Embed Size (px)
Citation preview

1
Metabolic engineering of Synechocystis 6803 for isobutanol 1
production 2
3
Arul M. Varman1, Yi Xiao1, Himadri B. Pakrasi1,2 and Yinjie J. Tang 1,a 4
5
1Department of Energy, Environmental and Chemical Engineering 6
2Department of Biology, Washington University, St. Louis, MO 63130, USA 7
aCorresponding author 8
9
(Running title: Synechocystis 6803 for isobutanol production) 10
Email addresses: 11
Arul M. Varman: [email protected] 12
Yi Xiao: [email protected] 13
Himadri B. Pakrasi: [email protected] 14
Yinjie J. Tang: [email protected] 15
16
17
18
19
Copyright © 2012, American Society for Microbiology. All Rights Reserved.Appl. Environ. Microbiol. doi:10.1128/AEM.02827-12 AEM Accepts, published online ahead of print on 26 November 2012
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

2
Abstract 20
Global warming and decreasing fossil fuel reserves have prompted great interest in the 21
synthesis of advanced biofuels from renewable resources. In an effort to address these concerns, 22
we have performed metabolic engineering of the cyanobacterium Synechocystis sp. PCC 6803 to 23
develop a strain that can synthesize isobutanol under both autotrophic and mixotrophic 24
conditions. With the expression of two heterologous genes from the Ehrlich Pathway, the 25
engineered strain can accumulate 90 mg/L of isobutanol from 50 mM bicarbonate in a gas-tight 26
shaking flask. This strain does not require any inducer (i.e., IPTG: Isopropyl β-D-1-27
thiogalactopyranoside) or antibiotics to maintain its isobutanol production. In the presence of 28
glucose, isobutanol synthesis is only moderately promoted (titer = 114 mg/L). Based on 29
isotopomer analysis, we find that compared to the wild-type strain, the mutant significantly 30
reduced its glucose utilization and mainly employed autotrophic metabolism for biomass growth 31
and isobutanol production. Since isobutanol is toxic to the cells and may also be degraded 32
photochemically by hydroxyl radicals during the cultivation process, we employed in situ 33
removal of the isobutanol using oleyl alcohol as a solvent trap. This resulted in a final net 34
concentration of 298 mg/L of isobutanol under mixotrophic culture conditions. 35
Key words: bicarbonate, Ehrlich pathway, inducer, isobutanol, isotopomer analysis, oleyl alcohol 36
37
38
39
40
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

3
Introduction 41
Global energy needs continue to increase rapidly due to industrial and development 42
demands, furthering environmental concerns. Much of the worldwide energy consumption 43
comes from the burning of fossil fuels, which produces about 6 gigatons of CO2 annually (27). 44
Increasing CO2 levels may act as a feedback loop to increase the soil emissions of other 45
greenhouse gases such as methane and nitrous oxide, heightening global temperature (32). For 46
energy security and environmental concerns, there is an urgent demand for the development of 47
bioenergy. Bioethanol is the most common biofuel, but it also has low energy density and 48
absorbs moisture. Isobutanol (IB) is a better fuel because it is less water soluble and has an 49
energy density / octane value close to that of gasoline (18, 20). Amongst the next generation 50
biofuels synthesized from pyruvate, IB possesses fewer reaction steps (5 reaction steps from 51
pyruvate to IB) in contrast to the synthesis of 1-butanol or biodiesel. IB is less toxic to microbes 52
(1) so that it may achieve higher product titer and yield (7, 33). For example, a maximum titer of 53
50.8 g/L of IB can be achieved in an engineered E. coli (3). 54
On the other hand, cyanobacteria can not only convert CO2 into bio-products, but also 55
can play an important role in environmental bioremediations. The photosynthetic efficiency of 56
cyanobacteria (3~9%) is high compared to higher plants (≤0.25~3%) (11, 40). Furthermore, 57
some species of cyanobacteria are amenable to genetic engineering. Table 1 lists the various 58
biofuels that have been synthesized through the metabolic engineering of cyanobacteria. 59
Autotrophic IB production in cyanobacteria was first demonstrated in Synechococcus 7942 (2). 60
Moreover, a model cyanobacterium Synechocystis sp. PCC 6803 is capable of growing under 61
both photoautotrophic and mixotrophic conditions, while the presence of glucose can 62
significantly promote biomass and bioproduct synthesis (26). Thereby, we have engineered a 63
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

4
glucose tolerant Synechocystis 6803 strain with two key genes kivd and adhA of the Ehrlich 64
pathway (28) so that the cyanobacterial strain can convert CO2 into IB. Through both metabolic 65
engineering and bioprocess optimization, we have improved our strain’s IB production 66
capabilities. 67
Materials and methods 68
Chemicals and reagents. Restriction enzymes, T4 DNA ligase, DNase and a Revertaid first 69
strand cDNA synthesis kit were purchased from Fermentas or New England Biolabs. 70
Oligonucleotides were purchased from Integrated DNA Technologies. Toluene, IB, α-71
ketoisovaleric acid, phenol and chloroform were purchased from Sigma-Aldrich (St. Louis, MO). 72
KlenTaq-LA (4) was purchased from DNA Polymerase Technology (St. Louis, MO). TRI 73
Reagent® was purchased from Ambion, USA. 13C labeled glucose was purchased from 74
Cambridge Isotope Laboratories. 75
Culture medium and growth conditions. A glucose tolerant wild-type strain of Synechocystis 76
6803 (WT) and the recombinant strain AV03 were grown at 30°C in liquid BG-11 medium or 77
solid BG-11 medium at a light intensity of 50 µmol of photons m-2 s-1 in ambient air. 78
Kanamycin at a concentration of 20 µg/mL was added to the BG 11 medium when required. 79
Growth of the cells was monitored by measuring OD730 of the cultures on an Agilent Cary 60 80
UV-Vis Spectrophotometer. Cultures for the synthesis of IB were grown in 10 mL medium 81
(Initial OD730 of 0.4) in 50 mL shake flasks for 4 days. The mid-log phase cultures were then 82
closed with rubber caps to prevent the loss of IB during incubation, and the cultures were 83
supplemented with 50 mM NaHCO3 as an inorganic carbon source. Mixotrophic cultures of 84
Synechocystis 6803 were started in a BG-11 medium containing a known amount of glucose as 85
an organic carbon source. E. coli strain DH10B was the host for all plasmids constructed in this 86
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

5
study. E. coli cells were grown in falcon tubes containing Luria-Bertani (LB) medium at 37°C 87
under continuous shaking. Ampicillin (100 µg/mL) or kanamycin (50 µg/mL) was added to the 88
LB medium when required, for the propagation of plasmids in E. coli. 89
Plasmid construction and transformation of Synechocystis 6803. The vector pTAC-KA 90
containing an ampicillin resistance cassette (AmpR) and two genes (kivd and adhA from 91
Lactococcus lactis) was constructed as described (36). The pTAC-KA vector was modified using 92
the following steps to convert it into a Synechocystis 6803 vector. To clone the flanking regions 93
of a potential neutral site into pTAC-KA, a Synechocystis 6803 vector pSL2035 containing both 94
the flanking regions and the kanamycin resistance cassette (KmR) was used as a template. 95
pSL2035 is a Synechocystis 6803 vector designed to integrate any foreign DNA into the genome 96
of Synechocystis 6803 by replacing the psbA1 gene and its promoter. psbA1 is a member of psbA 97
gene family and is found to be silent under most conditions (24, 25). pSL2035 was constructed 98
by cloning the flanking regions for the psbA1 gene and the KmR into pUC118. The 5’ flanking 99
region from pSL2035 was PCR amplified along with KmR by respective primers (Table 2) and 100
cloned into the PciI and Bsu36I site of pTAC-KA, resulting in the vector pTKA2. The 3’ 101
flanking region was PCR amplified from pSL2035 by the respective primers and inserted into the 102
AhdI site of pTKA2, disrupting the native AmpR and henceforth creating the vector pTKA3. 103
Transformation was performed by using a double homologous recombination system, and 104
the genes were integrated into the target site of the Synechocystis genomic DNA. Specifically, 2 105
mL of Synechocystis 6803 from a mid-log phase (1~3×108 cells mL-1) culture was centrifuged at 106
10,000 × g for 2 min. The pellet was suspended in a fresh BG-11 medium (200 µL) to a final cell 107
density of 1~3×109 cells mL-1. Plasmid DNA was added to a final DNA concentration of 5~10 108
µg/mL (38) to this dense Synechocystis 6803 cell culture. The mixture was then incubated under 109
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

6
normal light conditions (50 µE m-2 s-1) overnight. The culture was then spread onto a BG-11 agar 110
plate containing 20 µg/mL of kanamycin. Recombinant colonies usually appear between 7 and 111
10 days. Colonies were propagated on a fresh BG-11 plate containing kanamycin, and a colony 112
PCR was performed to verify successful integration of the insert into the genomic DNA of the 113
recombinant. The positive colonies were propagated continuously onto BG-11 plates containing 114
kanamycin, to get a high segregation of the insert in the recombinant (34). To verify the integrity 115
of the promoter and gene sequences, the heterologous DNA integrated into the genome of the 116
mutant AV03 was PCR amplified and sent for sequencing with the respective primers. 117
Reverse transcription PCR (RT-PCR). Total RNA isolation of Synechocystis 6803 was 118
performed using a TRI Reagent® (Ambion, USA) by following the manufacturer protocol with 119
modifications. 1 mL of RNAwiz was prewarmed to 70°C and pipetted into the frozen cells 120
directly. Immediately, the mixture was vortexed and incubated for 10 min at 70°C in a heater 121
block. 0.2 mL of chloroform was added to the mixture and mixed vigorously followed by 122
incubation at room temperature for 10 min. The aqueous and the organic phase were separated 123
by centrifugation at 10,000 × g at 4°C. The RNA containing aqueous phase was transferred into 124
an eppendorf tube, to which, equal volumes of phenol and chloroform were added. The mixture 125
was mixed vigorously followed by centrifugation to separate the aqueous and the organic phase. 126
The aqueous phase was removed to a clean tube, to which 0.5 mL of diethyl pyrocarbonate 127
(DEPC) treated water was added. RNA in the solution was precipitated by the addition of room 128
temperature isopropanol and centrifuged at 10,000 × g at 4°C to pellet the RNA. The RNA was 129
washed with ethanol and resuspended in a fresh 50 µL of DEPC treated water. The quantity and 130
quality of the isolated RNA was determined using a Nanodrop ND-1000 (Thermo Scientific, 131
USA). The RNA was incubated at room temperature with DNase to degrade any genomic DNA, 132
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

7
if present in the RNA sample. Synthesis of cDNA was performed by utilizing a Reverse 133
transcriptase enzyme from Fermentas along with dNTPs and random primers in a reaction buffer. 134
The mixture was incubated at 42°C for 60 min. The synthesized cDNA was used as a template 135
for the PCR, to detect the expression of the mRNA of interest. 136
Isobutanol quantification assay. IB synthesized in the culture was quantified using a gas 137
chromatograph (Hewlett Packard model 7890A, Agilent Technologies, equipped with a DB5-MS 138
column, J&W Scientific) and a mass spectrometer (5975C, Agilent Technologies). IB extraction 139
was done using a modified procedure (5). Samples of the cyanobacterial culture (400 µL) were 140
collected and centrifuged at 10000 × g for 5 min. IB was extracted from the supernatant by 141
vortexing for 1 min with 400 µL of toluene, and methanol was used as the internal standard. A 1 142
µL sample of the organic layer was injected into the gas chromotagraph (GC) with helium as the 143
carrier gas. The GC oven was held at 70°C for 2 min and then raised to 200°C with a 144
temperature ramp of 30°C min-1, and the post run was set at 300°C for 6 min. The range of the 145
mass spectrometer (MS) scan mode was set between m/z of 20 and 200. The concentration of IB 146
present in the culture was determined based on a calibration curve prepared with known 147
concentrations of IB ranging from 25 mg/L to 400 mg/L. 148
13C-experiment to detect carbon contribution of glucose. The 13C-abundance of some 149
important metabolites was measured for both the wild-type and the mutant strain AV03, to 150
estimate the carbon contribution of both glucose (fully labeled by 13C) and nonlabeled 151
bicarbonate for biomass and IB synthesis. Mixotrophic cultures of both the wild-type 152
Synechocystis 6803 and the mutant AV03 were grown in BG-11 medium (with 50mM 153
nonlabeled NaHCO3), which contained 0.5% glucose (U-13C, Cambridge Isotope Laboratories, 154
MA). Cultures were collected on day 3, 6 and 9, and proteinogenic amino acids were hydrolyzed 155
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

8
and then derivatized with TBDMS (N-(tert-butyldimethylsilyl)-N-methyl-trifluoroacetamide, 156
Sigma-Aldrich). The derivatized amino acids were analyzed for their mass isotopomer 157
abundance by GC-MS, as described before (13, 37). The m/z ion [M-57]+, which corresponds to 158
the entire amino acid, was used to calculate the 13C abundance in amino acids [m0 m1…. mn]. 159
The fraction of carbon (FA) derived from fully labeled glucose for each amino acid was estimated 160
based on the following equation: n
miF
n
i iA =
×= 1 (Equation 1) 161
where i is the number of labeled carbons, mi is the mass fraction for different isotopomers of the 162
corresponding amino acid and n represents the total number of carbons in the corresponding 163
amino acid. The m/z of [M-15]+ was used only for leucine and isoleucine, since their [M-57]+ 164
overlaps with other mass peaks (35). IB extraction was performed for samples obtained from the 165
above cultures and was analyzed using the GC-MS. The fraction of carbon derived from glucose 166
for isobutanol (FIB) was estimated based on the isobutanol MS peak abundances: 167
=
=×
=4
0
4
1
4i i
i iIB
A
AiF (Equation 2) 168
where Ai is the abundance of the mass-to-charge ratio peaks for the various isobutanol 169
isotopomers (i.e., A0~A4 for m/z=74~78). 170
Results 171
Construction of an isobutanol producing Synechocystis 6803 strain. IB synthesis in 172
Synechocystis 6803 requires the expression of two heterologous genes of the Ehrlich pathway. 173
The enzymes 2-keto-acid decarboxylase and alcohol dehydrogenase can convert 2-keto acids 174
into alcohols. In this work, we constructed a plasmid pTKA3 containing the genes kivd and adhA 175
from Lactococcus lactis under the control of an IPTG inducible promoter, Ptac. The plasmid was 176
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

9
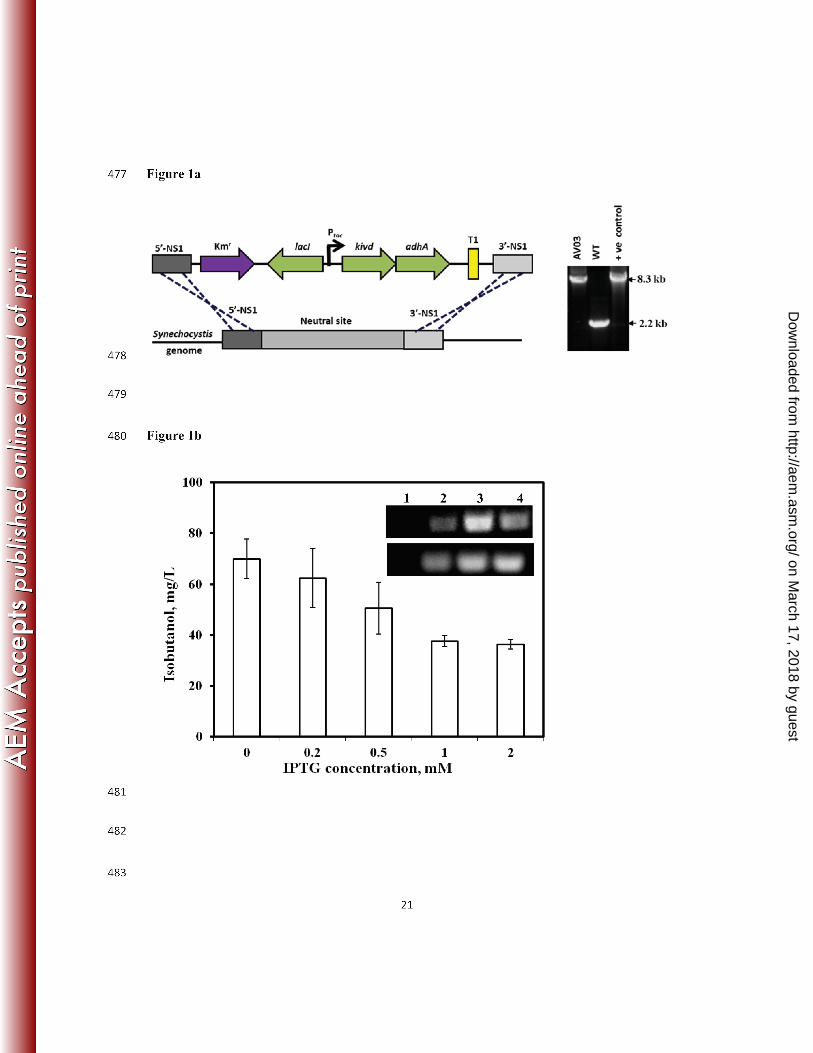
designed to integrate the genes into a neutral site in the genome of Synechocystis 6803, along 177
with a kanamycin resistance cassette (Fig. 1a - Left). The wild-type strain of Synechocystis 6803 178
was transformed with pTKA3, resulting in the recombinant strain AV03. The integration of the 179
insert genes into the genome was verified by a colony PCR after several rounds of segregation 180
(Fig. 1a - Right). 181
To identify the optimal IPTG concentration required for IB synthesis, the AV03 strain 182
was grown under different concentrations of IPTG. IB analysis from the cultures indicated that 183
IB was highly synthesized even without the addition of IPTG (Fig. 1b). To verify if this 184
observation was an artifact of any mutations that might have occurred in lacI or the promoter, the 185
foreign DNA integrated into the chromosome of AV03 was sequenced. Sequencing results for 186
the lacI and the promoter Ptac in the genome of AV03 revealed that the nucleotide sequence was 187
completely intact. There have been reports of leaky expressions with IPTG inducible promoters 188
(17). Besides, Fig. 1b indicates that as the concentration of IPTG went higher than 1mM, the IB 189
synthesis reduced. The OD730 of the different cultures indicated that the addition of IPTG did 190
not apparently interfere with the growth rate of the culture. RT-PCR showed the expression 191
levels of the genes kivd and adhA under different IPTG concentrations. The result of RT-PCR 192
experiment (Fig. 1b - Inset) indicated that the levels of kivd and adhA mRNA synthesized in the 193
mutant were higher with IPTG than without. Henceforth, lower expression of the two genes is 194
sufficient for IB synthesis, possibly because the Ehrlich pathway may not be the rate-limiting 195
step for IB production. 196
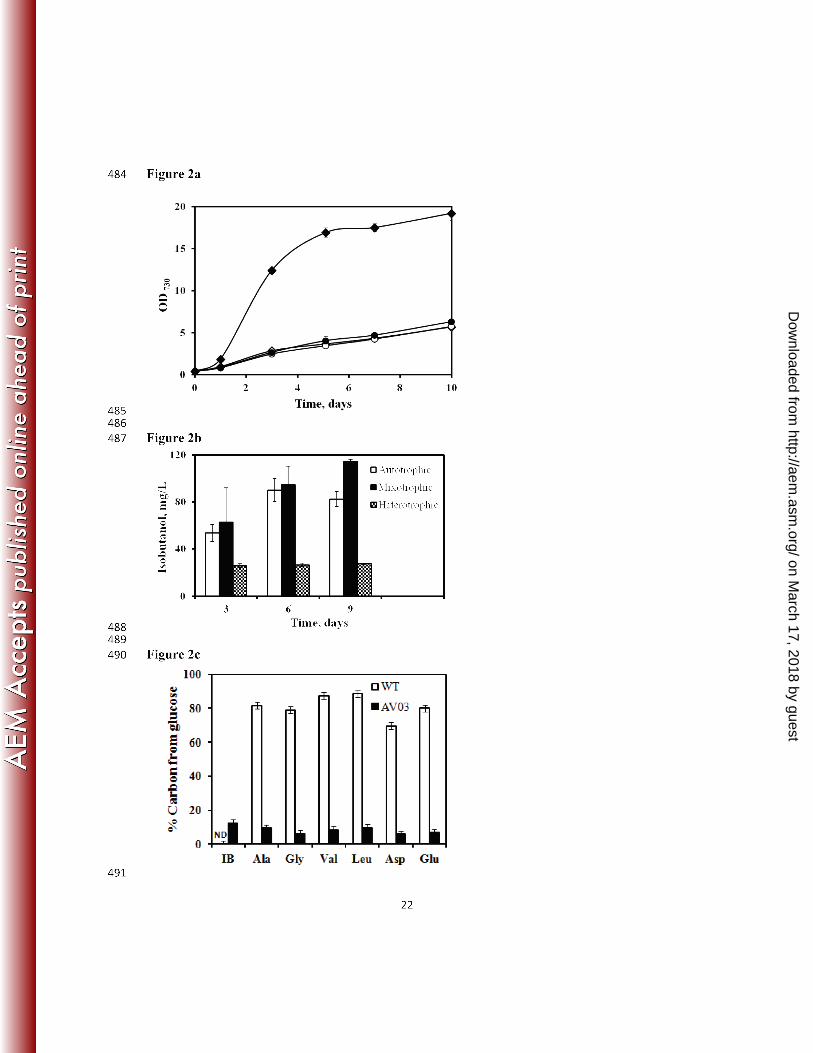
Isobutanol synthesis under autotrophic and mixotrophic growth. Under autotrophic 197
conditions, Synechocystis 6803 utilizes light as an energy source (ATP and NADPH) for the 198
conversion of CO2 into biomass and IB. Fig. 2a compares the autotrophic growth of the mutant 199
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

10
and the wild strain. Under autotrophic conditions, we found that the growth rate of the mutant 200
AV03 remains unaltered as compared to the wild-type strain. IB accumulation in the mutant was 201
tested under autotrophic condition (Fig. 2b), and the strain was found to synthesize a maximum 202
of 90 mg/L of IB (the only extracellular product detected by GC-MS) in a 6-day culture. In a 203
sealed shaking flask, NaHCO3 in the medium (50mM) was consumed by AV03 within six days, 204
and then both the biomass and IB started declining. 205
The wild-type strain of Synechocystis 6803 grows about 5 times faster under mixotrophic 206
conditions compared to autotrophic conditions (Fig. 2a). However, our mutant AV03 did not 207
exhibit an increased growth rate under mixotrophic conditions. To measure the glucose 208
utilization by wild-type and mutant AV03, we fed cells with 0.5% fully labeled glucose and 209
nonlabeled bicarbonate. Isotopomer analysis of 13C-abundance in cell metabolites (Fig. 2c) 210
showed that the wild-type synthesized 70~90% of its amino acids using carbons from glucose, 211
whereas the mutant produced biomass only using 5~10% carbon from glucose, and 12% of the 212
carbon of IB was labeled (i.e., derived from glucose). These results indicated that the mutant 213
tended to limit glucose metabolism for IB production. The AV03 strain was found to synthesize 214
a maximum of 114 mg/L of IB mixotrophically after 9 days (Fig. 2b), whereas cells with only 215
glucose (heterotrophic without bicarbonate or CO2) synthesized a maximum of 27 mg/L of IB. 216
This result suggests that the Synechocystis 6803 mutant is unable to take significant advantage of 217
its glucose metabolism to have a fast rate of IB production. 218
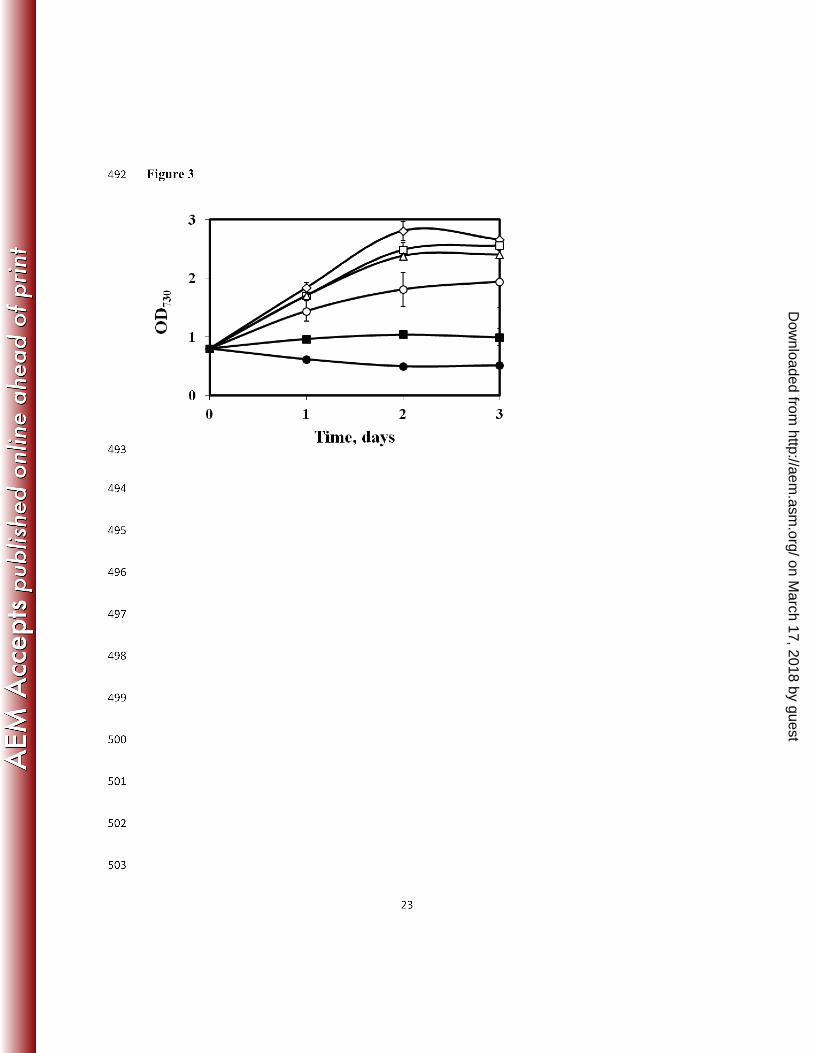
In situ alcohol concentrating system using a solvent trap. IB is toxic to the cells and our 219
study revealed that IB inhibited Synechocystis 6803 growth at external concentrations of only 2 220
g/L (Fig. 3). Moreover, our control experiments indicated the loss of IB after 9 days of 221
continuous incubation. IB can be slowly degraded by photo-chemically-produced hydroxyl 222
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

11
radicals in aerobic cyanobacterial cultures (6, 16, 31). Therefore, a system with continuous 223
removal of the synthesized alcohol products will be beneficial (19, 29). We have demonstrated 224
the use of an in situ alcohol removal system by using oleyl alcohol (12, 15) as a solvent trap for 225
increasing IB titer. In previous studies, gas stripping is one efficient way for IB recovery, but it 226
requires an expensive cooling system due to very low concentrations of IB from photo-227
bioreactors. Here, inside each cultivation flask, we placed a small glass vial containing 0.5 or 1 228
mL oleyl alcohol solvent, so that oleyl alcohol was not mixed with the culture solution (Fig. 4). 229
Volatile IB in the headspace can be trapped in the solvent vial because of the high solubility of 230
IB in oleyl alcohol. This method will effectively trap the IB while the solvent will not directly 231
interfere with light and cell culture conditions. 232
To test the effect of oleyl alcohol on IB productivity, we did a 3-week time-course study 233
(Fig. 4) by adding 50 mM NaHCO3 intermittently (every 4 days). During the cultivation, the pH 234
of the cultures was also adjusted to be between 8 and 9 before the addition of excess bicarbonate. 235
Every 3 days, the oleyl alcohol in the vials was taken out for IB measurement, and then replaced 236
with fresh oleyl alcohol. The mixotrophic cultures with alcohol trap (0.5 mL) reached the highest 237
net IB concentration of 298 mg/L. The autotrophic cultures with 0.5 mL and 1 mL oleyl alcohol 238
had a maximum net IB titer of 180 mg/L and 240 mg/L, respectively, whereas the autotrophic 239
cultures without the oleyl alcohol trap were able to achieve a maximum of only 108 mg/L of IB. 240
IB levels in the organic phase reached concentrations of up to 500 mg/L with only 3 days of 241
trapping. 242
Discussion 243
Isobutanol (IB) is a promising biofuel for the replacement of gasoline. So far, E. coli has 244
remained the most successful microbial host for IB production. In this study, we have focused 245
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

12
our efforts on a cyanobacterial species, Synechocystis 6803, which can grow on both CO2 and 246
glucose. The mixotrophic cultivation may offer industrial flexibility and economic benefits 247
because the gas-liquid mass transfer of CO2 is often a rate-limiting step in efficient 248
photobioreactor operations. Attempts in creating a stable strain of Synechococcus 7942 that can 249
transport and utilize glucose has been barely successful (39). The glucose tolerant strain of 250
Synechocystis 6803 unlike other cyanobacterial strains, can perform both autotrophic and 251
mixotrophic metabolisms. In our work, we found that the wild-type strain of Synechocystis 6803 252
under mixotrophic conditions grew at a rate 5 times faster than the autotrophic condition. 253
Moreover, the engineered Synechocystis 6803 strain accumulated 90 mg/L of IB, whereas the 254
Synechococcus 7942 strain expressing the same two enzymes (keto-acid decarboxylase and 255
alcohol dehydrogenase) only accumulated a maximum of 18 mg/L (2). Switching the condition 256
from autotrophic to mixotrophic for the mutant AV03 increased the maximum IB titer to 114 257
mg/L. Interestingly, the mutant tended to grow autotrophically and had minimal glucose 258
utilization compared to the wild-type strain (Fig 2c). 259
IB can be inhibitory to cell physiologies. Moreover, our experiments also observed IB 260
degradation (by hydroxyl radicals) during the incubation process. Thereby, efforts in coming up 261
with product recovery are important to improve IB titer in cyanobacterial culture. This work 262
employed an in situ IB removal system by growing cultures in shake flasks with vials containing 263
oleyl alcohol. Mixotrophic growth of AV03 along with in situ IB removal synthesized a 264
maximum of 298 mg/L IB, which is lower than the highest IB titer (450 mg/L) reported in 265
Synechococcus 7942 mutant expressing 3 more genes of the keto acid pathway. On the other 266
hand, our strain design has two apparent advantages for industrial application. First, our strain 267
does not require any antibiotics to maintain its IB production because the two heterologous genes 268
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

13
in the mutant show good stability during normal cultivation conditions. Second, the strain does 269
not need any inducer (IPTG) for IB production, which can significantly reduce the industrial 270
costs. 271
Overexpressing the keto acid pathway can increase the IB titer in Synechococcus 7942 272
(2). Furthermore, optimizing CO2 and light conditions of the cyanobacterial strain can also 273
increase the final titer and productivity. Liu et. al., (23) have reported a doubling time of 7.4 274
hours for Synechocystis 6803, by growing them under 140 µmol of photons m-2 s-1 of light and 275
by bubbling 1% CO2 enriched air. Therefore, our strain can serve as a springboard for future 276
development of higher performance Synechocystis 6803 strains with increased IB titer and 277
productivity. 278
In summary, IB synthesis under autotrophic conditions in a cyanobacterium 279
Synechocystis 6803 was demonstrated by the expression of two heterologous genes. It was 280
further demonstrated that mixotrophic cultures of the mutant can significantly increase IB 281
synthesis with minimal glucose consumption. The mechanism behind the reduced glucose-282
utilizing metabolism of AV03 compared to the wild-type strain remains unclear. A possible 283
explanation is that the cells tend to avoid the intracellular metabolic imbalance or IB 284
intermediate inhibition by down-regulating glucose uptake. Using oleyl alcohol as a simple 285
solvent trap, IB production can be improved by 2~3 times. Therefore, in situ IB recovery may 286
reduce the product loss and separation cost. We have also demonstrated that a simple expression 287
of the Ehrlich pathway with bioprocess modification can synthesize IB without other major 288
waste products, while still achieving comparable levels of IB to an extensive genetically 289
modified Synechococcus 7942 strain (Table 1) (2). 290
291
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

14
Acknowledgment 292
The authors wish to thank Dr. Abhay Singh and Dr. Anindita Bandyopadhyay for their 293
technical guidance. The authors would also like to thank Amelia Nguyen and Amelia Chen for 294
their assistance with data collection for a few experiments. This research was funded by an NSF 295
Career Grant to YT (MCB0954016), a grant to HBP from the Office of Science (BER), U.S. 296
Department of Energy, and a grant to HBP from the Consortium for Clean Coal Utilization at 297
Washington University. We would also like to thank Seema Mukhi Dahlheimer from the 298
Engineering Communication Center of Washington University in St. Louis for her close reading 299
of this manuscript. 300
Authors’ contributions 301
YJT initiated this study. HP, YJT and AV designed the experiments. AV and YX performed the 302
experiments. AV wrote the manuscript. All authors revised and approved the final manuscript. 303
References 304
1. Atsumi, S., T. Hanai, and J. C. Liao. 2008. Non-fermentative pathways for synthesis of 305 branched-chain higher alcohols as biofuels. Nature 451:86-9. 306
2. Atsumi, S., W. Higashide, and J. C. Liao. 2009. Direct photosynthetic recycling of 307 carbon dioxide to isobutyraldehyde. Nature Biotechnology 27:1177-1180. 308
3. Baez, A., K.-M. Cho, and J. C. Liao. 2011. High-flux isobutanol production using 309 engineered Escherichia coli: a bioreactor study with in situ product removal. Applied 310 Microbiology and Biotechnology 90:1681-1690. 311
4. Barnes, W. M. 1994. PCR amplification of up to 35 kb DNA with high fidelity and high 312 yield from bacteriophage templates. Proceedings of the National Academy of Sciences of 313 the United States of America 91:2216-2220. 314
5. Bond-Watts, B. B., R. J. Bellerose, and M. C. Chang. 2011. Enzyme mechanism as a 315 kinetic control element for designing synthetic biofuel pathways. Nat Chem Biol 7:222-7. 316
6. Cameron, J. C., and H. B. Pakrasi. 2011. Glutathione Facilitates Antibiotic Resistance 317 and Photosystem I Stability during Exposure to Gentamicin in Cyanobacteria. Applied 318 and Environmental Microbiology 77:3547-3550. 319
7. Colletti, P., Y. Goyal, A. Varman, x. Feng, B. Wu, and Y. Tang. 2011. Evaluating 320 factors that influence microbial synthesis yields by linear regression with numerical and 321 ordinal variables. Biotechnology and Bioengineering 108:893-901. 322
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

15
8. Deng, M. D., and J. R. Coleman. 1999. Ethanol synthesis by genetic engineering in 323 cyanobacteria. Applied and Environmental Microbiology 65:523-528. 324
9. Dexter, J., and P. Fu. 2009. Metabolic engineering of cyanobacteria for ethanol 325 production. Energy & Environmental Science 2:857-864. 326
10. Ducat, D. C., G. Sachdeva, and P. A. Silver. 2011. Rewiring hydrogenase-dependent 327 redox circuits in cyanobacteria. Proceedings of the National Academy of Sciences 328 108:3941-3946. 329
11. Ducat, D. C., J. C. Way, and P. A. Silver. 2011. Engineering cyanobacteria to generate 330 high-value products. Trends in Biotechnology 29:95-103. 331
12. Ezeji, T., C. Milne, N. D. Price, and H. P. Blaschek. 2010. Achievements and 332 perspectives to overcome the poor solvent resistance in acetone and butanol-producing 333 microorganisms. Applied Microbiology and Biotechnology 85:1697-1712. 334
13. Feng, X., A. Bandyopadhyay, B. Berla, L. Page, B. Wu, H. B. Pakrasi, and Y. J. 335 Tang. 2010. Mixotrophic and photoheterotrophic metabolism in Cyanothece sp. ATCC 336 51142 under continuous light. Microbiology 156:2566-74. 337
14. Gao, Q., W. Wang, H. Zhao, and X. Lu. 2012. Effects of fatty acid activation on 338 photosynthetic production of fatty acid-based biofuels in Synechocystis sp. PCC 6803. 339 Biotechnology for Biofuels 5. 340
15. Green, E. M. 2011. Fermentative production of butanol - the industrial perspective. 341 Current Opinion in Biotechnology 22:337-343. 342
16. Grosjean, D. 1997. Atmospheric chemistry of alcohols. Journal of the Brazilian 343 Chemical Society 8:433-442. 344
17. Huang, H.-H., D. Camsund, P. Lindblad, and T. Heidorn. 2010. Design and 345 characterization of molecular tools for a Synthetic Biology approach towards developing 346 cyanobacterial biotechnology. Nucleic acids research 38:2577-2593. 347
18. Keasling, J. D., and H. Chou. 2008. Metabolic engineering delivers next-generation 348 biofuels. Nat Biotech 26:298-299. 349
19. Kraemer, K., A. Harwardt, R. Bronneberg, and W. Marquardt. 2010. Separation of 350 butanol from acetone-butanol-ethanol fermentation by a hybrid extraction-distillation 351 process. Computers & Chemical Engineering 35:949-963. 352
20. Lamsen, E. N., and S. Atsumi. 2012. Recent progress in synthetic biology for microbial 353 production of C3-C10 alcohols. Frontiers in Microbiology 3. 354
21. Lan, E. I., and J. C. Liao. 2012. ATP drives direct photosynthetic production of 1-355 butanol in cyanobacteria. Proceedings of the National Academy of Sciences of the United 356 States of America 109:6018-6023. 357
22. Lan, E. I., and J. C. Liao. 2011. Metabolic engineering of cyanobacteria for 1-butanol 358 production from carbon dioxide. Metabolic Engineering 13:353-363. 359
23. Liu, X. Y., J. Sheng, and R. Curtiss. 2011. Fatty acid production in genetically 360 modified cyanobacteria. Proceedings of the National Academy of Sciences of the United 361 States of America 108:6899-6904. 362
24. Mohamed, A., J. Eriksson, H. D. Osiewacz, and C. Jansson. 1993. Differential 363 expression of the psbA genes in the cyanobacterium Synechocystis 6803. Molecular & 364 General Genetics 238:161-168. 365
25. Mohamed, A., and C. Jansson. 1989. Influence of light on accumulation of 366 photosynthesis-specific transcripts in the cyanobacterium Synechocystis 6803. Plant 367 Molecular Biology 13:693-700. 368
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

16
26. Okamoto, S., M. Ikeuchi, and M. Ohmori. 1999. Experimental analysis of recently 369 transposed insertion sequences in the cyanobacterium Synechocystis sp. PCC 6803. DNA 370 research : an international journal for rapid publication of reports on genes and genomes 371 6:265-73. 372
27. Rittmann, B. E. 2008. Opportunities for renewable bioenergy using microorganisms. 373 Biotechnology and Bioengineering 100:203-212. 374
28. Sentheshanmuganathan, S., and S. R. Elsden. 1958. The mechanism of the formation 375 of tyrosol by Saccharomyces cerevisiae. The Biochemical journal 69:210-8. 376
29. Shi, Z. P., C. Y. Zhang, J. X. Chen, and Z. G. Mao. 2005. Performance evaluation of 377 acetone-butanol continuous flash extractive fermentation process. Bioprocess and 378 Biosystems Engineering 27:175-183. 379
30. Tan, X., L. Yao, Q. Gao, W. Wang, F. Qi, and X. Lu. 2011. Photosynthesis driven 380 conversion of carbon dioxide to fatty alcohols and hydrocarbons in cyanobacteria. 381 Metabolic Engineering 13:169-176. 382
31. Tichy, M., and W. Vermaas. 1999. In Vivo Role of Catalase-Peroxidase 383 inSynechocystis sp. Strain PCC 6803. Journal of Bacteriology 181:1875-1882. 384
32. van Groenigen, K. J., C. W. Osenberg, and B. A. Hungate. 2011. Increased soil 385 emissions of potent greenhouse gases under increased atmospheric CO2. Nature 475:214-386 216. 387
33. Varman, A., Y. Xiao, E. Leonard, and Y. Tang. 2011. Statistics-based model for 388 prediction of chemical biosynthesis yield from Saccharomyces cerevisiae. Microbial Cell 389 Factories 10:45. 390
34. Vermaas, W. 1996. Molecular genetics of the cyanobacterium Synechocystis sp. PCC 391 6803: Principles and possible biotechnology applications. Journal of Applied Phycology 392 8:263-273. 393
35. Wahl, S. A., M. Dauner, and W. Wiechert. 2004. New tools for mass isotopomer data 394 evaluation in (13)C flux analysis: mass isotope correction, data consistency checking, and 395 precursor relationships. Biotechnol Bioeng 85:259-68. 396
36. Xiao, Y., X. Feng, A. M. Varman, L. He, H. Yu, and Y. J. Tang. 2012. Kinetic 397 modeling and isotopic investigation of isobutanol fermentation by two engineered 398 Escherichia coli strains. Industrial & Engineering Chemistry Research Under Review. 399
37. You, L., L. Page, X. Feng, B. Berla, H. B. Pakrasi, and Y. J. Tang. 2012. Metabolic 400 Pathway Confirmation and Discovery Through 13C-labeling of Proteinogenic Amino 401 Acids. J Vis Exp:e3583. 402
38. Zang, X. N., B. Liu, S. M. Liu, K. Arunakumara, and X. C. Zhang. 2007. Optimum 403 conditions for transformation of Synechocystis sp. PCC 6803. Journal of Microbiology 404 45:241-245. 405
39. Zhang, C. C., R. Jeanjean, and F. Joset. 1998. Obligate phototrophy in cyanobacteria: 406 more than a lack of sugar transport. Fems Microbiology Letters 161:285-292. 407
40. Zhou, J., and Y. Li. 2010. Engineering cyanobacteria for fuels and chemicals 408 production. Protein & Cell 1:207-210. 409
410 411 412 413 414
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

17
Figure Legend 415 416 Figure 1: (a) Schematic representation to show the integration of the genes kivd and adhA into 417
the genome of Synechocystis 6803. Colony PCR performed to verify the integration of the insert 418
into the genomic DNA of the mutant (AV03). The vector ptka3 was used as a template for the 419
positive control and wild-type cells were used as negative control. Colony PCR of AV03, 420
showed the presence of a band (8.3kb) the same size as the positive control (+ve) and the 421
absence of the negative control (WT) band. (b) IB synthesized by engineered Synechocystis 6803 422
under different IPTG concentrations (n=3). (Inset) Result of an RT-PCR performed to detect the 423
expression of the heterologous genes kivd (Top: 500bp from kivd) and adhA (Bottom: 200bp 424
from adhA). Lane 1, wild-type 6803 (WT); Lane 2, AV03 with 0 mM IPTG; Lane 3, AV03 425
with 0.5mM IPTG; Lane 4, AV03 with 1mM IPTG. 426
427
Figure 2: (a) Growth curves of Synechocystis 6803 WT and AV03 (n=3, shake flask cultures): 428
WT under autotrophic, ♦ WT under mixotrophic, ○ AV03 under autotrophic and ● AV03 under 429
mixotrophic conditions (note: growth curve of AV03 under mixotrophic condition overlaps with 430
autotrophic growth curves of AV03 and WT). (b) IB synthesized in AV03 under autotrophic 431
conditions (only HCO3), heterotrophic (only glucose) and mixotrophic (both HCO3 and glucose) 432
conditions (n=3, shake flask cultures with closed caps). (c) Percentage carbon contribution of 433
glucose for synthesizing amino acids and isobutanol in the wild-type (WT) and the mutant strain 434
(AV03) as measured on day 9 (shake flask cultures with closed caps). Isotopomer analysis 435
(TBDMS based method) of proteinogenic amino acids confirms the low 13C-glucose utilization 436
by the mutant. The error bar represents the 2% technical error of the instrument. 437
438
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

18
Figure 3: Toxic effects of IB on the growth of Synechocystis 6803. IB was added to a final 439
concentration (g/L, n=2) of ◊ 0, □ 0.2, Δ 0.5, ○ 1, ■ 2 and ● 5 to a Synechocystis 6803 culture 440
with an initial OD730 ~ 0.8. 441
Figure 4: Net concentration of IB synthesized (columns) and biomass growth (curves) by the 442
AV03 culture under different conditions (n=3). a – IB with 0.5 mL oleyl alcohol (Autotrophic); b 443
– IB with 1 mL oleyl alcohol (Autotrophic); c – IB with 0.5 mL oleyl alcohol and glucose 444
(Mixotrophic); d – IB with no oleyl alcohol (Autotrophic, negative control); a1 – OD730 with 0.5 445
mL oleyl alcohol (Autotrophic); a2 – OD730 with 1 mL oleyl alcohol (Autotrophic); a3 – OD730 446
with 0.5 mL oleyl alcohol and glucose (Mixotrophic); a4 – OD730 with no oleyl alcohol 447
(Autotrophic, negative control). (Inset) Schematic representation of the in situ IB removal 448
system used to increase the production of IB. 449
450
451
452
453
454
455
456
457
458
459
460
461
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from
19
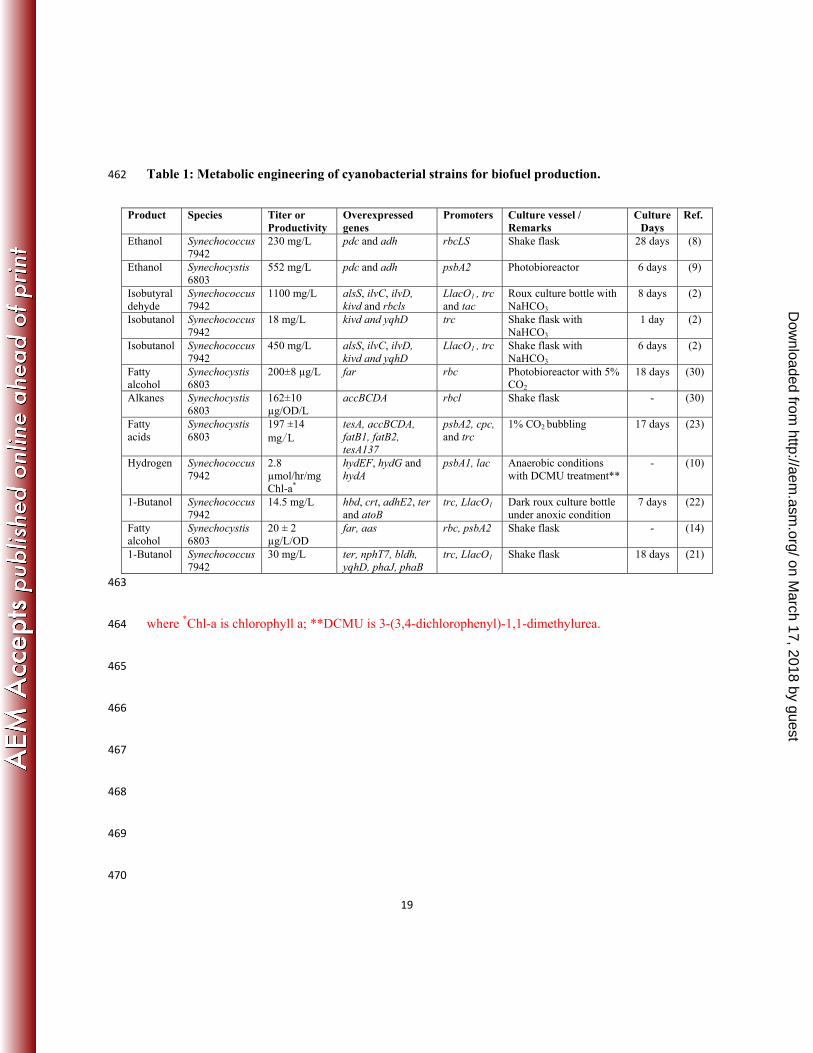
Table 1: Metabolic engineering of cyanobacterial strains for biofuel production. 462
Product Species Titer or Productivity
Overexpressed genes
Promoters Culture vessel / Remarks
Culture Days
Ref.
Ethanol Synechococcus 7942
230 mg/L pdc and adh
rbcLS Shake flask 28 days (8)
Ethanol Synechocystis 6803
552 mg/L pdc and adh
psbA2 Photobioreactor 6 days (9)
Isobutyraldehyde
Synechococcus 7942
1100 mg/L alsS, ilvC, ilvD, kivd and rbcls
LlacO1 , trc and tac
Roux culture bottle with NaHCO3
8 days (2)
Isobutanol Synechococcus 7942
18 mg/L kivd and yqhD
trc Shake flask with NaHCO3
1 day (2)
Isobutanol Synechococcus 7942
450 mg/L alsS, ilvC, ilvD, kivd and yqhD
LlacO1 , trc Shake flask with NaHCO3
6 days (2)
Fatty alcohol
Synechocystis 6803
200±8 µg/L far rbc Photobioreactor with 5% CO2
18 days (30)
Alkanes Synechocystis 6803
162±10 µg/OD/L
accBCDA rbcl Shake flask - (30)
Fatty acids
Synechocystis 6803
197 ±14 mg∕L
tesA, accBCDA, fatB1, fatB2, tesA137
psbA2, cpc, and trc
1% CO2 bubbling 17 days (23)
Hydrogen Synechococcus 7942
2.8 µmol/hr/mg Chl-a*
hydEF, hydG and hydA
psbA1, lac Anaerobic conditions with DCMU treatment**
- (10)
1-Butanol Synechococcus 7942
14.5 mg/L hbd, crt, adhE2, ter and atoB
trc, LlacO1 Dark roux culture bottle under anoxic condition
7 days (22)
Fatty alcohol
Synechocystis 6803
20 ± 2 µg/L/OD
far, aas rbc, psbA2 Shake flask - (14)
1-Butanol Synechococcus 7942
30 mg/L ter, nphT7, bldh, yqhD, phaJ, phaB
trc, LlacO1 Shake flask 18 days (21)
463
where *Chl-a is chlorophyll a; **DCMU is 3-(3,4-dichlorophenyl)-1,1-dimethylurea. 464
465
466
467
468
469
470
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from

20
Table 2: Primer sequences used in this study. 471
Name Sequence (5’→3’)
AMV14F GCGCACATGTCGGAACAGGACCAAGCCTTGAT
AMV15R GCGC CCTGAGGCCTTTACCATGACCTGCAGGG
AMV16F GCGCGACGGGGAGTCAATTGTGCCATTGCCATAACTGCTTTCG
AMV17R GCGCGACTCCCCGTCTTTGACTATCCTTTTTAGGATGGGGCA
ps1_up_fwd TACCGGAACAGGACCAAGCCTT
AMV01 GCGCCATATGTATACAGTAGGAGATTACCTATTAGAC
AMV12 GCAGCAGCAACATCAACTGGTAAG
AdhA-TMs TCAACTAGTGGTACCAGGAGATATAATATGAAAGCAGCAGTAGTAAG
adhA_RTr GACAATTCCAATTCCTTCATGACCAAG
Rnpbr CGGTATTTTTCTGTGGCACTGTCC
Rnpbf CAGCGGCCTATGGCTCTAATC
AV03_6F GAATCCGTAATCATGGTCATAGCTG
AV03_4R GCCAAAGCTAATTATTTCATGTCCTGT
AV03_1R TGTCGGGGCGCAGCCATGA
AV03_2F AGAGGATCCTTCTGAAATGAGCTG
AV03_3F CAGAGCCTAATCTTAAAGAATTCGTGG
AV03_4F GGGTAAACTATTTGCTGAACAAAATAAATC
AV03_2R CCGCTTCTGCGTTCTGATTTAATC
AV03_5F GTTGATCGGCGCGAGATTTAATCG
AV03_7F CCGTTGAAATTGACCGAGTACTTTCT
AV03_8F CAGTCGAAAGAGAAATTCATGGACC
AV03_5R CGCTACGGCGTTTCACTTCTG
472
473
474
475
on March 17, 2018 by guest
http://aem.asm
.org/D
ownloaded from








![Synechocystis sp. PCC 6803 in a bio-photoelectrolysis cell ... · 4-resu e efficienc ncentration ated. )6] 4-. Applic ells) did no lted in an i ies of H2 s and tem ation of a t produce](https://img.dokumen.tips/doc/110x75/5f80e05ab8e29d00bc4e36a8/synechocystis-sp-pcc-6803-in-a-bio-photoelectrolysis-cell-4-resu-e-efficienc.jpg)











![Benda Asing Di Esofagus1 [6803]](https://img.dokumen.tips/doc/110x75/577c77fa1a28abe0548e3655/benda-asing-di-esofagus1-6803.jpg)
![Dnevni avaz [broj 6803, 20.7.2014]](https://img.dokumen.tips/doc/110x75/577cc7011a28aba7119fbeb2/dnevni-avaz-broj-6803-2072014.jpg)








