Embed Size (px)
Citation preview

Seite1 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
MethodenderMolekularbiologie
NachweisvonDNA-Agarose-Gelelektrophorese
Seite2 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
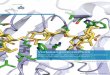
DNA-Analyse:Agarose-Gelelektrophorese
MethodezurTrennungvonDNA-MoleküleneinesGemischesanhandihrerGröße
Grundlage:
DNAdurchPhosphatrestenegaPvgeladen
DNAwandertdaherinelektrischenFeldernzurAnode(+Pol)
+ + + +
− − − −

Seite3 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNA-Analyse:Agarose-Gelelektrophorese
Agarose:PolysaccharidausDimerenvonD-Galaktoseund3,6-Anhydro-L-Galaktose
Prinzip:TrennungnachGrößedurchRetenPonineinemAgarosegel(NetzausAgarosefasern)ineinemElektrischenFeld

Seite4 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
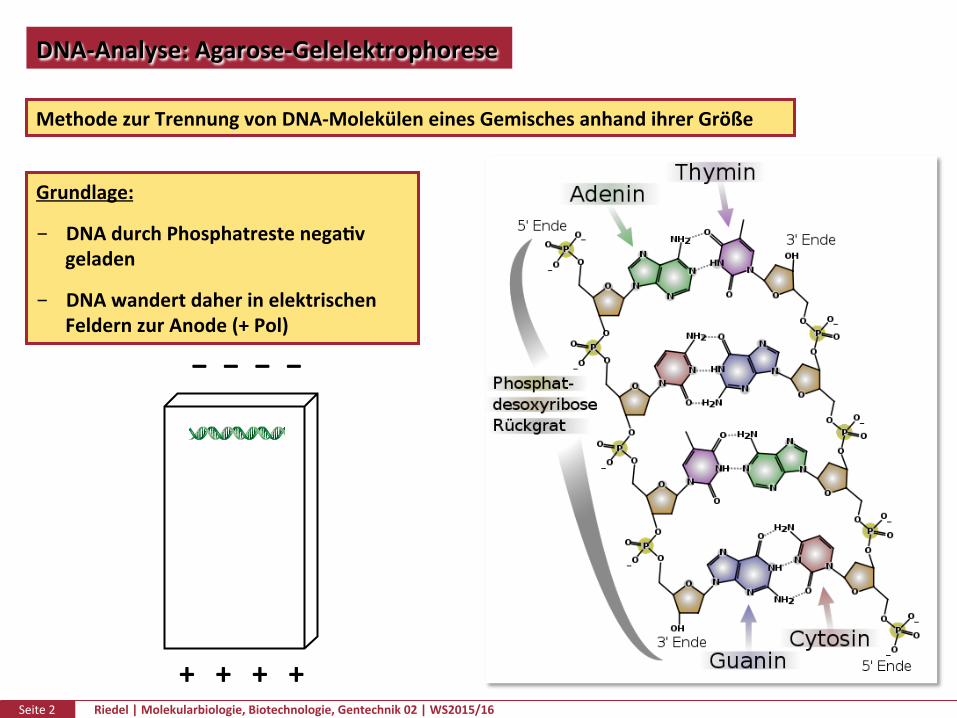
DNAQuanPtätundQualität
Prinzip:TrennungnachGrößedurchRetenPonineinemAgarosegel(NetzausAgarosefasern)ineinemElektrischenFeldDurchdasGelwerdendabeigrößereFragmentestärkerzurückgehaltenalskleine.
+ + + +
− − − −

Seite5 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNAQuanPtätundQualität
Prinzip:TrennungnachGrößedurchRetenPonineinemAgarosegel(NetzausAgarosefasern)ineinemElektrischenFeldDurchdasGelwerdendabeigrößereFragmentestärkerzurückgehaltenalskleineZugabevonFarbstoffenermöglichenesdenEndpunktderElektrophoresezubesPmmen
DünneGele=geringereDiffusion=schärferBanden
Seite6 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNAQuanPtätundQualität
Prinzip:TrennungnachGrößedurchRetenPonineinemAgarosegel(NetzausAgarosefasern)ineinemElektrischenFeldDurchdasGelwerdendabeigrößereFragmentestärkerzurückgehaltenalskleineZugabevonFarbstoffenermöglichenesdenEndpunktderElektrophoresezubesPmmenDieAgarosekonzentraPonbesPmmtdiePorengrößeunddamitdieAuflösungdesGels
%Agarose Auflösung(Bp)
0,6 20000-1000
0,7 10000-800
0,9 7000-500
1,2 6000-400
1,5 3000-200
2 2000-50
1%Agarose=150nmPoren
0,16%Agarose=500nmPoren
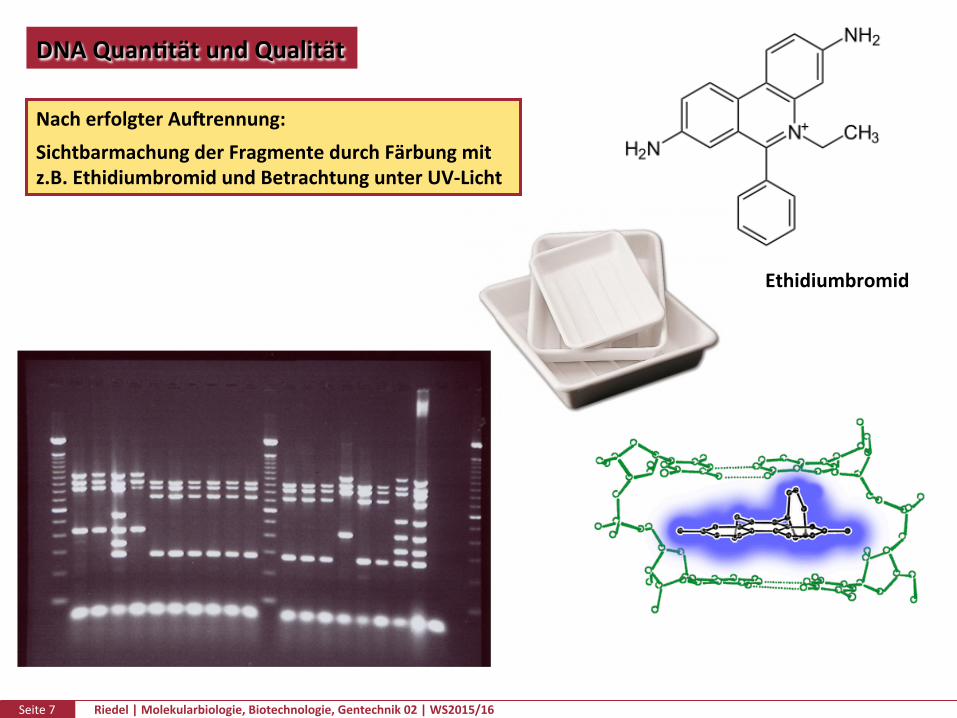
Seite7 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
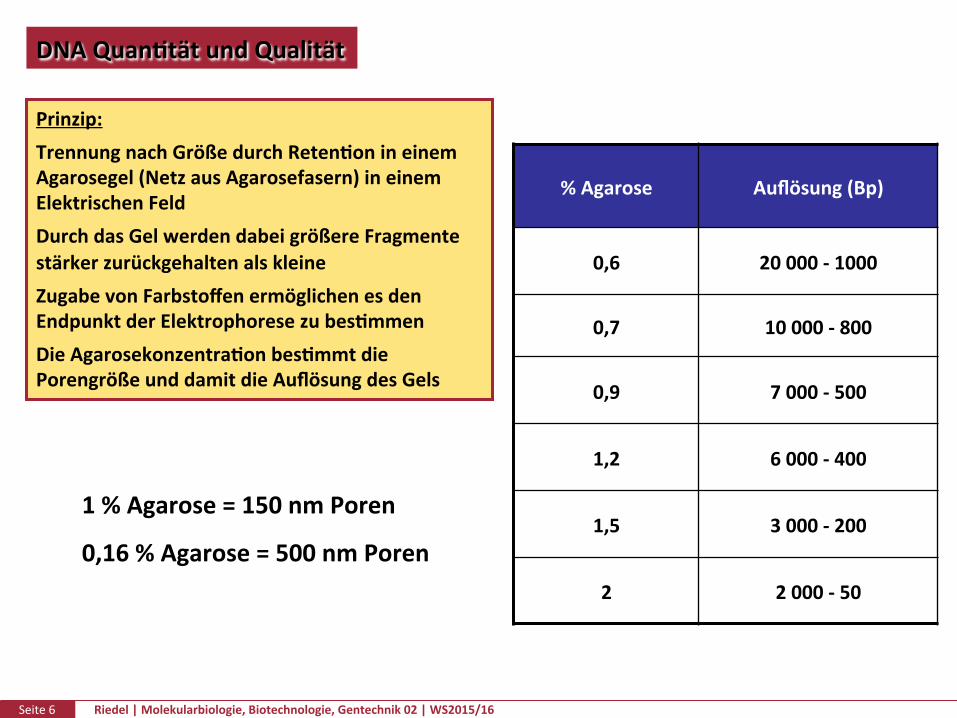
DNAQuanPtätundQualität
NacherfolgterAufrennung:SichtbarmachungderFragmentedurchFärbungmitz.B.EthidiumbromidundBetrachtungunterUV-Licht
Ethidiumbromid
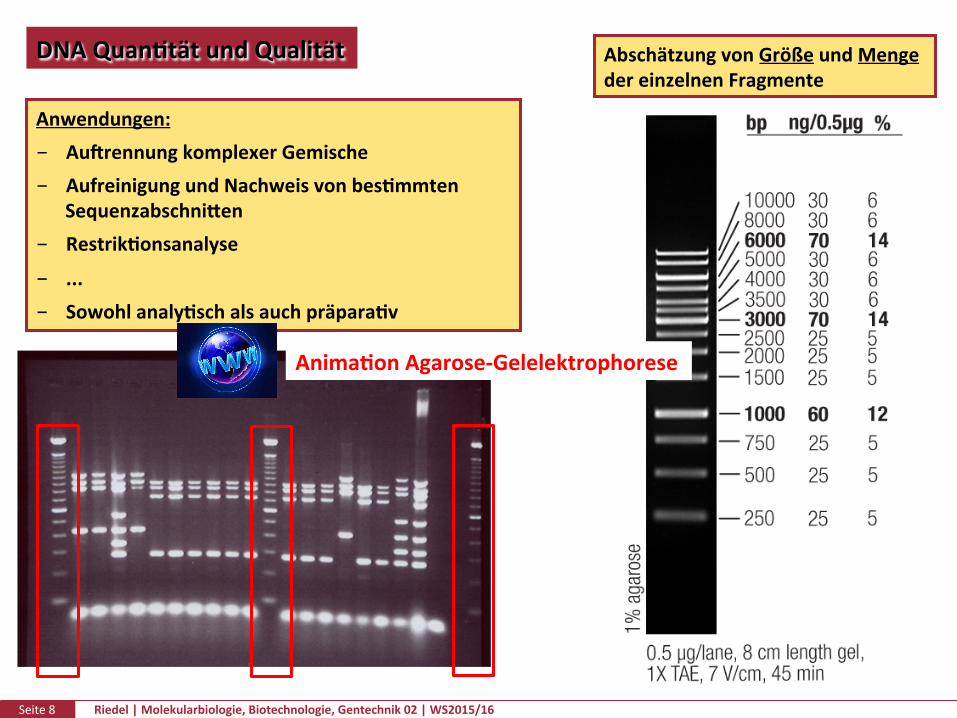
Seite8 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNAQuanPtätundQualität AbschätzungvonGrößeundMengedereinzelnenFragmenteProtocol for Loading
Step 1: Mix gently Step 2: Load 1 µl per 1 mm gel lane
Recommendations x Do not heat before loading. x Dilute your DNA sample with the 6X DNA Loading Dye
(#R0611, supplied with the ladder): mix 1 volume of the dye solution with 5 volumes of the DNA sample;
x Load the same volumes of the DNA sample and the DNA ladder;
x For quantification, adjust the concentration of the sample to equalize it approximately with the amount of DNA in the nearest band of the ladder.
x For DNA band visualization with SYBR® Green, GelRed and other intercalating dyes, do not add the dyes into the sample, use gel staining after electrophoresis or include dyes into agarose gel to avoid aberrant DNA migration.
NOTICE TO PURCHASER: LIMITED LICENSE This product or its use is covered by at least one claim of U.S. Patent Nos. 5,834,201, 6,680,378, and/or 7,132,520 owned by Invitrogen Corporation. The purchase of this product conveys to the buyer the non-transferable right to use the purchased amount of the product in internal research conducted by the buyer. The buyer cannot use this product or materials made by the employment of this product for Commercial Purposes. Commercial Purposes means any activity for consideration and may include, but is not limited to: (1) use of the product in manufacturing; (2) use of the product to provide a service, information, or data; (3) use of the product for therapeutic, diagnostic or prophylactic purposes; or (4) resale of the product, whether or not the product is resold for use in research. Further information on purchasing licenses under the above patents may be obtained by contacting the Business Development Department, Invitrogen Corporation, 5791 Van Allen Way, Carlsbad, CA 92008. Email: [email protected].
GeneRuler 1 kb DNA Ladder, ready-to-use
PRODUCT USE LIMITATION This product is developed, designed and sold exclusively for research purposes and in vitro use only. The product was not tested for use in diagnostics or for drug development, nor is it suitable for administration to humans or animals. Please refer to www.thermoscientific.com/onebio for Material Safety Data Sheet of the product.
© 2012 Thermo Fisher Scientific Inc. All rights reserved. SYBR is a registered trademark of Molecular Probes, Inc. All other trademarks are the property of Thermo Fisher Scientific Inc. and its subsidiaries.
Anwendungen: AufrennungkomplexerGemische AufreinigungundNachweisvonbesPmmten
Sequenzabschniken RestrikPonsanalyse
... SowohlanalyPschalsauchpräparaPv
AnimaPonAgarose-Gelelektrophorese

Seite9 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
MethodenderMolekularbiologie
ModifikaPonvonDNA:RestrikPonsenzyme
Seite10 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsendonukleasen
BisherigeMethodenfürArbeitenmitDNA: IsolaPon QualitätskontrolleundQuanPfikaPon VervielfälPgung(PCR) Agarose-Gelelektrophorese
WeitereArbeitsschrike,diefürdieKlonierungvonDNAwichPgsind: Schneiden
RestrikPonsenzyme=molekulareScheren,dieDNAschneiden
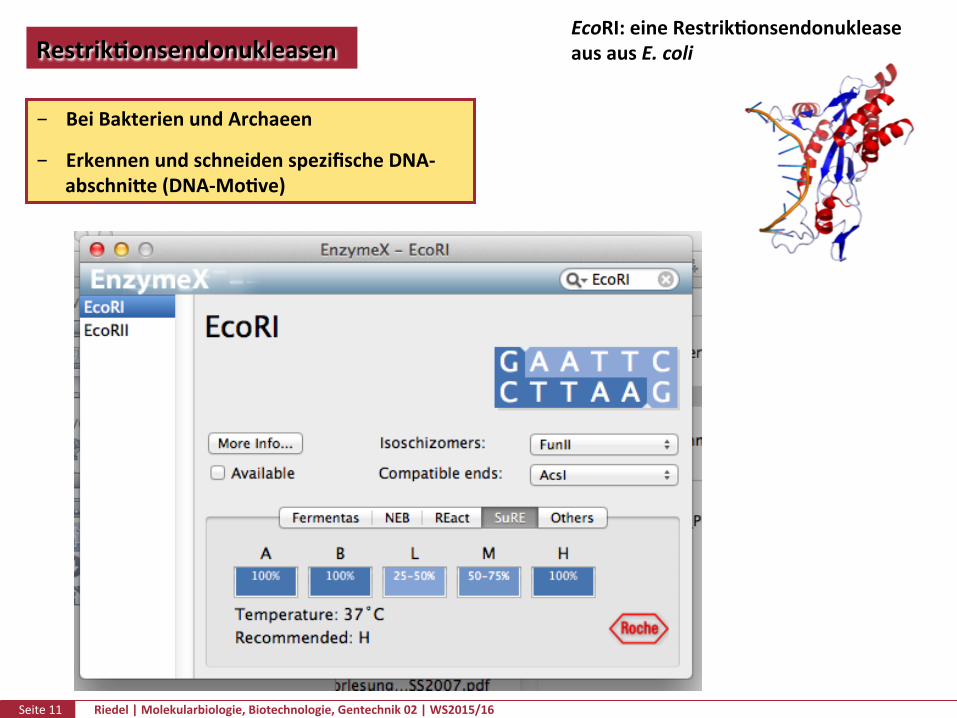
Seite11 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsendonukleasenEcoRI:eineRestrikPonsendonukleaseausausE.coli
BeiBakterienundArchaeen
ErkennenundschneidenspezifischeDNA-abschnike(DNA-MoPve)
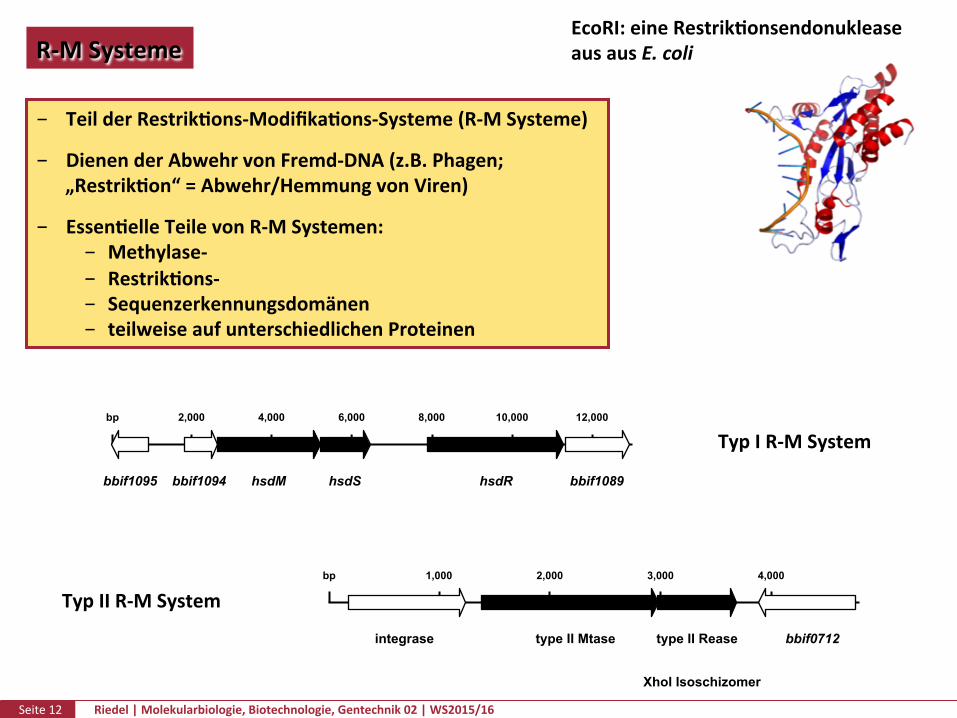
Seite12 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
R-MSysteme
TeilderRestrikPons-ModifikaPons-Systeme(R-MSysteme)
DienenderAbwehrvonFremd-DNA(z.B.Phagen;„RestrikPon“=Abwehr/HemmungvonViren)
EssenPelleTeilevonR-MSystemen: Methylase- RestrikPons- Sequenzerkennungsdomänen teilweiseaufunterschiedlichenProteinen
bbif1095 bbif1094 hsdM hsdS hsdR bbif1089
2,000 4,000 6,000 8,000 10,000 12,000 bp
integrase type II Rease bbif0712 type II Mtase
1,000 2,000 3,000 4,000 bp
TypIR-MSystem
TypIIR-MSystem
EcoRI:eineRestrikPonsendonukleaseausausE.coli
XhoI Isoschizomer

Seite13 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
R-MSysteme
Methylase(MTase)und(REase)RestrikPonsendonukleaseeinesBekteriumserkennendieselbenDNA-MoPve
MTase➜MethylierungdesMoPvs
REaseschneidetnurwenndieerkanntenDNA-MoPveNICHTmethyliertsind
BeigleichzeiPgerExpressionvonMTaseundRease:eigeneDNAgeschützt,Fremd-DNAwirdabgebaut
M 1 2 3 4 5
NachweiseinesXhoI-ähnlichenR-MSystems:
M Größenmarker(1kbLeiter)
1 PlasmidX
2 PlasmidX+XhoI
3 ChromosomaleDNA
4 ChromosomaleDNA+Xho
5 ChromosomaleDNA+EcoRI

Seite14 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
R-MSysteme
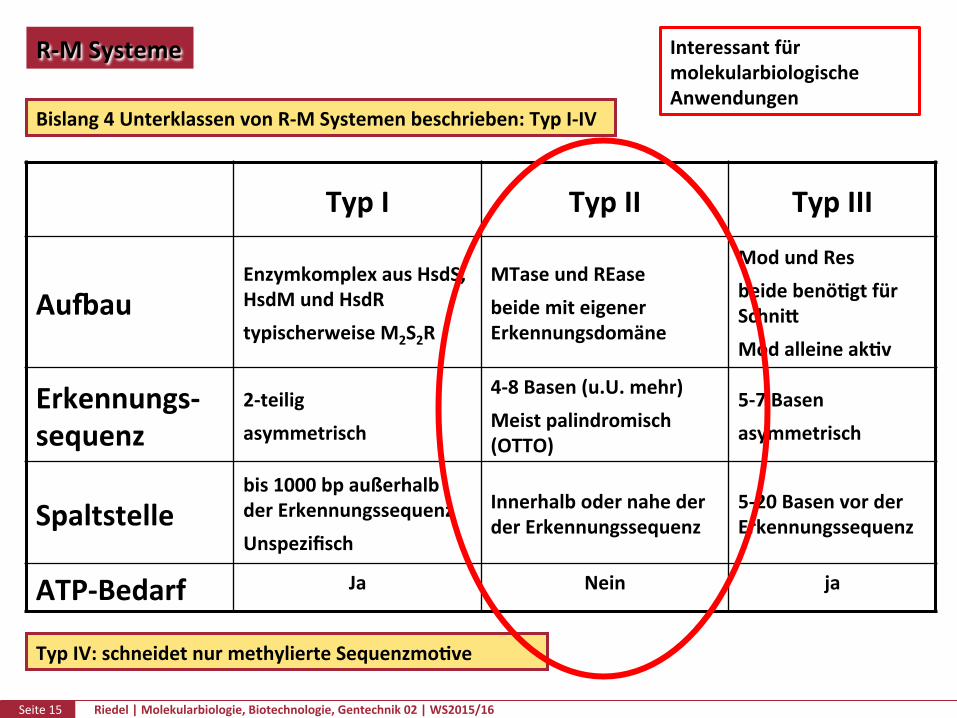
Bislang4UnterklassenvonR-MSystemenbeschrieben:TypI-IV
!"#$%& '() !"**'#&
' +,-.+/01234. 5,4 4.6247/27,+ .+89-.6: )('-.2;90241+65.416.6: ;,-7+< .+=,+3/0.16.6 1+= 2;.74<.+.6#7/;14= >? #,@.426A: *140.+. B.05,42C: D7-,2;9 B.62,4E: '6;,F !? B;1<G12H:
D;,-16 '? B7/F0.I: >3412. B727+172.: #,@.42 *? B03-.+2;10J: !.4<.9 K;? ).<2914.LM:
)1L7= D? N? )49=.+O: K.L7+ )9@L7<P: K.72; N74-1+Q: %0781L.21 !? R4,-,L1CS:
#7/;14= T? R3-U,42CC: !2.U;.+ %? V105,4=CE: !21+0.9 V122-1+CH: >,6.U; V.72-1+CI:
)1L7= W? V,4+@9CJ: '4L9=16 >1+3017276CM: '[email protected] >.026/;CO: >922. >,6.U;6.+CP:
'+210 K766CQ: D,== #? K01.+;1--.4ES: T/;78, K,@1916;7EC: V37-7+ K,+<:
).20.L V? K43X<.4EE: !1+5,4= Y1/F6EH: *1427+ R? *147+36EI: *7/;7F, *791;141EJ:
#7/;14= )? *,4<1+: (,4..+ %? *34419EM: $101F3+Z1 (1<141Z1EO: '+=48.Z W7.F14,G7/8EP:
'054.= W7+<,3=CO: %[email protected]; #10.7<;: ).674183 (? #1,EO: (,[email protected] #.7/;EQ:
$01=7-74 %? #.U7+HS: %47/ "? !.0F.4HC: W1+<[\;37 !;1GHE: )1+7.0 \? !2.7+HH:
B1449 Y? !2,==14=HI: ]1/01G !89@106F7HJ: D;,-16 '? D4132+.4HM: >1-.6 Y? $1+ %22.+HO:
>,4<. *? B? $72,4HP: R.,554.9 R? ]706,+ 1+= !;31+<[9,+< ^3
!"# $%&'(%) *+,'(-./ *"0"1'2/ 34 56768/ 9:4/ 63,'";<'(1 ="%">+;. ?1,&1(@/ !"# A,1B :>(>" C"D(1>@"%> ,EF"('>G/ 4'-(%2/ !A 6HH56IH55H/ 9:4/ H="%">+;. (%) C"0"',D@"%>/ J,'<@-+( 9%+0"1.+>2/ !"# A,1B/ !A 655KH/9:4/ KC"D(1>@"%> ,E JG"@+.>12/ L(2%" :>(>" 9%+0"1.+>2/ C">1,+>/ 3M NOH5H/ 9:4/ NC"D(1>@"%> ,E 3+;1,-+,',&2/*+,P"%>1<@/ 9%+0"1.+>(> *(."'/ JFIN58Q *(."'/ :#+>P"1'(%)/ 8?1,&1(@ +% *+,+%E,1@(>+;. (%) ?1,>",@+;.R="%,@+;./3")+;(' J,''"&" ,E SG+,/ T,'"),/ SF NKQ77I555O/ 9:4/ Q:+-$%P2@"/ QK5575 !,0,.+-+1.B/ U<..+(/ V:;G,,' ,EJG"@+.>12/ 9%+0"1.+>2 ,E $)+%-<1&G/ TG" W+%&X. *<+')+%&./ $)+%-<1&G $F7 KYY/ 9W/ OC"D(1>@"%> ,E ="%">+;./9%+0"1.+>2 ,E 4'(-(@( (> *+1@+%&G(@/ *+1@+%&G(@/ 4Z K8H7N/ 9:4/ 7*+,DG2.+;. Z(-,1(>,1+"./ :;G,,' ,E *+,',&+;(':;+"%;"./ 9%+0"1.+>2 ,E ?,1>.@,<>G/ ?,1>.@,<>G ?S6 HCT/ 9W/ 654[![ *"',P"1.B2 M%.>+><>" ,E ?G2.+;,IJG"@+;('*+,',&2/ 3,.;,# :>(>" 9%+0"1.+>2/ 66777H 3,.;,#/ U<..+(/ 66TG" 9%+0"1.+>2 ,E M''+%,+. J,''"&" ,E 3")+;+%"/91-(%(/ MZ Q6O56IKQ5H/ 9:4/ 6HC"D(1>@"%> ,E *+,;G"@+.>12/ 9%+0"1.+>2 ,E *1+.>,' 3")+;(' :;G,,'/ *1+.>,' *:O 6TC/9W/ 6KC"D(1>@"%> ,E *+,',&2/ 9%+0"1.+>2 ,E U,;G".>"1/ U,;G".>"1/ !A 6NQHVI5H66/ 9:4/ 6NF,#(1) F<&G".3")+;(' M%.>+><>"/ C<B" 9%+0"1.+>2 3")+;(' J"%>"1/ C<1G(@/ !J HVV65/ 9:4/ 68C"D(1>@"%> ,E 3,'";<'(1 *+,',&2(%) *+,>";G%,',&2/ 9%+0"1.+>2 ,E :G"E\"')/ ]+1>G J,<1>/ L".>"1% *(%B/ :G"E\"') :65 HT!/ 9W/ 6QM%.>+><>" ,E*+,>";G%,',&2/ ZTIH5HO ^+'%+<./ Z+>G<(%+(/ 6VM%.>+><> E<_1 *+,;G"@+"/ Y<.><.IZ+"-+&I9%+0"1.+>(_ >/ CIK8K7H =+".."%/="1@(%2/ 6OC"D(1>@"%> ,E C(+12 (%) ],,) :;+"%;"/ U,2(' ^">"1+%(12 (%) 4&1+;<'><1(' 9%+0"1.+>2/ CWI678O]1")"1+B.-"1& J/ C"%@(1B/ 67M%.>+><>" ,E *+,;G"@+.>12/ *UJ/ FIQV56 :P"&")/ F<%&(12/ H5C"D(1>@"%>. ,E ],,):;+"%;" (%) 3+;1,-+,',&2/ !,1>G J(1,'+%( :>(>" 9%+0"1.+>2/ U('"+&G/ !J HVQ78IVQHN/ 9:4/ H6C"D(1>@"%> ,E3,'";<'(1 *+,',&2/ M%.>+><>" ,E 3")+;(' :;+"%;"/ 9%+0"1.+>2 ,E T,B2,/ NIQI6/ :G+1,B(%")(+/ 3+%(>,IB</T,B2, 65OIOQK7/ Y(D(%/ HHM%.>+><> E<_1 ^+1,',&+"IJG(1+>"/ F<@-,')> 9%+0"1.+>(>/ CI6557O *"1'+%/ ="1@(%2/HK*1,,BG(0"% !(>+,%(' Z(-,1(>,12/ 9D>,%/ !A 667VKI8555/ 9:4/ HNC"D(1>@"%> ,E ?G(1@(;,',&2/ 9%+0"1.+>2 ,E3(..(;G<.">>. 3")+;(' :;G,,'/ L,1;".>"1/ 34 56Q88/ 9:4/ H8!(>+,%(' M%.>+><>" ,E F"('>G :;+"%;"./ 6I6OI6/W(@+2,&(/ :">(&(2(IB</ T,B2, 68OIO856/ Y(D(%/ HQM%.>+><>" ,E J"'' (%) 3,'";<'(1 *+,',&2/ 9%+0"1.+>2 ,E $)+%-<1&G/TG" W+%&X. *<+')+%&./ $)+%-<1&G $F7 KYU/ 9W/ HVC"D(1>@"%> ,E 3+;1,-+,',&2 (%) J"'' *+,',&2/ M%)+(% M%.>+><>" ,E:;+"%;"/ M!I8Q556H *(%&(',1"/ M%)+(/ HOM%.>+><>" ,E 3+;1,-+,',&2/ L(1.(# 9%+0"1.+>2/ 3+";P%+B,#( 6/ 5HI57QL(1.(#/ ?,'(%)/ H79%+0"1.+>2 ,E J('+E,1%+(/ :(%>( *(1-(1(/ :(%>( *(1-(1(/ J4 7K65QI5556/ 9:4/ K5:>(>"U"."(1;G J"%>"1 ,E ^+1,',&2 (%) *+,>";G%,',&2 `^"B>,1X/ W,'>.,0,/ !,0,.+-+1.B 1"&+,% QKK587/ U<..+(/ K6M%.>+><>",E 3,'";<'(1 *+,',&2/ 9%+0"1.+>2 ,E S1"&,%/ $<&"%"/ SU 7VN5K/ 9:4/ KHC"D(1>@"%> ,E *+,;G"@+.>12/ TG" JG+%"."9%+0"1.+>2 ,E F,%& W,%&/ F,%& W,%&/ KKC"D(1>@"%> ,E 3+;1,-+,',&2/ 9%+0"1.+>2 ,E 3(12'(%)/ J,''"&" ?(1B/3C H5VNH/ 9:4/ KN]1") F<>;G+%.,% J(%;"1 U"."(1;G J"%>"1/ :"(>>'"/ L4 7O657/ 9:4/ K83;41)'" Z(-,1(>,12/
!"# $%#& '#(()*+#,-),') *%#./- 0) 1--()**)-2 ")/3 45 678 697 ::89; <1=3 45 678 695 5>97; ?&1@/3 (#0)(A*B,)02'#&
!"#$%&# '#&() *%)%+,#-. /001. 23$4 15. !34 6 !"#$%!"!&789: 50450;1<=+,<>?>/6@
!"#$%&# '#&() *%)%+,#-. 23$4 15 !34 6 ! 8AB3,( C=&D%,)&EF G,%)) /001H +$$ ,&>-E) ,%)%,D%(
at Universitaetsbibliothek U
lm on January 24, 2013
http://nar.oxfordjournals.org/D
ownloaded from
Seite15 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
R-MSysteme
Bislang4UnterklassenvonR-MSystemenbeschrieben:TypI-IV
TypI TypII TypIII
AuuauEnzymkomplexausHsdS,HsdMundHsdRtypischerweiseM2S2R
MTaseundREasebeidemiteigenerErkennungsdomäne
ModundResbeidebenöPgtfürSchnikModalleineakPv
Erkennungs-sequenz
2-teiligasymmetrisch
4-8Basen(u.U.mehr)Meistpalindromisch(OTTO)
5-7Basenasymmetrisch
Spaltstellebis1000bpaußerhalbderErkennungssequenzUnspezifisch
InnerhalbodernahederderErkennungssequenz
5-20BasenvorderErkennungssequenz
ATP-Bedarf Ja Nein ja
TypIV:schneidetnurmethylierteSequenzmoPve
InteressantfürmolekularbiologischeAnwendungen
Seite16 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
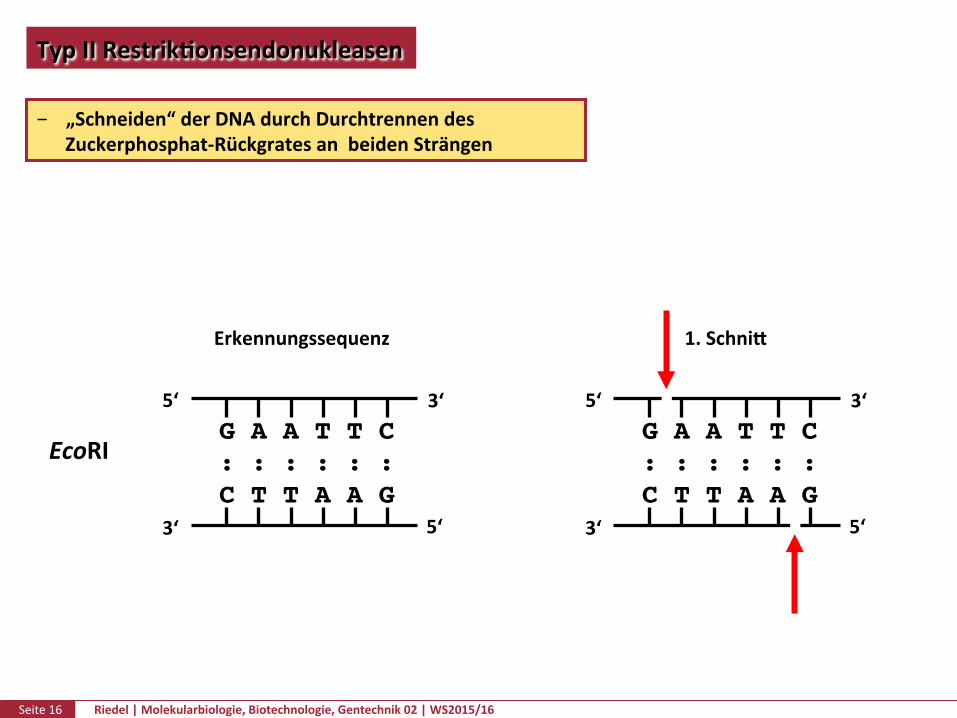
TypIIRestrikPonsendonukleasen
„Schneiden“derDNAdurchDurchtrennendesZuckerphosphat-RückgratesanbeidenSträngen
EcoRI
Erkennungssequenz 1.Schnik
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
Seite17 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
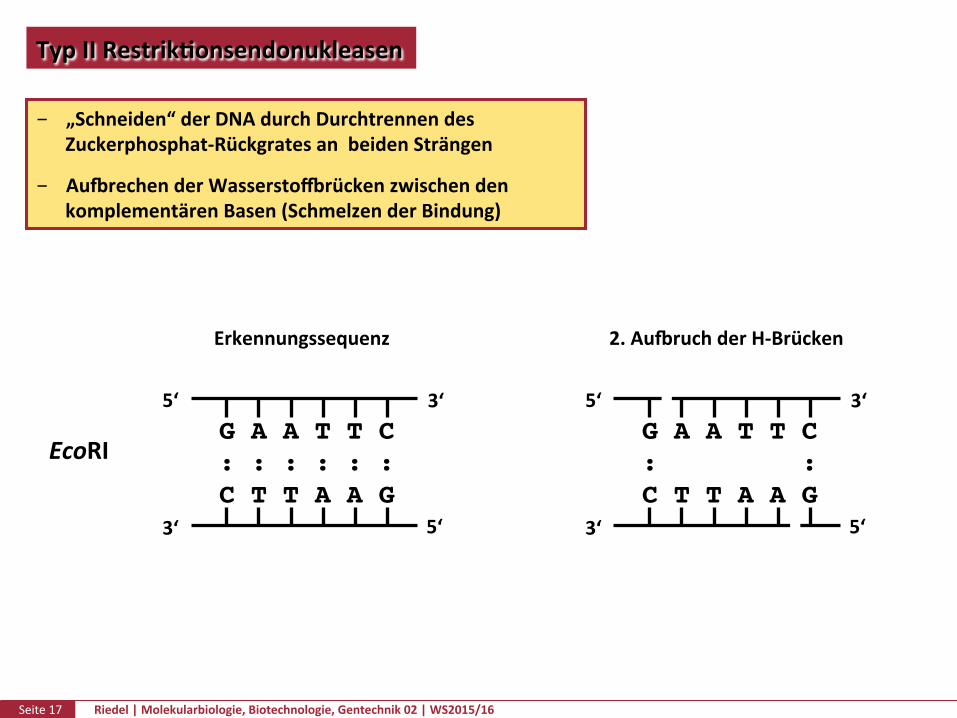
TypIIRestrikPonsendonukleasen
„Schneiden“derDNAdurchDurchtrennendesZuckerphosphat-RückgratesanbeidenSträngen
AuurechenderWasserstozrückenzwischendenkomplementärenBasen(SchmelzenderBindung)
EcoRI
Erkennungssequenz 2.AuuruchderH-Brücken
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
Seite18 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
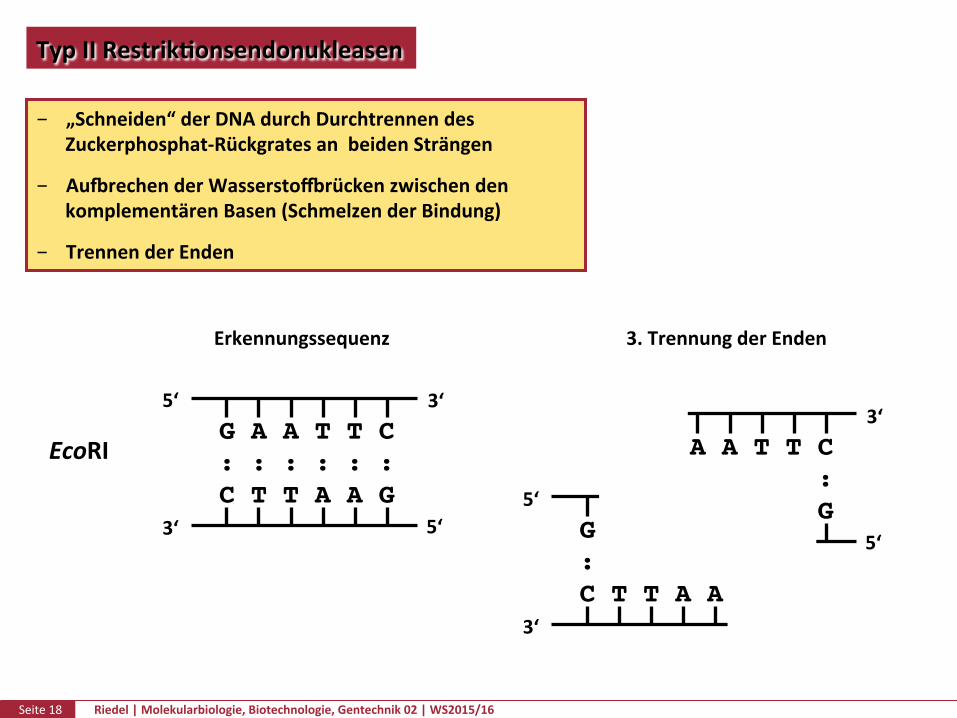
TypIIRestrikPonsendonukleasen
„Schneiden“derDNAdurchDurchtrennendesZuckerphosphat-RückgratesanbeidenSträngen
AuurechenderWasserstozrückenzwischendenkomplementärenBasen(SchmelzenderBindung)
TrennenderEnden
EcoRI
Erkennungssequenz 3.TrennungderEnden
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
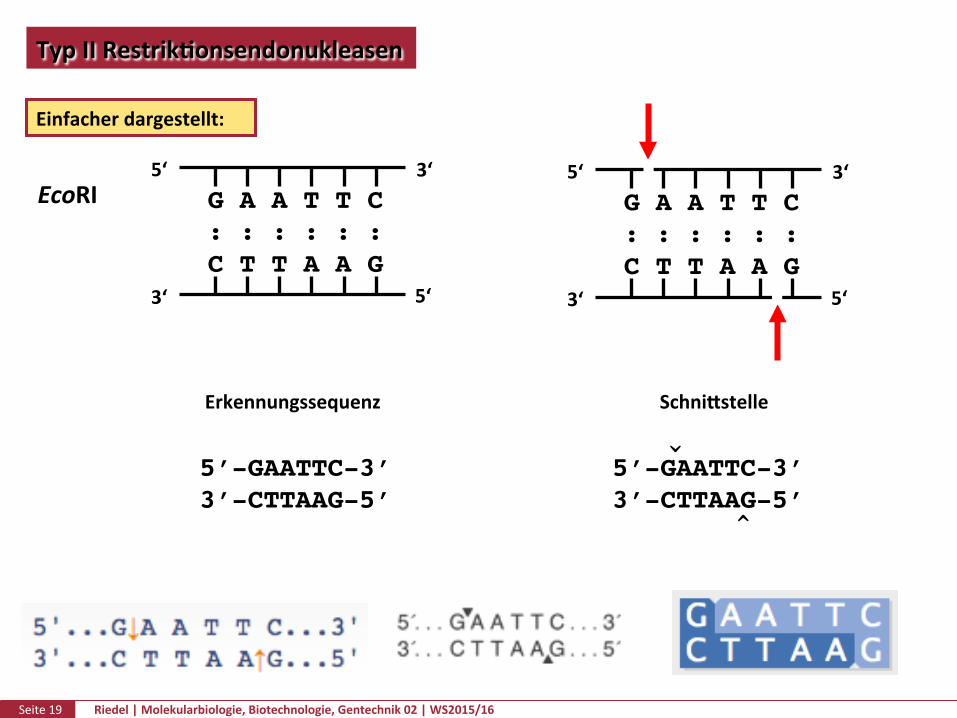
Seite19 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
TypIIRestrikPonsendonukleasen
Einfacherdargestellt:
EcoRI
Erkennungssequenz Schnikstelle
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
5’-GAATTC-3’3’-CTTAAG-5’
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
5’-GAATTC-3’3’-CTTAAG-5’
^
^
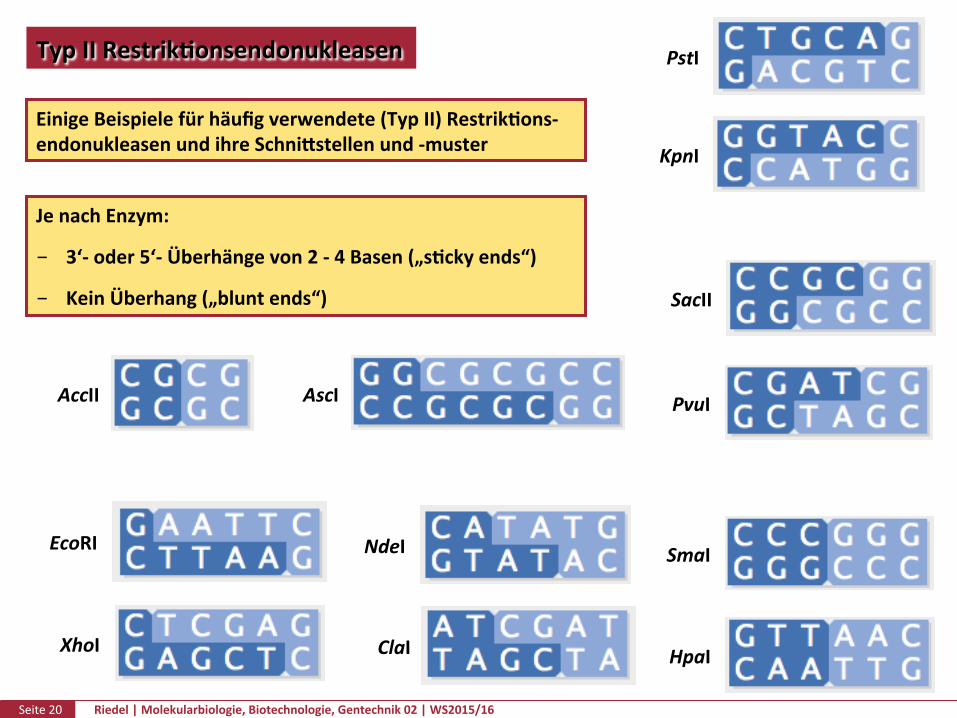
Seite20 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
TypIIRestrikPonsendonukleasen
EinigeBeispielefürhäufigverwendete(TypII)RestrikPons-endonukleasenundihreSchnikstellenund-muster
EcoRI
XhoI
PstI
NdeI
ClaI HpaI
SmaI
PvuI
KpnI
SacII
JenachEnzym:
3‘-oder5‘-Überhängevon2-4Basen(„sPckyends“)
KeinÜberhang(„bluntends“)
AccII AscI
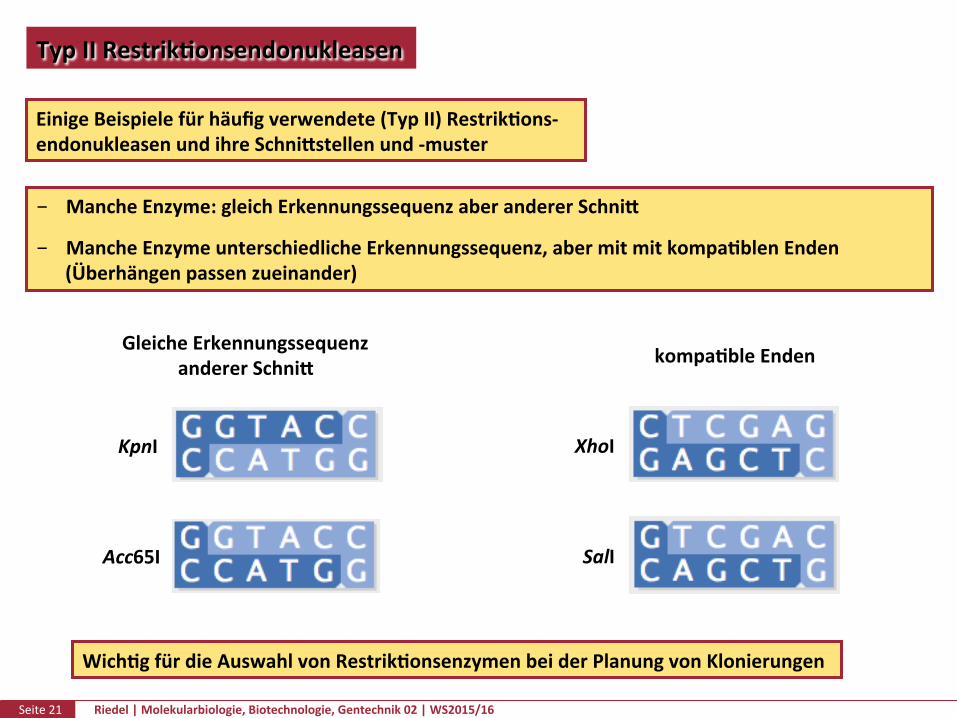
Seite21 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
TypIIRestrikPonsendonukleasen
EinigeBeispielefürhäufigverwendete(TypII)RestrikPons-endonukleasenundihreSchnikstellenund-muster
MancheEnzyme:gleichErkennungssequenzaberandererSchnik
MancheEnzymeunterschiedlicheErkennungssequenz,abermitmitkompaPblenEnden(Überhängenpassenzueinander)
XhoI
SalI
kompaPbleEndenGleicheErkennungssequenzandererSchnik
KpnI
Acc65I
WichPgfürdieAuswahlvonRestrikPonsenzymenbeiderPlanungvonKlonierungen
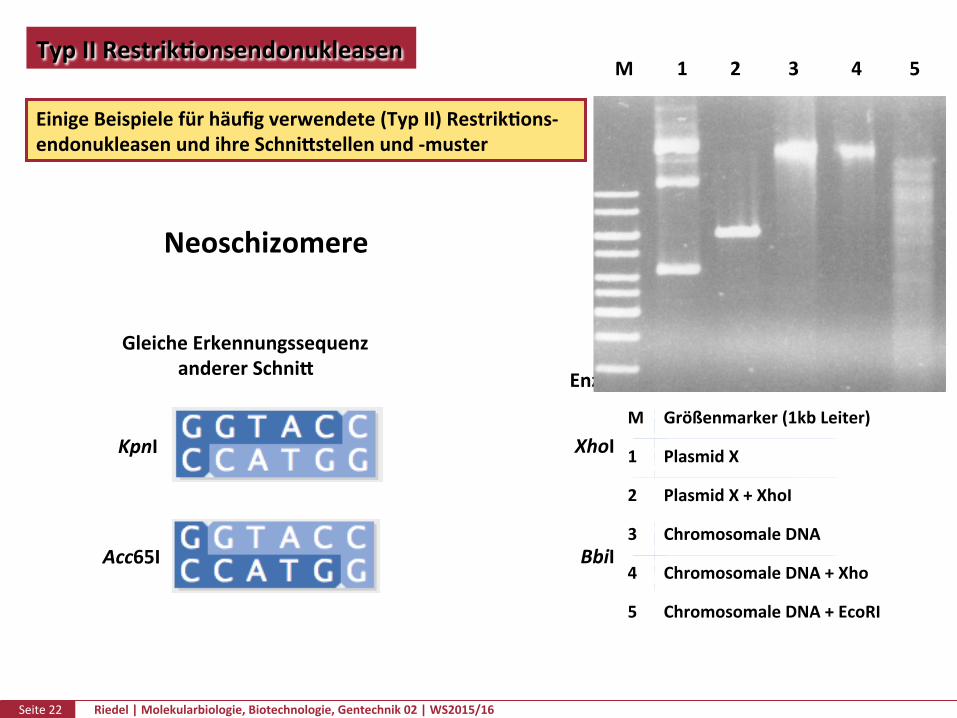
Seite22 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
TypIIRestrikPonsendonukleasen
EinigeBeispielefürhäufigverwendete(TypII)RestrikPons-endonukleasenundihreSchnikstellenund-muster
XhoI
BbiI
GleicheErkennungssequenzandererSchnik
KpnI
Acc65I
Neoschizomere Isoschizomere
GleicheErkennungssequenzgleicherSchnik
EnzymaberausanderemOrganismus
M 1 2 3 4 5
M Größenmarker(1kbLeiter)
1 PlasmidX
2 PlasmidX+XhoI
3 ChromosomaleDNA
4 ChromosomaleDNA+Xho
5 ChromosomaleDNA+EcoRI
Seite23 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsverdauinderMolekularbiologie
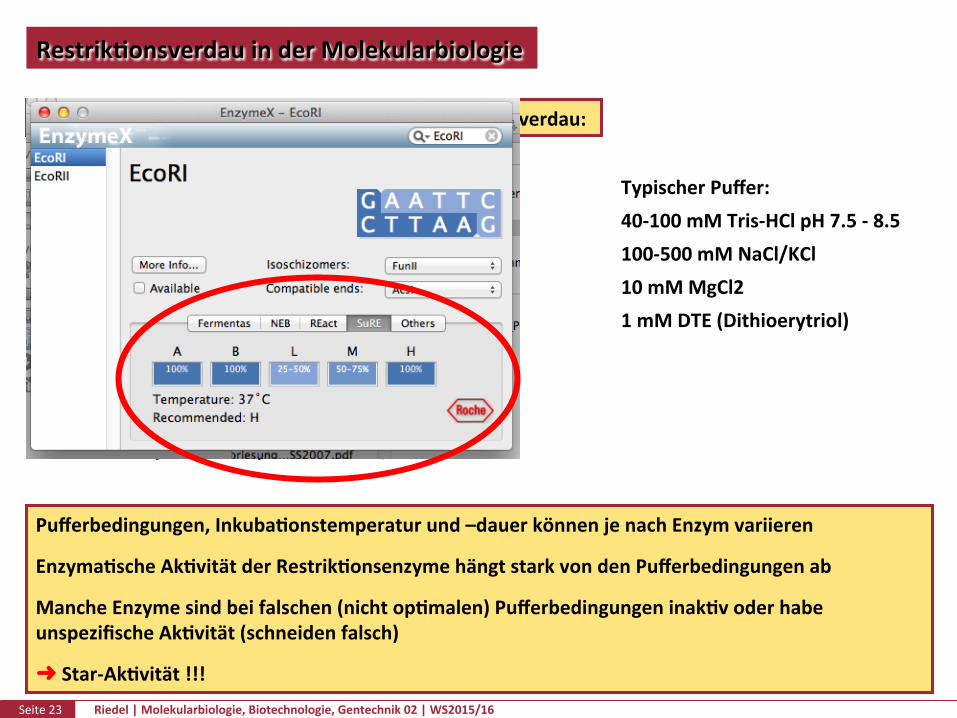
InderPraxis:AnsatzfüreinenanalyPschenResrikPonsverdau:
1µlDNA-lösung(1µg)2µl10xPuffer1µlRestrikPonsendonuklease(2U)16µlH2O
InkubaPonfür1-2h,inderRegelbei37°C
TypischerPuffer:40-100mMTris-HClpH7.5-8.5100-500mMNaCl/KCl10mMMgCl21mMDTE(Dithioerytriol)
Pufferbedingungen,InkubaPonstemperaturund–dauerkönnenjenachEnzymvariieren
EnzymaPscheAkPvitätderRestrikPonsenzymehängtstarkvondenPufferbedingungenab
MancheEnzymesindbeifalschen(nichtopPmalen)PufferbedingungeninakPvoderhabeunspezifischeAkPvität(schneidenfalsch)
➜Star-AkPvität!!!
AkPvität(Menge)derEnzymeinUnits(U)Standard-Enzym:1U=1µmolSubstrat/min
Seite24 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsverdauinderMolekularbiologie
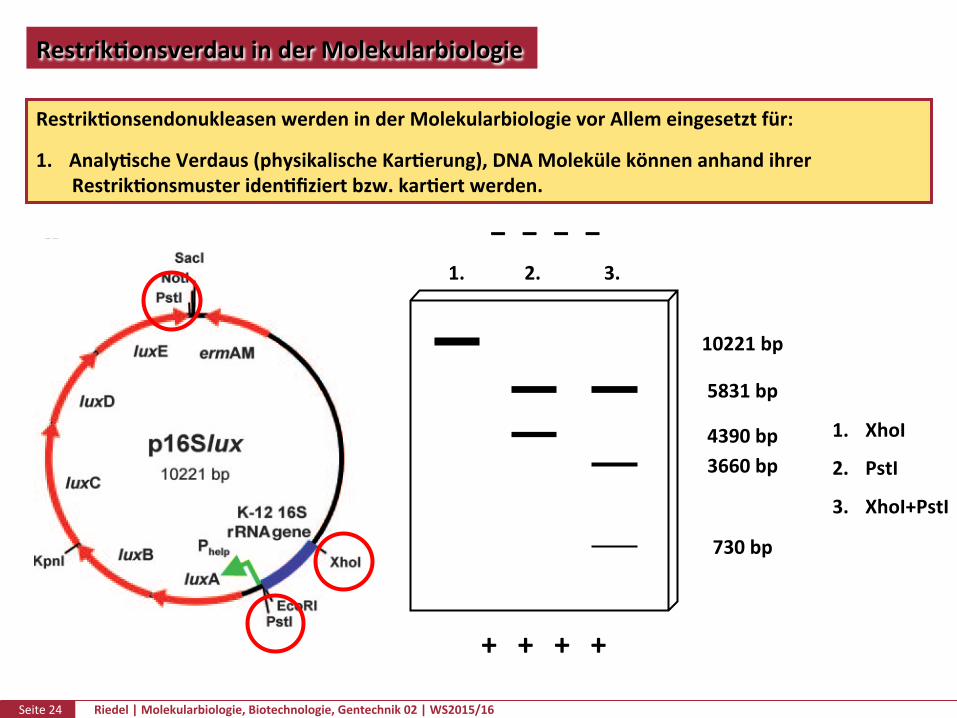
RestrikPonsendonukleasenwerdeninderMolekularbiologievorAllemeingesetztfür:
1. AnalyPscheVerdaus(physikalischeKarPerung),DNAMolekülekönnenanhandihrerRestrikPonsmusteridenPfiziertbzw.karPertwerden.
cence are critical (e.g., in comparisons of wild-type and mutantstrains), relative levels of luminescence or copy numbers ofintegrants should be verified. Eryr colonies were checked forlight emission, and the integration of p16Slux was confirmedby PCR using primers 16S_rev_XhoI and 16S_fwd_int (5!-ATTAGCTAGTAGGTGGGGTAACGGCTCACCTAGG-3!).16S_fwd_int anneals upstream of 16S_fwd_Econew in the 16SrRNA gene sequence and yields a 1,150-bp PCR fragment onlyif p16Slux is integrated at the correct site in the chromosome.By using this protocol, the following organisms were renderedbioluminescent: E. coli DH10B, C. rodentium ICC169, Salmo-nella enterica serovar Typhimurium UK-1, P. aeruginosa PAO1,Enterobacter sakazakii DPC6440, Shigella flexneri 2a ATCC700930, and Y. enterocolitica NCTC13174 (Fig. 1B). In all lux-tagged strains, plasmid integration was stable in the absence ofantibiotic for at least 50 generations. All lux-labeled strainswere tested for differences in growth on complex media in aSpectraMax M2, 96-well plate reader (Molecular Devices,Sunnyvale, CA). At the time points indicated in Fig. 1, lumi-nescence was measured in relative light units (RLU, in photonss"1) with the IVIS100 imaging system. As shown in Fig. 1C forC. rodentium and S. enterica serovar Typhimurium UK-1, noneof the tagged strains exhibited a significant difference from thewild-type strain in growth rate and final optical density at 600nm on LB media (Fig. 1C and data not shown).
All gram-negative strains presented here produced intense lu-minescence (#1 $ 107 to 5 $ 107 RLU) in LB broth in 96-wellplates (Fig. 1C and data not shown). Luminescence in the gram-negative strains was approximately 10-fold higher than the lumi-nescence observed in L. monocytogenes EGDe::pPL2luxPhelp,
which harbors the same luxPhelp construct as a single-copy chro-mosomal integration (10). Also, luminescence in the gram-nega-tive strains was about 10-fold higher than that in the gram-positivestrains containing p16Slux replicating as a plasmid (data notshown). Additionally, while luminescence in L. monocytogenes(10) and other gram-positive bacteria (data not shown) decreaseddramatically in stationary phase, luminescence in all gram-nega-tive bacteria tested remained relatively stable throughout the en-tire growth curve (Fig. 1C and data not shown). Whether thesedifferences are due to the availability of the substrate of thereaction or to differences in the intracellular conditions remainsto be elucidated.
To assess the functionality of the Phelp-driven expression ofluxABCDE in gram-negative bacteria in vivo, three of the la-beled strains were tested in murine models of infection in 6- to8-week-old conventional female BALB/c mice and comparedto their wild-type strains. C. rodentium, S. enterica serovarTyphimurium, and P. aeruginosa were chosen as models fornoninvasive intestinal, invasive intestinal, and invasive pulmo-nary pathogens, respectively. Animals were kept in a conven-tional animal colony, and all experiments were approved by theanimal ethics committee of University College Cork. Animalswere infected by gavage (C. rodentium and S. enterica serovarTyphimurium) or intranasally (P. aeruginosa) with bacteriafrom overnight cultures washed with phosphate-buffered saline(PBS). Doses used were 2 $ 109 CFU/animal in 100 %l of PBSfor C. rodentium, 1 $ 107 CFU/animal in 100 %l PBS for S.enterica serovar Typhimurium, and 1 $ 106 CFU/animal in 10%l PBS for P. aeruginosa. At various time points during infec-tion (Fig. 2), animals were anesthetized with isoflurane and
FIG. 1. (A) Plasmid map of p16Slux with relevant restriction sites and arrangement of the E. coli DH10B 16S sequence (blue), the Phelp promoterregion (green arrow), and luxABCDE (red arrows). (B) Gram-negative strains tagged by chromosomal integration of p16Slux (i, E. coli DH10B::p16Slux;ii, C. rodentium ICC169::p16Slux; iii, S. enterica serovar Typhimurium UK-1::p16Slux; iv, P. aeruginosa PAO1::p16Slux; v, E. sakazakii DPC6440::p16Slux;vi, S. flexneri 2a ATCC 700930::p16Slux; vii, Y. enterocolitica NCTC13174::p16Slux). The color bar indicates bioluminescence signal intensity (in photonss"1 cm"2). Strains were grown on LB agar plates containing erythromycin under nonpermissive conditions and imaged using the Xenogen IVIS100imaging system. min, minimum; max, maximum. (C) Growth (symbols) and luminescence (bars) of wild-type (wt) and lux-tagged strains of C. rodentiumICC169 (C. rod) and S. enterica serovar Typhimurium UK-1 (S. typh) on LB medium. Data for luminescence are presented as mean RLU & standarddeviations of results for four wells, and the results from one representative of three independent experiments are shown. The top panel showsrepresentative wells containing the indicated strain in LB broth at the indicated time points.
VOL. 73, 2007 BIOLUMINESCENT IMAGING OF GRAM-NEGATIVE BACTERIA 7093
+ + + +
− − − −1. 2. 3.
1. XhoI2. PstI
3. XhoI+PstI
10221bp
3660bp
5831bp
4390bp
730bp
Seite25 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsverdauinderMolekularbiologie
Seite26 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsverdauinderMolekularbiologie
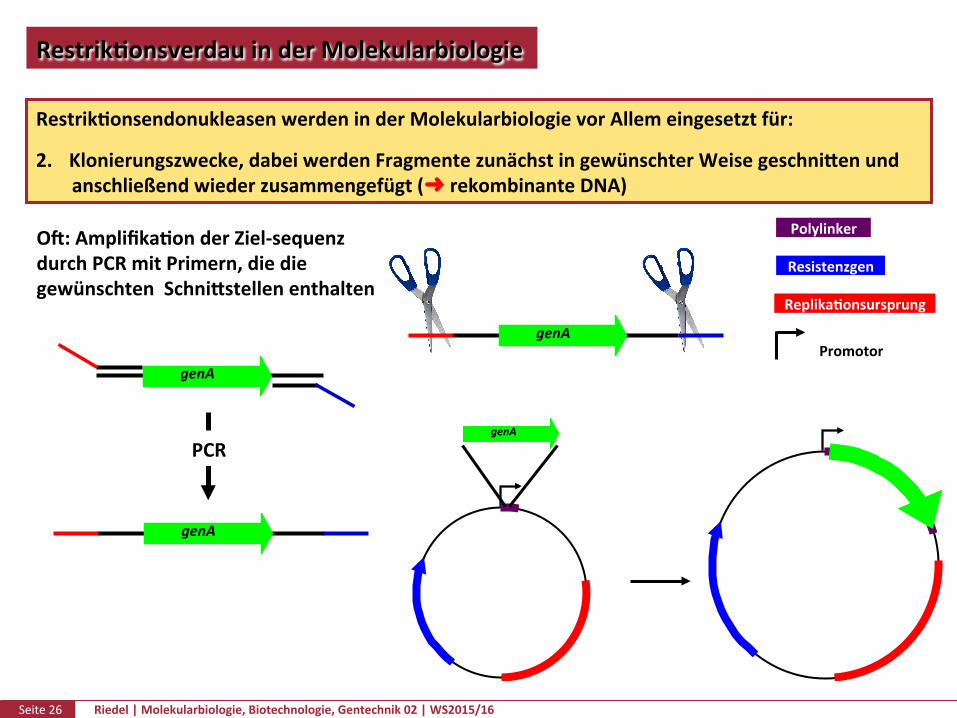
RestrikPonsendonukleasenwerdeninderMolekularbiologievorAllemeingesetztfür:
2. Klonierungszwecke,dabeiwerdenFragmentezunächstingewünschterWeisegeschnikenundanschließendwiederzusammengefügt(➜rekombinanteDNA)
genA%
Resistenzgen)
Replika.onsursprung)
Polylinker)
Promotor)genA
Of:AmplifikaPonderZiel-sequenzdurchPCRmitPrimern,diediegewünschtenSchnikstellenenthalten
genA
PCR
genA

Seite27 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
MethodenderMolekularbiologie
ModifikaPonvonDNA:LigaPon
Seite28 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsendonukleasen
WeitereArbeitsschrike,diefürdieKlonierungvonDNAwichPgsind: VerbindenvonFragmenten
Ligase=molekularerKleberfür(kompaPble)DNA-Enden
BisherigeMethodenfürArbeitenmitDNA: IsolaPon QualitätskontrolleundQuanPfikaPon VervielfälPgung(PCR) Agarose-Gelelektrophorese
Schneiden(RestrikPonsverdau
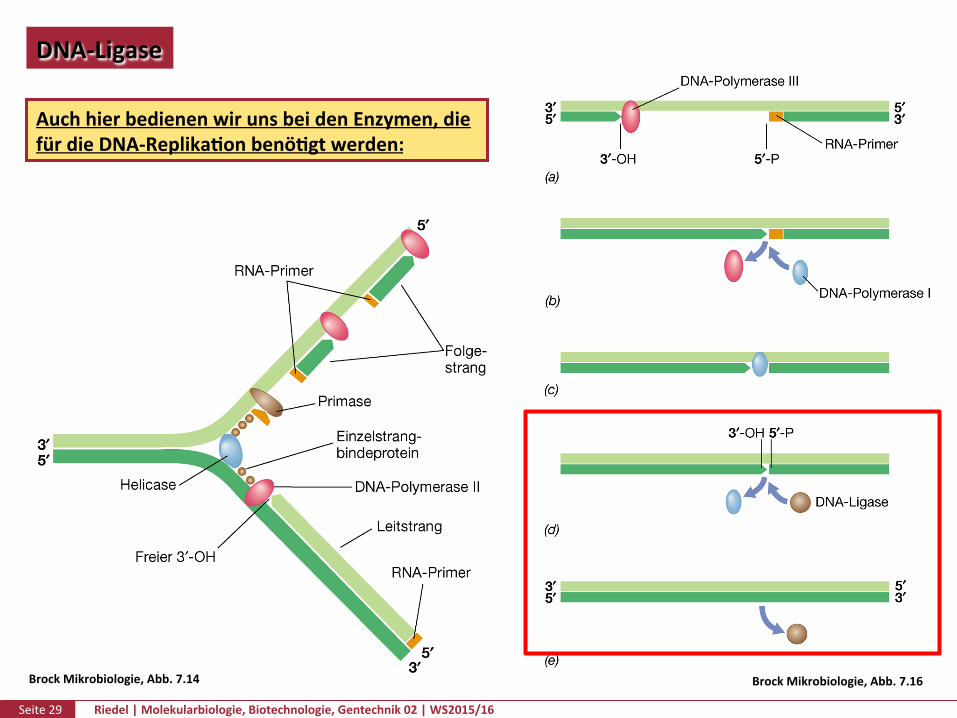
Seite29 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNA-Ligase
AuchhierbedienenwirunsbeidenEnzymen,diefürdieDNA-ReplikaPonbenöPgtwerden:
BrockMikrobiologie,Abb.7.16BrockMikrobiologie,Abb.7.14
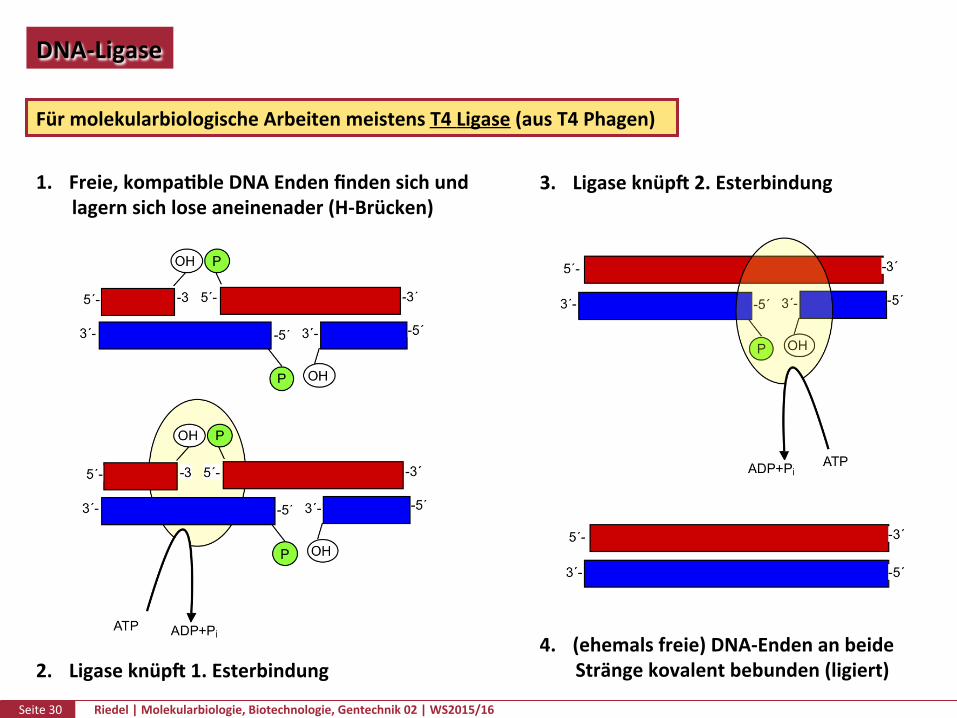
Seite30 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNA-Ligase
FürmolekularbiologischeArbeitenmeistensT4Ligase(ausT4Phagen)
1. Freie,kompaPbleDNAEndenfindensichundlagernsichloseaneinenader(H-Brücken)
2. Ligaseknüpf1.Esterbindung
3. Ligaseknüpf2.Esterbindung
4. (ehemalsfreie)DNA-EndenanbeideSträngekovalentbebunden(ligiert)
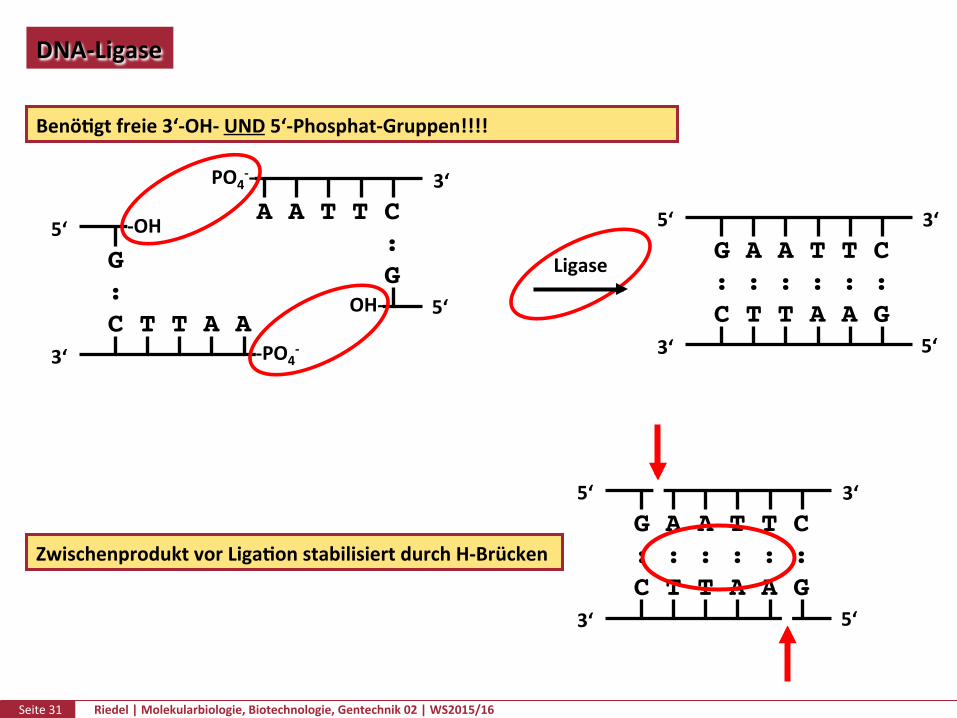
Seite31 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNA-Ligase
ZwischenproduktvorLigaPonstabilisiertdurchH-Brücken
BenöPgtfreie3‘-OH-UND5‘-Phosphat-Gruppen!!!!
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
-OH
-PO4-
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
OH-
PO4--
Ligase
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
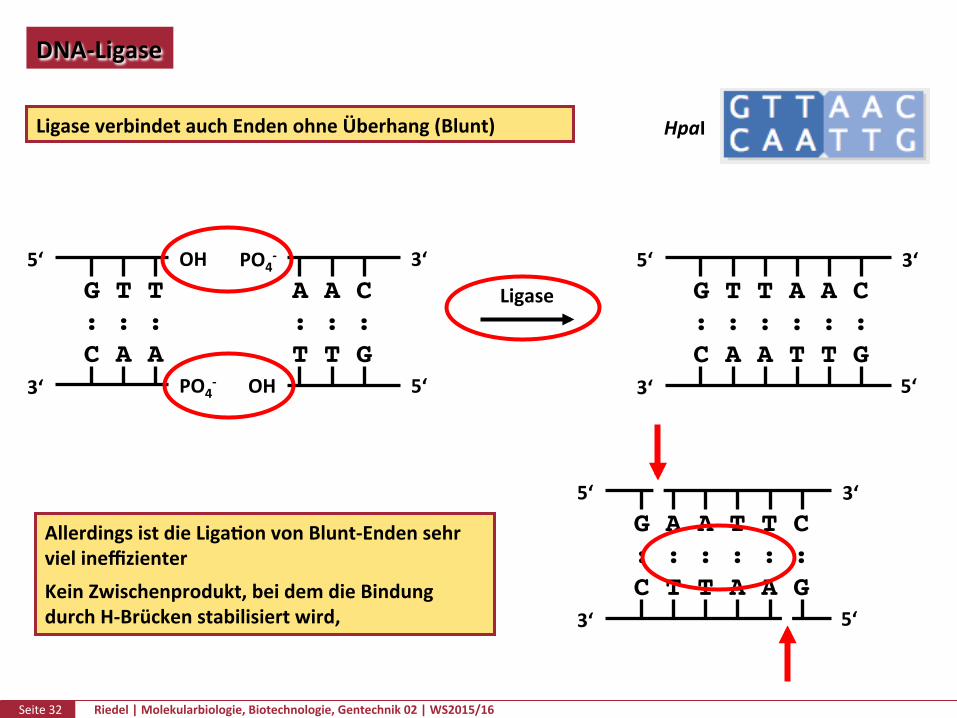
Seite32 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
DNA-Ligase
LigaseverbindetauchEndenohneÜberhang(Blunt)
3‘
5‘
PO4-
OHG T T: : : C A A
HpaI
OH 5‘
3‘A A C: : : T T G
PO4-
Ligase
3‘
5‘
5‘
3‘G T T A A C: : : : : :C A A T T G
AllerdingsistdieLigaPonvonBlunt-EndensehrvielineffizienterKeinZwischenprodukt,beidemdieBindungdurchH-Brückenstabilisiertwird, 3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G

Seite33 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
MethodenderMolekularbiologie
ModifikaPonvonDNA:weitere
Seite34 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
RestrikPonsendonukleasen
BisherigeMethodenfürArbeitenmitDNA: IsolaPon QualitätskontrolleundQuanPfikaPon VervielfälPgung(PCR) Agarose-Gelelektrophorese
RestriPonsverdau LigaPon
ModifikaPonfreierEnden
ModifikaPonfreierEnden
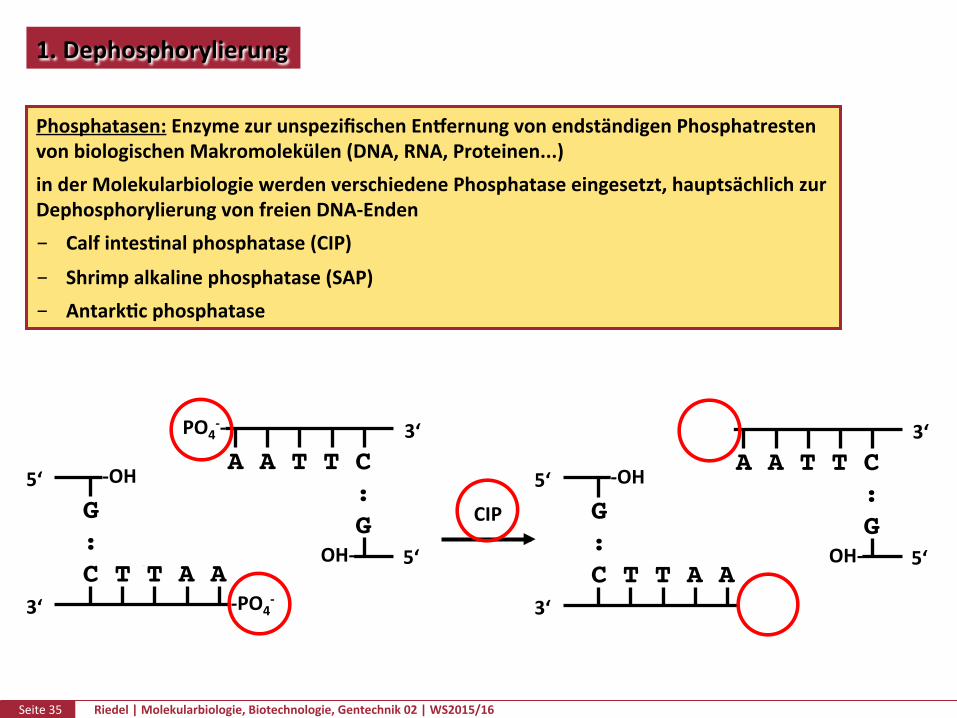
Seite35 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
1.Dephosphorylierung
Phosphatasen:EnzymezurunspezifischenEn�ernungvonendständigenPhosphatrestenvonbiologischenMakromolekülen(DNA,RNA,Proteinen...)inderMolekularbiologiewerdenverschiedenePhosphataseeingesetzt,hauptsächlichzurDephosphorylierungvonfreienDNA-Enden CalfintesPnalphosphatase(CIP)
Shrimpalkalinephosphatase(SAP) AntarkPcphosphatase
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
-OH
-PO4-
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
OH-
PO4--
CIP
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
-OH
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
OH-
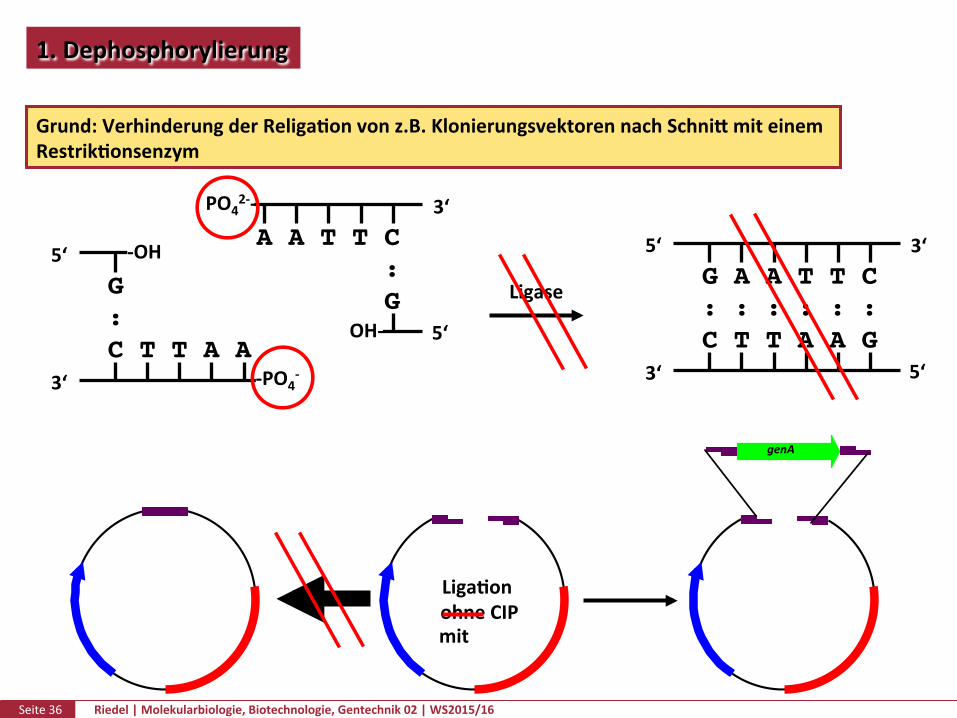
Seite36 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
1.Dephosphorylierung
Grund:VerhinderungderReligaPonvonz.B.KlonierungsvektorennachSchnikmiteinemRestrikPonsenzym
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
-OH
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
OH-
Ligase
3‘
5‘
5‘
3‘G A A T T C: : : : : :C T T A A G
genA%
LigaPonohneCIPmit
PO42--
-PO4-
Seite37 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
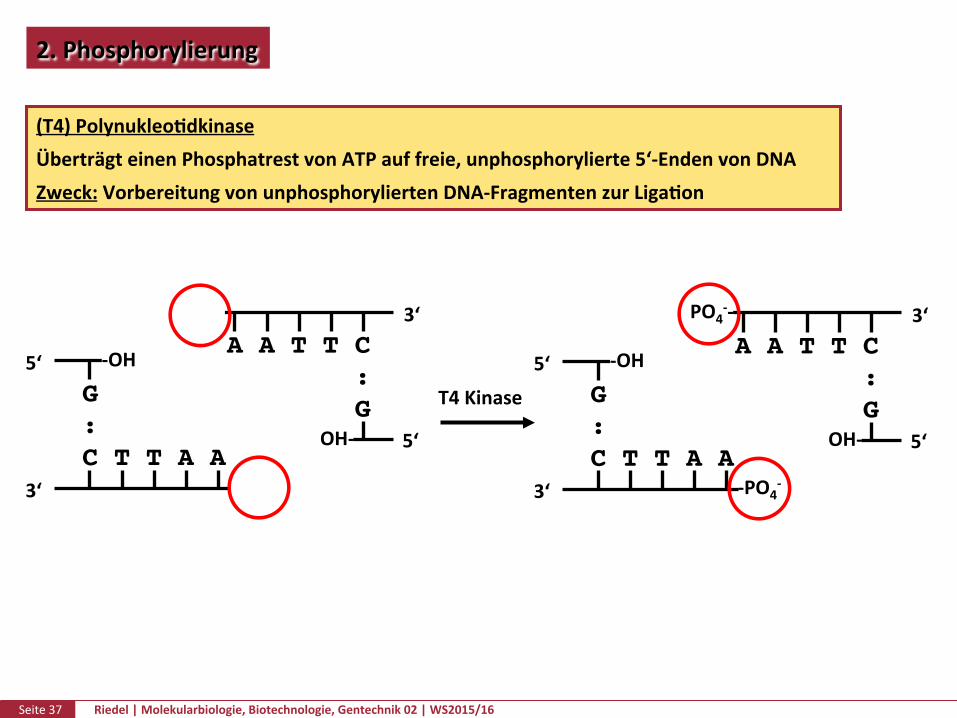
2.Phosphorylierung
(T4)PolynukleoPdkinaseÜberträgteinenPhosphatrestvonATPauffreie,unphosphorylierte5‘-EndenvonDNAZweck:VorbereitungvonunphosphoryliertenDNA-FragmentenzurLigaPon
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
-OH
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
OH-
T4Kinase
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
-OH
3‘
5‘
5‘
3‘G A A T T C: :C T T A A G
OH-
PO4--
-PO4-
Seite38 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
2.Phosphorylierung
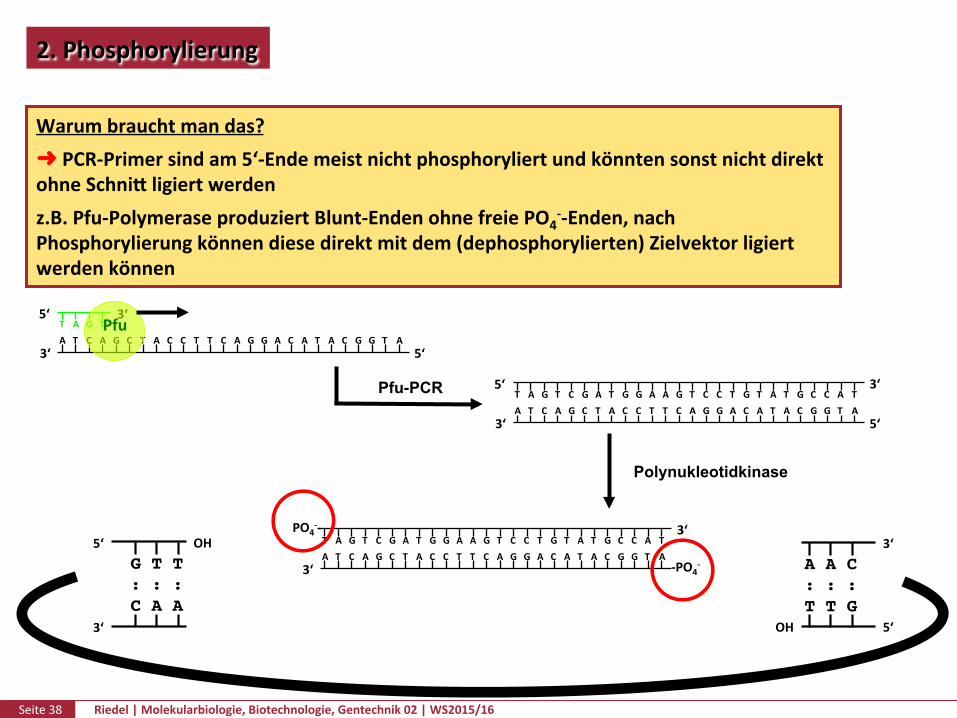
Warumbrauchtmandas?➜PCR-Primersindam5‘-EndemeistnichtphosphoryliertundkönntensonstnichtdirektohneSchnikligiertwerdenz.B.Pfu-PolymeraseproduziertBlunt-EndenohnefreiePO4
--Enden,nachPhosphorylierungkönnendiesedirektmitdem(dephosphorylierten)Zielvektorligiertwerdenkönnen
A" T" C" A" G" C" T" A" C" C" T" T" C" A" G" G" A" C" A" T" A" C" G" G" T" A"T" A" G" T"
3‘"
5‘"
5‘"
3‘"Pfu"
A" T" C" A" G" C" T" A" C" C" T" T" C" A" G" G" A" C" A" T" A" C" G" G" T" A"T" A" G" T" C" G" A" T" G" G" A" A" G" T" C" C" T" G" T" A" T" G" C" C" A" T"
3‘"
5‘"
5‘"
3‘"Pfu-PCR
A" T" C" A" G" C" T" A" C" C" T" T" C" A" G" G" A" C" A" T" A" C" G" G" T" A"T" A" G" T" C" G" A" T" G" G" A" A" G" T" C" C" T" G" T" A" T" G" C" C" A" T"
3‘"
PO4++"
+PO4+"
3‘"
Polynukleotidkinase
OH# 5‘#
3‘#A A C!: : : !T T G !
3‘#
5‘# OH#G T T!: : : !C A A !
Seite39 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
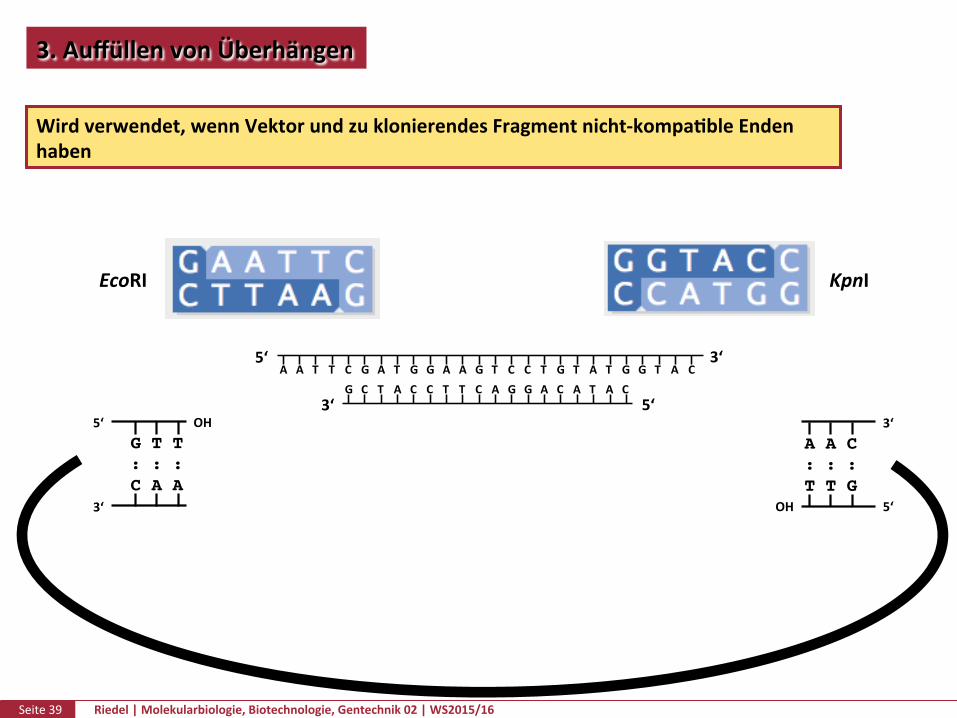
3.AuffüllenvonÜberhängen
Wirdverwendet,wennVektorundzuklonierendesFragmentnicht-kompaPbleEndenhaben
OH# 5‘#
3‘#A A C!: : : !T T G !
3‘#
5‘# OH#G T T!: : : !C A A !
EcoRI KpnI
G" C" T" A" C" C" T" T" C" A" G" G" A" C" A" T" A" C"A" A" T" T" C" G" A" T" G" G" A" A" G" T" C" C" T" G" T" A" T" G" G" T" A" C"
3‘"
5‘"
5‘"
3‘"

Seite40 Riedel|Molekularbiologie,Biotechnologie,Gentechnik02|WS2015/16
3.AuffüllenvonÜberhängen
Wirdverwendet,wennVektorundzuklonierendesFragmentnicht-kompaPbleEndenhaben
Enzym:Klenow-Fragment=dasgrößerederbeidenProteinfragmentederDNA-PolymeraseIausE.colinachenzymaPscherSpaltungmitSubPlisin
Besitzt2enzymaPscheAkPvitäten:
1. 5‘→3'Polymerase-AkPvität2. 3‘→5'Exonuklease-AkPvität(proofreading)
UrsprünglichesEnzymzurAmplifikaPonvonDNAdurchPCR(vorVerwendungvonTaq)
2. 3‘→5'Exonuklease-AkPvität
G C T A C C T T C A G G A C A T A CA A T T C G A T G G A A G T C C T G T A T G G T A C
3‘
5‘
5‘
3‘
AT AT
3‘
3‘
1. 5‘→3'Polymerase-AkPvität
PhosphorylierungundLigaPon