodernecologicalrelationshipscanbeextrapolatedtointerpretthefossilrecordinaparticulararea(DizandFrances,2009).TheoccurrenceofbenthicforaminiferainHolocenemarinesedimentsofthenorthernBuenosAirescoastalplainisrelativelywelldocumented(Laprida,1997,1999;LapridaandBertels-Psotka,2003),butmodernlittoralassemblageshavereceivedlittleattention.Themostdetailedstudiespub-lishedtodatearethoseofBoltovskoy(1970,1976),whichdealwithdistributionoflittoralforaminiferainAgentina,UrurguayandSouthernBrazil,buttheauthorfailstotakequantitativeanalysisofthemicrofaunaintoaccount.
Accurateapplicationofforaminiferatoresolvepa-leoenvironmentalproblemsrequiresataphonomicevalu-ationoffossilassemblages(Barbieri,1996;GoldsteinandWatkins,1999)sincebiostratinomic/taphonomicdatareflect conditions of depositional environments, represent-ingavaluableadditionalsourceofinformationoncoastaldynamics(Brandt,1989;LapridaandBertels-Psotka,2003).Taphonomicsignatures(sensuFürsichandOschmann,1993)mayvaryinapredictablefashionalonganenviron-mentalgradient(e.g.,shallowtodeepwater)andhencetaphonomicdatacanbepowerfultoolsinpaleoenviron-mentalstudiesespeciallywhensedimentologicdataarelackingordubious.
Inthisstudywecharacterizetotal(live+dead)as-semblages recovered from superficial sediments of selected environmentsofnorthernBuenosAiresAtlanticlittoralfrombothataxonomicandataphonomicpointofview,andwecomparethemwithfossilassemblagesrecoveredfromamid-Holoceneshellybeachridgebelongingtothe

Modern foraminifera from coastal settings, northern Argentina 47
URUGUAY
de la Plata river
Punta Rasa
Punta Médanos
Mar del Plata57º W
57º W
36ºS36ºS
BuenosAires
Coastal plains
ARGENTINA
AtlanticOcean N
Punta Piedras
Mar de Ajó
Km187
**
Atlan
ticOc
ean
Samborombón bay
0 50 km
siliciclasticsandbeds(Laprida,1997;LapridaandBertels-Psotka,2003).Thesedepositswereinterpretedasbeachridgesandbarrierislandsrelatedtohighenergycoastalenvironments(TonniandFidalgo,1978;Bertels-PsotkaandLaprida,1998).Tothesouth,betweenPuntaRasaandPuntaMédanos,mid-HolocenecoastalmarinesedimentsarerepresentedbytheMardeAjóMemberofthePozoN°17Formation(Parker,1979).
ThemodernlittoralareaadjacenttotheHolocenede-posits corresponds to tidal flats in the northern Samborombón Bayarea–betweenPuntaPiedrasandPuntaRasa–andtosandybeachesoftheArgentineSea,southofPuntaRasa.BuenosAirescoastsbetweenPuntaRasaandPuntaMédanos(Figure1)areexposedtowaveaction,andtheyarestormandwavedominated.Itisanopencoast,straight,withNorth-Southshorelineorientation.PrevailingdirectionsofincidentwavesarefromtheSandSSE,producinganorth-wardlittoraldriftcurrent.Thetidalregimeissemidiurnalwithdiurnalinequalitieswithmeanamplituderangingfrom1.37m(springtides)to0.78m(neaptides)(ServiciodeHidrografíaNaval,2001).Extremevaluesrangefrom0to2.40mforhightidesandfrom0.40to1.60mforlowtides(Perillo,1979).Whenoccurringjointlywithspringtides,stormsreachthedunebaseandcauseerosionalescarpments.Thebeachvariesinwidthfrom40to140mandhasameanslopeof2°.Thebeachcorrespondstotheintermediatetypewithbarsandchannels(Sunamura,1988),wherestableandnonstable(seasonal)bermswereobserved.Swashbarsarefrequentlyobservedattheforeshore,andrangefrom15to25minwidthand0.25to0.50minheight.Twobreakerlinescanbedistinguishednexttotheshore,between40and100mandbetween140and160mfromthemeantideline,respectively.Themeanwaveheightis0.70m,whilethemaximumwaveheightis2m.Themeanwaveperiodis8.4seconds(Speroniet al.,1999).
MATERIALS AND METHODS
Formodernforaminiferalanalysis,twentysixsuper-ficial sediment samples were collected from two different littorallocalitiesofMardeAjó,MunicipalityofLaCosta(Figure1),during(austral)summerof2002and2007.Samplesweretakenalongmeasuredwest-easttransectsperpendiculartothecoastlineandspacedabout2km,at36º43’44.1”S-56º40’24.4”W(Southerntransect,12samples)andat36º42’37.4”S-56º40’30.7”W(Northerntransect, 14 samples). Transects were defined from the top oftheduneintheforedunetothelowershorefaceataround-10mdepth.Sampleswerecollectedmanuallywhilewalk-ingandwadingalongtheintertidalzoneandthebackshore.AVanVeengrabsamplerwasusedtorecoversedimentsatthesediment/waterinterfaceinsubtidalsettings.Fromeachsample,onlytheuppermostlayerofthesediment(about5-10cm,accordingtoDenneandSenGupta,1989)wasscrapedoffandkeptinalcohol70%.Asolutionof
CerrodelaGloriaMemberofLasEscobasFormation,southernSamborombónBay.Totalpopulationspresentamorehomogeneousspatialdistributioncomparedtolivingonesasaconsequenceoflateralandverticalmixingoftestsbybiostratinomicprocesses(ScottandMedioli,1980;Scottet al.,2001),andhencetheyaremoreadequatetocomparewithfossilassemblages.Consequently,thepresentstudyprovidesinsightnotonlyintothepresentdistributionoftotallittoralforaminiferalassemblagesofnortheasternBuenosAiresProvincebutalsointotheoriginandevolutionofmid-Holocenebeachridgesofthebonaerensiancoastalplain.
STUDY AREA
ThestudyareaislocatedalongtheAtlanticOcean,intheeasterncoastalplainofnortheasternBuenosAiresprovince,Argentina(Figure1).Thiscoastalareaisanaccresive coast characterized by superficial mid-Holocene marinesedimentsoverlyingPliocenetolatePleistocene“pampean sediments” (Fidalgo et al., 1975). In theSamborombónBayarea,mid-HolocenesedimentsarerepresentedbytheLasEscobasFormation(Fidagoet al.,1975),whichincludestwomembers:CerrodelaGloriaandCanal18.TheCerrodelaGloriaMembercomprisesawell-stratified succession of white, fine to coarse-grained shelly gravels and light brown, fine-grained siliciclastic sands(Spallettiet al.,1987).Thegravelfractionismainlycomposedofunarticulatedandbrokenbivalveshells.Calcareousmicrofossils,especiallybenthicforaminiferaandostracods,areabundantinboththeshellygraveland
Figure1.Locationmapofthestudyarea.

Laprida et al.48
2ppmRoseBengalinethanolwasusedforstaininglivespecimens.
Thenortherntransect(Table1)lacksseasonalberms,butastablebermapproximately30minwidthand0.25minheightisobserved.Thebeachwidthvariesbetween90 and 140 m and the average grain size is medium to fine sand,whileintheintertidalsectortheaverageismediumtocoarsesand.Inthesoutherntransect(Table1)nostableorseasonalbermsareobserved.Thebeachwidthvariesbetween55and65mandtheaveragegrainsizeismediumto fine sand.
ForHoloceneforaminiferalanalysis,twentyninesedi-ments samples were collected from an outcrop, an artificial quarryof3.90mlocatedattheleftmarginoftheprovincialRoute N° 11, in the milestone numbered km 187 (Profile KM187).TheoutcropbelongstotheCerrodelaGloriaMemberofLasEscobasFormation(Fidalgoet al.,1975).Twentyninesamplesof500gweretakenfromeachidenti-fied bed (Figure 2) irrespective of its grain size, geometry and internal structure. KM187 profile comprises a well stratified succession of bioclastic deposits with subordinated light brown siliciclastic sands, which are well defined by
sharpchangesingrainsize.Bioclastsaremainlycomposedofunarticulatedandfragmentedshellsofbivalvesandpebbles of caliche. Subhorizontal stratification is dominant in shelly gravels, whereas hummocky and trough stratifica-tionarealsopresent,thelatterasisolatedsetsintercalatedin the subhorizontal stratificated facies.
Modernandfossilsedimentsampleswerewetwashedinthelaboratorythrougha0.075mmsieve.Foraminiferswereseparatedfromthesand-sizedfractionunderastereo-microscope.Onthebasisofpreviousexperience(Laprida,1997,1999;LapridaandBertelsPsotka,2003),aprox.7gofdrysedimentforeachsamplewereanalyzedorap-proximately300individualswerepicked,whichevercamefirst. According to Phleger (1960), Buzas (1990) and Scott et al. (2001), this amount provides sufficient accuracy for mostquantitativeexaminations.GenericassignmentoftaxafollowsLoeblichandTappan(1988),whilespeciesidentification follows Boltovskoy (1954a, 1954b, 1957, 1958),Boltovskoyet al.(1980)andLaprida(1997,1999).Taphonomic modification of tests in both modern and fossil assemblageswereobservedinordertoobtainadditional(paleo)environmentalinformation.Inordertoavoidso-
Sample Nº Distance from swash zone (m)
Height above sea level (m)
Sediment compositionSubenvironment
Silt-clay Sand Gravel
Northern transectNST-1 1700 -10.5 96.67 3.33 0.00 ShorefaceNST-2 1500 -9 87.41 12.59 0.00 ShorefaceNST-3 1300 -7.5 76.07 23.93 0.00 ShorefaceNST-4 837 -7 51.85 48.15 0.00 ShorefaceNST-5 564 -5.5 10.42 89.58 0.00 ShorefaceNST-6 379 -4.5 8.27 91.73 0.00 ShorefaceNST-7 50 -1.2 0.06 99.09 0.85 ShorefaceNST-8 30 -1.5 0.04 99.66 0.30 ShorefaceNET-1 0 0 0.01 95.55 4.44 ForeshoreNET-2 -35 0.5 0.04 99.96 0.00 ForeshoreNET-3 -59 1 0.02 99.98 0.00 ForeshoreNET-4 -84 1.5 0.06 99.94 0.00 BackshoreNET-5 -109 1.3 0.01 99.99 0.00 BackshoreNET-6 -131 3.7 0.01 99.99 0.00 Foredune
Southern transect
SST-1 1616 -9.3 6.67 93.33 0.00 ShorefaceSST-2 1415 -8.7 41.30 58.70 0.00 ShorefaceSST-3 1237 -8.3 93.73 6.27 0.00 ShorefaceSST-4 1041 -8.1 49.29 50.71 0.00 ShorefaceSST-5 490 -4.7 12.82 87.1 0.00 ShorefaceSST-6 314 -3.85 43.20 56.80 0.00 ShorefaceSST-7 162 -1.95 9.05 90.95 0.00 ShorefaceSET-1 0 0 0.01 97.81 2.18 ForeshoreSET-2 -15 0.25 0.01 99.99 0.00 ForeshoreSET-3 -31 0.6 0.02 99.98 0.00 BackshoreSET-4 -65 1 0.05 99.95 0.00 BackshoreSET-5 -67 3.5 0.17 99.83 0.00 Foredune
Table1.Sedimentologicalcharacteristics,locationandsubenvironmentsofsamplesfromthenorthernandsoutherntransects.

Modern foraminifera from coastal settings, northern Argentina 49
darkbrown;d)abradedandscratchedshells;ande)brokenandsometimesfracturedshells.Fragmentedshellsdenoteshellswithlessthan50%ofitsoriginalshellmaterialpre-served.Abradedshellsincludeerodedshellswithpolishedmorphologicalfeaturesandintenselyeroded,completelyabradedshells.Bothabrasion(physicalagents)anddis-solution(chemical/biologicalprocesses)cancreatesimilartaphonomicsignatures,anddistinguishingbetweenthemisusuallynotpossible.Therefore,abrasionandfragmentationwereestablishedbytallyingbreaksindependentoftheirorigin(physical,chemicaland/orbiological).
Abundanceoftestsofmodernandfossilassemblageswascalculatedfromtheextrapolationofthenumberofindi-vidualspickedoutofeachsampleto10gofdry,untreatedsediment.MultivariatestatisticalanalyseswereappliedtobothsetsofsamplesusingPASTsoftware(version1.71)(Hammeret al.,2001)inordertoidentifypatternsandcor-relationsamongthedifferentparametersmeasured(speciescomposition, taphonomical modifications of tests, water depthandgrainsize).Rarespeciesthatoccurat<1%ofthewholeassemblagewereexcludedfromthestatisticalanalysis.SampleKM187-1,whichonlyyieldedtwoin-dividualsofBuccella peruviana (D´Orbigny,1839a),wasexcludedfromthedatabasebecauseofitslittlereliability.Anunconstrainedclusteranalysisbasedontheunweightedpair-groupaveragealgorithmandtheBray-Curtissimilar-ityindexwasperformedonthetaxonomiccomposition(speciesrelativecontribution)ofmodernandfossilas-semblagesconsideredjointly.Likewise,anunconstrainedclusteranalysisbasedontheunweightedpair-groupaveragealgorithmandtheEuclideandistancesimilarityindexwasperformedonthetaphonomicsignaturesofthewholesetof samples. Clusters were defined by the branches cut by alinedrawnperpendicularlytothemid-lengthofthelon-gestdistancebetweensuccessivenodesofthedendogram.Whenanodefellveryclosetothisline,itwasredrawnsoastoincludebothgroups,sinceinviewofourobjectivesandthenatureofthisinvestigationunderestimationduetothelossofinformationwasdeemedworsethanoveresti-mationduetoacceptanceofasomewhatfeeblebut,inourunderstanding,meaningfulgrouping.Atotalofsixlineardiscriminantanalysis(LDA)wereperformedonbothsetsofrecentsamples,inordertoevaluatewhethersubtidalandsupratidalbeachsamples,andnorthernandsoutherntran-sectsamples,couldbedistinguishedfromeachotherwithrespecttotheirspeciescomposition,taphonomicsignaturesandgrainsize.Aleave-one-outvalidationwasperformedoneachanalysisbyexcludingeachsampleinturnfromthecalculationofthediscriminantfunctionandclassifyingwiththeresultingalgorithm.Finally,thecorrelationbetweengrainsize,preservationalstateandspeciescompositionineachtransectwasassessedbymeansoftheManteltestinordertoevaluatethepossibilitythatbothsiteswereundergo-ingdifferentprocesses.ThespeciescompositionsimilaritymatriceswerecomputedwiththeBray-Curtisindex,whiletheotherswerecalculatedusingtheEuclideandistanceas
Figure 2. Scheme of the profile KM187 (Cerro de la Gloria Member, Las EscobasFormation,BuenosAiresProvince),andlocationofthesamplesstudied.
calledobservererror(Rothfus,2004),taphonomicobser-vationsweredonebythesameoperator,usingastereomi-croscope at 100x magnification. Taphonomic modifications weredeterminedforeachspecimenobserved.Accordingtotype and degree of taphonomic modification, five categories were defined: a) Well preserved shells, lacking at the most thelast(morefragile)chambers;b)partiallyortotallyrecrystallized shells; c) “black” shells with chamber fill-ingsand/orwithapolishedcoatinginshadesofblackand

Laprida et al.50
asimilarityindex.SincethreeManteltestswereperformedoneachsetofsamples,aBonferronicorrectionwasappliedto the significance level, so that the p-valueofeachtestwascompared with an adjusted alpha αad = α / 3 = 0.017. PAST softwarewasalsousedtocalculateecologicalparameterssuchasspeciesnumber(S)andShannon-Weaverdiversityindex(H)inbothmodernandfossilassemblages.
Sampleswerenamedaccordingto:NST(NorthernSubmergedTransect)forsubmerged(subtidal)sitesoftheNortherntransect;NET(NorthernEmergedTransect)foremerged(intertidalandsupratidal)sitesoftheNortherntran-sect;SST(SouthernSubmergedTransect)forsubmerged(subtidal)sitesoftheSoutherntransect;SET(SouthernEmergedTransect)foremerged(intertidalandsupratidal)sites of Southern transect; and KM187 for KM187 profile. Ordinalsindicaterelativepositionineachtransectandstratigraphical level in the Mid-Holocene profile.
RESULTS
Foraminifera from modern environments
Allsamplesborebenthicforaminifera,eventhoseoftheforedune(baseandtopofthedune).Atotalof78foraminiferspeciesand13generaleftinopennomencla-ture were identified, of which only 13 species showed a relativeabundancehigherthan1%(Table2).Thenumberoftestsin10gofsedimentvariedfromabout95to3,559,thenumberofspeciesineachsamplevariedfrom5to38(Table3),andtheShannon-Weaverdiversityindex,H(S),variedbetween2.67and0.85.AssemblagesintheNortherntransecttendtobemorediverseandabundantthanintheSoutherntransect.AsBoltovskoy(1970)statedpreviouslyforlittoralsettings,veryfewlivingspecimenswerepresentanditwasimpossibletousethemforstatisticalanalysis.Thus,thefollowingstudyisbasedontotal(live+dead)as-semblages.Hyalinecalcareousspeciesaredominant.Eventhoughthemajorityoftestsaremoderatelywellpreserved,someshoweddifferenttypesanddegreesofalteration.Insome samples several unidentifiable quinqueloculinid mili-olidsweremostlybrokenorfragmented,andhencewerelumpedinQuinqueloculinaspp..
Modernlittoralsamplesaredominatedbyafewpro-lific species of benthic foraminifera, particularly Buccella peruviana,Quinqueloculina seminula (Linné,1758),Ammonia beccarii(D´Orbigny,1839a),Discorbis william-soni(ChapmanandParr,inParr,1932),andElphidium dis-coidale(D´Orbigny,1839a).Speciesthatarelesscommon,butareneverthelessconsistentelementsofthefauna,includeQuinqueloculina patagonica D´Orbigny,1839b,Textularia gramenD´Orbigny,1846,Elphidium gunteriCole,1931,Quinqueloculina milleti (Wiesner,1912),Quinqueloculinasp.cf.Q. implexaTerquem,inTerquemandTerquem,1886,Trochammina ochracea(Williamson,1858),Miliolinella subrotunda(Montagu,1803)andCibicidoidessp.cf.C.
fletcheri(GallowayandWissler,1927).Bolivina striatulaCushman,1922islocallyabundantintheSoutherntransectin samples adjacent to the shore and confined to subtidal settings.
Table3showsthedataobtainedfor theNorherntransect.Thebenthicforaminiferalnumber(specimens/10gdrysediment),speciesnumberandtheShannon-Weaverdiversity indexH(S)arerelatedwithwaterdepthanddecreaseonshore.Lowernumberofspecimens,speciesanddiversityoccuratthebackshoreandtheforedune.Consideringthe totalnumberofspeciesandthe totalassemblages identified in theNorthern transect,onlyBuccella peruviana,Ammonia becariiandElphidium discoidale(theBAEgroup)canbeconsideredaswidelydistributedalongthetransect,withBuccella peruvianabeingbyfarthedominantspecies(relativecontributionbetween63–36%).Theonlyexceptionisalowershorefacesamplesbelow-7.5mwhereQuinqueloculina seminuladominates,accountingfor21–35%oftheassemblages.Discorbis williamsoniisdistributedinallsubtidalsamplesbutitisvirtuallyabsentinsupratidalsamples.Ammonia beccariiandElphidium discoidale,whicharealsodistributedthroughout,becomedominantonlyintheuppershorefaceandinthebackshoreandforedune,wheretheBAEgrouprepresentsmorethan50%ofassemblages.SpeciessuchasTrochammina ochracea,Quinqueloculina millettiandQuinqueloculinasp.cfQ. implexaarelocallyabundantinwaterdepthgreaterthan-7mbuttheycanbecategorizedassporadic,andtheydonotrepresentmorethan12%ofeachassemblage.
Thefollowingspeciesareclassifiedasaccessoryspecies:Quinqueloculina patagonica,Quinqueloculina milletti,Elphidium gunteri,Cibicidoidessp.cf.C. fletcheriandMiliolinella subrotunda.Otherspeciesoccurinrela-tiveabundancelowerthan1%oftheassemblageandareconsideredtoberareoraccidentalspecies.
Taphonomic modifications of shells in subtidal settings aredirectlyrelatedwithwaterdepth(Figure3a).Wellpreservedtestsrepresentmorethan70%ofassemblagesinthelowershoreface,andvarybetween62–42%intheuppershoreface.Fragmentedtestsvarybetween30–18%inthelowershorefaceandincreaseto36–54%intheuppershoreface.Intheforeshore,fragmentedshellsprevail(51–56%)andabrasionandrecrystalizationarelow(2–6%).Inthebackshore,wellpreservedtestsvarybetween38–58%,andfragmentationisintense(26–50%).Abradedandrecrystalizedshellsarerelativelymoreabundantinthebackshore,especiallyinthebaseofthedune,whereupto9%oftheshellsarerecrystalized,andatthetopofthedune,whereabradedtestsaccountfor12%oftheassemblage.Thenumberoftestsleftinopennomenclatureduetotaphonomicmodification varies between 8 % and 27 %.
Tables2and4showsthedataconcerningtheSoutherntransect.Highestspeciesnumber,abundanceanddiversityarefoundintheshorefaceanddecreaseintheforeshore,andarelowestinthebackshoreandtheforedune.Considering

Modern foraminifera from coastal settings, northern Argentina 51
Species
Sample/ level A
mm
onia
bec
cari
i
Buc
cella
pe
ruvi
ana
Cib
icid
oide
s sp.
cf.
C. fl
etch
eri
Dis
corb
is
will
iam
soni
Elp
hidi
um
disc
oida
le
Elp
hidi
um g
unte
ri
Mili
olin
ella
su
brot
unda
Qui
nque
locu
lina
mill
etti
Qui
nque
locu
lina
pata
goni
ca
Qui
nque
locu
lina
sem
inul
a
Qui
nque
locu
lina
sp. c
f. Q
. im
plex
a
Text
ular
ia g
ram
en
Troc
ham
min
a oc
hrac
ea
NST-1 11 29 1 9 1 - 8 33 21 112 22 - 34NST-2 11 35 1 12 2 - 5 1 8 73 3 - 4NST-3 12 65 - 21 2 3 2 17 13 71 19 - 1NST-4 13 163 - 27 5 4 4 5 2 25 1 - -NST-5 22 192 7 17 27 5 - - 4 11 - 5 -NST-6 23 218 10 14 24 7 4 1 4 7 - 1 -NST-7 19 76 2 3 13 6 1 - - 6 8 3 -NST-8 31 218 7 9 28 15 3 - 12 9 1 - -NET-1 10 34 1 2 6 - - - 1 4 - 3 -NET-2 10 34 1 2 7 2 - - 4 2 - - -NET-3 8 40 1 1 7 3 - - - 3 - - -NET-4 6 48 1 2 5 2 - - 1 1 - - -NET-5 16 59 - - 4 4 - - 1 - - - -NET-6 6 29 1 - 2 2 - - 1 1 - - -SST-1 67 23 - 5 7 2 - - 4 11 - - 3SST-2 15 46 1 7 2 4 - - - 5 - - 7SST-3 37 36 - 3 8 1 - 1 5 12 - - 2SST-4 20 140 - 18 5 2 3 5 7 39 2 - -SST-5 3 54 - 24 6 - 16 2 6 22 5 47 1SST-6 1 46 3 16 9 1 1 - 1 5 - 14 -SST-7 - 16 5 3 4 - - - - 2 - 2 -SET-1 4 25 2 - - 1 - - 2 2 - - -SET-2 11 54 2 1 7 - 1 - 1 2 - 3 -SET-3 6 32 - - 1 3 - - - 1 - - -SET-4 4 37 - - 3 4 - - - 1 - - -SET-5 6 34 - - 3 - - - 2 1 - - -KM187-1 - 2 - - - - - - - - - - -KM187-2 28 150 2 - 27 - - - - - - - -KM187-3 142 242 - - 89 8 - - - - - - -KM187-4 62 112 - - 26 1 - - - 1 - - -KM187-5 69 131 - - 42 3 - - - - - - -KM187-6 73 76 - - 56 5 - - 1 - - - -KM187-7 19 22 - - 8 1 - - - - - - -KM187-8 70 109 - - 60 4 - - - 4 - - -KM187-9 56 60 - - 118 26 - - - - - - -KM187-10 73 85 - - 107 10 - - - - - - -KM187-11 72 75 2 - 93 9 - - - - - - -KM187-12 98 162 - - 30 3 - - - 2 - - -KM187-13 86 150 - - 41 - - - - 2 - - -KM187-14 42 124 1 - 38 2 - - - - - - -KM187-15 96 74 - - 88 - - - - - - - -KM187-16 47 195 1 1 60 4 - - - 4 - - -KM187-17 43 140 - - 50 4 - - - - - - -KM187-18 56 132 - - 37 - - - - - - - -KM187-19 35 60 1 - 35 3 - - - - - - -KM187-20 59 107 - - 34 3 - - - - - - -KM187-21 55 80 1 - 50 6 - - - - - - -KM187-22 56 198 - - 77 3 - - 1 - - - -KM187-23 40 88 - - 92 4 - - - - - - -KM187-24 60 151 1 - 74 6 - - - - - - -KM187-25 33 127 - - 90 - - - - - - - -KM187-26 26 196 - - 88 4 - - 1 4 - - -KM187-27 43 163 - - 94 3 - - - - - - -KM187-28 - 160 1 - 88 - - - - 3 - - -KM187-29 41 189 - - 120 4 - - - 12 - - -
Table2.Distributionchartofspecieswitharelativeabundancehigherthan1%inmodernandHoloceneenvironments.

Laprida et al.52
Sample Principal species S Abundance (ind./10g) H (S) Indeterminable
individuals (%)Taphonomic condition (%)
WP R A F F&C
NST-1 Quinqueloculina seminula:26.48%Trochammina ochracea:8.04%Quinqueloculina milletti:7.80%Buccella peruviana:6.86%
26 3559 2.25 26.71 69.98 0.00 0.00 30.02 0.00
NST-2 Quinqueloculina seminula:35.44%Buccella peruviana:16.99%Discorbis williamsoni:5.83%
28 418 2.28 8.74 81.55 0.00 0.00 18.45 0.00
NST-3 Quinqueloculina seminula:21.26%Buccella peruviana:19.46%Discorbis williamsoni:6.29%Quinqueloculina sp. cf. Q. implexa:5.69%
25 1550 2.25 23.65 62.87 0.00 0.30 36.83 0.00
NST-4 Buccella peruviana:53.44%Discorbis williamsoni:8.85%Quinqueloculina seminula:5.20%
16 6734 1.54 12.13 58.69 0.33 0.00 40.98 0.00
NST-5 Buccella peruviana:55.49%Elphidium discoidale:7.80%Ammonia beccarii:6.36%Discorbis williamsoni:4.91%
16 3130 1.40 14.16 56.06 0.58 0.87 42.49 0.00
NST-6 Buccella peruviana:60.06%Elphidium discoidale:6.61%Ammonia beccarii:6.34%
18 2101 1.33 11.02 60.05 0.28 0.83 38.84 0.00
NST-7 Buccella peruviana:44.71%Ammonia beccarii:11.18%Elphidium discoidale:7.65%
19 305 1.84 12.94 51.19 1.76 1.76 45.29 0.00
NST-8 Buccella peruviana:49.66%Ammonia beccarii:7.06%Elphidium discoidale:6.38%
19 899 1.51 20.73 42.60 1.82 1.14 54.44 0.00
NET-1 Buccella peruviana:36.17%Ammonia beccarii:10.64%Elphidium discoidale:6.38%
14 160 1.88 24.47 46.81 2.13 0.00 51.06 0.00
NET-2 Buccella peruviana:41.46%Ammonia beccarii:12.20%Elphidium discoidale:8.54%
16 149 1.93 12.20 50.00 6.10 2.44 41.46 0.00
NET-3 Buccella peruviana:54.05%Ammonia beccarii:10.81%Elphidium discoidale:9.46%
11 151 1.45 9.46 33.78 6.76 2.70 56.76 0.00
NET-4 Buccella peruviana:63.16%Ammonia beccarii:7.89%Elphidium discoidale:6.58%
9 157 1.11 11.84 38.16 2.63 9.21 50.00 0.00
NET-5 Buccella peruviana:60.82%Ammonia beccarii:16.49%Elphidium discoidale:4.12%
10 181 1.16 8.25 58.77 9.28 5.15 26.80 0.00
NET-6 Buccella peruviana:52.73%Ammonia beccarii:10.91%Elphidium discoidale:3.64%
11 110 1.41 16.36 34.55 5.45 12.73 47.27 0.00
thetotalnumberofspeciesandthetotalassemblagesiden-tified in the Southern transect, only Buccella peruviana,Quinqueloculina seminula,Ammonia becariiandElphidium discoidalecanbeconsideredaswidelydistributedthrough-outthetransect.Buccella peruvianaisbyfarthedominantspeciesinsupratidalsettings(relativecontributionpersample>36%)andinalmostallsubtidalareas.Ammonia
beccarii, Bolivina striatula, Quinqueloculina seminula, and Textularia gramencanbelocallyabundantinsubtidalset-tings.D. williamsoniisdistributedinallsubtidalsamplesbutitisabsentinsamplesfromthebackshore.Intheforeshoreandthebackshore,thespeciesoftheBAEgrouprepresentsmorethan70%ofassemblages.
The following species are classified as accessory:
Table3.Mainspecies,numberofspecies(S),abundance,DiversityIndex[H(S)],andtaphonomicconditionofassemblagesfromthenortherntransect.
WP: Well preserved; R: recrystallized; A: abraded; F: fragmented; F&C: filled and coated.

Modern foraminifera from coastal settings, northern Argentina 53
Sample Principal species S Abundance (ind./10g) H (S) Indeterminable
individuals (%)Taphonomic condition (%)
WP R A F F&C
SST-1 Ammonia beccarii:40.85%Bolivina striatula:14.02%Buccella peruviana:14.02%
12 156 1.725 9.76 84.76 0.00 0.00 15.24 0.00
SST-2 Buccella peruviana:45.10%Ammonia beccarii:14.71%Discorbis williamsoni:6.86%Trochammina ochracea:6.86%
16 140 1.871 4.90 76.47 0.00 1.96 21.57 0.00
SST-3 Ammonia beccarii:28.91%Buccella peruviana:28.13%Quinqueloculina seminula:9.38%Elphidium discoidale:6.25%
15 481 1.915 9.38 70.31 0.00 0.78 28.91 0.00
SST-4 Buccella peruviana:47.30%Quinqueloculina seminula:13.18%Ammonia beccarii:6.76%Discorbis williamsoni:6.08%
23 939 1.743 12.16 72.30 0.00 0.00 27.70 0.00
SST-5 Buccella peruviana:18.12%Textularia gramen:15.77%Pyrgo ringens:8.72%Discorbis williamsoni:8.05%
38 330 2.647 15.10 73.82 0.00 0.34 25.84 0.00
SST-6 Buccella peruviana:31.72%Discorbis williamsoni:11.03%Textularia gramen:9.66%Elphidium discoidale:6.21%
19 274 2.001 24.83 60.69 0.00 0.00 39.31 0.00
SST-7 Buccella peruviana:34.04%Cibicidoides sp. cf. C. fletcheri:16.64%Elphidium discoidale:8.51%Discorbis williamsoni:6.38%
11 55 1.881 21.28 44.68 0.00 0.00 55.32 0.00
SET-1 Buccella peruviana:36.23%Ammonia beccarii:5.80%Cibicidoides sp. cf. C. fletcheri:2.90%
11 95 1.514 40.58 23.19 2.90 10.14 63.77 0.00
SET-2 Buccella peruviana:52.43%Ammonia beccarii:10.68%Elphidium discoidale:6.80%
11 192 1.397 15.53 44.65 5.83 7.77 41.75 0.00
SET-3 Buccella peruviana:64.00%Ammonia beccarii:12.00%Elphidium gunteri:6.00%
5 102 0.8554 14.00 38.00 6.00 10.00 46.00 0.00
SET-4 Buccella peruviana:63.79%Ammonia beccarii:6.90%Elphidium gunteri:6.90%Elphidium discoidale:5.17%
5 115 0.8716 15.52 22.42 17.24 17.24 43.10 0.00
SET-5 Buccella peruviana:61.82%Ammonia beccarii:10.91%Elphidium discoidale:5.45%Quinqueloculina patagonica:3.64%
5 109 0.8867 16.36 38.18 5.45 12.73 43.64 0.00
Quinqueloculina patagonica,Elphidium gunteri,Elphidium galvestonense Kornfeld,1931,Cibicidoidessp.cf.C. fletcheri,Cribrorotalia meridionalis (CushmanandKellett,1929),Pyrgo ringens (Lamarck,1804),andMiliolinella subrotunda.Otherspeciesoccurinrelativeabundancelowerthan1%oftheassemblageandareconsideredtoberareoraccidentalspecies.
Taphomomic modifications are directly related with waterdepthinsubtidalsettings(Figure3b).Wellpreserved
testsrepresentmorethan70%ofassemblages in thelowershoreface,andvarybetween60–44%intheuppershoreface.Fragmentedtestsvarybetween29–15%inthelowershorefaceandclearlyincreaseonshore.Intheuppershoreface,fragmentedtestsrepresent39–55%,andintheforeshore,fragmentedshellsattain63–41%andabradedshellsincreaseto8–10%.Inthebackshore,wellpreservedtestsvarybetween22–38%,butfragmentationisintense(40–43%).Abradedandrecrystalizedshellsare
Table4.Mainspecies,numberofspecies(S),abundance,DiversityIndex[H(S)],andtaphonomicconditionofassemblagesfromthesoutherntransect.
WP: Well preserved; R: recrystallized; A: abraded; F: fragmented; F&C: filled and coated.

Laprida et al.54
-10
-8
-6
-4
-2
0
1
2
3
80 700
102030405060
H.W.L.W.
S.H.W.
450 950 19501450
SST-1
SST-2
SST-3
SST-4
SST-5
SST-6SET-1
SST-7SET-2
SET-3
SET-4
SET-5
Distance from swash zone (m)
Hei
ghta
bove
sea l
evel
(m)
4
Well preserved Recrystallized Abraded Fragmented
Emerged beach Submerged beach
a)
b)
presentalmostexclusivelyintheemergedbeachandarerelativelyabundantinthebackshore,andespeciallyintheforedune,wheretheyattain17%.Thenumberoftestleft in open nomenclature due to taphonomic modification ishighest in the intertidalzone(40%)anddecreasesoffshore,whereasintheforeshoreandbackshoretendstoberatherconstant(~18%).
Foraminifera from Holocene environments
Tables2and5showdataobtainedfromtheKM187profile, where a total of 7,340 individuals were picked out. Speciesnumberineachsamplevariesbetween5and13,theShannon-Weaverdiversityindexvariesbetween0.73and1.48,andabsoluteabundancevariesbetween2(inthe
Figure 3. Schemes of the beach transects analysed and cumulative frequency diagrams based on taphonomic modifications of shells for each subenvironment.a)Northerntransect;b)southerntransect.

Modern foraminifera from coastal settings, northern Argentina 55
Tabl
e 5.
Lev
el th
ickn
ess,
litho
logi
cal c
hara
cter
istic
s, m
ain
spec
ies a
nd ta
phon
omic
con
ditio
n of
ass
embl
ages
from
the
KM
187
profi
le. S
: num
ber o
f spe
cies
, H(S
): D
iver
sity
Inde
x.
Lev
elT
hick
ness
(m)
Des
crip
tion
Prin
cipa
l spe
cies
SA
bund
ance
(in
d./1
0g)
H(S
)Ta
phon
omic
con
ditio
n (%
)
WP
RA
FF&
C
KM
187-
10.
40So
ilw
ith4
0%b
iocl
astic
par
ticip
atio
nco
nsis
ting
ofc
oars
esa
nd-s
ized
cru
shed
val
ves.
Bucc
ella
per
uvia
na:1
00%
12
00.
000.
000.
0050
.00
50.0
0K
M18
7-2
0.13
Poor
ly se
lect
ed y
ello
wis
h-w
hite
fine
gra
ined
bio
clas
tic g
rave
ls w
ith sa
nd-s
ized
bi
ocla
stic
mat
rix.
Bucc
ella
per
uvia
na:7
4.18
%El
phid
ium
dis
coid
ale:
13.
62%
Amm
onia
bec
cari
i:9.
86%
821
10.
919
50.2
30.
471.
8814
.08
33.3
3
KM
187-
30.
10Po
orly
sele
cted
yel
low
ish-
whi
te fi
ne g
rain
ed b
iocl
astic
sand
s; so
me
com
plet
e va
lves
up
to0
.7c
mlo
ng.
Bucc
ella
per
uvia
na:4
7.75
%Am
mon
ia b
ecca
rii:
32.6
2%El
phid
ium
dis
coid
ale:
16.
78%
648
51.
130
31.6
80.
002.
3619
.62
46.3
4
KM
187-
40.
07Po
orly
sele
cted
bro
wn
clas
ticsa
ndsw
ithc
onsi
dera
ble
amou
nto
fver
yco
arse
sand
-si
zed
bioc
last
icm
ater
ial.
Bucc
ella
per
uvia
na:5
4.71
%Am
mon
ia b
ecca
rii:
28.2
5%El
phid
ium
dis
coid
ale:
13.
00%
820
81.
137
41.2
60.
906.
2818
.39
33.1
8
KM
187-
50.
21Po
orly
sele
cted
yel
low
ish-
brow
nbi
ocla
stic
gra
vel,
with
10%
sand
cla
stic
mat
rix;
inci
pien
tcon
solid
atio
n.Bu
ccel
la p
eruv
iana
:50.
77%
Amm
onia
bec
cari
i:26
.92%
Elph
idiu
m d
isco
idal
e:1
8.46
%
525
01.
125
34.6
22.
695.
3812
.69
44.6
2
KM
187-
60.
06Po
orly
sele
cted
bro
wn
coar
se-g
rain
edb
iocl
astic
sand
swith
sand
-siz
edc
last
icm
atrix
.Bu
ccel
la p
eruv
iana
:36.
36%
Amm
onia
bec
cari
i:33
.64%
Elph
idiu
m d
isco
idal
e:2
5.45
%
721
51.
282
25.9
13.
1810
.91
10.4
549
.55
KM
187-
70.
24W
ells
elec
ted
yello
wis
h-w
hite
bio
clas
ticg
rave
lsw
ith5
%sa
ndc
last
icm
atrix
;com
plet
eva
lves
up
to1
cm
inle
ngth
.Bu
ccel
la p
eruv
iana
:46.
88%
Amm
onia
bec
cari
i:31
.25%
Elph
idiu
m d
isco
idal
e:1
5.63
%
553
1.25
621
.88
3.13
14.0
610
.94
50.0
0
KM
187-
80.
18Po
orly
sele
cted
ligh
t bro
wn
fine
bioc
last
ic g
rave
ls w
ith sa
nd c
last
ic m
atrix
and
cro
ss
lam
inat
ion;
inci
pien
tcon
solid
atio
n.Bu
ccel
la p
eruv
iana
:38.
66%
Amm
onia
bec
cari
i:27
.51%
Elph
idiu
m d
isco
idal
e:2
4.91
%
926
21.
418
33.4
61.
128.
1811
.15
46.1
0
KM
187-
90.
09Ye
llow
ish-
brow
nbi
ocla
stic
coa
rse
sand
with
10%
sand
cla
stic
mat
rix;c
ross
la
min
atio
n;a
few
com
plet
eva
lves
up
to1
cm
inle
ngth
.El
phid
ium
dis
coid
ale:
43.
36%
Bucc
ella
per
uvia
na:2
4.48
%Am
mon
ia b
ecca
rii:
19.5
8%El
phid
ium
gun
teri
:9.7
9%
726
31.
318
17.1
31.
407.
343.
8570
.28
KM
187-
100.
19W
hitis
h-br
own
bioc
last
ic fi
ne g
rave
l with
abu
ndan
t san
d cl
astic
mat
rix; c
ross
la
min
atio
n,e
ntire
val
vesu
pto
1c
min
leng
th,m
oder
atel
yw
ellp
rese
rved
.The
upp
er
part
show
sinc
ipie
ntc
onso
lidat
ion.
Elph
idiu
m d
isco
idal
e:3
8.36
%Bu
ccel
la p
eruv
iana
:30.
82%
Amm
onia
bec
cari
i:24
.32%
728
41.
335
14.3
83.
4210
.96
3.77
67.4
7
KM
187-
110.
09Po
orly
sele
cted
ligh
tbro
wn
coar
seb
iocl
astic
sand
with
cro
ssla
min
atio
n;a
few
co
mpl
ete
valv
esu
pto
0.5
cm
inle
ngth
.El
phid
ium
dis
coid
ale:
32.
19%
Amm
onia
bec
cari
i:27
.40%
Bucc
ella
per
uvia
na:2
5.34
%
728
41.
486
9.25
4.11
9.59
3.08
73.9
7
KM
187-
120.
17Po
orly
sele
cted
bro
wn
clas
ticsa
ndsw
ithc
ross
lam
inat
ion;
poo
rlyp
rese
rved
val
vesu
pto
2c
mlo
ngd
omin
ate
the
uppe
rpar
toft
hele
vel.
Bucc
ella
per
uvia
na:5
2.13
%Am
mon
ia b
ecca
rii:
31.8
0%El
phid
ium
dis
coid
ale:
13.
11%
629
81.
054
24.2
64.
9213
.77
7.21
49.8
4
KM
187-
13Íd
emK
M18
7-12
.Bu
ccel
la p
eruv
iana
:52.
63%
Amm
onia
bec
cari
i:27
.96%
Elph
idiu
m d
isco
idal
e:1
6.45
%
628
61.
114
30.5
93.
9510
.86
8.55
46.0
5
KM
187-
140.
12Po
orly
sele
cted
yel
low
ish-
whi
teb
iocl
astic
gra
vels
with
abu
ndan
t,w
ell-p
rese
rved
va
lves
up
to2
cm
inle
ngth
.Bu
ccel
la p
eruv
iana
:55.
31%
Amm
onia
bec
cari
i:22
.12%
Elph
idiu
m d
isco
idal
e:1
9.03
%
921
21.
132
25.2
22.
2112
.39
7.96
52.2
1

Laprida et al.56Ta
ble
5 (c
ontin
ued)
. Lev
el th
ickn
ess,
litho
logi
cal c
hara
cter
istic
s, m
ain
spec
ies a
nd ta
phon
omic
con
ditio
n of
ass
embl
ages
from
the
KM
187
profi
le. S
: num
ber o
f spe
cies
, H(S
): D
iver
sity
Inde
x.
Lev
elT
hick
ness
(m)
Des
crip
tion
Prin
cipa
l spe
cies
SA
bund
ance
(in
d./1
0g)
H(S
)Ta
phon
omic
con
ditio
n (%
)
WP
RA
FF&
C
KM
187-
150.
05Po
orly
sele
cted
yel
low
ish-
brow
nbi
ocla
stic
sand
s;p
oorly
pre
serv
edv
alve
sup
to1
.5
cmin
leng
thAm
mon
ia b
ecca
rii:
34.5
6%El
phid
ium
dis
coid
ale:
33.
09%
Bucc
ella
per
uvia
na:2
7.94
%
526
81.
230
16.9
12.
5717
.28
5.15
58.0
9
KM
187-
160.
09Ye
llow
ish-
brow
nbi
ocla
stic
gra
velw
ith2
0%c
last
icsa
ndm
atrix
;val
vesw
ell
pres
erve
dup
to1
.5c
min
leng
thBu
ccel
la p
eruv
iana
:58.
17%
Elph
idiu
m d
isco
idal
e:1
8.84
%Am
mon
ia b
ecca
rii:
14.1
3%
1333
41.
302
52.0
81.
665.
8213
.30
27.1
5
KM
187-
170.
07Ye
llow
ish
whi
te,v
ery
thic
kbi
ocla
stic
sand
s,w
ith1
0%o
fcla
stic
par
ticip
atio
nco
nsis
ting
ofm
ediu
msa
nd;s
carc
eco
mpl
ete
valv
esu
pto
0.5
cm
inle
ngth
.Bu
ccel
la p
eruv
iana
:51.
79%
Elph
idiu
m d
isco
idal
e:2
0.36
%Am
mon
ia b
ecca
rii:
17.5
0%
725
21.
212
33.9
31.
439.
6412
.86
42.1
4
KM
187-
180.
25Po
orly
sele
cted
,yel
low
ish
whi
teti
nged
with
pin
k,so
mew
hatc
onso
lidat
edb
iocl
astic
gr
avel
.Mat
rixc
onsi
stin
gof
bio
clas
ticsa
nd;c
ross
lam
inat
ion;
few
lith
icc
last
sup
to
0.5
cmlo
nga
ndc
ompl
ete
valv
esu
pto
1c
mlo
ngto
war
dsth
eto
pof
the
leve
l.
Bucc
ella
per
uvia
na:5
4.32
%Am
mon
ia b
ecca
rii:
23.0
5%El
phid
ium
dis
coid
ale:
16.
87%
623
51.
118
32.1
01.
659.
4710
.70
46.0
9
KM
187-
19Id
emK
M18
7-18
Bucc
ella
per
uvia
na:4
4.68
%El
phid
ium
dis
coid
ale:
27.
66%
Amm
onia
bec
cari
i:21
.28%
713
91.
313
34.0
40.
0013
.48
12.0
640
.43
KM
187-
200.
08W
ells
elec
ted
brow
ncl
astic
sand
,with
littl
epa
rtici
patio
nof
bio
clas
ticm
ater
ial;
wel
lpr
eser
ved
disa
rticu
late
dbi
valv
esa
ndg
astro
pods
up
to2
cm
(the
form
er)a
nd5
cm
lo
ng(t
hela
tter)
.
Bucc
ella
per
uvia
na:5
1.07
%Am
mon
ia b
ecca
rii:
27.0
4%El
phid
ium
dis
coid
ale:
18.
45%
520
41.
084
38.6
33.
0021
.03
12.4
524
.89
KM
187-
210.
08Po
orly
sele
cted
bro
wn
clas
ticsa
ndw
ith3
0%o
fcla
stic
par
ticip
atio
n;p
aral
lel
lam
inat
ion;
poo
rlyp
rese
rved
com
plet
eva
lves
up
to2
cm
long
and
coa
rse
sand
-siz
ed
valv
efr
agm
ents
.
Bucc
ella
per
uvia
na:4
0.70
%Am
mon
ia b
ecca
rii:
28.1
4%El
phid
ium
dis
coid
ale:
26.
63%
619
31.
235
12.0
63.
0214
.57
2.51
67.8
4
KM
187-
220.
12M
oder
atel
yse
lect
edb
row
ncl
astic
sand
with
5%
ofb
iocl
astic
par
ticip
atio
n;p
aral
lel
lam
inat
ion;
poo
rlyp
rese
rved
com
plet
eva
lves
up
to2
cm
long
and
coa
rse
sand
-siz
ed
valv
efr
agm
ents
.
Bucc
ella
per
uvia
na:5
7.87
%El
phid
ium
dis
coid
ale:
22.
75%
Amm
onia
bec
cari
i:15
.73%
834
31.
116
49.1
61.
978.
719.
8330
.34
KM
187-
230.
15Ye
llow
ish
brow
nbi
ocla
stic
coa
rse
sand
swith
40%
ofc
last
icp
artic
ipat
ion;
cro
ss
lam
inat
ion;
abu
ndan
tlith
icc
last
sof0
.2c
min
dia
met
er.
Elph
idiu
m d
isco
idal
e:4
0.00
%Bu
ccel
la p
eruv
iana
:36.
80%
Amm
onia
bec
cari
i:19
.60%
722
91.
214
30.4
03.
2019
.60
6.00
40.8
0
KM
187-
240.
08Ye
llow
ish
brow
ncl
astic
sand
with
15%
ofb
iocl
astic
par
ticip
atio
n;c
ross
lam
inat
ion;
sa
nd-s
ized
coa
rse
valv
esfr
agm
ents
.Bu
ccel
la p
eruv
iana
:50.
16%
Elph
idiu
m d
isco
idal
e:2
6.56
%Am
mon
ia b
ecca
rii:
20.3
3%
629
41.
146
39.3
42.
9516
.07
10.1
631
.48
KM
187-
250.
08B
row
nco
arse
-gra
ined
bio
clas
ticsa
ndsw
ith1
0%o
fcla
stic
par
ticip
atio
n;a
bund
ant
lithi
ccl
asts
up
to0
.5c
min
dia
met
era
nda
few
com
plet
eva
lves
up
to2
cm
inle
ngth
.Bu
ccel
la p
eruv
iana
:51.
83%
Elph
idiu
m d
isco
idal
e:3
5.55
%Am
mon
ia b
ecca
rii:
10.3
0%
425
11.
001
47.5
10.
6610
.96
10.6
330
.23
KM
187-
260.
15Po
orly
sele
cted
yel
low
ish
bioc
last
icg
rave
lsw
ithli
ttle
bioc
last
icm
atrix
;com
plet
eva
lves
up
to1
.5c
min
leng
tha
ndfe
wc
oars
ecl
asts
ofu
pto
2.5
cm
ind
iam
eter
.Bu
ccel
la p
eruv
iana
:58.
65%
Elph
idiu
m d
isco
idal
e:2
6.39
%Am
mon
ia b
ecca
rii:
7.62
%
933
51.
165
48.0
90.
2911
.73
14.0
825
.81
KM
187-
270.
14Pi
nk-ti
nged
whi
te, fi
ne-g
rain
ed b
iocl
astic
gra
vel w
ith li
ttle
sand
y m
atrix
; a fe
w
lithi
ccl
asts
up
to0
.5c
min
dia
met
er;n
umer
ousw
ellp
rese
rved
com
plet
eva
lves
and
ga
stro
pods
up
to2
cm
(the
form
er)a
nd7
cm
long
(the
latte
r).
Bucc
ella
per
uvia
na:5
0.94
%El
phid
ium
dis
coid
ale:
30.
00%
Amm
onia
bec
cari
i:14
.06%
531
51.
142
41.2
50.
9410
.94
14.3
832
.50
KM
187-
280.
44M
oder
atel
yse
lect
edy
ello
wis
hbr
own
bioc
last
icsa
ndsw
ithli
ttle
parti
cipa
tion
of
clas
ticm
atrix
;irr
egul
arli
thic
cla
stsu
pto
1c
min
dia
met
er.
Bucc
ella
per
uvia
na:5
2.38
%El
phid
ium
dis
coid
ale:
29.
21%
Amm
onia
bec
cari
i:13
.33%
425
20.
730
53.6
51.
2710
.16
13.9
720
.95
KM
187-
29Id
emK
M18
7-28
Bucc
ella
per
uvia
na:4
9.37
%El
phid
ium
dis
coid
ale:
32.
58%
Amm
onia
bec
cari
i:10
.03%
838
31.
265
56.3
90.
7511
.28
11.7
819
.80

Modern foraminifera from coastal settings, northern Argentina 57
Well preserved Recrystallized AbradedFragmented Chamber fillings and
polished coating shells
0 10 20 30 40 50 60 70 80 90 100
KM187-1KM187-2KM187-3KM187-4KM187-5KM187-6KM187-7KM187-8KM187-9
KM187-10KM187-11KM187-12KM187-13KM187-14KM187-15KM187-16KM187-17KM187-18KM187-19KM187-20KM187-21KM187-22KM187-23KM187-24KM187-25KM187-26KM187-27KM187-28KM187-29
%
Leve
l
Figure 4. Cumulative frequency diagram based on taphonomic modifi-cations of shells of the Profile KM187.
topmostsample)and485specimensin10gdrysediment.OnlyBuccella peruviana,Ammonia becariiandElphidium discoidalecanbeconsideredaswidelydistributedinallsamples.Infact,theBAEgrouprepresentsnear95%oftheassemblageinsomesamples.Buccella peruvianaisbyfarthedominantspecies(48%ofthewholeassemblage).Elphidium discoidale represents24%andAmmonia bec-carii21%ofthetotalassemblage,andonlyElphidium guntericanbeconsideredasanaccessoryspecies.Allotherspeciesoccurinrelativeabundancelowerthan1%ofeachassemblageandareconsideredtoberareorac-cidentalspecies.
AssemblagesfromKM187aremoderatelywelltopoorlypreserved.Inthebase(KM187-29/22),themajorityofshellsarewellpreserved(Figure4),whileinthemiddleof the section (KM187-21/12) shells with chamber fillings and/ordarkpolishedcoatingstendtoincreaseupward(KM187-11/9) and finally constitute more than 70 % of the assemblages.Inthetopmostsamples(KM187-8/1),wellpreserved shells and shells with chamber fillings and/or dark polishedcoatingsareequallyrepresented,whilefragmentedshellstendtoincreaseandabradedshellstodecreaseup-ward.Remarkably,taphonomicconditionofassemblagesdoesnotseemtoberelatedwithlithology(Figure2).
Statistical analyses
Theclusteranalysisperformedonthewholesetofsampleswithregardtospeciescomposition(cofeneticcor-relation coefficient: 0.8568) yielded a dendogram (Figure 5) where five clusters and one independent entity –namely asamplethatdoesn’tgroupwithanyothers(Herrera-Moreno,2000)–wererecognized.Thelargestgrouping(Cluster5,Table6)comprisesallassemblagesfromtheKM187 profile, with the exception of sample KM187-2, consistentwiththeoverwhelmingdominanceoftheBAEgroup.SampleKM187-2differsbecausethedominanceof Buccella peruvianaisevenmoremarked,accountingfor70%oftheassemblage,whichlinksthissamplewithmod-ernassemblagesfromthebackshoretothelowershoreface(Cluster4,Table6).There,B. peruvianaisatleastfourtimesasabundantasA. beccarii,whichinturnisovertwiceasabundantasE. discoidale.Theremainingclusterscomprisesubmergedsamplesinwhichotherspeciesareasabundantas,orevenmorethanB. peruviana,A. beccarii orE. discoidale.Thelowershorefacefromthenortherntran-sectischaracterizedbythedominanceofQuinqueloculina seminula;bycontrast,intheremainingsubmergedsamplesfromthistransect(Cluster4),thisspeciesbecomes10timeslessabundant.Quinqueloculina seminula isalsorelevantasanaccompanyingspeciesinCluster6,madeupoftwoofthelowershorefacesamplesfromthesoutherntransectdominatedbyA. beccarii,andinthesampleSST-5,anindependententity(Cluster2).SampleSST-5standsaloneonaccountofbeingtheonlyonewhereB. peruvianaisco-
dominantandwhereTextularia gramenmakesup25%oftheforaminiferalfauna.Finally,Cluster3groupsthetwoshallowestsamplesoftheuppershorefacefromthesoutherntransect–SST6andSST7-,whereB. peruvianabecomesagaindominantandisaccompaniedbyE. discoidale, T. gramen, andDiscorbis williamsoni.
Theclusteranalysisperformedonthewholesetwithregard to taphonomic modifications (cofenetic correlation coefficient: 0.8611) (Figure 6) led to the recognition of sixclustersandtwoindependententities(Table7).MostsamplesfromthebaseofKM187aregroupedinCluster1,characterizedbyyieldingthegreatestamountofwell-preservedtests(almost50%inaverage)ofthewholesetofHolocenesamples.InCluster2,whichincludesseveralsamplesmostlyfromthetopandmiddlelayersofKM187,filled or coated shells are nearly twice more abundant in averagethaninCluster1.Thefourremainingsamplesofthe profile consist of almost 70 % of filled or coated shells andaregroupedinCluster3.Samplesfrommodernenviron-ments, which lack filled or coated shells, fall entirely apart fromHoloceneonesandaregroupedinaccountoftheirpositionalongtheforeshoreandtheuppershoreface.Some

Laprida et al.58
0
6
12
18
24
30
36
42
48
54
0.32 0.4 0.48 0.56 0.64 0.72 0.8 0.88 0.96
Similarity
NST-1NST-2NST-3SST-5SST-6SST-7SST-4NST-4SET-1SET-2NET-3NST-5NST-6NET-4NST-8SET-3SET-4NET-5NET-6SET-5KM187-2SST-2NST-7NET-2NET-1KM187-3KM187-5KM187-20KM187-4KM187-13KM187-18KM187-14KM187-17KM187-22KM187-16KM187-24KM187-6KM187-8KM187-19KM187-21KM187-7KM187-25KM187-29KM187-27KM187-28KM187-26KM187-12KM187-9KM187-10KM187-11KM187-15KM187-23SST-3SST-1
123
4
5
6
0
6
12
18
24
30
36
42
48
54
-64 -56 -48 -40 -32 -24 -16 -8Similarity
NST-1
NST-2
NST-3
SST-5
SST-6
SST-7
SST-4
NST-4
SET-1SET-2
NET-3
NST-5
NST-6
NET-4
NST-8
SET-3
SET-4
NET-5
NET-6SET-5
KM187-2
NST-7NET-2
NET-1
KM187-3KM187-5
KM187-20
KM187-4
KM187-13KM187-18
KM187-14
KM187-17
KM187-22KM187-16
KM187-24
KM187-6
KM187-8
KM187-19
KM187-21
KM187-7
KM187-25
KM187-29
KM187-27KM187-28
KM187-26
KM187-12
KM187-9KM187-10
KM187-11
KM187-15
KM187-23
SST-3SST-1
SST-2
1
2
3
4
56
7
8
ofthelowershorefacesamplesfalltogetherinCluster4,whichshowsthehighestproportionofwellpreservedtests(75%onaverage)ofthecompleteset.Cluster7comprisesmostoftheuppershorefaceofthenortherntransectandsomeforeshoresamples,allofwhichpresentarelativelyhighamountoffragmentedshells.Finally,mostemergedanduppermostshorefacesamplesaregroupedinCluster8where50%oftestsarefragmented.SamplesSET-1andSET-4areindependententities;theformer,locatedatthebaseoftheforeshore,becauseofthehighproportionofbrokenshells–thehighestinthewholeset–;andthelatter,locatedatthebaseoftheforedune,becauseoftherelativelyhighproportionofrecrystallizedandabradedtests.
Clusteranalysissuggeststhattherearesomemean-ingfuldifferencesamongsamplesfromdifferentlocations,whichledustoperformaseriesoflineardiscriminantanalysesonthemoderndatasetinordertoevaluatetheconsistenceofthesedifferences.TheresultsareshowninTables8and9.Thetestsuccessfullydiscriminatessamples
frombothtransects:however,itdoesn´tperformaswellindistinguishingsupralittoralfromsublittoralsamples,exceptwithregardtotaxonomiccomposition.Additionally,thecorrelationbetweengrainsize,preservationstateandspeciescompositionwasassessedbymeansoftheManteltest.Theresults(Table9)showthatspeciescompositionis marginally but significantly correlated with taphonomic signatureinbothsites,whereasspeciescompositionandtaphonomic signature are significantly correlated with grain sizeonlyinthenortherntransect.
DISCUSION
Littoral foraminifera from modern beaches
ThebenthicforaminiferafromcoastalsettingsofnorthernBuenosAiresAtlanticcoastaremoderatelydiverse.Shannon-Wienerindexvaluesobtainedinthisstudyareconsistentwiththosetypicalofmarginalmarineenviron-ments,accordingtoMurray(2006).Thenumberofbenthic
Figure5.Dendogramofsamplesgroupedbyspeciescomposition.Thedashedlineindicatesthemid-lengthofthelongestdistancebetweensuccessivenodes.
Figure 6. Dendogram of samples grouped by taphonomic modifications. Thedashedlineindicatesthemid-lengthofthelongestdistancebetweensuccessivenodes.

Modern foraminifera from coastal settings, northern Argentina 59
foraminiferainsubtidalsettingsissimilartothoseobtainedfromtheinnershelfelsewhere(Murray,2006;Mendeset al.,2004).Afewsampleshaveamodestcontributionofmiliolidswithporcelaneouswalls,asMurray(2006)pointedoutforthewholesouthwesternSouthAtlantic.Agglutinatedshellstendtobedestroyeddifferentiallyeveninsubtidalset-tings(DenneandSenGupta,1989;Murray,2006;Barbieri,1996),andasaconsequence,hyalinecalcareousshellsdominatethewholemodernbenthicassemblage.
SpeciesrecordedinnorthernBuenosAireslittoralfol-lowthelargeregionaltrendofthelittoralbenthicforamin-iferafromtheSouthwestSouthAtlantic,butclearlydifferfrom the inner shelf (>40 m depth) assemblages defined byBoltovskoyandTotah(1985)intermsoftaxonomiccomposition, dominance and diversity. All identified spe-ciesdwellinsouthernBrazilandUruguayneighboringlit-toralareas(Closs,1963;ClossandBarberena,1962;ClossandLopes-Madeira,1968;Boltovskoy,1970).Amongthedominantspecies,Buccella peruvianaisthemostconstantandcommonforaminiferaintheSouthAtlanticlittoralareaanddominatesthroughoutthewholeArgentineshelf(Boltovskoy,1970).Ammonia beccarii,Quinqueloculina
Cluster / independent entity
Samples Species composition (% )Buccella
peruvianaAmmonia beccarii
Elphidium discoidale
Quinqueloculina seminula
Textularia gramen
Discorbis williamsoni
1 NST-1/3 20.55 5.44 0.84 39.46 0.00 6.75
2 SST-5 29.03 1.61 3.23 11.83 25.27 12.90
3 SST-6,7 48.71 0.52 10.89 5.70 10.34 12.93
4 SST-2,4;NST-4/8;SET-1/5;NET-1/6;KM187-2
62.93 15.35 8.81 3.46 0.56 2.34
5 KM187-3/29 48.67 24.03 25.23 0.00 0.00 0.00
6 SST-1/3 26.57 45.08 6.68 10.22 0.00 3.48
Table6.Samplegroupingaccordingto(average)percentageofspeciescomposition.Onlyspecieswhichrepresentmorethan10%ofaclusterandindependententitiesareshown.
seminulaandDiscorbis williamsoniarewidelydistributedallalongArgentinelittoralandinnershelf(Boltovskoy1970;BoltovskoyandWright,1976;Boltovskoyet al.,1980).Elphidium discoidale is typicallyaneurytopic,nearshorespeciesinhabitingSouthAmericanwarmwatersnorthof41°S(Boltovskoy,1970;HippensteelandMartin,1999).
Livingforaminiferausuallyshowapronouncedlypatchygeographicdistributioninshallowwaters(Schaffer,1971;BoltovskoyandWright,1976).Consequently,detailedtaxonomiccompositionofmoderntotalassemblagesisfrequentlydeterminedbylocal(smallscale)environmentalparameters.StrongdominanceofAmmonia beccariiandBolivina striatulainthelowershorefaceofthesoutherntransectseemstoberelatedtomuddysubstrates,sincethesespecies prefer organic-rich, fine-grained sediments (Alve andMurray,1999).Abundanceandtaxonomiccompositionof total littoral assemblages reflect biological preferences ofspeciesbutalsotaphonomicprocessesoperatinginthecoastalareas(Barbieri,1996;MurrayandAlve,1999).This could account for the fact that, although our findings confirm the general taxonomic composition consigned by
Cluster / independent entity
Samples Taphonomic signature (%)Well
preservedFragmented Filled/
coatedAbraded Recrystallized
1 KM187-2,4,16,20,22,24,25/29 47.05 13.00 28.15 10.44 1.352 KM187-3,5/8,12-15,17/19,23 28.85 10.41 47.10 11.34 2.313 KM187-9/11,21 13.21 3.30 69.89 10.62 2.994 NST-1,2
SST-1/575.60 23.96 0.00 0.44 0.00
5 SET-4 22.42 43.10 0.00 17.24 17.246 SET-1 23.19 63.77 0.00 10.14 2.907 NST-3/7;NET-2,5;SST-6;SET-2 55.89 39.31 0.00 2.12 2.688 NST-8;NET-1,3-4,6;SST-7;SET-3,5 39.60 50.56 0.00 6.06 3.78
Table7.Samplegroupingaccordingtoaveragetaphonomicsignatureofsamples.

Laprida et al.60
Boltoskoy(1970),small,fragileand/orscarceandrarespecieshavenotbeenfoundinourlittoralsamples.Mostinnercontinentalshelfassemblagesareparauthochtonous(Callenderet al., 1992), and potential taphonomic modifi-cationsobservedaretransportoftestsfromthepositionoflife to be deposited elsewhere, damage of tests and, finally, totaldestructionoftests(Murray,2006).Thisisevidentinthebackshoreandforedunewherethenumberofspeci-mensanddiversityofassemblagesarestronglyreducedasaconsequenceofbiostratinomicprocesses(mainlyshelldestructionrelatedwithstormwaves,windsandsubaerialexposure)duetopreferentialpreservation/destructionofmoreresistant/fragilespecies.
TheMantelTestindicatesthatintheNortherntransect,taphonomicsignaturesandtaxonomiccompositionoftotalassemblagesarehighlycorrelatedtograinsize,andhence,withsedimentaryprocessesinthelittoral.Inturntapho-nomicsignaturesofassemblagesarecorrelatedtotaxonomiccomposition,indicatingthattaphonomic(mainlybiostratin-omic) processes are crucial in defining the specific composi-tionofassemblagesinthenortherntransect.Totalabundanceand diversity confirm this hypothesis since they are related towaterdepthinsubtidalsettingsandwithpositionfromtheshorelineinthebackshore.However,theserelationshipsarenotasstraightforwardintheSoutherntransect,whereonly taphonomic modifications are related with water depth andpositionfromtheshoreline.FortheSoutherntransect,the Mantel test does not show any significant correlation of thetaphonomicsignaturesortaxonomiccompositionwithgrainsize,althoughtaphonomicsignaturesandtaxonomiccompositionshowaweak(marginal)correlation.Aplau-sibleexplanationwouldbethattaphonomicprocessesotherthan purely biostratinomics are crucial in defining the spe-cific compositions of assemblages in the southern transect, sincevariationsinshellpreservationarenotlinkedonlywithbathymetrically-dependentenvironmentalvariables(Loubereet al.,1993;MurrayandAlve,1999).However,additionalenvironmentaldataisneededtoidentifywhichprocessesareoperatinginthegenerationofdeath(andhencetotal)assemblagesintheSoutherntransect.
Althoughlocationofsamplescanbeamajorfac-
toraffectingfaunaltrendseveninasmallscale,relativeabundance, abundance and taphonomic modification of shellsseemtoberathersimilarinbothtransects.Clusteranalysessuggestthattherearesomemeaningfuldifferencesinspeciescompositionamongmodernsamplesfromsimilardepths,especiallyinsubtidalsettings.Infact,taxonomiccomposition does not allow bathymetrical classification ofsampleswhenallsamplesareconsideredtogether.ThislendsweighttoBoltovskoy’s(1970)statementthatforaminiferalassemblagesbetweentheintertidalzoneand-15mdepth“areverysimilar”(sic.).However,therearesomebathymetrictendenciesintaxonomiccompositionespeciallyinthelowershoreface(Table6).There,Buccella peruvianarepresents“only”20–26%ofassemblagesandQuinqueloculina seminularepresentsbetween10–39%.Intheuppershoreface,theBAEgroupincreasesitsdominance,anduppershoreface,foreshore,backshoreandforeduneassemblagesarerathersimilarintermsoftaxonomiccom-positionduetotheoutstandingdominanceofBuccella pe-ruviana.Thereisnodoubtthatbiologicalandtaphonomicalprocessesdeterminethebenthicforaminiferalassemblagesinthelowershoreface,buttaphonomicprocessesdominatenotonlyintheemergedbeach,butalsoinuppershorefacesettings.Intermsofwaveenergy,emergedbeachisonlyinfluenced by storm waves and winds, and sediments can beperiodicallyremobilized.Shellsderivedfromshorefacesettingsaccumulateonbackshoreandforedune(Murray,2006),andultimatelytheyreturntotheforeshoreandup-pershorefacebyoffshoretransportofsandduringepisodesofcoastalerosion(AlbertzartandWilkinson,1990).Thiscoastaldynamicstrengthenstheargumentthatsupratidalanduppershorefaceassemblagesaretaxonomicallysimilarduetotaphonomicprocessesoperatinginthebeach.
Somespeciesshowpatchydistribution,amajorfactorindistinguishingbetweentransectsasassessedbylineardiscriminantanalysis(LDA)performedontaxonomiccomposition.Trochammina ochracea,Quinqueloculina milletti,Bolivina striatula, andTextularia gramenarelo-callyabundantinsubmergedsites,probablyrelatedtolocalenvironmentalparameters.AsimilarpatchydistributionofforaminiferaoffArgentinecoastswascitedbyBoltovskoyandLena(1969).Additionally,LDAalsodistinguished
Emerged vs. submerged beach
Northern vs. southern transect
NorthernTransect
SouthernTransect
Emergedbeach
Submergedbeach
Grainsize
92.3% 98.6% 57% 71.6%
Taphonomicsignature
92.3% 100% 74.4% 72%
Speciescomposition
100% 100% 100% 100%
Table8.Comparisonofaveragepercentagecorrectclassificationofsamplesaccordingtolineardiscriminantfunctionconsideringgrainsize,taphonomicsignaturesandspeciescomposition.
Northern transect Southern transectTaphonomic
signatureSpecies
compositionTaphonomic
signatureSpecies
composition
Grain size R=0.4931p=0.0024*
R=0.9267p<0.0001*
R=0.0864p=0.2928
R=0.0787p=0.2616
Speciescomposition
R=0.4683p=0.0106* -
R=0.3684p=0.0102* -
Table9.Manteltestsshowingthecorrelationbetweengrainsize,tapho-nomicsignaturesandtaxonomiccompositionineachtransect.Statisticalsignificance (*) is granted to the tests with associated p-value lower than alpha αad=0.017(numberofpermutationsN=5000).

Modern foraminifera from coastal settings, northern Argentina 61
successfullybetweensublittoralandsupralittoralassem-blages.Inotherwords,evenwhentaxonomiccompositionofmodernassemblagesdifferinbothtransects,subtidalandsupratidalassemblagescouldbeeasilydiscriminated.ThispointisparticularlyrelevantbecauseitimpliesthatmodernbenthicforaminiferalassemblagesfromsandybeachesofnorthernBuenosAiresProvincearepotentiallyanadequategeneralbaselinetodiscriminateQuaternarybeachsubenvironmentsifbothtaxonomicandtaphonomicfeaturesareconsideredtogether.
Asnotedabove,LDAdistinguishedverywellbetweennorthernandsoutherntransect,whichsuggeststhatbiologi-calandsedimentologicalprocessesoperatingonbothsitesdiffer,especiallyinsubtidalsettings.Particularenviron-mentalconditions,expressedasdifferentgeomorphologicalfeaturesandgranulometrictendenciesbetweennorthernandsoutherntransectsmaybeatleastinpartresponsibleforpatchydistributionofspecies.Conversely,foredune,backshore,anduppershorefaceassemblagestendtogroupwhentaxonomiccompositionisconsidered.Since,intheemergedbeach,taphonomicprocesseslargelydominateoverbiologicalones,itcanbesuggestedthattaphonomicprocesses are crucial in defining the taxonomic composition ofmoderntotalassemblagesfromtheuppershorefacetotheforedune,whilebiologicalprocesses,whicharedeterminedbylocalenvironmentalparametersinthesubmergedbeach,areresponsiblefordeterminingtaxonomiccompositionofmoderntotalassemblagesinthelowershoreface.
Taphonomicsignaturesarenotuniformlydistributedalongthebathymetricgradient.Notably,taphonomicmodi-fications allow the recognition between lower shoreface, uppershorefaceandforeshore/backshore/foreduneassem-blages,indicatingthattaphonomicprocessesaremainlydeterminedbywaterdepth,anddependmainlyonwaterenergy,aerialexposure,andbeachdynamics,thusshowinghighpotentialtobeusedinpaleoenvironmentalanalyses.
Morphologicalfeaturesofthespeciesinvolvedcanexplaintheirselectivepreservationinthedifferentsubset-tings.TheBAEgroupdominatesfromtheuppershorefacetotheforedune.Elphidium discoidalehasaveryheavy,strong,biconvexplanospiralshell,whileBuccella peruvianaandAmmonia beccariihavestrong,biconvex,trochospiralshell.AccordingtoPeeblesandLewis(1991),largetestsaremoreresistanttoabrasionthansmallertests.Thesespeciesarehencepreferentiallypreservedinthesedimentsoftheforeshoreandtheforedune.BeachesinthenortheastofBuenosAiresProvinceareexposedtostrongwaveac-tion,andtheyarestormandwavedominated,thusitisnotsurprisingthatthisthreelargespeciesdominatetotalas-semblagesespeciallyathighenergysettings,wherefragilespeciesarepreferentiallyremovedand/ordestroyed.Inthelowershoreface,fragilespeciessuchasQuinqueloculina seminula,Discorbis williamsoni and Textularia gramen aremoderatelywellrepresented,buttheyareselectivelydestroyedinhighenergyandsupratidalsettings.Intheuppershorefaceandintheemergedbeachthesespecies
representinaveragelessthan4%oftotalassemblages,whereasinsubtidalsettingstheycanattainmorethan10%.Hyalineshellsarelesssusceptibletodissolutionthanarenaceousshells(PeeblesandLewis,1991),andaccordingtoCorlissandHonjo(1981),hyalinetrochospiraltaxaaremoreresistanttodissolutionthanporcelanaceousspeciessuchasQuinqueloculina seminula.Textularia gramenhasabiserialtestwithagglutinatedwallwithrelativecoarsegrains,traversedbycanaliculithatmayopenasperforationsorbeclosedexternallybyathinagglutinatedlayer(LoeblichandTappan,1988).Inturn,thin-walledforaminiferawithahighdensityofporesaremoresusceptibletodissolution(CorlissandHonjo,1981).Discorbis williamsonihasasmalltestwithcalcareous,thinwallcoarselyperforatedin the umbilical side, with prominent umbilical flaps and chamberlets beneath the flaps.
Comparison between Holocene and modern assemblages
TaxonomiclistsarerathersimilarbetweenHoloceneandsupratidalmodernassemblages,buttheydifferwithrespecttodiversity,numericaldominanceoftaxaandta-phonomic modifications. In general, diversity is strongly reducedinHolocenesamples,whereonlythethreespeciesoftheBAEgroupoccurconsistentlyandabundantly.TheBAEgroupisalsocommoninpresentdayhabitatsandtheyareespeciallyabundantintheemergedbeach,buttheytendtobecomparativelylessdominant.Additionally,somekey species locally important in defining groups of modern totalassemblagesarevirtuallyabsentfromtheHolocenesamples.Forexample,Discorbis williamsoni isnearlyabsentfrombothemergedbeachandHolocenesamples,whileinsubtidalsettingsitusuallyrepresentsmorethan3%inaverage(Table6).Quinqueloculina seminulaiswellrepresentedinsubtidalsettingsofbothtransects(>10%average,Table6),butitisnearlyabsentfromHoloceneandsupratidalbeachsamples.Thesespecies,whichcanbelocallyimportantconstituentsofthebenthicassemblagesinsubtidalsettings,arefragileenoughtobedestroyedbybiostratinomicand/orfossildiageneticprocessesthattakeplaceintheemergedbeach.Tosumup,thedominanceoftheBAEgroupandtheabsenceofDiscorbis williamsoniandQuinqueloculina seminulainHoloceneassemblagesseemtoindicatethatKM187sectionrepresentsdepositioninabackshore,wave-dominatedsetting.ResemblancebetweenHoloceneandemergedmodern-beachtotalas-semblages seems to reflect the operation of some common taphonomicprocesses.InKM187sediments,theincreaseofabradedandrecrystalizedshellsandtheconcomitantdimi-nution of well preserved tests towards the top of the profile indicatethetransitionofanupperforeshoretobackshoreenvironments.TaxonomiccompositionandtaphonomicsignaturesofHoloceneassemblagesseemtoindicatethattheHolocenebeachesofthesouthernSamborombónBay

Laprida et al.62
werestormandwavedominated.Fossilassemblagesseemtobeinfactstronglyaffectedbytaphonomicprocessesrelatedtowaves,tidesandwind,whichhavedestroyedpreferentiallylittleand/orfragilespecies.Strongspeci-mensweredisplacedandconcentratedupshorebystormsurges,whichcarriedtotheupperbeachataphonomicallyandtaxonomicallydistinctiveassemblage,theBAEgroup,moderatelytopoorlypreserved,andstronglydominatedbyBuccella peruviana.Moreover,fromtheclusteranalysis,itisclearthattheHolocenesamplesaremoresimilartoeachotherthanareassemblagesfrommodernenviron-ments(bothsubtidalandsupratidal).ThisseemstoindicatethattaphonomicprocesseshavebeenparticularlyintenseduringtheMidHoloceneregression,andtheystronglyinfluenced not only the preservational states of shells, but alsodeterminedthetaxonomiccompositionanddiversityoffossilassemblages.
Although there are some similarities betweenthe taxonomiccompositionofHoloceneandmodernassemblagesfromemergedbeaches,theyarenotstrongenough todrawaperfectparallelbetweenHoloceneand modern beaches.As Holocene assemblages areoverwhelminglydominatedbytheBAEgroup,taphonomicanalysis is a more efficient tool than taxonomic composition toanalyzepaleoenvironmentalevolutioninHolocenesamples.Additionally,Holoceneandmodernassemblagesare not lumped when taphonomic modification of tests is considered,indicatingthattaphonomicprocessesprevailingintheHoloceneandmodernenvironmentsarequitedifferent.Themaindifferencebetweenthemisthepresenceofdarkshells with chamber fillings and/or with polished coatings inHolocenesediments.Darkshellsreflectdiageneticmodifications occurred probably in reducing, dysaerobic environments (Martin,1999) suchas saltmarshandswamps.Thus,theseshellsareallochthonous,andtheywereprobablyerodedfromLatePleistocene-EarlyHolocene?darkdepositssuchasthosedescribedbyNábel(1987)fromthe adjacent shelf. A similar taphonomic modification has beenobservedpreviouslyinwithin-habitattime-averagedassemblagesfromthetypesectionoftheCerrodelaGloriaMemberofLasEscobasFormation(LapridaandBertels-Psotka, 2003). In the base, shells with chamber fillings and/orwithpolishedcoatingsrepresentinaverage28%oftheassemblage(Cluster1,Table7),whileintheupperparttheyaccountformorethan47%(Clusters2and3,Table7).ThereductionintherateofsealevelriseduringtheMidHoloceneallowedonshoretransportprocessestodominatethelittoralsystemalongalowgradientinnershelfsuchasthebonaerensian(TaylorandStone,1996).Inthiscontext,thepreservationalstatesofshellsfromthetopoftheKM187sectionmayindicatethaterosionofrelicticcoastalsedimentwasmoreintense,probablyduetotherelativefallinsealevel.Thus,taphonomicsignaturesofbothmodernandHoloceneassemblagesindicatethatthesourcematerialandcoastaldynamicsofmodernandHolocenebeachesarenoticeablydifferent.SedimentsourcesofHolocenebeaches
ofthesouthernSamborombón(proto)BayincludedLatePleistocene-EarlyHolocenecoastalandmarginalmarineenvironmentsstronglysubjectedtoreworkingduringtheMid-lateHolocenesealevelfall.GrainsizedistributionofHolocenebeachridgessuggest that theywerebuiltunderconditionsthatarenotoperativetoday.Forexample,Holocene beach deposits are significantly more shell rich thanmodernbeaches,andwecanconcludethateithershellabundancewashigherorsandsupplywaslowerduringtheMid-to-LateHolocenethantoday.
CONCLUSION
EvenwhenmodernbeachesfromNorthernBuenosAiresProvinceareheterogeneousintermsofgeomorphol-ogyandtaxonomiccompositionofbenthicforaminiferalassemblages,discriminationbetweensubtidalandsupra-tidalmodernassemblagesisaccurateenoughtousetheminpaleoenvironmentalreconstructionofHolocenelittoralenvironmentswhentaxonomicandtaphonomicdataareconsideredtogether.Abundanceandtaxonomiccompositionof total littoral assemblages reflect biological preferences of speciesbutalsotaphonomicprocessesoperatinginlittoralenvironments.Biologicalandtaphonomicalprocessesde-terminethebenthicforaminiferalassemblagesinthelowershoreface,buttaphonomicprocesseslargelydominatenotonlyintheemergedbeach,butalsoinuppershorefacesettings.Asaconsequence,supratidalanduppershorefaceassemblagesaretaxonomicallysimilarduetotaphonomicprocessesoperatinginthebeach.Smallandfragilespeciesaredifferentiallydestroyed,thusstrongshellssuchasthoseofBuccella peruviana,Ammonia beccariiandElphidium discoidalebecomedominantinuppershorefaceandback-shorefacies.Taphonomicmodificationsoftestarenotuniformlydistributedalongthebathymetricgradient,whichallowstherecognitionoflowershoreface,uppershorefaceandforeshore/backshore/foreduneassemblages,indicatingthattaphonomicprocessesaremainlydeterminedbywaterdepth,anddependmainlyonwaterenergy,aerialexposure,andbeachdynamics,thusshowinghighpotentialtobeusedinpaleoenvironmentalanalyses.
TaxonomiccompositionandtaphonomicsignaturesofHoloceneassemblagesdonothavestricthomologousbetweenmodernsamples,indicatingthatHolocenebeachdynamicswereradicallydifferentfromthoseoperatinginthepresent.However,basedontaxonomiccompositionandsometaphonomicsignatures,wecanassertthattheKM187successionwasdepositedinuppershoreface-backshoreenvironments,probablyrelatedtostormsurgesinawave-dominatedcoast.Inaddition,taphonomicanalysisshowsthatsedimentsourcesofHolocenebeachesofthesouthernSamborombón(proto)bay includedLatePleistocene-EarlyHolocenecoastalandmarginalmarineenvironmentsstronglysubjectedtoreworkingduringtheMid-to-LateHolocenesealevelfall.

Modern foraminifera from coastal settings, northern Argentina 63
REFERENCES
Aguirre,M.L.,Whatley,R.C.,1995,LateQuaternarymarginalmarinedepositsandpalaeoenvironmentsfromnortheasternBuenosAiresProvince,Argentina:Areview:QuaternaryScienceReviews,14(3),223-254.
Albertzart,L.S.,Wilkinson,B.H.,1990,BarrierBackbeachShellAssemblagesfromtheCentralTexasGulfCoast:PALAIOS,5(4),346-355.
Alve,E.,MurrayJ.W.,1999,Marginalmarineenvironmentsof theSkagerrakandKattegat:abaselinestudyofliving(stained)benthicforaminiferalecology:Palaeogeography,Palaeoclimatology,Palaeoecology,146(1),171-193.
Barbieri,R.,1996,Syndepositionaltaphonomicbiasinforaminiferafromfossilintertidaldeposits,ColoradoDelta(BajaCalifornia,Mexico):JournalofForaminiferalResearch,26(4),331-341.
Bertels-Psotka,A.,Laprida,C.,1998,Ostracodos(Arthropoda,Crustacea)delMiembroCerrode laGloria,FormaciónLasEscobas(Holoceno),provinciadeBuenosAires,RepúblicaArgentina:RevistaEspañoladeMicropaleontología,30(1),103-128.
Boltovskoy,E.,1954a,ForaminíferosdelgolfoSanJorge:RevistadelInstitutoNacionaldeInvestigacióndelasCienciasNaturalesyMuseoArgentinodeCienciasNaturales“BernardinoRivadavia”,3(3),85-246.
Boltovskoy,E.,1954b,ForaminíferosdelabahíaSanBlas:RevistadelInstitutoNacionaldeInvestigacióndelasCienciasNaturalesyMuseoArgentinodeCienciasNaturales“BernardinoRivadavia”,3(4),247-300.
Boltovskoy,E.,1957,LosforaminíferosdelestuariodelRíodelaPlataysu zona de influencia: Revista del Instituto Nacional de Ciencias Naturales,Geología,6(1),1-76.
Boltovskoy,E.,1958,TheforaminiferafaunaoftheRíodelaPlataanditsrelationtotheCaribbeanarea:CushmanFoundationofForaminiferalResearch,Contribution9(1),17-21.
Boltovskoy,E.,1970,DistributionofthemarinelittoralForaminiferainArgentina,UruguayandSouthernBrazil:MarineBiology,6(4),335-344.
Boltovskoy,E.,1976,DistributionofRecentforaminiferaoftheSouthAmerican Region, in Hedley, R.H.,Adams, C.G. (eds.),Foraminifera2:London,AcademicPress,171-236.
Boltovskoy,E.,Lena,H.,1969,Seasonaloccurrences,standingcropandproductioninbenthicforaminiferaofPuertoDeseado:CushmanFoundationforForaminiferalResearch,Contribution20(3),87-95.
Boltovskoy,E.,Totah,V.,1985,Diversity,similarityanddominanceinbenthicforaminiferalfaunaalongonetransectoftheArgentineshelf:RevuedeMicropaléontologie,28(1),23-31.
Boltovskoy,E.,Wright,R.,1976,RecentForaminifera:TheHague,Netherlands,Dr.W.Junkb.v.Publishers,515pp.
Boltovskoy,E.,Giusani,G.,Watanabe,S.,Wright,R.,1980,AtlasofbenthicshelfForaminiferaoftheSouthwestAtlantic:TheHague,Netherlands,Dr.Junkb.v.Publishers,147pp.
Brandt, D.S., 1989, Taphonomic Grades as a Classification for Fossiliferous AssemblagesandImplicationsforPaleoecology:Palaios,4(4),303-309.
Buzas,M.A.,1990,Another lookatconfidence limits forspeciesproportion:.JournalofPaleontology,64(5),842-843.
Callender,W.R., Powell, E.N., Staff, G.M., Davies, D.J., 1992,Distinguishingautochthony,parautochthonyandallochthonyusing taphofaciesanalysis:Cancoldseepassemblagesbediscriminatedfromassemblagesofthenearshoreandcontinentalshelf?:Palaios,7(4),409-421.
Closs,D.,1963,ForaminiferosetecamebasdaLagoadosPatos(RGS):PortoAlegre,Brasil,UniversidadeFederaldoRioGrandedoSul,EscoladeGeologiadoPortoAlegre,Boletim11,1-130.
Closs,D.,Barberena,M.C.,1962,ForaminiferosRecentesdaspraiasdolitoralsulbrasileiro.1.ArroioChuí(R.G.S.)–Araranguá(S.C.):PortoAlegre,Brasil,UniversidadeFederaldoRioGrandedoSul,InstitutodeCiênciasNaturais,Boletim16,7-55.
Closs,D.,Lopes-Madeira,M.L.,1968,Seasonalvariationsofbrackish
foraminiferidaintheLosPatoslagoon,southernBrazil:SãoPaulo,Brasil,UniversidadeFederaldoRioGrandedoSul,EscoladeGeologia,PublicaçãoEspecial1,1-151.
Codignotto,J.O.,Aguirre,M.,1993,Coastalevolution,changesinsealevelandmolluscanfaunainnortheasternArgentinaduringtheLateQuaternary:MarineGeology,110(1-2),163-175.
Cole,W.S.,1931,ThePlioceneandPleistoceneforaminiferaofFlorida:Florida,U.S.A.,FloridaStateGeologicalSurvey,Bulletin6,79pp.
Corliss,BH.,Honjo,S.,1981,DissolutionofDeep-SeaBenthonicForaminifera:Micropaleontology,27(4),356-378.
Cushman,J.A.,1922,Shallow-waterforaminiferaoftheTortugasRegion:Washington,CarnegieInstitution,DepartmentofMarineBiology,papers17,1-85.
Cushman,J.A.,Kellett,B.,1929,RecentforaminiferafromwestcoastofSouthAmerica:U.S.NationalMuseumProceeding,75(25),11.
Denne,R.A.,SenGupta,B.K.,1989,EffectsoftaphonomyandhabitatontherecordofbenthicforaminiferainModernsediments:PALAIOS,4(5),414-423.
Diz,P.,Francés,G.,2009,PostmortemprocessesaffectingbenthicforaminiferalassemblagesintheRíadeVigo,Spain:implicationsforpaleoenvironmentalstudies:JournalofForaminiferalResearch,39(3),166-179.
D´Orbigny,A.,1839a,Foraminiféres,indelaSagra,R.(ed.),Histoirephysique,politiqueetnaturelledel´IlledeCuba:Paris,ArthusBertrand,224pp.
D’Orbigny,A.,1839b,Voyagedansl’Amériqueméridionale,Foraminifères:Strasbourg,P.Bertrand,5,1-86.
D´Orbigny,A.,1846,ForaminiféresfósilesdubassinTertiaredeVienne(Austriche):Paris,GideetCompagnie,312pp.
Fidalgo, F., de Francesco, F.O., Pascual, R., 1975, Geología superficial delallanurabonaerense,in6ºCongresoGeológicoArgentino,BahíaBlanca,BuenosAires,Argentina:Argentina,AsociaciónGeológicaArgentina,Relatorio,103-138.
Fürsich,F.T.,Oschmann,W.,1993,Shellbedsastoolsinbasinanalysis:theJurassicofKachchh,westernIndia:JournaloftheGeologicalSociety,150(1),169-185.
Galloway,J.J.,Wissler,S.G.,1927,PleistoceneforaminiferafromtheLomitaQuarry,PalosVerdesHills,California:JournalofPaleontology,1(1),35-87.
Goldstein, S.T.,Watkins, G.T., 1999,Taphonomy of salt marshforaminifera:anexamplefromcoastalGeorgia:Palaeogeography,Palaeoclimatology,Palaeoecology,149(1),103-114.
Hammer,Ø.,Harper,D.A.T.,RyanP.D.,2001,PAST:PaleontologicalStatisticsSoftwarePackageforEducationandDataAnalysis:PalaeontologiaElectronica,4(1),1-9.
Herrera-Moreno, A., 2000, La clasificación numérica y su aplicación en la ecología:RepúblicaDominicana,InstitutoTecnológicodeSantoDomingo,88pp.
Hippensteel,S.P.,Martin,R.E.,1999,Foraminiferaasanindicatorofoverwashdeposits,BarrierIslandsedimentsupply,andBarrierIslandevolution:FollyBeach,SouthCarolina:Palaeogeography,Palaeoclimatology,Palaeoecology,149(1),115-125.
Kornfeld,M.M.,1931,RecentlittoralForaminiferafromTexasandLouisiana:Stanford,California,U.S.A.,StanfordUniversity,DepartmentofGeology,Contributions1(3),77-101.
Lamarck,J.B.,1804,SuitedesMémoiressurlesfossilsdesénvironsdeParis:Paris,France,MuséumNationald´HistoireNaturelle,Annales5(a),179-188.
Laprida,C.,1997,Micropaleontología(OstrácodosyForaminíferos)ypaleoecologíadelCuaternarioTardíodelNordestedelaprovinciadeBuenosAires,RepúblicaArgentina:BuenosAires,Argentina,UniversidaddeBuenosAires,doctoralthesis,555pp.
Laprida,C.,1999,Micropaleontologicalassemblages(ForaminiferidaandOstracoda)fromLateQuaternarymarginalmarineenvironments(DestacamentoRíoSaladoFormation),SaladoBasin,Argentina:RevuedePaléobiologie,17(2),461-478.
Laprida, C., Bertels-Psotka,A., 2003, Benthic foraminifera andpaleoecologyofaHoloceneshellyconcentration,SaladoBasin,Argentina:GEOBIOS,36(5),559-572.

Laprida et al.64
Linné,C.,1758,SystemaNaturaeperregnatrianaturae:secundumclasses,ordines,genera,species,cumcharacteribus,diferentiis,synonimis,locis:Stockholm,Holmiae(LaurentiiSalvii),10thedition,1,823pp.
Loeblich,A.,Tappan,H., 1988,ForaminiferalGenera andTheirClassifications: New York, Van Nostrand Reinhold Company, 970pp.
Loubere,P.,Gary,A.,Lagoe,M.,1993,Generationof thebenthicforaminiferalassemblage:theoryandpreliminarydata:MarineMicropaleontology,20(3-4),165-181.
Martin,R.E.,1999,Taphonomy.AProcessApproach:Cambridge,CambridgeUniversityPress,508pp.
Mendes,I.,González,R.,Dias,J.M.A.,Lobo,F.,Martins,V.,2004,Factors influencing recent benthic foraminifera distribution on the Guadianashelf(SouthwesternIberia):MarineMicropaleontology,51(1-2),171-192
Montagu,G.,1803,TestaceaBritannica,orNaturalHistoryofBritishshells,marine,landandfresh-water,includingmostminute:Romsey,GreatBritain,J.S.Hollis,606pp.
Murray,J.W.,2006,EcologyandPalaeoecologyofBenthicForaminifera:CambridgeUniversityPress,Cambridge,426pp.
Murray,J.W.,Alve,E.,1999,Taphonomicexperimentsonmarginalmarineforaminiferalassemblages:howmuchecologicalinformationispreserved?:Palaeogeography,Palaeoclimatology,Palaeoecology,149(1),183-197.
Murray-Wallace,C.V.,Belperio,A.P.,Bouman,R.P.,Cann,J.H.,Price,D.M.,1999,Faciesarchitectureofalastinterglacialbarrier:amodelforQuaternarybarrierdevelopmentfromtheCoorongtoMountGambierCoastalPlain,SoutheasternAustralia:MarineGeology,158(1),177-195.
Nábel,P.,1987,Estudiopaleomagnéticoysedimentológicodesedimentosdeplataforma,provinciadeBuenosAires:RevistadelaAsociaciónGeológicaArgentina,42(3-4),377-387.
Parker,G.,1979,GeologíadelaPlaniciecosteraentrePinamaryMardeAjó,ProvinciadeBuenosAires:RevistaAsociaciónGeológicaArgentina,34(2),83-167.
Parr,W.J.,1932,VictorianandSouthAustralianshallow-waterforaminifera.PartII:ProceedingsoftheRoyalSocietyofVictoria,44(1),p.226.
Peebles,M.W.,Lewis,R.D.,1991,SurfacetexturesofbenthicforaminiferafromSanSalvador,Bahamas:JournalofForaminiferalResearch,21(4),285-292.
Perillo,G.M.,1979,CálculodelvolumendesedimentosdelaplayafrontaleneláreadePuntaMédanos,ProvinciadeBuenosAires:ActaOceanográfica Argentina, 2(2), 31-55.
Phleger,F.B.,1960,EcologyanddistributionofRecentForaminifera:JohnHopkinsPress,Baltimore,297pp.
Rine,J.M.,Tillman,R.W.,Culver,S.J.,Swift,D.J.P.,1991,GenerationoftheLateHolocenesandridgesonthemiddlecontinentalshelfofNewJersey,U.S.A.–Evidenceforformationinamiddleshelfsettingbasedoncomparisonwithanearshoreridge, in Swift,D.J.P.,Oertel,G.,Tillman,R.W.(eds.),ShelfSandandSandstoneBodies:Origin,FaciesandDistribution:NewYork,InternationalAssociationofSedimentologists,SpecialPublication1(14),359-423.
Rothfus,T.A.,2004,HowManyTaphonomistsSpoiltheData?MultipleOperatorsinTaphofaciesStudies:Palaios,19(5),514-519.
Schafer,C.T.,1971,Samplingandspatialdistributionofbenthonicforaminifera:LimnologyandOceanography,16(6),944-951.
Scott,D.B.,Medioli,F.S.,1980,Livingvs.totalforaminiferalpopulations:theirrelativeusefulnessinpaleoecology:JournalofPaleontology,54(4),814-831.
Scott,D.B.,Medioli,F.S.,Schafer,C.T.,2001,MonitoringofCoastalenvironmentsusingForaminiferaandThecamoebianindicators:UnitedKingdom,CambridgeUniversityPress,176pp.
ServiciodeHidrografíaNaval,2001,TablasdeMarea.CostaArgentina,incluyendoAntártidaArgentinayprincipalespuertosdeBrasil,UruguayyChile:BuenosAires,Argentina,ServiciodeHidrografíaNaval,PublicaciónNáuticaH-610.
Spalletti,L.,Mattheos,S.,Poiré,D.,1987,SedimentologyoftheHolocenelittoralridgesatSamborombómBay(centralBuenosAiresProvince,Argentina),inRabassa,J.(ed.),QuaternaryofSouthAmericaandAntarcticPeninsula:Rotterdam,A.A.Balkema,5,111-132.
Speroni,J.O.,Dragani,W.C.,Mazzio,C.A.,1999,ObservacionescosterasenMardeAjó,Pcia.deBuenosAires,descripcióndelambientelitoral:BuenosAires,Argentina,ServiciodeHidrografíaNaval,ReporteTécnico102/99,19pp.
Sunamura,T.,1988,Beachmorphologiesandtheirchange,inHorikawa,K.(ed.),NearshoreDynamicsandCoastalProcesses:Tokyo,UniversityofTokyo,136-157.
Taylor,M.,Stone,G.W.,1996,Beach-Ridges:AReview:JournalofCoastalResearch,12(3),612-621.
Terquem,O.,Terquem,E.,1886,Foraminifèresetostracodesdel´IslandeetduSuddelaNorvège:BulletindelaSociétéZoologiquedeFrance,11,328-339.
Tonni,E.P.,Fidalgo,F.,1978,ConsideracionessobreloscambiosclimáticosduranteelPleistocenoTardío-RecienteenlaprovinciadeBuenosAires.Aspectosecológicosyzoogeográficosrelacionados:Ameghiniana,15(1–2),235-253.
Wiesner,H.,1912,ZürSystematikadriatischerNubecularien,Spiroloculinen,MiliolinenundBiloculinen:ArchivfürProtistenkunde,25(2),201-239.
Williamson,W.C.,1858,OntheRecentforaminiferaofGreatBritain:London,TheRaySociety,108pp.
Manuscriptreceived:March17,2009Correctedmanuscriptreceived:July3,2010Manuscriptaccepted:September9,2010