Embed Size (px)
Citation preview

4Industrialization of Functional Mouse Genomics Technologiesfor Biopharmaceutical Drug Discovery and DevelopmentPeter Stadler, Jost Seibler, and Frieder Schwenk
4.1Introduction
The big challenge for pharmaceutical industry is to increase the number of newlylaunched blockbuster products per year to meet its requirements for revenuegrowth. To address this problem, pharmaceutical companies have focused on theinvestigation of differentially expressed genes to mine the human genome for newdrug target candidates in the 1990s of the last century. As it turned out, informationabout gene sequence and protein structure alone is not sufficient to be used as anefficient basis for drug target selection since it does not provide knowledge aboutgene function relative to a selected pathomechanism. Consequently, “functionalgenomics” was introduced as a powerful tool to overcome the previously describedlimitations of “structural genomics.” “Forward genetic screens” using genetic modelorganisms such as nematode worm, fruit fly, and zebra fish are used in pharma-ceutical research to identify and validate disease-related genes based on phenotypesof randomly mutagenized organisms. See a detailed review of different animalmodels and their use in (bio)pharmaceutical development by the editor elsewhere inthis book. In addition, “reverse genetics” in mice is broadly applied to furthervalidate the function of candidate genes in higher model organisms reflectinghuman physiology as closely as possible. We consider this strategy to be presentlythe most promising approach for filtering out those targets, which have a higherchance to enable the identification of candidate compounds that will finally make itinto successful clinical development [1].Among the most important technologies in advanced mouse genomics are
conditional gene knockout (inducible and/or tissue specific) and gene knockdownvia expression of interfering ribonucleic acid (RNAi) in transgenic animals for targetvalidation, including the validation on a disease background. Another importantapproach is the exchange of mouse genes by their human orthologs (“humanizationof mice”) for testing of drug target candidates that do not interact with mouseproteins but only with their human counterparts. Those models also allowperforming preclinical studies of humanized drug metabolism.
Modern Biopharmaceuticals: Recent Success Stories, First Edition. Edited by Jörg Knäblein.# 2013 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2013 by Wiley-VCH Verlag GmbH & Co. KGaA.
j77

While only a couple of years ago, the generation of a genetically modified mousemodel required �24 months, the time required today is less than half. Functionalanalysis of genes via RNAi in transgenic mice is meanwhile even possible in notmore than five months. Based on the expression of short hairpin RNA (shRNA),novel methodologies have been developed that will, for the first time, allow theswitching off and subsequently switching on again of selected genes in an adultmouse. These recently achieved major improvements in mouse genomics technol-ogy are based on optimization and a methodological redesign of every single step ofthe transgenic mice generation process, as well as on the development andintroduction of novel scientific methods. The result is a genetic and genomictoolbox, which is now widely used in an efficient way for biopharmaceutical andacademic research on an increasingly broader basis.
4.2The Mouse Genetics Story
Among the genetic model organisms, the laboratory mouse (Mus musculus) has apredominant role in the study of human disease and preclinical drug development.Apart from the high degree of sequence homology of mouse and human genomes,as well as similarities in many physiological aspects, advanced targeting technol-ogies make the crucial difference, providing unique tools for elucidating genefunction in vivo. The ability to manipulate the genome in embryonic stem (ES) cellsand mice was developed in the late 1980s. Since then, gene targeting has beenextensively used to study gene function in genetically modified mouse strains. Asinitially developed, the technique allows the disruption of a target gene in themurine germline by the insertion of a selectable marker. The vast majority ofpresently �4000 existing “knockout” (KO) mouse models have been createdfollowing this design. Many of these KO strains have given valuable informationon human physiology and disease processes. As “conventional” KOmice are usuallyhomozygous for a null allele in the germline, they provide an appropriate model forinherited diseases, leading to embryonic or early postnatal lethality in about 30% ofcases. Beyond this application, germline KO mice do not necessarily represent thebest technical approach for drug target validation and to study other aspects of genefunction in vivo, in particular, in adult mice. In many cases, the phenotype obtainedin a conventional KO strain reflects a developmental defect rather than genefunction in the mature organ. In addition, other gene products can compensatethe activity of a continuously inactivated gene, thereby veiling the KO phenotype.A refinedKO strategy, termed “conditional gene targeting,” has been developed that
permits target gene inactivation to be restricted to a particular organ and/or develop-mental stage. This is achieved by the expression of the site-specific deoxyribonucleicacid (DNA) recombinaseCre in conjunctionwith the introduction of two recombinaserecognition sequences (loxP) into the genomic locus of interest. These sites are usuallyplaced into introns such that recombination results in gene inactivation through thedeletion of the loxP flanked exon(s), whereas the unrecombined allele is fully active.
78j 4 Industrialization of Functional Mouse Genomics Technologies

Since the initial demonstration of this technology in 1994 [2], an increasing number ofconditional knockout experiments has been published, most of which employedtissue-specific promoters to control Cre expression. Spatially regulated geneinactivation has proven to be powerful in dissecting the role of different cell typesin a physiological process. Themethod of choice for precise gene function analyses inadult mice, however, is the temporal control of gene inactivation as it can preventimpaired embryonic development until the time of induction. Furthermore, itpermits the investigation of the effect of gene inactivation after the onset of a chronicor acute disease phenotype, simulating the activity of antagonistic drugs. This aspectis of particular interest for the validation of potential drug target genes inbiopharmaceutical drug development.
4.3Establishing Inducible Gene Targeting Tools
The first attempts to establish an inducible gene targeting approach were initiated byour scientific advisor Klaus Rajewsky (until 2001: University of Cologne, Germany;from 2002–2011HarvardMedical School Boston, MA; now:Max-Delbrück-Centrumfür Molekulare Medizin (MDC) Berlin-Buch in 1990. At that time, Cre was placedunder the control of the interferon-a/b inducible promoter of the mouse Mx1gene [3]. Mx, part of the defense to viral infections, is silent in healthy mice buttransiently activated to high levels of transcription upon administration of inter-feron. Mx-cre transgenic mice were tested using a loxP-flanked segment of the DNApolymerase � gene (one of the few “floxed” alleles that were available at this time).The success of this experiment was surprisingly resounding: almost completedeletion of the loxP-flanked target was achieved in the liver and lymphocytes withintwo days of interferon treatment, demonstrating for the first time that induced Cre-mediated deletion can proceed both rapidly and efficiently even in an organcomposed mainly of resting cells [3]. However, a significant degree of backgroundrecombination was detectable in these experiments, presumably because of theendogenous interferon generation. In addition, not all tissues responded equallywell to the inductor, limiting this system to particular biological questions [3].When Artemis was founded in 1998, the primary goal was to provide rapid,
efficient, and sophisticated techniques for drug development research. In particular,techniques for inducible gene targeting in all tissues of the mouse were highlydesired by our pharmaceutical collaboration partners for in vivo drug target valida-tion. Inducible mutagenesis in mice using site-specific recombinases requires, forprecision, a near complete absence of background recombination and a high-levelrecombinase activity during the short time of induction, mediating complete targetinactivation in all tissues. To accomplish these two objectives, we employed a fusionprotein consisting of Cre and part of the human estrogen receptor (ER). In theabsence of hormone, ERs are bound by heat shock proteins, which inactivatethe recombinase, presumably by sterical hindrance. The receptor is activated bythe addition of hormone that releases the heat shock proteins from the fusion
4.3 Establishing Inducible Gene Targeting Tools j79

protein. To avoid activation through endogenous steroids, we used a mutantER (ERT2) [4], which was supposed to be unresponsive to their natural ligandbut can be activated by synthetic hormone antagonists such as tamoxifen. TheCreERT2 coding region was inserted into the ubiquitously expressed Rosa26 locusthrough homologous recombination to obtain a configuration that allows expressionof CreERT2 in every single cells of themouse. By this strategy we expected an optimalCre expression since all control elements driving the transgene are present at theirnatural chromosomal environment. Upon induction of transgenicmice, the Rosa26-CreERT2 configuration achieved nearly complete inducible excision of a loxP-flankedgene segment in most organs, without detectable background recombination in theabsence of inductor. Thus, our system permitted, for the first time, a tight temporalcontrol of ubiquitous gene inactivation allowing precise gene function and drugtarget analyses in animals that have undergone normal embryonic development. Inthemeantime, the Rosa26-CreERT2
finds broad application in gene function analysisstudies by our customers in both academia and industry.The generation of genetically modified mouse models is a complex process,
combining a variety of disciplines such as molecular genetics, ES cell culture,microinjection techniques, and animal breeding. The integration of these differenttechniques and expertise in a streamlined process is extremely demanding. Multipleimprovements in project management and technology have to be implemented toensure quality and robustness of the industrial mouse generation process.The generation of a conditional gene targeting vector requires particular caution
since anymutation of the regulatory or coding regionmust be excluded, as itmay altertarget gene expression before Cre/loxP-recombination. So far, the generation of suchvectorsbypolymerasechain reaction (PCR)amplificationofgenomicDNAand ligationof restriction fragments was hindered by the high incidence of unwanted mutations,requiring extensive sequencing and recloning of DNA constructs. This problem hasbeen addressed by introducing the technique of recombineering pioneered by ourscientific advisor Francis Stewart in 1998 [5].Here, large pieces ofDNAare engineeredthroughhomologous recombination in transformedbacteria expressing theRac phageprotein pair RecE/RecT. This approach allows the precise and rapid assembly ofcomplex targeting vectors without the requirement of PCR amplification and in vitrotreatments with restriction enzymes or DNA ligases. Implementing this scheme hasreduced the time and resources required for vector construction to less than half.Another crucial point in the mouse model generation pipeline is the handling of
ES cells. Here, mistreatment will easily lead to differentiation from their pluripotentstate. In addition, spontaneous mutations or chromosomal aberration can affect theES cell quality. Consequently, cellsmay become unable to participate in developmentand, therefore, to derive chimeras. To minimize the risk of failure, we rigorouslyrestricted the number of in vitro ES cell culture passages before genetic manipula-tion. This is facilitated by the constant derivation and validation of new ES cell linesfor the working stock. In addition, we integrated multiple quality checkpoints intothe cell culture process, including mycoplasma control, PCR-based virus testing,and high-resolution karyotyping using multicolor fluorescent in situ hybridization(FISH). Finally, we implemented cell culture robotics for high throughput and
80j 4 Industrialization of Functional Mouse Genomics Technologies

contact-free ES cell handling, minimizing departure from optimal treatment. Owingto these measures, the quality of genetically modified ES lines has reached anoptimal level, supporting efficient mouse generation.The animal housing also needs particular notice to guarantee optimal breeding
condition, the well-being of animals, and to exclude any infection with pathogen. Wehave therefore sectioned our facilities into two physically separated barrier units,each supplied by its own “high-efficiency particulate air” (HEPA)-filtered air con-ditioning. In addition, all mice are housed in isolated ventilated cages (IVC),providing optimal husbandry at highest hygienic standard. The “specified patho-gen-free” (SPF) status of our animal facility is constantly monitored by an elaboratehealth-screening program with sentinel animals, exceeding the recommendationsof International Laboratory Animal Science Associations Federation of EuropeanLaboratory Animal Science Associations (FELASA) and Association for Assessmentand Accreditation of Laboratory Animal Care International (AAALAC). Theseprecautions are essential to meet the customer’s expectations in regard to qualityand health standard of animal models.The overall mouse generation process is guided by the proprietary TaconicArtemis
MousehouseDatabaseTM. AnyDNAvector receives a unique identification (ID) that isrecorded together with molecular details in this database. Likewise, each ES cellmanipulation, blastocyst injection, and embryo transfer experiment, as well as eachmouse inour facility obtains aunique IDnumber that is assigned to the correspondingproject. All information generated during the course of a project is linked in thedatabase, allowing automatic generation of updated project reports and milestonedocumentations at the push of a button. These features are available to support theinternal as well as the customer’s project management (Figure 4.1).
4.4RNAi – Talking About a Revolution?
RNA interference (RNAi) has been discovered some years ago as a tool for inhibitionof gene expression and the 2006 Nobel Prize for physiology/medicine was awardedto Andrew Z. Fire and Craig C. Mello “for their discovery of RNA interference – genesilencing by double-stranded RNA” [6,7].It is based on the introduction of molecules into cells, whereby one strand is
complementary to the coding region of a target gene. Through pairing of the specificmRNA with the introduced RNA molecule, the mRNA is degraded by a cellularmechanism. Since long double-stranded RNA (dsRNA) provokes an interferonresponse in mammalian cells, the technology was initially restricted to organismsor cells showing no interferon response. The finding that short interfering RNAs(siRNAs) (<30 bp) circumvent the interferon response extended the application tomammalian cells [8]. Sustained gene silencing has been achieved by the intracellulartranscription of small inverted repeats separated by a spacer region [7,6,9,10].For the expression of these short hairpin RNAs (shRNAs), the RNApolymerase III-
dependent promoters U6 and H1 have been used in most cases. These promoters
4.4 RNAi – Talking About a Revolution? j81

produce small transcripts lacking nonfunctional bases at the 50 and 30 ends. Theresulting shRNAs are processed in vivo into active “small interfering” RNAs.Given that RNAi-mediated gene knockdown have been reported to reach effi-
ciencies of 80% or more in mammalian cells, the technology promised greatpotential for gene function analysis in mice. Compared to the existing KO technol-ogy, RNAi requires just a single transgene to silence both alleles of the target gene.Furthermore, the effort for generating shRNA transgenes is much lower comparedto the construction of gene targeting vectors.
Figure 4.1 Streamlined mouse modelgeneration pipeline. The generation ofgenetically engineered mouse models is basedon optimized “standard operation procedures”(SOPs) and guided by multiple qualitycheckpoints to ensure quality of the process.Targeting vectors are precisely assembled usingimproved ET cloning techniques. Cell culturerobotics are implemented for high throughputand contact-free ES cell handling. Transgenic EScell clones are routinely tested using PCR-basedvirus assays and high-resolution karyotyping
using multicolor fluorescent in situhybridization. Mouse models are housed inisolated ventilated cages providing optimalbreeding conditions. The “specified pathogen-free” (SPF) status of our animal facility isconstantly monitored by an elaborate health-screening program. The overall mousegeneration process is traced by the proprietaryTaconicArtemis Mousehouse DatabaseTM,allowing automatic generation of updatedproject reports and milestone documentations.
82j 4 Industrialization of Functional Mouse Genomics Technologies
Sustained, shRNA-mediated RNAi in mice had been demonstrated in recentpublications using random transgenesis via pronuclear injection [11] and transfectionof plasmid-based shRNA constructs, as well as lentiviral infection [12–14]. Dependingon the site of transgene integration, the efficiency of gene silencing in those animalswas variable because of the inherent problem of position-dependent effects onexpression. Thus, a thorough characterization of the resulting mouse lines wasrequired to determine the potency of RNAi in all relevant organs, which is a time-consuming and laborious undertaking. To avoid these complications, we tried toidentify a genomic locus that allows strong and predictable expression of shRNAtransgenes inserted through homologous recombination. Again, we chose the Rosa26locus for this purpose, inspired by the impressing results of ourRosa26-CreERproject.The Rosa26 locus is ubiquitously expressed during development and in adult miceindicating anopen chromatin surroundingenablingpolymerase III-dependent expres-sion of shRNAs. Analysis of ES cell-derivedmice revealed that our single-copy shRNAconfiguration indeedmediates body-wide gene silencing with high efficiency, rangingbetween 80% and 95% knockdown in most organs [14]. This technology brings
600
Blo
od g
luco
se le
vels
(m
g/dl
)
500
400
300
200
100
00 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 19 21 22 23 24 25 28 29 30 31 days
+ + + + + + + + + + - - - - - - - - - - - - - - - - - dox
H1-tetO shRNAP itetRtet R
H1-tetO shRNAP itetR
tet R
+ Dox
- Dox
(a)
(b)
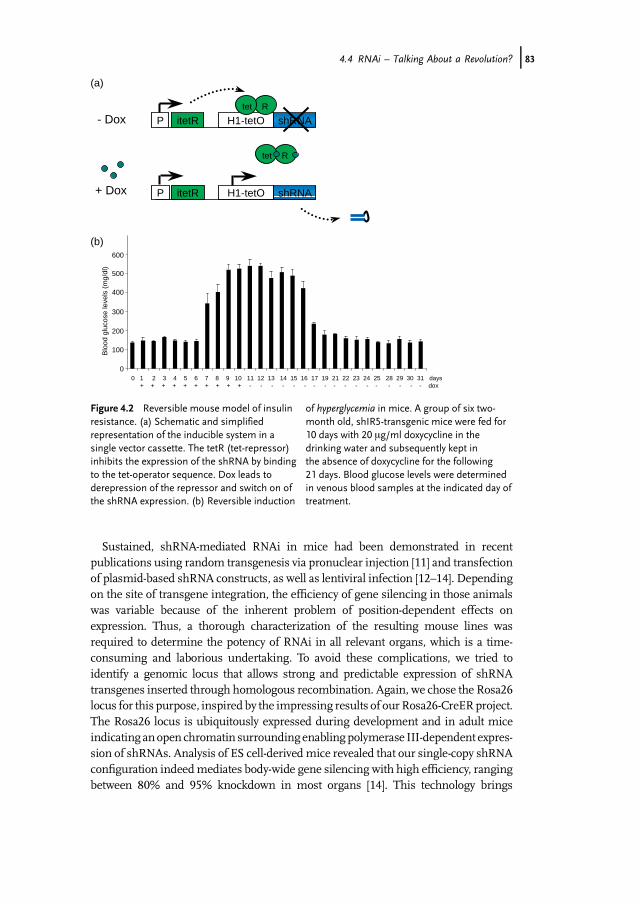
Figure 4.2 Reversible mouse model of insulinresistance. (a) Schematic and simplifiedrepresentation of the inducible system in asingle vector cassette. The tetR (tet-repressor)inhibits the expression of the shRNA by bindingto the tet-operator sequence. Dox leads toderepression of the repressor and switch on ofthe shRNA expression. (b) Reversible induction
of hyperglycemia in mice. A group of six two-month old, shIR5-transgenic mice were fed for10 days with 20 mg/ml doxycycline in thedrinking water and subsequently kept inthe absence of doxycycline for the following21 days. Blood glucose levels were determinedin venous blood samples at the indicated day oftreatment.
4.4 RNAi – Talking About a Revolution? j83

TaconicArtemis into the unique position to perform in vivo knockdown studies for ourpharmaceutical partners with manageable resources at large scale.As discussed above, the method of choice for precise gene function analysis in
adult mice is the temporal control of gene silencing. Optimally, such a system shouldprovide a reversible gene switch in every single cell of the body upon administrationof the inductor. In cultured cell lines, inducible expression of antisense or shorthairpin RNAs has been achieved using engineered, RNA polymerase III-dependentpromoters containing operator sequences (tetO) of the Escherichia coli tetracyclineresistance operon. Binding of the tetracycline resistance operon repressor (tetR)to tetO blocks transcription, whereas derepression is achievable by adding theinductor doxycycline (dox), causing release of TetR (Figure 4.2).We have recently shown that a codon-optimized tet repressor, but not the wt coding
region as employed in previous attempts, facilitates tight control of the H1 promoterwhenintegratedat therosa26locus [15].Wefoundefficient,dox-induciblegenesilencingwithin 10 days of doxycycline treatment, reaching 80–90% repression in most tissues.As expected, the system appeared to be reversible upon withdrawal of the inductor.From our point of view, RNAi indeed revolutionized the generation process as well
as the quality of genetically modified mouse models to an unexpected extent. Still,the RNAi field is rapidly moving and technologies need continuous improvement toassure state-of-the-art technologies. Current projects at TaconicArtemis include thetissue-specific activation of shRNA vectors and the simultaneous expression ofmultiple shRNA transgenes from a single locus (Figure 4.3).
4.5Further Shortening the Generation Timeline for RNAi Mouse Models
The standard protocol to derive mouse mutants currently requires the generation ofgermline chimeras from heterozygous-targeted embryonic stem (ES) cells, followedby at least one breeding step for germline transmission. Even the generation ofRNAi mouse models is still a time-intensive task, exceeding nine months before theanalysis of adult mutants. Owing to these extensive timelines, the impact oftransgenic mouse models in pharmaceutical research was limited for a longtime, creating a demand for a timesaving single-step procedure.To adapt the timelines to our customer’s needs, we established the generation of
RNAi-based mouse models through the introduction of diploid ES cells intotetraploid blastocysts. The latter provide an initial host environment for thedifferentiation of ES cells, but do not contribute to the embryo at later developmentalstages. Although the methodology to produce such “ES cell-tetraploid” (ES) micefrom inbred ES cell lines had already been described in 1990 [16], its application waslimited because of the extremely low frequency at which viable ES pups arerecovered. In the meantime, however, the technology was significantly improvedthrough the discovery of our scientific advisor Rudolf Jaenisch (Whitehead Institute,Cambridge, MA) that ES cell lines derived from hybrid mouse strains support thedevelopment of viable ES mice at 50-fold higher rate as compared to inbred ES cells
84j 4 Industrialization of Functional Mouse Genomics Technologies

[17]. Thus, ES mouse technology offered the opportunity to efficiently producetargeted mouse mutants directly from hybrid ES cell clones within the time of asingle mouse generation and without the requirement of further breeding. Wesuccessfully established the routine generation of ES mice using geneticallymodified ES cell clones, thereby saving three months for the mouse modelgeneration.To further speed up the generation of shRNA transgenic animals, we adapted
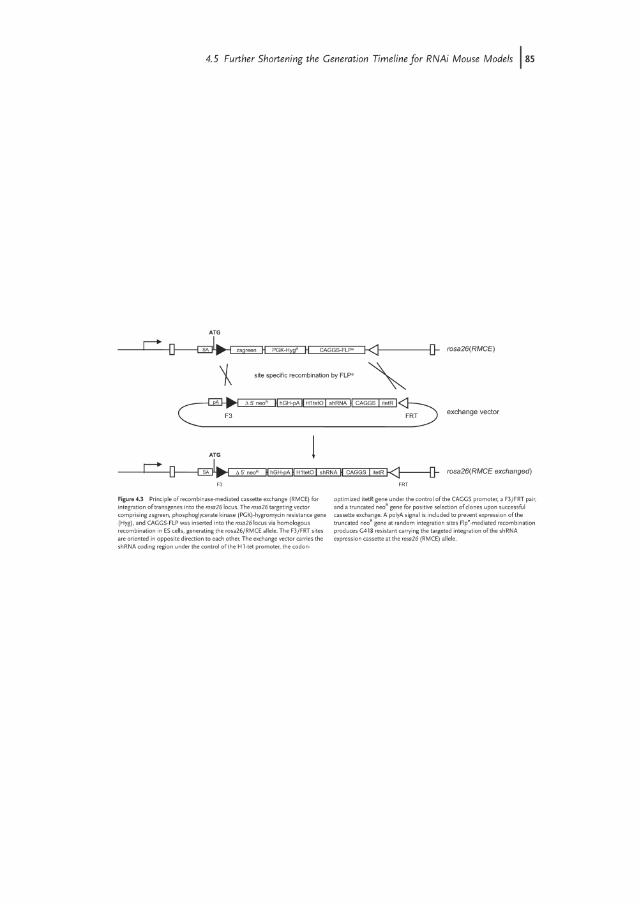
recombinase-mediated cassette exchange (RMCE) to the rosa26 locus. Here, theintegration of transgenes into a predefined locus is mediated by the site-specificDNA recombinase such as Cre or Flp. Although the RMCE method using the Flprecombinase has been already discovered in 1994 [18], so far only few examples ofsuccessful RMCE in ES cells had been described. In these experiments, randomintegration of the exchange vector as well as incomplete recombination frequentlyproduced unwanted transgene configurations. The efficiency of RMCE variedstrongly, depending on the choice of recombination sites, the selection strategy,and the chromosomal target. We combined a couple of the following features toaddress these problems (Figure 4.3):
1) We utilized Flp recombinase-mediated RMCE using a wild-type Flp target site(FRT) in combination with an inverted F3 site. The F3/F3 couple is recom-bined by FLP with the same efficiency as two wild-type recombinaserecognition sites (RRS), whereas recombination of a FRT/F3 pair is notcatalyzed. This characteristic contrasts other pairs of wild-type and mutantrecombinase recognition sites that were found to mediate a residualrecombination activity.
2) We added a constitutive FLP expression cassette on the targeting vector to providesufficient recombinase activity until successful RMCE of the exchange vector.Thus, incomplete recombination intermediates should be avoided.
3) A 50 truncated selection marker including a splice acceptor site but lacking afunctional promoter was inserted in the exchange vector. Expression of theselection marker should therefore be mediated by the endogenous rosa26promoter following successful RMCE, but not through random integration ofthe exchange vector.
We found that our RMCE strategy facilitates the insertion of shRNAtransgenes into the rosa26 locus with high efficiencies, consistently reachinga frequency of >90% correct clones [15]. Given the simple cloning strategy forthe insertion of shRNA transgenes into the RMCE exchange vector and thelimited screening efforts, our RMCE approach largely facilitates the generationof shRNA transgenic mice. By employing the tetraploid blastocyst comple-mentation approach for the direct generation of ES cell-derived mice, we havenow reached a timeline of less than five months for the generation of adultshRNA transgenic mouse models for our industrial partners (Figure 4.4).Moreover, the strategy should be applicable to other genetic model organismssuch as rat, in which homologous recombination is not yet established forroutine application.
86j 4 Industrialization of Functional Mouse Genomics Technologies

4.6Adapting the Mouse Genetics Toolbox for New Applications
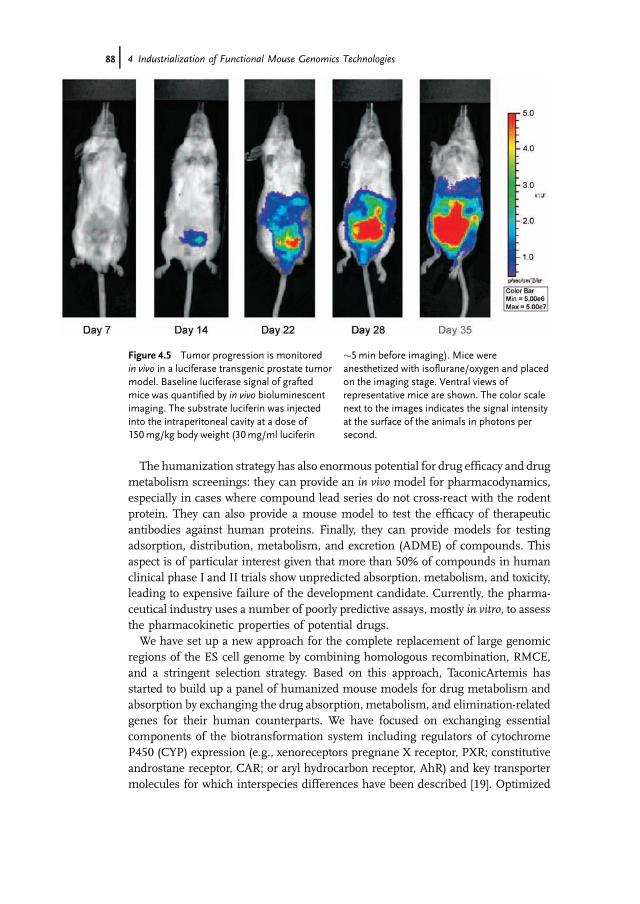
Beyond gene function analysis and drug target validation, genomic engineeringprovides a variety of additional opportunities to support the drug developmentprocess. In particular, the replacement of mouse genes by their human orthologshas opened up new avenues of research, where the genes and pathways altered inhuman disease can be reproduced in “humanized” animal models. In combinationwith our in vivo biophotonic imaging system for phenotypic characterization, theseimproved disease models provide powerful tools for preclinical drug testing,allowing the noninvasive studying disease progression in real time (Figure 4.5).
Figure 4.4 Generation of shRNA transgenicmouse models within less than five months.The targeted insertion of all elements requiredfor inducible shRNA expression into rosa26using RMCE, and the subsequent ES cellinjection using tetraploid blastocysts acceleratethe generation of conditional knockdown micesignificantly: four weeks for cloning of the
RMCE exchange vector, three weeks forgenerating targeted ES cell clones, two weeksfor verification of successful RMCE andexpansion of transgenic ES cell clones, andeight weeks for the generation of mice.Quantification of specific RNAi in cell lines andtissue samples is performed in parallel usingreal-time PCR.
4.6 Adapting the Mouse Genetics Toolbox for New Applications j87
The humanization strategy has also enormous potential for drug efficacy and drugmetabolism screenings: they can provide an in vivo model for pharmacodynamics,especially in cases where compound lead series do not cross-react with the rodentprotein. They can also provide a mouse model to test the efficacy of therapeuticantibodies against human proteins. Finally, they can provide models for testingadsorption, distribution, metabolism, and excretion (ADME) of compounds. Thisaspect is of particular interest given that more than 50% of compounds in humanclinical phase I and II trials show unpredicted absorption, metabolism, and toxicity,leading to expensive failure of the development candidate. Currently, the pharma-ceutical industry uses a number of poorly predictive assays, mostly in vitro, to assessthe pharmacokinetic properties of potential drugs.We have set up a new approach for the complete replacement of large genomic
regions of the ES cell genome by combining homologous recombination, RMCE,and a stringent selection strategy. Based on this approach, TaconicArtemis hasstarted to build up a panel of humanized mouse models for drug metabolism andabsorption by exchanging the drug absorption, metabolism, and elimination-relatedgenes for their human counterparts. We have focused on exchanging essentialcomponents of the biotransformation system including regulators of cytochromeP450 (CYP) expression (e.g., xenoreceptors pregnane X receptor, PXR; constitutiveandrostane receptor, CAR; or aryl hydrocarbon receptor, AhR) and key transportermolecules for which interspecies differences have been described [19]. Optimized
Figure 4.5 Tumor progression is monitoredin vivo in a luciferase transgenic prostate tumormodel. Baseline luciferase signal of graftedmice was quantified by in vivo bioluminescentimaging. The substrate luciferin was injectedinto the intraperitoneal cavity at a dose of150mg/kg body weight (30mg/ml luciferin
�5min before imaging). Mice wereanesthetized with isoflurane/oxygen and placedon the imaging stage. Ventral views ofrepresentative mice are shown. The color scalenext to the images indicates the signal intensityat the surface of the animals in photons persecond.
88j 4 Industrialization of Functional Mouse Genomics Technologies
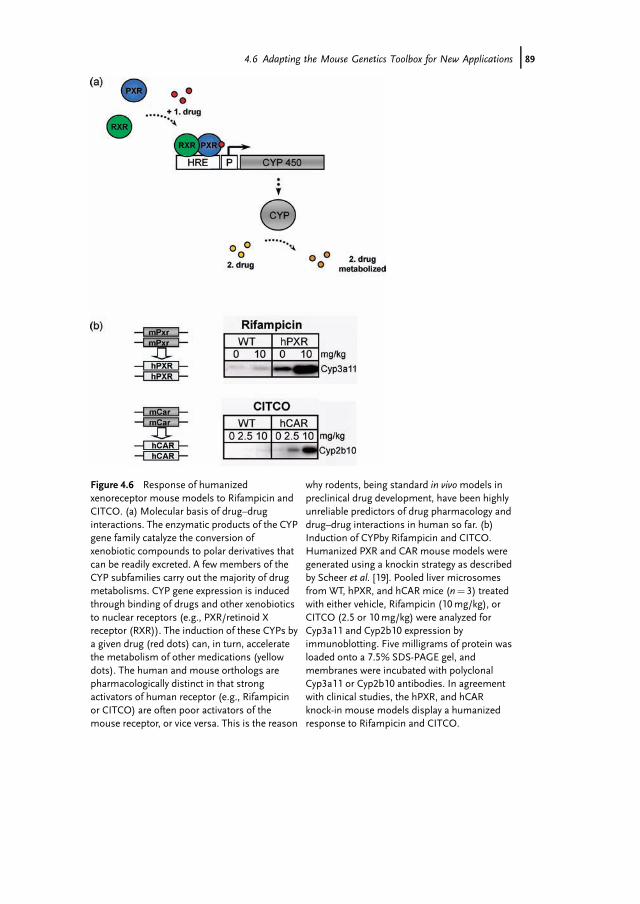
Figure 4.6 Response of humanizedxenoreceptor mouse models to Rifampicin andCITCO. (a) Molecular basis of drug–druginteractions. The enzymatic products of the CYPgene family catalyze the conversion ofxenobiotic compounds to polar derivatives thatcan be readily excreted. A few members of theCYP subfamilies carry out the majority of drugmetabolisms. CYP gene expression is inducedthrough binding of drugs and other xenobioticsto nuclear receptors (e.g., PXR/retinoid Xreceptor (RXR)). The induction of these CYPs bya given drug (red dots) can, in turn, acceleratethe metabolism of other medications (yellowdots). The human and mouse orthologs arepharmacologically distinct in that strongactivators of human receptor (e.g., Rifampicinor CITCO) are often poor activators of themouse receptor, or vice versa. This is the reason
why rodents, being standard in vivomodels inpreclinical drug development, have been highlyunreliable predictors of drug pharmacology anddrug–drug interactions in human so far. (b)Induction of CYPby Rifampicin and CITCO.Humanized PXR and CAR mouse models weregenerated using a knockin strategy as describedby Scheer et al. [19]. Pooled liver microsomesfrom WT, hPXR, and hCAR mice (n¼ 3) treatedwith either vehicle, Rifampicin (10mg/kg), orCITCO (2.5 or 10mg/kg) were analyzed forCyp3a11 and Cyp2b10 expression byimmunoblotting. Five milligrams of protein wasloaded onto a 7.5% SDS-PAGE gel, andmembranes were incubated with polyclonalCyp3a11 or Cyp2b10 antibodies. In agreementwith clinical studies, the hPXR, and hCARknock-in mouse models display a humanizedresponse to Rifampicin and CITCO.
4.6 Adapting the Mouse Genetics Toolbox for New Applications j89

models of human drug metabolism are created by combining several of suchmodified loci in a single mouse strain. In addition, we are currently generatingimproved reporter mouse models allowing to monitor CYP induction in ahumanized manner. We expect that these novel models will allow to assess thepharmacology of compounds in an in vivo setting, far more predictive, and muchearlier in the course of the research and development (R&D) process than currentassays (Figure 4.6).The rat is also an important animal model in biomedical research. However,
tools to generate loss-of-function mutation in rats are limited. In particular, genetargeting using homologous recombination in embryonic stem cells is still notestablished. To exploit the advantages of this model organism even so, we adaptedour inducible RNAi techniques to the rat using lentiviral infection and pro-nucleus injection strategies. Two recent publications illustrate the application ofinducible gene knockdown in rat for generating disease models [20,21]. Anotherpromising approach to quickly create transgenic rats are designed DNA nucle-ases (like zinc finger-, TALE- and Meganucleases Alfred Pingoud, George H.Silva, and Wolfgang Wende (2013)” “Precision Genome Surgery With Mega-nucleases: A Promising Biopharmaceutical for Gene Therapy” in “ModernBiopharmaceuticals – Recent Success Stories”, volume V. Editor: J. Kn€ablein.Publisher: Wiley-VCH. In press).Thus, the creation of animal models recapitulating human diseases will boost the
development of modern biopharmaceuticals and speedup the entire process.
References
1 See also Brakebusch, C. (2005) Geneticallymodified mice in medical andpharmaceutical research, inModernBiopharmaceuticals – Design, Developmentand Optimization, vol. 3, part V (ed. J.Kn€ablein), Wiley-VCH Verlag GmbH,Weinheim, pp. 1301–1320.
2 Gu, H., Marth, J.D., Orban, P.C.,Mossmann, H., and Rajewsky, K. (1994)Deletion of a DNA polymerase beta genesegment in T cells using cell type-specificgene targeting. Science, 265 (5168),103–106.
3 K€uhn, R., Schwenk, F., Aguet, M., andRajewsky, K. (1995) Inducible genetargeting in mice. Science, 269, 1427–1429.
4 Seibler, J., Zevnik, B., Kuter-Luks, B.,Andreas, S., Kern, H., Hennek, T., Rode,A., Heimann, C., Faust, N., Kauselmann,G. et al. (2003) Rapid generation ofinducible mouse mutants. Nucleic AcidsRes., 31, e12.
5 Zhang, Y., Buchholz, F., Muyrers, J.P., andStewart, A.F. (1998) A new logic for DNAengineering using recombination inEscherichia coli. Nat. Genet., 20 (2),123–128.
6 See also Boese, Q. and Khvorova, A.(2005) Rational siRNA design for RNAinterference: optimization fortherapeutic use, in ModernBiopharmaceuticals – Design, Developmentand Optimization, vol. 1, part I (ed. J.Kn€ablein), Wiley-VCH Verlag GmbH,Weinheim, pp. 243–264.
7 See also Bagowski, C. (2005) Targetvalidation: an important early step inpharmaceutical development, inModernBiopharmaceuticals – Design, Developmentand Optimization, vol. 2, part III (ed. J.Kn€ablein), Wiley-VCH Verlag GmbH,Weinheim, pp. 621–645.
8 Elbashir, S.M., Harborth, J., Lendeckel, W.,Yalcin, A., Weber, K., and Tuschl, T. (2001)
90j 4 Industrialization of Functional Mouse Genomics Technologies

Duplexes of 21-nucleotide RNAs mediateRNA interference in cultured mammaliancells. Nature, 411 (6836), 494–498.
9 See also Szymanski, M., Barciszewski, J.,and Erdmann, V.A. (2005) Nonprotein-coding RNAs and their potential asbiopharmaceuticals, inModernBiopharmaceuticals – Design, Developmentand Optimization, vol. 1, part I, (ed. J.Kn€ablein), Wiley-VCH Verlag GmbH,Weinheim, pp. 213–223.
10 See also Morris, K.V. and Rossi, J.J. (2005)Combinatorial RNA-based therapies forHIV-1, inModern Biopharmaceuticals –Design, Development and Optimization,vol. 2, part II, (ed. J. Kn€ablein), Wiley-VCHVerlag GmbH, Weinheim, pp. 569–577.
11 See also Hwang, W.S. (2005) The firstcloned human embryo: an unlimitedsource of stem cells, inModernBiopharmaceuticals – Design, Developmentand Optimization, vol. 1, part I (ed. J.Kn€ablein), Wiley-VCH Verlag GmbH,Weinheim, pp. 269–279.
12 Carmell, M.A., Zhang, L., Conklin, D.S.,Hannon, G.J., and Rosenquist, T.A. (2003)Germline transmission of RNAi in mice.Nat. Struct. Biol., 10, 91–92.
13 Rubinson, D.A., Dillon, C.P., Kwiatkowski,A.V., Sievers, C., Yang, L., Kopinja, J.,Rooney, D.L., Ihrig, M.M., McManus, M.T.,and Gertler, F.B. (2003) A lentivirus-basedsystem to functionally silence genes inprimary mammalian cells, stem cellsand transgenic mice by RNA interference.Nat. Genet., 33, 401–406.
14 Seibler, J., Kuter-Luks, B., Kern, H.,Streu, S., Plum, L., Mauer, J., Kuhn, R.,Bruning, J.C., and Schwenk, F. (2005)Single copy shRNA configuration forubiquitous gene knockdown in mice.Nucleic Acids Res., 33, e67.
15 Seibler, J., Kleinridders, A., K€uter-Luks, B.,Niehaves, S., Br€uning, J.C., and Schwenk,F. (2007) Reversible gene knockdown inmice using a tight, inducible shRNAexpression system. Nucleic Acids Res., 35,e54.
16 Nagy, A., Gocza, E., Diaz, E.M., Prideaux,V.R., Ivanyi, E., Markkula, M., and Rossant,J. (1990) Embryonic stem cells alone areable to support fetal development in themouse. Development, 110, 815–821.
17 Eggan, K., Akutsu, H., Loring, J., Jackson-Grusby, L., Klemm, M., Rideout, W.M.3rd,Yanagimachi, R., and Jaenisch, R. (2001)Hybrid vigor, fetal overgrowth, and viabilityof mice derived by nuclear cloning andtetraploid embryo complementation. Proc.Natl. Acad. Sci. USA, 98, 6209–6214.
18 Schlake, T. and J. Bode (1994) Use ofmutated FLP recognition target (FRT) sitesfor the exchange of expression cassettes atdefined chromosomal loci. Biochemistry,33 (43), 12746–12751.
19 Scheer, N., Ross, J., Rode, A., Zevnik, B.,Niehaves, S., Faust, N., and Wolf, C.R.(2008) A novel panel of mouse models toevaluate the role of human pregnane Xreceptor and constitutive androstanereceptor in drug response. J. Clin. Invest.,118 (9), 3228–3239.
20 Herold, M.J., van den Brandt, J., Seibler, J.,and Reichardt, H.M. (2008) Inducible andreversible gene silencing by stableintegration of an shRNA-encodinglentivirus in transgenic rats. Proc. Natl.Acad. Sci. USA, 105 (47), 18507–18512.
21 Kotnik, K., Popova, E., Todiras, M., Mori,M.A., Alenina, N., Seibler, J., and Bader, M.(2009) Inducible transgenic rat model fordiabetes mellitus based on shRNA-mediated gene knockdown. PLoS One,4 (4), e5124.
References j91





























