Embed Size (px)
Citation preview

APPLIED MICROBIOLOGY, Nov., 1965Copyright © 1965 American Society for Microbiology
Vol. 13, No. 6Printed in U.S.A.
Microbiological Assay for Organic Compoundsin Seawater
I. Quantitative Assay Procedures and Biotin DistributionCAROL D. LITCHFIELD' AND DONALD W. HOOD2
Department of Oceanography and Meteorology, Texas A & M University, College Station, Texas
Received for publication 28 May 1965
ABSTRACTLITCHFIELD, CAROL D. (Texas A & M University, College Station), AND DONALD W.
HOOD. Microbiological assay for organic compounds in seawater. I. Quantitative assayprocedures and biotin distribution. Appl. Microbiol. 13:886-894. 1965.-A method forthe quantitative identification of organic compounds in seawater has been developed.When auxotrophic mutants of Serratia marinorubra were incubated at 21 to 24 C for 72hr with constant agitation, standard bioassay reference curves were obtained. Sodiumglycerophosphate (400 mg per liter), ammonium dibasic citrate (5 g per liter), andglycerol (25 ml per liter) supplied the needed nutrients for maximal growth with alimited concentration of the required metabolite. Data are presented for the micro-biological assay for biotin in waters of the Gulf of Mexico and adjacent bays. The rangeof sensitivity for the biotin mutant A1lYV is 5 to 12 m,ug per liter in seawater, with a
growth response from 2 to 16 m,ug per liter of seawater. The possible ecological andchemical significance of biotin occurrence in spring-summer off-shore water is dis-cussed.
For over 60 years oceanographers have beenaware of the presence of organic compounds inseawater. Except for occasional interest (Putter,1909; Waksman and Carey, 1935; Saunders, 1957;Johnston, 1955; Stephens, personal communica-tion), the utility of organic compounds to themarine biosphere has not been studied. Un-doubtedly, the primary cause for this has been alack of knowledge about the composition of thisorganic fraction. Chemical studies have beenhampered by the small amount of organic matterpresent compared with the other fractions ofseawater, and, thus, analyses have been limitedprimarily to total organic carbon determinationsor to major organic fractions such as lipids andamino acids (Hood, 1963).The technique of microbiological assay seemed
particularly well suited to the marine environ-ment if assay organisms could be found withspecific organic requirements, a high degree ofsensitivity to the compound, and a fair toleranceto salinity changes. Provasoli and Pintner (1953)found that by using the alga Euglena gracilisvar. bacillaris they could assay for vitamin B12
I Present address: Department of Biochemistryand Nutrition, Texas A & M University, CollegeStation.
2 Present address: Institute of Marine Science,University of Alaska, College.
in seawater. Since that time much additionalwork on the occurrence of vitamin B12 in seawaterhas been reported. Summaries of these studieshave been published by Provasoli (1960, 1963)and Belser (1963). Most of the cited work, how-ever, has involved extraction (Cowey, 1956),desalting (Provasoli and Pintner, 1953), dilution(Daisley, 1958), and algal assays (Provasoli1963), which required long incubation periods.Comparatively little interest has been shown
in the other vitamins that may be present inseawater. Burkholder and Burkholder (1956),Provasoli and Gold (1959), Vishniac and Riley(1959), and Vishniac (1961) have investigatedthe occurrence of thiamine, as well as B12, in themarine environment. Biotin has been sought byProvasoli and Gold (1959) by use of the algaGyrodinium cohnii, whereas Burkholder andBurkholder (1956) used a biotin-requiring marinebacterium for the assay. Antia (1963) also used amarine bacterium requiring biotin to establish astandard biotin bioassay curve in the range of 5to 10 m,g of biotin per liter of seawater. To date,though, this has not been extensively applied tomarine samples.
Belser (1958) first proposed irradiation withultraviolet light to induce mutations in the ma-rine bacterium Serratia marinorubra so that cul-tures could be obtained with specific organic re-
886
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

BIOTIN DISTRIBUTION IN SEAWATER
quirements. This organism exhibited a highdegree of tolerance to salinity variations. The re-maining criterion to be satisfied was the sensi-tivity of the mutants to specific organic com-pounds.
Belser (1959) has reported qualitative dataobtained with mutants requiring biotin, uracil,isoleucine, "purine," glycine, tryptophan, andthreonine. Carlucci and Belser (1963) found 10 ,gof biotin per liter and 100 Mg of isoleucine perliter in water off the pier at Scripps Institution ofOceanography. A summary of the work withthese biochemically deficient strains of S. marino-rubra has been published by Belser (1963).To date, no studies have been reported on the
microbiological assay for specific organic metabo-lites in waters of the Gulf of Mexico. This paperpresents the methods developed for the quanti-tative microbiological assay of organic metabo-lites by use of the marine bacterium S. marino-rubra. In addition, the data obtained during a2-year period of analysis for biotin in waters ofthe Gulf of Mexico and adjacent bays arereported.
MATERIALS AND METHODSChemicals. Synthetic seawater (30 to 32%) was
prepared from Rila Marine Mix (Utility ChemicalCorp., Paterson, N.J.) supplemented with 30 mgof ethylenediaminetetraacetic acid per liter of sea-water.The best grades of organic chemicals available
from Calbiochem and Nutritional BiochemicalsCorp., Cleveland, Ohio, were used. Where stereo-isomers were available, only the natural form wasused. All other chemicals were reagent grade. Theagar, Proteose Peptone, Casitone, and yeast ex-tract were purchased from Difco Laboratories.Cystine Trypticase Agar (BBL) was used for stockculture maintenance.
Stock solutions of those organic compoundsthat were soluble in seawater were prepared inNorit-cleaned synthetic seawater and frozen untilused. We have found the average life of thesefrozen solutions to be 6 to 9 months before decom-position begins. No preservative was added.Pyrex tubes (13 by 100 mm) capped with Bacti-
Cappalls (Scientific Products, Inc., Detroit,Mich.) were used in all bioassay analyses. Allglassware and polypropylene items were firstwashed in 7X Solution (Linbro Chemical Corp.,New Haven, Conn.), rinsed, and drained dry. Theywere then soaked for 1 to 2 hr in concentratedHNO3, rinsed eight times in distilled water, andsoaked for 30 min in each of three rinses in doubleglass-distilled water. Metal-containing apparatus,Swinney filters, syringes, etc., were cleaned in asimilar manner, except that a dilute acid wash wasused without the soaking period.
Sterilization. The appropriate method for steri-lization, either dry heat or steam, was used for allequipment.
All samples and large volumes of media, exceptfor stock culture media, were filter-sterilizedthrough an all glass Millipore filter apparatus byuse of a 0.22-,u filter. A vacuum pump was used toobtain a more rapid filtration rate. Small volumesof specific growth factors and other heat-labilesubstances were passed through a Swinney filteradapter fitted with a 0.22-,u Millipore filter.
Sterile solutions, including samples, were eitherfrozen or stored at 0 to 4 C until used.Media. Enriched synthetic seawater (SWE) was
composed of: (NH4)2HC6H507, 5.0 g; sodiumglycerophosphate, 0.25 g; glycerol, 25.00 ml; Pro-teose Peptone, 2.00 g; Casitone, 2.00 g; yeast ex-tract, 2.00 g; synthetic seawater, 975.00 ml. ThepH was adjusted to 7.0 + 0.2 with NaOH or HCl,or both. Agar, if needed, was added at a concen.tration of 1.5%. This is a slight modification of themedium proposed by Belser (1958) and permitsautoclaving without a precipitate forming.Aged seawater was prepared from near-surface
water samples collected from the Sigsbee Deeparea of the Gulf of Mexico. The samples were fil-tered into 5-gal glass carboys, and these werestored in the dark for 4 to 6 weeks to provide agednatural seawater samples for comparative testing.
Norit-cleaned synthetic seawater was preparedby cleaning 6 liters of the synthetic seawater with1% activated Norit A charcoal according to theprocedures of Antia (personal communication).After filtration to remove the charcoal fines, nopreservative was added, but the cleaned seawaterwas placed in acid-cleaned polypropylene bottlesand stored at -27 C until used. Maximal storagetime was 3 months.
Fortified seawater base (FSWB) was composedof: (NH4)2HC6H507, 5.00 g; sodium glycerophos-phate, 0.40 g; glycerol, 25.00 ml; Norit-cleanedsynthetic sea-water, 915.00 ml; pH, 7.0 d 0.1.This medium was prepared slightly concentratedfor later adjustment with the necessary growthfactors or sterile seawater. This method preventedany variation in the concentration of the essentialnutrients from one tube to another. The mediumwas always filter-sterilized.
Culture. Three of the cultures in use were ob-tained from William Belser at Scripps Institute ofOceanography. They were the biotin mutant[SIO No. 38 (TAMU No. AlOlV)], the revertedpurine-pyrimidine [SIO No. P+ (TAMU No. A120)]which was used in irradiation experiments con-ducted in our laboratory, and the uracil mutant[SIO No. 22 (TAMU No. A100P)] which was usedin assay experiments to be reported later(Litchfield and Hood, in preparation).
All mutants were carried on SWE slants andCystine Trypticase Agar stabs prepared withsynthetic seawater. Transfers were made approxi-mately every 3 months, and when growth appearedon the slants or stabs the cultures were stored at0 to 4 C. One attempt at freezing cell suspensionsat -30 to -40 C, according to the procedure out-lined by Tanguay (1959), was found to be success-
VOL. 13, 1965 887
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
888 LITCHFIELD AND HOOD
ful after only 3 months. This technique is stillunder investigation.The strain to be used in an experiment was pre-
grown for 18 to 24 hr in SWE broth. The culturewas centrifuged for 7 min, the supernatant fluidwas discarded, and the cells were washed threetimes in 10-ml samples of sterile Norit-cleanedsynthetic seawater. After the final wash, the cellswere suspended to a concentration of 80 to 85% Tas read on the Beckman model DU spectrophotom-eter. This turbidity corresponded to approxi-mately 6 X 107 to 108 cells per milliliter. A Preci-sion AVari-Hi-Speed Centricone centrifuge, tablemodel, at a standard speed of 3,500 to 4,500 rev/min was used for the inoculum preparations.
Spectrophotometric determinations. For standardassay curve and seawater sample analyses, growthwas determined turbidimetrically with the Beck-man model DU spectrophotometer. A slit openingof 0.035 mm was used with a wavelength setting of530 m,u. Each culture tube was shaken vigorouslyfor 5 sec by hand prior to pouring into 1-cmmatched cuvettes, tolerance + 1% T. The rever-sion controls, no specific metabolite added, wereread first against a sterile uninoculated control,and the instrument was then readjusted to 100%T with an average of the reversion-control read-ings. All test sets were then read; thus, any growthdue to carry-over of growth factor or true rever-sion was automatically subtracted from thereadings.
Incubation conditions. All experiments wereincubated in a standard BOD incubator whichpermitted fairly constant humidity and tempera-ture control below room temperature.Early work showed a temperature range of 4 to
38 C, with an optimum of 20 to 24 C. Less pro-diginosin was excreted into the culture medium bythe cells at temperatures below 30 C. Conse-quently, we arbitrarily selected 24 C i 2 C forthe incubation temperature.Two rotary table model shakers describing a
circular motion of 1.9-cm diameter in the hori-zontal plane were installed in the incubator. Thespeed on both shakers was maintained at 130 to135 rev/min.
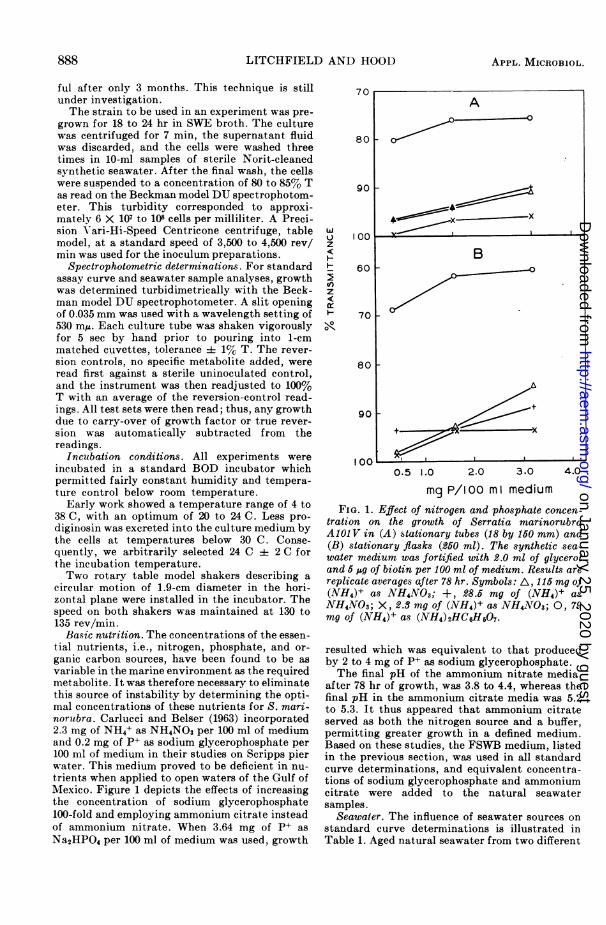
Basic nutrition. The concentrations of the essen-tial nutrients, i.e., nitrogen, phosphate, and or-ganic carbon sources, have been found to be asvariable in the marine environment as the requiredmetabolite. It was therefore necessary to eliminatethis source of instability by determining the opti-mal concentrations of these nutrients for S. mari-norubra. Carlucci and Belser (1963) incorporated2.3 mg of NH4+ as NH4NO3 per 100 ml of mediumand 0.2 mg of P+ as sodium glycerophosphate per100 ml of medium in their studies on Scripps pierwater. This medium proved to be deficient in nu-trients when applied to open waters of the Gulf ofMexico. Figure 1 depicts the effects of increasingthe concentration of sodium glycerophosphate100-fold and employing ammonium citrate insteadof ammonium nitrate. When 3.64 mg of P+ asNa2HPO4 per 100 ml of medium was used, growth
70
z
Hz
HO-OO-0
APPL. MICROBIOL.
0.5 1.0 2.0 3.0 4.0
mg P/loo m I mediumFIG. 1. Effect of nitrogen and phosphate concen-
tration on the growth of Serratia marinorubraA101V in (A) stationary tubes (18 by 160 mm) and(B) stationary flasks (250 ml). The synthetic sea-water medium was fortified with 2.0 ml of glyceroland 5 ,u of biotin per 100 ml of medium. Results arereplicate averages after 78 hr. Symbols: A, 115 mg of(NH4)+ as NH4NO3; +, 28.5 mg of (NH4)+ asNH4NO3; X, 2.8 mg of (NH4)+ as NH4NO3; 0, 79mg of (NH4)+ as (NH4) 2HC6H507.
resulted which was equivalent to that producedby 2 to 4 mg of P+ as sodium glycerophosphate.The final pH of the ammonium nitrate media,
after 78 hr of growth, was 3.8 to 4.4, whereas thefinal pH in the ammonium citrate media was 5.2to 5.3. It thus appeared that ammonium citrateserved as both the nitrogen source and a buffer,permitting greater growth in a defined medium.Based on these studies, the FSWB medium, listedin the previous section, was used in all standardcurve determinations, and equivalent concentra-tions of sodium glycerophosphate and ammoniumcitrate were added to the natural seawatersamples.
Seawater. The influence of seawater sources onstandard curve determinations is illustrated inTable 1. Aged natural seawater from two different
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
BIOTIN DISTRIBUTION IN SEAWATER
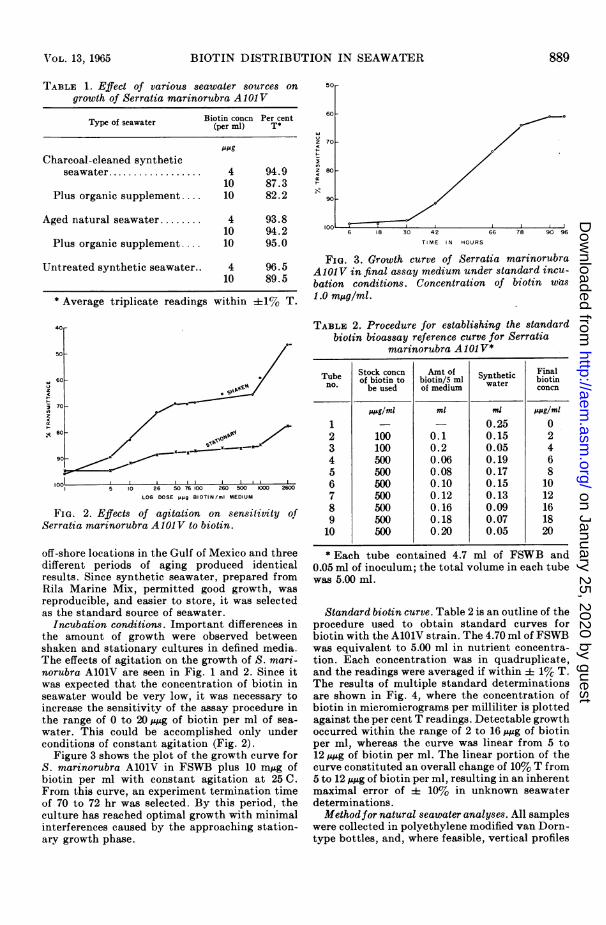
TABLE 1. Effect of various seawater sources ongrowth of Serratia marinorubra A101V
Type of seawater Biotin concn Per centTypeofeawater(per ml) T*
MSgCharcoal-cleaned synthetic
seawater .................. 4 94.910 87.3
Plus organic supplement 10 82.2
Aged natural seawater........ 4 93.810 94.2
Plus organic supplement .... 10 95.0
Untreated synthetic seawater.. 4 96.510 89.5
* Average triplicate readings within ±1% T.
40
W 60
Z #
> 70_
4z
90
1001 II I I1015 10 26 50 76 100 260 500 1000 2600
LOG DOSE Ig 0BIOTIN/m MEDIUM
FIG. 2. Effects of agitation on sensitivity ofSerratia marinorubra AlOJ V to biotin.
off-shore locations in the Gulf of Mexico and threedifferent periods of aging produced identicalresults. Since synthetic seawater, prepared fromRila Marine Mix, permitted good growth, wasreproducible, and easier to store, it was selectedas the standard source of seawater.
Incubation conditions. Important differences inthe amount of growth were observed betweenshaken and stationary cultures in defined media.The effects of agitation on the growth of S. mari-norubra AlOlV are seen in Fig. 1 and 2. Since itwas expected that the concentration of biotin inseawater would be very low, it was necessary toincrease the sensitivity of the assay procedure inthe range of 0 to 20,u,g of biotin per ml of sea-water. This could be accomplished only underconditions of constant agitation (Fig. 2).
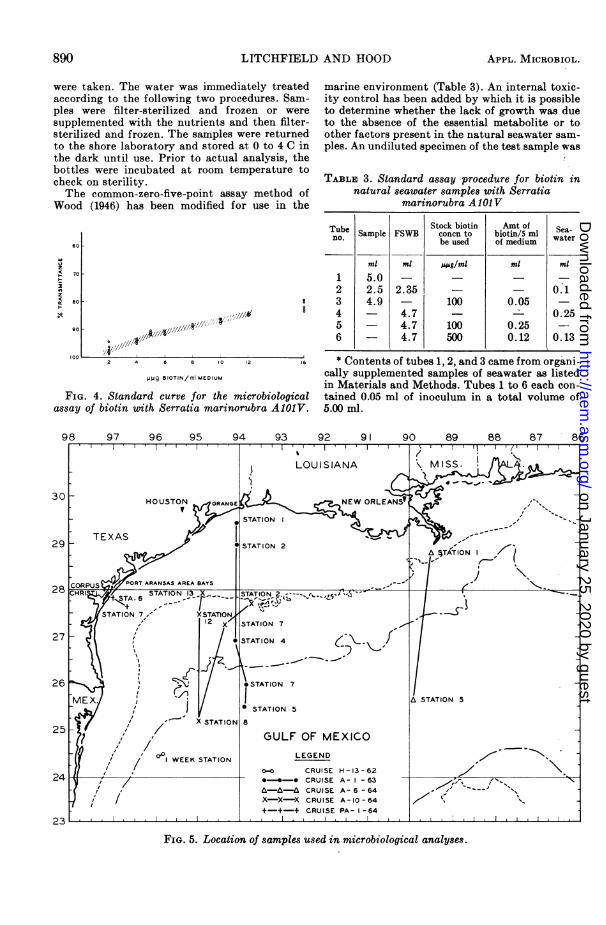
Figure 3 shows the plot of the growth curve forS. marinorubra A1O1V in FSWB plus 10 m;g ofbiotin per ml with constant agitation at 25 C.From this curve, an experiment termination timeof 70 to 72 hr was selected. By this period, theculture has reached optimal growth with minimalinterferences caused by the approaching station-ary growth phase.
60
z 70
82
6 18 30 42 66 78 90 96
TIME IN HOURS
FIG. 3. Growth curve of Serratia marinorubraA1OJV in final assay medium under standard incu-bation conditions. Concentration of biotin was1.0 m,g/ml.
TABLE 2. Procedure for establishing the standardbiotin bioassay reference curve for Serratia
marinorubra AJl0V*
Tube Stock concn Amt of Synthetic Finaln.
of biotin to biotin/5 ml Syntheti biotinno. be used of medium water concn
ppg/ml ml ml ivWg/ml1 _ 0.25 02 100 0.1 0.15 23 100 0.2 0.05 44 500 0.06 0.19 65 500 0.08 0.17 86 500 0.10 0.15 107 500 0.12 0.13 128 500 0.16 0.09 169 500 0.18 0.07 1810 500 0.20 0.05 20
* Each tube contained 4.7 ml of FSWB and0.05 ml of inoculum; the total volume in each tubewas 5.00 ml.
Standard biotin curve. Table 2 is an outline of theprocedure used to obtain standard curves forbiotin with the AlOlV strain. The 4.70 ml of FSWVBwas equivalent to 5.00 ml in nutrient concentra-tion. Each concentration was in quadruplicate,and the readings were averaged if within i 1%G T.The results of multiple standard determinationsare shown in Fig. 4, where the concentration ofbiotin in micromicrograms per milliliter is plottedagainst the per cent T readings. Detectable growthoccurred within the range of 2 to 16 sl,g of biotinper ml, whereas the curve was linear from 5 to12 j.sg of biotin per ml. The linear portion of thecurve constituted an overall change of 10% T from5 to 12 .uMg of biotin per ml, resulting in an inherentmaximal error of ± 10% in unknown seawaterdeterminations.Methodfor natural seawater analyses. All samples
were collected in polyethylene modified van Dorn-type bottles, and, where feasible, vertical profiles
889l'OL. 13, 1965
5or-
,---"O-o
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
LITCHFIELD
were taken. The water was immediately treatedaccording to the following two procedures. Sam-ples were filter-sterilized and frozen or weresupplemented with the nutrients and then filter-sterilized and frozen. The samples were returnedto the shore laboratory and stored at 0 to 4 C inthe dark until use. Prior to actual analysis, thebottles were incubated at room temperature tocheck on sterility.The common-zero-five-point assay method of
Wood (1946) has been modified for use in the
AND HOOD APPL. MICROBIOL.
marine environment (Table 3). An internal toxic-ity control has been added by which it is possibleto determine whether the lack of growth was dueto the absence of the essential metabolite or toother factors present in the natural seawater sam-ples. An undiluted specimen of the test sample was
TABLE 3. Standard assay procedure for biotin innatural seawater samples with Serratia
marinorubra Al01V
ppg BIOTIN/ml MEDIUM
FIG. 4. Standard curve for the microbiologicalassay of biotin with Serratia marinorubra AlOIV.
Tube Stock biotin Amt of Seano. Sample FSWB concn to biotin/5 ml water
beused of medium wae
ml ml jWglmi ml ml
1 5.0 _2 2.5 2.35 0.13 4.9 100 0.054 4.7 0.255 4.7 100 0.256 - 4.7 500 0.12 0.13
* Contents of tubes 1, 2, and 3 came from organi-cally supplemented samples of seawater as listedin Materials and Methods. Tubes 1 to 6 each con-tained 0.05 ml of inoculum in a total volume of5.00 ml.
FIG. 5. Location of samples used in microbiological analyses.
890
60
z
z
Fx
70
so 88
"//,///*
/ " " 4, ",
'AllllflllllI/i/I
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
BIOTIN DISTRIBUTION IN SEAWATER
fortified with a known minimal concentration ofbiotin. Less growth than that expected from thebiotin addition indicated toxicity, whereas moregrowth indicated possible concentration of biotinbelow the minimal level readable from the stand-ard curve.
RESULTS
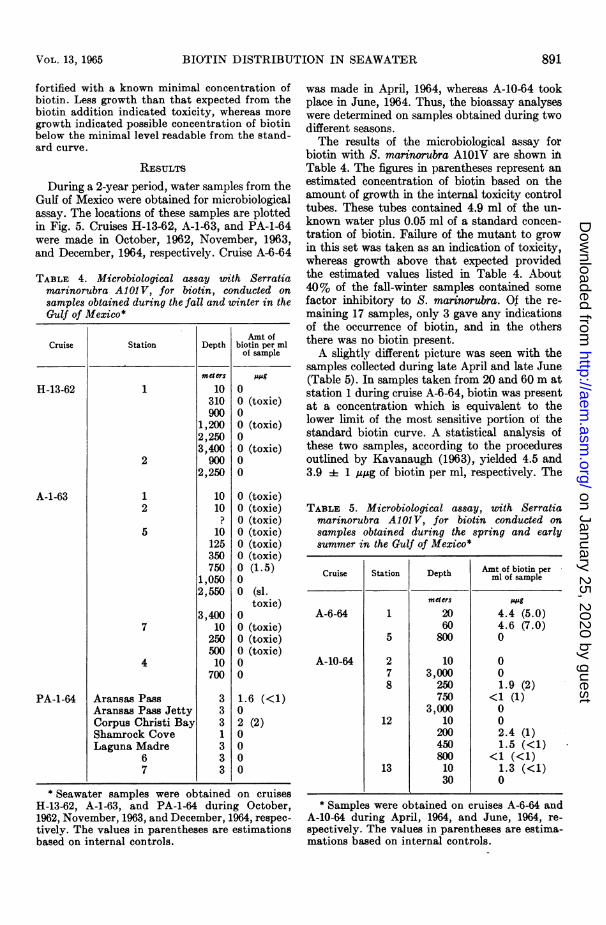
During a 2-year period, water samples from theGulf of Mexico were obtained for microbiologicalassay. The locations of these samples are plottedin Fig. 5. Cruises H-13-62, A-1-63, and PA-1-64were made in October, 1962, November, 1963,and December, 1964, respectively. Cruise A-6-64
TABLE 4. Microbiological assay with Serratiamarinorubra A10 V, for biotin, conducted onsamples obtained during the fall and winter in theGulf of Mexico*
Amt ofCruise Station Depth biotin per ml
of sample
H-13-62
A-1-63
PA-1-64
1
2
12
5
7
4
Aransas PassAransas Pass JettyCorpus Christi BayShamrock CoveLaguna Madre
67
meters10
310900
1,2002,2503,400900
2,250
1010
10125350750
1,0502,550
3,40010
25050010
700
3331333
00 (toxic)00 (toxic)00 (toxic)00
0 (toxic)0 (toxic)0 (toxic)0 (toxic)0 (toxic)0 (toxic)0 (1.5)00 (81.
toxic)00 (toxic)0 (toxic)0 (toxic)00
1.6 (<1)02 (2)0000
was made in April, 1964, whereas A-10-64 tookplace in June, 1964. Thus, the bioassay analyseswere determined on samples obtained during twodifferent seasons.The results of the microbiological assay for
biotin with S. marinorubra AlOlV are shown inTable 4. The figures in parentheses represent anestimated concentration of biotin based on theamount of growth in the internal toxicity controltubes. These tubes contained 4.9 ml of the un-known water plus 0.05 ml of a standard concen-tration of biotin. Failure of the mutant to growin this set was taken as an indication of toxicity,whereas growth above that expected providedthe estimated values listed in Table 4. About40% of the fall-winter samples contained somefactor inhibitory to S. marinorubra. Of the re-maining 17 samples, only 3 gave any indicationsof the occurrence of biotin, and in the othersthere was no biotin present.A slightly different picture was seen with the
samples collected during late April and late June(Table 5). In samples taken from 20 and 60 m atstation 1 during cruise A-6-64, biotin was presentat a concentration which is equivalent to thelower limit of the most sensitive portion of thestandard biotin curve. A statistical analysis ofthese two samples, according to the proceduresoutlined by Kavanaugh (1963), yielded 4.5 and3.9 -+ 1 ,ug of biotin per ml, respectively. The
TABLE 5. Microbiological assay, with Serratiamarinorubra AIO V, for biotin conducted onsamples obtained during the spring and earlysummer in the Gulf of Mexico*
Cruise Station Depth Amt of biotin perml of sample
meters SWA-6-64 1 20 4.4 (5.0)
60 4.6 (7.0)5 800 0
A-10-64 2 10 07 3,000 08 250 1.9 (2)
750 <1 (1)3,000 0
12 10 0200 2.4 (1)450 1.5 (<1)800 <1 (<1)
13 10 1.3 (<1)30 0
* Samples were obtained on cruises A-6-64 andA-10-64 during April, 1964, and June, 1964, re-spectively. The values in parentheses are estima-mations based on internal controls.
* Seawater samples were obtained on cruises11-13-62, A-1-63, and PA-1-64 during October,1962, November, 1963, and December, 1964, respec-tively. The values in parentheses are estimationsbased on internal controls.
VOL. 13, 1965 891
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from
LITCHFIELD AND HOOD
values listed in Table 5 were read directly fromthe standard curve with a i 10% confidence level.
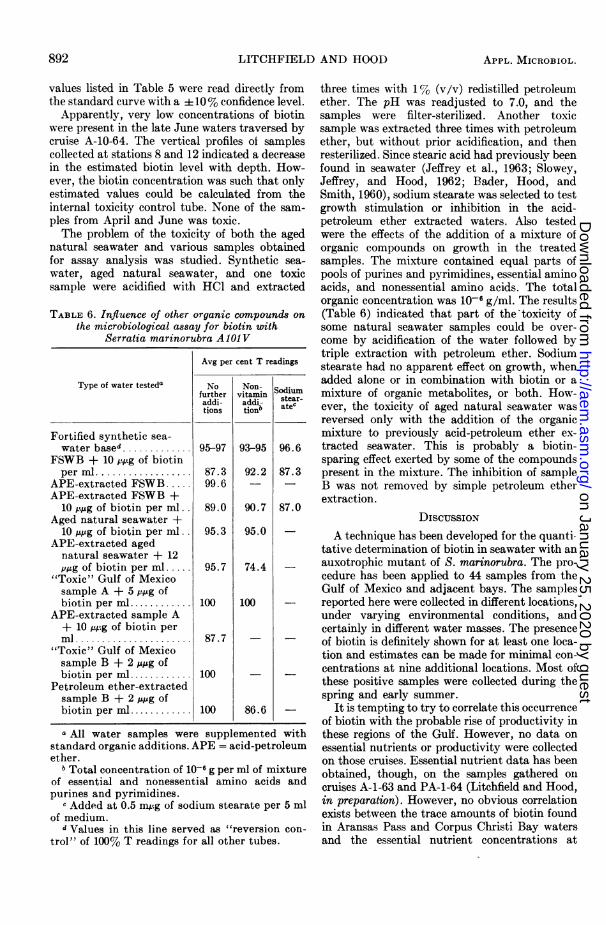
Apparently, very low concentrations of biotinwere present in the late June waters traversed bycruise A-10-64. The vertical profiles of samplescollected at stations 8 and 12 indicated a decreasein the estimated biotin level with depth. How-ever, the biotin concentration was such that onlyestimated values could be calculated from theinternal toxicity control tube. None of the sam-ples from April and June was toxic.The problem of the toxicity of both the aged
natural seawater and various samples obtainedfor assay analysis was studied. Synthetic sea-water, aged natural seawater, and one toxicsample were acidified with HCl and extracted
TABLE 6. Influence of other organic compounds onthe microbiological assay for biotin with
Serratia marinorubra A101 V
Avg per cent T readings
Type of water testeda No Non- Sodiumfurther vitamin
seraddi- addi- sertions tionb atee
Fortified synthetic sea-water base ............. 95-97 93-95 96.6
FSWB + 10 pp.g of biotinper ml .................. 87.3 92.2 87.3
APE-extracted FSWB..... 99.6APE-extracted FSWB +10,ptggof biotin per ml. 89.0 90.7 87.0
Aged natural seawater +10,u,g of biotin per ml. 95.3 95.0
APE-extracted agednatural seawater + 12p,ug of biotin per ml ..... 95.7 74.4
"Toxic" Gulf of Mexicosample A + 5,v,ug ofbiotin per ml ............ 100 100
APE-extracted sample A+ 10 gpg of biotin perml .................... 87.7 -
"Toxic" Gulf of Mexicosample B + 2 /Aug ofbiotin per ml ............ 100 -
Petroleum ether-extractedsample B + 2 Ipg ofbiotin per ml ............ 100 86.6
a All water samples were supplemented withstandard organic additions. APE = acid-petroleumether.
b Total concentration of 10-6 g per ml of mixtureof essential and nonessential amino acids andpurines and pyrimidines.
c Added at 0.5 m,.g of sodium stearate per 5 mlof medium.
d Values in this line served as "reversion con-trol" of 100% T readings for all other tubes.
three times with 1% (v/v) redistilled petroleumether. The pH was readjusted to 7.0, and thesamples were filter-sterilized. Another toxicsample was extracted three times with petroleumether, but without prior acidification, and thenresterilized. Since stearic acid had previously beenfound in seawater (Jeffrey et al., 1963; Slowey,Jeffrey, and Hood, 1962; Bader, Hood, andSmith, 1960), sodium stearate was selected to testgrowth stimulation or inhibition in the acid-petroleum ether extracted waters. Also testedwere the effects of the addition of a mixture oforganic compounds on growth in the treatedsamples. The mixture contained equal parts ofpools of purines and pyrimidines, essential aminoacids, and nonessential amino acids. The totalorganic concentration was 10-6 g/ml. The results(Table 6) indicated that part of the toxicity ofsome natural seawater samples could be over-come by acidification of the water followed bytriple extraction with petroleum ether. Sodiumstearate had no apparent effect on growth, whenadded alone or in combination with biotin or amixture of organic metabolites, or both. How-ever, the toxicity of aged natural seawater wasreversed only with the addition of the organicmixture to previously acid-petroleum ether ex-tracted seawater. This is probably a biotin-sparing effect exerted by some of the compoundspresent in the mixture. The inhibition of sampleB was not removed by simple petroleum etherextraction.
DISCUSSIONA technique has been developed for the quanti-
tative determination of biotin in seawater with anauxotrophic mutant of S. marinorubra. The pro-cedure has been applied to 44 samples from theGulf of Mexico and adjacent bays. The samplesreported here were collected in different locations,under varying environmental conditions, andcertainly in different water masses. The presenceof biotin is definitely shown for at least one loca-tion and estimates can be made for minimal con-centrations at nine additional locations. Most ofthese positive samples were collected during thespring and early summer.
It is tempting to try to correlate this occurrenceof biotin with the probable rise of productivity inthese regions of the Gulf. However, no data onessential nutrients or productivity were collectedon those cruises. Essential nutrient data has beenobtained, though, on the samples gathered oIncruises A-1-63 and PA-1-64 (Litchfield and Hood,in preparation). However, no obvious correlationexists between the trace amounts of biotin foundin Aransas Pass and Corpus Christi Bay watersand the essential nutrient concentrations at
892 APPL. MICROBIOL.
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

BIOTIN DISTRIBUTION IN SEAWATER
these two positions; neither can a correlation befound between the lack of biotin in waters sam-pled during cruise A-1-63 and the essentialnutrient data. To keep the data in perspective,one must realize that the information obtainedrepresents finite times and places and should notbe extrapolated to any annual or overall pattern.Much additional testing over prolonged periodsot time needs to be done before any productivity-metabolite relationship can be drawn.The inhibitors found in many of the winter
samples present yet another area for considera-tion. An examination of the trace metal data ob-tained by Hood and Slowey (1964) on portions ofthe bioassay samples from cruise A-1-63 revealeda p)ossible correlation between high concentra-tions of zinc (>1.5 jig per liter) and manganese(>0.5 ,ug per liter) and the toxicity of S. marino-rubra. This high concentration of trace metalscould also indicate the presence of a very differentwater mass which might contain a substance toxicto S. marinorubra. At station 5 no inhibition oc-curred at depths below 750 m where there was adefinite break in the levels of zinc and manganese.However, the fact that the toxicity may some-times be removed by acid-petroleum ether treat-ment further complicated this picture. Among thecurrent research problems in our laboratory is aninvestigation of the possible existence of lipid-trace metal complexes in natural waters.
Although no overall conclusions can be drawnas to the significance of biotin in the marine en-vironment, it is reasonable to say that a quanti-tative microbiological assay system has been de-veloped and extensively tested in the Gulf ofMexico waters. Further work should include atwo- to threefold concentration of samples. SinceS. marinorubra is highly salt tolerant, up to 10%NaCl, the resultant increase in salinity from aconcentration step should not interfere with theassay. The benefit of such an additional stepwould be to raise the biotin level to a point withinthe most sensitive portions of the standardreference curve. Thus, an even more accuratepicture of the occurrence of biotin in seawatercould be obtained.
ACKNOWLEDGMENTS
This investigation was supported by grantG 19809 from the National Science Foundation.We are indebted to W. L. Belser for supplying
us with the PI strain and no. 38 mutant, the biotinmutant, from the Scripps Institute of Oceanog-raphy Serratia marinorubra mutant collection. Weare also most appreciative for his assistance andencouragement during the initial phases of thiswork. We also wish to thank J. M. Prescott for hishelpful suggestions during the research and manu-
script preparation. Finally, we wish to acknowl-edge the excellent technical assistance of GarlandSparks throughout much of this study.
LITERATURE CITED
ANTIA, N. J. 1963. A microbiological assay forbiotin in sea water. Can. J. Microbiol. 9:403-409.
BADER, R. G., D. W. HOOD, AND J. B. SMITH. 1960.Recovery of dissolved organic matter in seawater and organic sorption by particulatematerial. Geochim. Cosmochim. Acta 19:236-243.
BELSER, W. L. 1958. Possible application of abacterial bioassay in productivity studies. Spec.Sci. Rept. Fisheries 279:55-58.
BELSER, W. L. 1959. Bioassay of organic micro-nutrients in the sea. Proc. Natl. Acad. Sci. U.S.45:1535-1542.
BELSER, W. L. 1963. Bioassay of trace substances,p. 220-231. In M. N. Hill [ed.], The sea, vol. 2.Interscience Publishers, Inc., New York.
BURKHOLDER, P. R., AND L. M. BURKHOLDER.1956. Microbiological assay of vitamin B12 inmarine solids. Science 123:1071-1073.
CARLUCCI, A. F., AND W. L. BELSER. 1963. Amethod for the bioassay of metabolites in seawater with Serratia marinorubra. Bacteriol.Proc., p. 3.
COWEY, C. B. 1956. A preliminary investigationof the variation of vitamin B12 in oceanic andcoastal waters. J. Marine Biol. Assoc. U.K.35:609-620.
DAISLEY, K. W. 1958. Measurement of vitaminB12 in sea water. J. Marine Biol. Assoc. U.K.37:673-681.
HoOD, D. W. 1963. Chemical oceanography, p. 129-155. In H. Barnes [ed.], Oceanography andmarine biology, annual review, vol. 1. GeorgeAllen and Unwin Ltd., London.
HoOD, D. W., AND J. F. SLOWEY. 1964. The chem-istry and analysis of trace metals in sea water.AEC Ann. Rept., 64-27A.
JEFFREY, L. M., B. F. PASBY, B. STEVENSON, ANDD. W. HOOD. 1963. Lipids of ocean water, p. 1-23.In U. Colombo and G. D. Hobson fed.], Ad-vances in organic geochemistry. PergamonPress, Inc., New York.
JOHNSTON, R. 1955. Biologically active compoundsin the sea. J. Marine Biol. Assoc. U.K. 34:185-195.
KAVANAGH, F. 1963. Elements of photometricassaying, p. 142-217. In F. Kavanagh [ed.], Ana-lytical microbiology. Academic Press, Inc., NewYork.
PROVASOLI, L. 1960. Growth factors in marineorganisms, p. 385-403. In A. A. Buzzati-Traverso[ed.], Proc. Symp. Perspectives in MarineBiology. Univ. California Press, Berkeley.
PROVASOLI, L. 1963. Organic regulation of phyto-plankton fertility, p. 165-219. In M. N. Hill[ed.), The sea, vol. 2. Interscience PublishersInc., New York.
893\' OL. 13, 1965
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from

894 LITCHFIELD AND HOOD
PROVASOLI, L., AND I. J. PINTNER. 1953. Assay ofvitamin B12 in sea water. Proc. Soc. Protozool.4:10.
PROVASOLI, L., AND K. GoLD. 1959. Gyrodiniumcohnii, a bioassay organism for biotin and thia-mine in sea water. Intern. Oceanogr. Congr.,New York, p. 931-933.
PVTTER, A. 1909. Die Ernahrung der Wassertiereund der Staffhaushalt der Gewasser. VEBGustav Fischer Verlag, Jena, Germany.
SAUNDERS, G. 1957. Interrelationships of dissolvedorganic matter and phytoplankton. Botan. Rev.23:389-409.
SLOWEY, J. F., L. M. JEFFREY, AND D. W. HOOD.1962. The fatty acid content of ocean water.Geochim. Cosmochim. Acta 26:607-616.
STEPHENS, G. C. 1964. Dissolved organic materialas a nutritional source for marine and estuarine
APPL. MICROBIOL.
invertebrates. Preprint, Congress on Estuaries.Jekyll Island, Ga., 3 April 1964.
TANGUAY, A. E. 1959. Preservation of microbio-logical assay organisms by direct freezing. Appl.Microbiol. 7:84-88.
VISHNIAC, H. S. 1961. A biological assay for thia-mine in sea water. Limnol. Oceanogr. 6:31-35.
VISHNIAC, H. S., AND G. RILEY. 1959. B12 andthiamine in Long Island Sound: patterns ofdistribution and ecological significance. Intern.Oceanogr. Congr., New York, p. 942-944.
WAKSMAN, S. A., AND C. L. CAREY. 1935. Decom-position of organic matter in sea water by bac-teria. II. Influence of addition of organic sub-stances upon bacterial activities. J. Bacteriol.29:545-561.
WOOD, E. D. 1946. The theory of certain analyti-cal procedures, with particular reference' tomicro-biological assays. Analyst 71:1-14.
on January 25, 2020 by guesthttp://aem
.asm.org/
Dow
nloaded from





























