Embed Size (px)
Citation preview

Mass Spectrometry-based Peptidomics for the Discovery of Antimicrobial Peptides
Tsukasa Osaki*, Kazuki Sasaki and Naoto Minamino
Department of Molecular Pharmacology, National Cerebral and Cardiovascular Center Research Institute, 5-7-1 Fujishirodai, Suita, Osaka 565-8565, Japan
Most antimicrobial peptides (AMPs) are cationic molecules with a high content of basic amino acids. They often undergo C-terminal amidation that may be related to their antimicrobial activity. Here we describe a mass spectromety (MS)-based approach for identifying novel AMPs. MS is used to profile peptides secreted by endocrine cells. Peptide sequences characteristic of AMPs are selected to test their antimicrobial activity. Our ‘sequence first approach’ has revealed two novel AMPs whose activity against certain microbes are comparable to that of cathelicidin (CATH) or β-defensin-2 (BD2). The antimicrobial activity and natural occurrence of the peptides, designated antimicrobial peptide derived from insulin-like growth factor-binding protein-5 (AMP-IBP5) and VGF [554-577]-NH2, are described in this review.
Keywords antimicrobial activity; bioactive peptides; insulin-like growth factor-binding protein 5; mass spectrometry; neurosecretory protein VGF; peptidomics.
1. MS-based AMP discovery
For applications to humans, it is important to discover endogenous AMPs from mammalians including humans. In addition, it is necessary to discover AMPs with physiological significances. Degradation products of common proteins such as hemoglobins have been described as AMPs [1]; however, their physiological implications remain unclear. It is thus important to discover physiologically relevant AMPs when considering their application to humans. We therefore identified endogenous peptides generated by specific processing in physiolgical conditions and selected AMP candidates from the identified peptides, considering chemical properties of AMPs, e.g. high contents of basic amino acids and C-terminal amidation.
1.1. MS profiling of secretory peptides for AMP discovery
Advances in MS, accompanied by an increase in sequenced genomes, have enabled a comprehensive study of naturally occurring peptides [2, 3]. The MS method for identifying endogenous peptides is tandem mass spectrometry (MS/MS), where each peptide is dissociated to a sequence ladder of fragment ions in a mass spectrometer. Basically, MS identification procedures are so affected by sample heterogeneity that sample preparation needs to be optimized for each specific application. To identify bioactive peptides efficiently, we analyzed peptides released by endocrine cells that are stimulated to induce exocytosis [4]. Released peptides were harvested for MS. This protocol has proved effective, as we found that more than 95% of the peptides identified from endocrine cell lines originated from precursor proteins localized in secretory granules [4, 5]. Since endocrine cells secrete AMPs through proteolytic processing of granin precursor proteins, e.g. chromofungin processed from chromogranin A [6], we expected that in-depth MS profiling of secretory peptides might uncover novel AMPs that have escaped conventional identification strategies using either activity-guided biochemical purification or homology-based cDNA cloning.
1.2. Selecting AMP candidates with reference to sequence features
In our “sequence first” approach, a number of peptide sequences are revealed by liquid chromatography (LC)-MS/MS analysis. Bioactivity of a peptide is identified only after it is tested with an assay system. Since it is practically impossible to synthesize all the identified peptides, we should select peptides for assessing antimicrobial activity. To select candidate AMPs, we have considered interspecies homology, basic amino acid contents and posttranslational modifications associated with known AMPs. Interspecies homology is especially critical because sequences involved in biological activity are highly conserved. Second, the occurrence of multiple basic amino acids is common in most AMPs. For instance, BD2 and CATH, well-characterized mammalian AMPs, both bear a net charge of +6 at pH 7. The overall positive charge in physiological condition is important for AMPs to adhere to negatively charged membranes of pathogens and to disturb lipids to alter the membrane structure [7]. Third, we also have taken into account post-translational modifications, especially C-terminal amidation. Some AMPs have the C-terminal amide group that plays a significant role in their antimicrobial property [8, 9]. Mechanistically, it is known that amidated C-termini, rather than common carboxy termini, help to enhance static interactions between the peptides and target bacterial membranes [8]. Based on these characteristics observed in many AMPs, we selected a total of 18 candidate peptides to assess antimicrobial activity.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1145

1.3. Antimicrobial activity screening
We measured antimicrobial activity of the 18 candidates against the Gram-positive Staphylococcus aureus (S. aureus) 209P and Gram-negative Escherichia coli (E. coli) K12 and fungus Pichia pastoris (P. pastoris) GS115 using a metabolic indicator alamarBlue. Most peptides even at 10 μM did not have antimicrobial activity with a survival rate of over 90% against these microbes tested (data not shown). We found that two of the 18 peptides were antimicrobial with a survival rate of under 50%. One was derived from IGFBP-5 and the other derived from the neurosecretory protein VGF (non-acronymic), neither of which has previously been described. The former, IGFBP-5 [193-214]-NH2 was designated AMP-IBP5, because of its potent antimicrobial activity. The latter was VGF [554-577]-NH2; VGF is now established as a bioactive peptide precursor. These two novel AMPs are described in the following sections.
2. AMP-IBP5
2.1. Introduction
AMP-IBP5 is a 22-amino-acid C-terminally amidated peptide with an intramolecular disulfide linkage [10]. At the amino acid sequence level, the peptide is identical among placental mammals and highly conserved between placental and non-placental mammals (Fig. 1). The peptide is highly rich in basic amino acids, which confers a net charge of +7 at pH 7. AMP-IBP5 is named after its antimicrobial activity. It is comparable to CATH and BD-2 with regard to antimicrobial spectrum and potency. These points are detailed in the following sections.
Fig. 1 Alignment of deduced mature amino acid sequences and the N- and C-terminal flanking sequences of AMP-IBP5. Conserved amino acids are shown with large font. C-terminally amidated residues are underlined. Flanking sequences are in italic and shaded boxes. Heparin-binding motif (YKRKQCKP) is boxed. Bold letters represent basic amino acids. Double underlined cysteines are disulfide-linked.
2.2. Discovery
The peptide was originally identified in our effort to analyze cationic peptides. Fig. 2 illustrates the scheme for peptide identification. First, a human pancreatic islet cell line was stimulated for exocytosis to recover supernatant (secretopeptidome). A pool of basic peptides, eluted over a high ammonium formate concentration of 0.9 M, pH 3.8, from a cation-exchange column, was isolated. The basic peptide-rich pool was subjected to LC-matrix-assisted laser desorption ionization (MALDI) MS analysis. This peptidomic survey has led to the identification of AMP-IBP5. It should be noted that no specific chemical or biochemical selection was conducted to identify these C-terminally amidated peptides, although we used endocrine cell lines secreting known C-terminally amidated peptide hormones.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1146

Fig. 2 Peptide identification scheme. A peptidomic survey of cationic peptide fractions resulted in the identification of a new AMP designated AMP-IBP5.
2.3. Naturally occurring form
Natural occurrence of the peptide was examined in rat tissues, since it was originally identified in a cell line. We prepared antibody and characterized immunoreactive (IR)-peptide(s) in rat brain and small intestine extracts using chromatography and MS. In gel filtration chromatography, IR-AMP-IBP5 in both extracts occurred as a distinct peak in the region of a molecular mass less than 4 kDa and separated into two major IR-peaks by reverse-phase high performance liquid chromatography (HPLC). Judged by retention time, the earlier eluted peak corresponded to a synthetic 22-amino-acid AMP-IBP5. This HPLC fraction was immunoprecipitated with a specific antibody for MS analysis, which detected a distinctive peak at m/z 2654.4 ([M+H]+ ion), the value consistent with synthetic AMP-IBP5 being a disulfide-linked peptide. These data indicate that AMP-IBP5 occurs in normal rat tissues in the molecular form identical to that found in the human cell conditioned medium. Assessed with a radioimmunoassay (RIA) system using the same antibody, IR-AMP-IBP5 in the rat brain, pituitary gland and small intestine was 2.1, 6.2 and 1.5 pmol/g wet tissues, respectively. In other tissues such as heart, it was below a detection limit for quantitative measurement (0.6 pmol/g wet tissue) [10]. On the other hand, intact IGFBP-5 protein in the brain, pituitary gland, heart and small intestine was 18.5, 15.9, 30.9 and 5.5 nmol/g wet tissues, respectively [10]. These data suggest tissue-to-tissue heterogeneity in IGFBP-5 processing; the precursor appears to be processed in the pituitary to a greater extent than in the heart.
2.4. Peptide processing sites
The peptide is C-terminally flanked by a canonical C-terminal amidation motif commonly found in peptide hormones and neuropeptides. The sequence 214RGRKRG219 fits the consensus motif ((R/K)-(X)n-(R/K)↓, where n = 0, 2, 4, or 6 and X is any amino acid other than Cys [11]) for prohormone convertase (PC) 1/3 and 2, major endoproteolytic proteases responsible for bioactive peptide processing. The precursor IGFBP-5 is therefore supposed to be first endoproteolytically cleaved C-terminal to 218R by PC1/3 or PC2, followed by carboxypeptidase E-mediated removal of 216RKR218. The upstream glycine (Fig. 1, underlined) is targeted by peptidyl-glycine alpha-amidating monooxygenase (PAM) [12] to generate the arginine amide structure. These processing enzymes were all expressed by the cell line we used (data not shown). In contrast, the N-terminal cleavage site of AMP-IBP5 (189MVPR↓A193) is an atypical monobasic site, which does not agree with the PC1/3 or PC2 consensus motif. However, some monobasic sites have been described to be recognized by PC1/3 or PC2 [13]. For instance, PC2 is involved in N-terminal processing of neuromedin C, MYPR↓G [13]. Since AMP-IBP5 is similar to neuromedin C in the N-terminal cleavage site, PC2 may also participate in the processing. Overall we suggest that AMP-IBP5 is generated by specific processing proteases involved in bioactive peptide processing.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1147
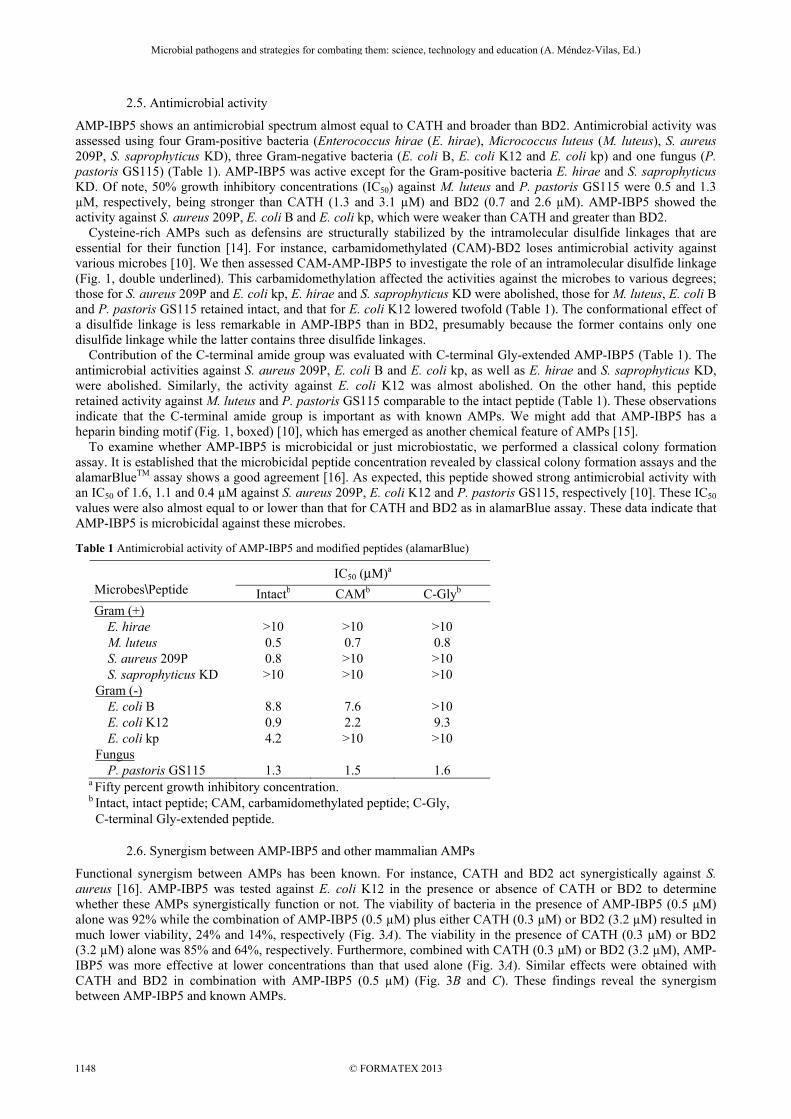
2.5. Antimicrobial activity
AMP-IBP5 shows an antimicrobial spectrum almost equal to CATH and broader than BD2. Antimicrobial activity was assessed using four Gram-positive bacteria (Enterococcus hirae (E. hirae), Micrococcus luteus (M. luteus), S. aureus 209P, S. saprophyticus KD), three Gram-negative bacteria (E. coli B, E. coli K12 and E. coli kp) and one fungus (P. pastoris GS115) (Table 1). AMP-IBP5 was active except for the Gram-positive bacteria E. hirae and S. saprophyticus KD. Of note, 50% growth inhibitory concentrations (IC50) against M. luteus and P. pastoris GS115 were 0.5 and 1.3 µM, respectively, being stronger than CATH (1.3 and 3.1 µM) and BD2 (0.7 and 2.6 µM). AMP-IBP5 showed the activity against S. aureus 209P, E. coli B and E. coli kp, which were weaker than CATH and greater than BD2. Cysteine-rich AMPs such as defensins are structurally stabilized by the intramolecular disulfide linkages that are essential for their function [14]. For instance, carbamidomethylated (CAM)-BD2 loses antimicrobial activity against various microbes [10]. We then assessed CAM-AMP-IBP5 to investigate the role of an intramolecular disulfide linkage (Fig. 1, double underlined). This carbamidomethylation affected the activities against the microbes to various degrees; those for S. aureus 209P and E. coli kp, E. hirae and S. saprophyticus KD were abolished, those for M. luteus, E. coli B and P. pastoris GS115 retained intact, and that for E. coli K12 lowered twofold (Table 1). The conformational effect of a disulfide linkage is less remarkable in AMP-IBP5 than in BD2, presumably because the former contains only one disulfide linkage while the latter contains three disulfide linkages. Contribution of the C-terminal amide group was evaluated with C-terminal Gly-extended AMP-IBP5 (Table 1). The antimicrobial activities against S. aureus 209P, E. coli B and E. coli kp, as well as E. hirae and S. saprophyticus KD, were abolished. Similarly, the activity against E. coli K12 was almost abolished. On the other hand, this peptide retained activity against M. luteus and P. pastoris GS115 comparable to the intact peptide (Table 1). These observations indicate that the C-terminal amide group is important as with known AMPs. We might add that AMP-IBP5 has a heparin binding motif (Fig. 1, boxed) [10], which has emerged as another chemical feature of AMPs [15]. To examine whether AMP-IBP5 is microbicidal or just microbiostatic, we performed a classical colony formation assay. It is established that the microbicidal peptide concentration revealed by classical colony formation assays and the alamarBlueTM assay shows a good agreement [16]. As expected, this peptide showed strong antimicrobial activity with an IC50 of 1.6, 1.1 and 0.4 µM against S. aureus 209P, E. coli K12 and P. pastoris GS115, respectively [10]. These IC50 values were also almost equal to or lower than that for CATH and BD2 as in alamarBlue assay. These data indicate that AMP-IBP5 is microbicidal against these microbes.
Table 1 Antimicrobial activity of AMP-IBP5 and modified peptides (alamarBlue)
Microbes\Peptide
IC50 (μM)a
Intactb CAMb C-Glyb Gram (+)
E. hirae >10 >10 >10 M. luteus 0.5 0.7 0.8 S. aureus 209P 0.8 >10 >10 S. saprophyticus KD >10 >10 >10
Gram (-) E. coli B 8.8 7.6 >10 E. coli K12 0.9 2.2 9.3 E. coli kp 4.2 >10 >10
Fungus P. pastoris GS115 1.3 1.5 1.6
a Fifty percent growth inhibitory concentration. b Intact, intact peptide; CAM, carbamidomethylated peptide; C-Gly, C-terminal Gly-extended peptide.
2.6. Synergism between AMP-IBP5 and other mammalian AMPs
Functional synergism between AMPs has been known. For instance, CATH and BD2 act synergistically against S. aureus [16]. AMP-IBP5 was tested against E. coli K12 in the presence or absence of CATH or BD2 to determine whether these AMPs synergistically function or not. The viability of bacteria in the presence of AMP-IBP5 (0.5 µM) alone was 92% while the combination of AMP-IBP5 (0.5 µM) plus either CATH (0.3 µM) or BD2 (3.2 µM) resulted in much lower viability, 24% and 14%, respectively (Fig. 3A). The viability in the presence of CATH (0.3 µM) or BD2 (3.2 µM) alone was 85% and 64%, respectively. Furthermore, combined with CATH (0.3 µM) or BD2 (3.2 µM), AMP-IBP5 was more effective at lower concentrations than that used alone (Fig. 3A). Similar effects were obtained with CATH and BD2 in combination with AMP-IBP5 (0.5 µM) (Fig. 3B and C). These findings reveal the synergism between AMP-IBP5 and known AMPs.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1148

Endogenous AMPs are a key regulator in innate immunity that provides a rapid response against invading pathogens [17-19]. In terms of antimicrobial spectrum and potency, AMP-IBP5 is comparable to potent AMPs CATH and BD2; AMP-IBP5 was effective against six microbes including E. coli K12 in the eight organisms tested. As described above, AMP-IBP5 was abundant in the brain and intestine, where many AMPs including CATH and BD2 have also been described [20-21]. Taken together, these findings suggest that AMP-IBP5 functions in cooperation with other AMPs in these tissues.
Fig. 3 Synergistic antimicrobial activity of AMP-IBP5 with BD-2 or CATH against E. coli K12. A, Increasing concentrations of AMP-IBP5 (0.1-1.3 μM) in the absence (●) or presence of BD-2 (▲, 3.2 μM) or CATH (■, 0.3 μM) were mixed and incubated as in (A) with the bacterium and incubated for 1 h at 37°C. B, Increasing concentrations of BD-2 (0.8-5.8 μM) in the absence (●) or presence of AMP-IBP5 (▲, 0.5 μM) were mixed and incubated as in (A). C, Increasing concentrations of CATH (0.1-0.6 μM) in the absence (●) or presence of AMP-IBP5 (▲, 0.5 μM) were mixed. Data are mean ± standard deviation (SD) of three experiments.
3. VGF [554-577]-NH2
3.1. Introduction
VGF [554-577]-NH2 is a C-terminally amidated peptide derived from the neurosecretory protein VGF [22]. VGF, selectively expressed in neurons and endocrine cells, has been known to be a precursor to multiple bioactive peptides. This peptide, however, is the only AMP arising from the precursor. The peptide sequence is highly conserved among placental mammals, including their N- and C-terminal processing sites (Fig. 4). This peptide contains four arginines, three of which are contiguous, bearing a net charge of +5.
Fig. 4 Alignment of deduced mature amino acid sequences and the N- and C-terminal flanking sequences of VGF [554-577]-NH2. Conserved amino acids are shown with large font. C-terminally amidated residues are underlined. Flanking sequences are in italic and shaded boxes. Bold letters represent basic amino acids.
3.2. Discovery
The peptide was identified in our study to uncover peptides using electron transfer dissociation (ETD), a recently introduced MS/MS method. The most important aspect of MS/MS sequencing is that every peptide is dissociated to a degree sufficient enough to provide sequence information. Although collision-induced dissociation (CID) remains a standard method of choice, most bioactive peptides are, due to their internal basic residues, resistant to CID than typical tryptic peptides [23]. Actually, VGF [554-577]-NH2 has escaped identification by CID. In the proteomics community, the benefit of ETD in bottom-up proteomics has been widely acknowledged, but endogenous peptides remain largely unexplored, despite the general assumption that ETD generates a more complete series of fragment ions than CID [24-26]. Using CID and ETD, we conducted a large-scale identification of endogenous secretory peptides, ranging from 1000 to 15000 Da [22]. A total of 795 peptides were identified by CID and 569 peptides by ETD, with an overlap of 397. Although outperformed by CID in total peptide identifications, an appreciable number of peptides, including VGF [554-577]-NH2, were uncovered only by ETD. CID and ETD MS/MS spectra of VGF [554-577]-NH2 are shown in Fig. 5 (at the same charge). Compared with CID, ETD yielded consecutive fragment ions. The observed fragment ions produced by ETD well matched to theoretical ions across the entire sequence, with an expectation value of 7.2 x 10-8, which was calculated by MS/MS search software
0
20
40
60
80
100
0 2 4 6
%V
iabi
lity
BD-2 (μM)
0
20
40
60
80
100
0 0.2 0.4 0.6 0.8
%V
iabi
lity
CATH (μM)
0
20
40
60
80
100
0 0.5 1 1.5
%V
iabi
lity
AMP-IBP5 (μM)
B CA
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1149

Mascot. This was evident when assessed by the percent fragmentation defined by Good et al. [26]; quadruply charges ions underwent fragmentation with a mean value of 24% for CID and that of 62% for ETD in triplicate analyses. It has been shown that N-Cα bonds N-terminal to proline are not cleaved by ETD [25]. Consistent with previous work, no product ion corresponding to the proline cleavage was present in the ETD spectrum (Fig. 5). Excluding the five proline residues, the peptide has 18 theoretically cleavable bonds. Notably, 17 of the 18 bonds were actually cleaved, so that ETD achieved confident identification. Overall, our study is the first demonstration that ETD is a powerful tool in peptidomics as a complement to conventional CID. The study, however, does not necessarily detract from the utility of CID, as shown by our observation that N-terminally truncated 13- and 14-residue peptides sharing the C-terminal amide structure were solely identified by CID [22].
Fig. 5 CID (left panel) and ETD (right panel) MS/MS spectra of pentuply charged m/z 578.12. It was unequivocally identified as VGF [554-577]-NH2 by ETD. Ions matching the theoretical ions with a mass error of less than 0.025 Da are labelled. Note that in Mascot calculation the smaller expectation value shows the better match between observed and calculated fragment ions.
3.3. Naturally occurring form
Major components present in culture medium were VGF [554-577]-NH2 and [565-577]-NH2 [22], as assessed by MALDI analysis of immunoprecipitates using the antibody that specifically recognizes the proline amide structure. However, we recently got evidence that a set of larger N-terminally extended peptides are released by endocrine cell lines [22].
3.4. Peptide processing sites
Unlike AMP-IBP5, VGF [554-577]-NH2 is C-terminally not followed by a typical amidation motif harboring dibasic amino acids. However, the sequence 574RHYPGRE580 corresponds to the PC1/3 or PC2 consensus motif. It is therefore presumed that the precursor undergoes endoproteolytic processing C-terminal to 579R, followed by a series of processing steps as in classical C-terminally amidated peptides. The amidation motif is shared by primate VGF sequences, but not found in other mammals (Fig. 4), suggesting that some biological role unique to primates is attributed to the peptide. The N-terminus of human VGF [554-577]-NH2, 554T, is shared by a set of ETD-identified peptides, including VGF [554-577]-NH2, [554-583] and [554-615] [22]. The threonine corresponds to 556T in the mouse/rat sequence. In rat brain, VGF [556-617]/ TLQP-62 is a major peptide recognized by antibody against the C-terminus of the protein [27]. Given the sequence identity between human and rat, this cleavage site (550YIPR↓TLQP557), though not typical for the PC recognition motif, may represent a major processing site also in human (Fig. 4). We suggest that the peptide would follow a series of processing reactions common to neuropeptide processing.
3.5. Antimicrobial activity of VGF [554-577]-NH2 and related peptides
VGF [554-577]-NH2 showed strong antimicrobial activity with an IC50 of 1.8 and 4.5 µM against M. luteus and P. pastoris GS115, respectively (Table 2), comparable to BD2 and CATH. We also tested related VGF [554-578], VGF [554-583] and VGF [565-577]-NH2, because they were detected in the original pancreatic cell line, and their N- and C-terminal cleavage sites fit a consensus motif for PC1/3 and PC2 cleavage sites. N-terminally truncated VGF [565-577]-NH2 did not show antimicrobial activity even at 10 μM (Table 2), suggesting that the N-terminal 11 residues are essential for activity. On the other hand, C-terminal extension, assessed with VGF [554-578] and [554-583], reduced or abolished activity against M. luteus, or P. pastoris GS115, respectively (Table 2). This observation suggests that the C-terminal amide structure is also important. However, C-terminally amidated peptides are not always antimicrobial, such as other VGF-derived C-terminally amidated peptides, VGF [565-577]-NH2, Neuroendocrine regulatory peptides (NERPs)-1 and -2 [28].
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1150

To examine whether VGF [554-577]-NH2 is microbiocidal or just microbiostatic, we also performed a classical colony formation assay. The peptide showed strong antimicrobial activity in the colony assay with an IC50 of 1.9 and 1.4 µM against M. luteus and P. pastoris GS115, respectively, indicating that it is bactericidal.
Table 2 Antimicrobial activity of VGF [554-577]-NH2 and related peptides
Microbes \VGF peptide
IC50 (μM)a
[554-577]-NH2 [554-578] [554-583] [565-577]-NH2
Gram (+)
M. luteus 1.8 5.6 5.8 >10
Fungus
P. pastoris GS115 4.5 >10 >10 >10 a Fifty percent growth inhibitory concentration.
4. Implication of peptidomics in AMP identification
Our MS profiling study allows secretory peptide identification on a large scale never achieved before. The two novel AMPs were identified in a non-biased MS/MS analysis of the peptides released by human PAM-expressing endocrine cells. and selected as candidate AMPs solely on the basis of their sequence characteristics. We should also underscore the importance of studying peptides in their native form, rather than in vitro digestion with enzyme (e.g. trypsin) which is routinely applied in conventional proteomics. It should be stressed that both AMP-IBP5 and VGF [554-577]-NH2 are generated through a regulated mechanism of proteolytic processing and distinct from protein degradation fragments. IGFBP-5 is known to modulate the cellular availability of its binding partners insulin-like growth factors (IGFs), but recently IGF-independent actions have also been described [29]. Until the discovery of AMP-IBP5, IGFBP-5 was not considered as a bioactive peptide precursor. The peptide has thus added to our knowledge about IGFBP-5 biological functions. The neurosecretory protein VGF constitutes an important member of the granin family; however, because of low abundance in normal endocrine tissues, VGF-derived peptides have received much less attention than classical granins such as chromogranins A and B. Cysteines are important components in AMPs. In general, cysteine-containing peptides are reductive alkylated to facilitate MS/MS sequencing. Accordingly, at the initial MS profiling stage, it was not certain that the cysteines of AMP-IBP5 would be disulfide-linked in its internal sequence. As described in Section 2.3, we have reported that AMP-IBP5 has a molecular mass of 2653.4 Da in physiological condition, consistent with the presence of an intramolecular disulfide linkage (C199-C210) [10]. Using MS techniques, Nili et al. recently determined all disulfide linkages of the mouse IGFBP-5 overexpressed in mouse embryonic fibroblasts with an adenovirus vector [30]. They reported that C172-C199, C210-C221, and C223-C243 were disulfide-linked in the IGFBP-5 sequence. Notably, they identified C199-C210 as a very minor component. We should keep in mind, however, that the study investigated an overexpressed recombinant protein, not an endogenous protein. On the other hand, we recently got evidence to support the original finding that AMP-IBP5 is secreted in the molecular form with the linkage C199-C210. This was revealed by ETD, without reductive alkylation, using the culture supernatant of the pancreatic cell line that endogenously expresses IGFBP-5 (Fig. 6). Although the issue of natural disulfide linkage remains to be clear, IGFBP-5 may exist in two different conformations. In any case, bioinformatics-assisted prediction of AMP-IBP5 would be practically impossible, whatever the canonical amidation motif GRKR. In conclusion, the present study points to the impact of peptidomics for uncovering novel AMPs. Our MS-based peptidomic approach is applicable to any other species with available genome sequences.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1151

Fig. 6 ETD MS/MS spectrum of quadruply charged native AMP-IBP5. Note that the peptide was subjected to MS/MS without reductive alkylation.
References
[1] Machado A, Sforça ML, Miranda A, Daffre S, Pertinhez TA, Spisni A, Miranda MT. Truncation of amidated fragment 33-61 of bovine alpha-hemoglobin: effects on the structure and anticandidal activity. Biopolymers 2007; 88: 413-426.
[2] Clynen E, Baggerman G, Veelaert D, Cerstiaens A, Van der Horst D, Harthoorn L, Derua R, Waelkens E, De Loof A, Schoofs L. Peptidomics of the pars intercerebralis-corpus cardiacum complex of the migratory locust, Locusta migratoria. Eur J Biochem 2001; 268: 1929-1939.
[3] Schrader M, Schultz-Knappe P. Peptidomics technologies for human body fluids. Trends Biotechnol 2001; 10: S55-60. [4] Sasaki K, Satomi Y, Takao T, Minamino N. Snapshot peptidomics of the regulated secretory pathway. Mol Cell Proteomics
2009; 8: 1638-1647. [5] Sasaki K, Takahashi N, Satoh M, Yamasaki M, Minamino N. A peptidomics strategy for discovering endogenous bioactive
peptides. J Proteome Res 2010; 9: 5047-5052. [6] Metz-Boutigue MH, Goumon Y, Strub JM, Lugardon K, Aunis D. Antimicrobial chromogranins and proenkephalin-A-derived
peptides. Antibacterial and antifungal activities of chromogranins and proenkephalin-A-derived peptides. Ann NY Acad Sci 2003; 992: 168-178.
[7] Zasloff M. Antimicrobial peptides of multicellular organisms. Nature 2002; 415: 389-395. [8] Dos Santos Cabrera MP, Arcisio-Miranda M, Broggio Costa ST, Konno K, Ruggiero JR, Procopio J, Ruggiero Neto J. Study of
the mechanism of action of anoplin, a helical antimicrobial decapeptide with ion channel-like activity, and the role of the amidated C-terminus. J Pept Sci 2008; 14: 661-669.
[9] Suetake T, Aizawa T, Koganesawa N, Osaki T, Kobashigawa Y, Demura M, Kawabata S, Kawano K, Tsuda S, Nitta K. Production and characterization of recombinant tachycitin, the Cys-rich chitin-binding protein. Protein Eng 2002; 15: 763-769.
[10] Osaki T, Sasaki K, Minamino N. Peptidomics-based discovery of an antimicrobial peptide derived from insulin-like growth factor-binding protein 5. J Proteome Res 2011; 10: 1870-1880.
[11] Fricker LD. Neuropeptide-processing enzymes: applications for drug discovery. AAPS J. 2005; 7: E449-455. [12] Pan H, Che FY, Peng B. Steiner DF, Ointar JE, Fricker LD. The role of prohormone convertase-2 in hypothalamic neuropeptide
processing: a quantitative neuropeptidomic study. J Neurochem 2006; 98: 1763-1777. [13] Zhang X, Pan H, Peng B, Steiner DF, Pintart JE, Fricker LD. Neuropeptidomic analysis establishes a major role for prohormone
convertase-2 in neuropeptide biosynthesis. J Neurochem 2010; 112: 1168-1179. [14] Bauer F, Schweimer K, Klüver E, Conejo-Garcia JR, Forssmann WG, Rösch P, Adermann K, Sticht H. Structure determination
of human and murine beta-defensins reveals structural conservation in the absence of significant sequence similarity. Protein Sci 2001; 10: 2470-2479.
[15] Osaki T, Omotezako M, Nagayama R, Hirata M, Iwanaga S, Kasahara J, Hattori J, Ito I, Sugiyama H, Kawabata S. Horseshoe crab hemocyte-derived antimicrobial polypeptides, tachystatins, with sequence similarity to spider neurotoxins. J Biol Chem 1999; 274: 26172-26178.
[16] DeForge LE, Billeci KL, Kramer SM. Effect of IFN-γ on the killing of S. aureus in human whole blood. Assessment of bacterial viability by CFU determination and by a new method using alamarBlue. J Immunol Methods 2000; 245: 79-89.
[17] Ganz T. Defensins: antimicrobial peptides of innate immunity. Nat Rev Immunol 2003; 3: 710-720. [18] Boman HG. Peptide antibiotics and their role in innate immunity. Annu Rev Immunol 1995; 13: 61-92. [19] Nizet V, Ohtake T, Lauth X, Trowbridge J, Rudisill J, Dorschner RA, Pestonjamasp V, Piraino J, Huttner K, Gallo RL. Innate
antimicrobial peptide protects the skin from invasive bacterial infection. Nature 2001; 414: 454-457. [20] Wehkamp J, Schauber J, Stange EF. Defensins and cathelicidins in gastrointestinal infections. Curr Opin Gastroenterol 2007;
23: 32-38
m/z
AVYLPNCDRKGFYKRKQCKPSR-NH2
Mascot score 36expt. 0.75
% Fragmentation= 23/42 (55%)
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1152

[21] Gallo RL, Kim KJ, Bernfield M, Kozak CA, Zanetti M, Merluzzi L, Gennaro R. Identification of CRAMP, a cathelin-related antimicrobial peptide expressed in the embryonic and adult mouse. J Biol Chem 1997; 272: 13088-13093.
[22] Sasaki K, Osaki T, Minamino N. Large-scale identification of endogenous secretory peptides using electron transfer dissociation mass spectrometry. Mol Cell Proteomics 2013; 12: 700-709.
[23] Tabb DL, Huang Y, Wysocki VH, Yates JR 3rd. Influence of basic residue content on fragment ion peak intensities in low-energy collision-induced dissociation spectra of peptides. Anal Chem 2004; 76: 1243-1248.
[24] Syka JE, Coon JJ, Schroeder MJ, Shabanowitz J, Hunt DF. Peptide and protein sequence analysis by electron transfer dissociation mass spectrometry. Proc Natl Acad Sci U S A. 2004; 101: 9528-9533.
[25] Mikesh LM, Ueberheide B, Chi A, Coon JJ, Syka JE, Shabanowitz J, Hunt DF. The utility of ETD mass spectrometry in proteomic analysis. Biochim Biophys Acta 2006; 1764: 1811-1822.
[26] Good DM, Wirtala M, McAlister GC, Coon JJ. Performance characteristics of electron transfer dissociation mass spectrometry. Mol Cell Proteomics 2007; 6: 1942-1951.
[27] Bartolomucci A, Possenti R, Mahata SK, Fischer-Colbrie R, Loh YP, Salton SR. The extended granin family: structure, function, and biomedical implications. Endocr Rev 2011; 32: 755-797.
[28] Yamaguchi H, Sasaki K, Satomi Y, Shimbara T, Kageyama H, Mondal MS, Toshinai K, Date Y, González LJ, Shioda S, Takao T, Nakazato M, Minamino N. Peptidomic identification and biological validation of neuroendocrine regulatory peptide-1 and -2. J Biol Chem 2007; 282: 26354-26360.
[29] Mohan S, Baylink DJ. IGF-binding proteins are multifunctional and act via IGF-dependent and -independent mechanisms. J Endocrinol 2002; 175: 19-31.
[30] Nili M, Mukherjee A, Shinde U, David L, Rotwein P. Defining the disulfide bonds of insulin-like growth factor-binding protein-5 by tandem mass spectrometry with electron transfer dissociation and collision-induced dissociation. J Biol Chem 2012; 287: 1510-1519.
Microbial pathogens and strategies for combating them: science, technology and education (A. Méndez-Vilas, Ed.)
© FORMATEX 2013
____________________________________________________________________________________________
1153





























