Embed Size (px)
Citation preview

Aus dem Institut für Anatomie und Zellbiologie der Universität Würzburg Lehrstuhl für Anatomie II
Vorstand: Professor Dr. med. D. Drenckhahn
Lokalisierung und Charakterisierung von Zellkontaktproteinen im olfaktorischen Epithel
INAUGURAL-DISSERTATION zur Erlangung der Doktorwürde der
Medizinischen Fakultät der
Bayerischen Julius-Maximilians-Universität zu Würzburg vorgelegt von
Anna Sofia Meier-Stiegen aus Hamburg
Würzburg, August 2004

Referent: Prof. Dr. med. E. Asan Korreferent: Prof. Dr. med. D. Drenckhahn Dekan: Prof. Dr. med. S. Silbernagl Tag der mündlichen Prüfung: 25. Januar 2005 Die Promovendin ist Assistenzärztin

Die Einsicht oder Ansicht, dass das Vollkommene und die Wissenschaft
Stückwerk ist, darf niemanden daran hindern, doch stets weiterzubauen
und eben doch das Mögliche zu erreichen.
(Hermann Hesse, unveröffentlichte Briefe)

Inhaltsverzeichnis
Inhaltsverzeichnis
1. Einleitung 1
1.1 Topographie und Morphologie der Riechschleimhaut 2
1.2 Interzellularkontakte der Riechschleimhaut 9
1.3 Die Familie der Cadherine 13
1.4 Zielsetzung der vorliegenden Arbeit 14
2. Material 15
2.1 Geräte und übrige Materialien 15
2.2 Chemikalien 15
2.3 Puffer 16
2.4 Antiseren 16
2.5 Größenstandard 19
2.6 Peptid 86-2 19
3. Methoden 20
3.1 Perfusion von Geweben 20
3.2 LR-White-Einbettung 20
3.3 Epon-Einbettung 21
3.4 Trimmen 21
3.5 Semidünnschnitte 22
3.6 Lichtmikroskopie 22
3.7 Ultradünnschnitte 22
3.8 Kontrastierung von Ultradünnschnitten 22
3.9 Immunelektronenmikroskopie 23
3.10 Silberverstärkung nach Danscher 23
3.11 Herauslösen von Epon aus Semidünnschnitten 24
3.12 Fluoreszenzimmunhistochemie 24
3.13 Herstellung eines Peptidantikörpers 25
3.13.1 Koppelung von Peptid an KLH 25
3.13.2 Aufbereitung von Dialyseschläuchen 25
3.13.3 Dialyse der Lösung von KLH-gekoppeltem Peptid gegen PBS 26
3.13.4 Immunisierung eines Kaninchens 26

Inhaltsverzeichnis
3.13.5 Aufbereitung und Testung des Immunserums 27
3.13.6 Affinitätsreinigung des Immunserums 27
3.14 Amidoschwarz-Proteinbestimmung 28
3.15 Bradford-Proteinbestimmung 29
3.16 SDS-Polyacrylamid-Gelelektrophorese (SDS-Page) 29
3.17 Immunreplikanalyse (Western-Blot) 30
4. Ergebnisse 33
4.1 Darstellung von Proteinen der Zonula occludens im olfaktorischen
Epithel 33
4.2 Darstellung von Proteinen der Zonula adherens im olfaktorischen
Epithel 35
5. Diskussion 56
6. Zusammenfassung 64
7. Literaturverzeichnis 65
Danksagung und Lebenslauf


Einleitung 1
1. Einleitung
Zellen eines Organismus bilden Kontakte zu zellulären und extrazellulären
Komponenten ihrer Umgebung aus, um vielfältigste Funktionen in so unterschiedlichen
Bereichen wie der Entwicklung von Zellen und Geweben, der Gerinnung und der
Immunantwort zu erfüllen (Vaughn et al., 1996; Petruzzelli et al., 1999). Im Verlauf der
letzten Jahre konnten zahlreiche Erkenntnisse über die daran beteiligten
Kontaktproteine gewonnen werden. Insbesondere die Möglichkeit, diese
Kontaktproteine auf molekularer Ebene zu charakterisieren, erlaubte deren Einteilung in
die Familie der Cadherine, der Immunglobuline, der Integrine und der Selektine
(Petruzzelli et al., 1999).
Eine der interessantesten Aufgaben der Kontaktproteine ist ihre Beteiligung an der
Entwicklung des zentralen Nervensystems. Hierbei sind von besonderer Bedeutung die
Proteine der Cadherin-Superfamilie. Sie wurden als Bestandteile von Synapsen
identifiziert und scheinen als solche an Bildung und Plastizität synaptischer
Verbindungen beteiligt zu sein (Suzuki et al., 1997; Tang et al., 1998; Manabe et al.,
2000; Tanaka et al., 2000).
Ein einzigartiger Abschnitt des zentralen Nervensystems ist das olfaktorische System.
Die ersten Neurone der Riechbahn sind bipolare Sinneszellen, die in der
Riechschleimhaut, einem Neuroepithel, lokalisiert sind. Sie sind die einzigen Neurone,
die in unmittelbarem Kontakt zur Außenwelt stehen. Durch die Ausbildung
epitheltypischer Kontakte zu umgebenden Zellen sind sie an der Epithelbarriere
beteiligt. Sie differenzieren sich aus neuronalen Vorläuferzellen, die im basalen
Kompartiment des olfaktorischen Epithels gelegen sind. Diese Basalzellen besitzen ein
Leben lang die Fähigkeit zur Zellteilung und Differenzierung. Damit verbunden ist das
Wachstum und die Zielfindung von Axonen sowie die Ausbildung synaptischer
Verbindungen.
Aufgrund dieser hohen Plastizität ist das Riechepithel ein ideales Modell zur
Untersuchung von Zellkontakten insbesondere der Nervenzellen (Miragall et al., 1994;
Alenius et al., 1997).

Einleitung 2
1.1 Topographie und Morphologie der Riechschleimhaut
Die Riechschleimhaut des Nagers (Pars olfactoria tunicae mucosae nasi) hat eine Fläche
von ungefähr 62 mm2 und kleidet beidseits die obere Hälfte der Nasenscheidewand und
der lateralen Nasenwand sowie das Nasendach aus (Mendoza, 1993). Ihre Abgrenzung
zur respiratorischen Schleimhaut (Pars respiratoria tunicae mucosae nasi) ist unscharf.
Es finden sich auch versprengte Anteile des mehrreihigen Flimmerepithels inmitten des
sensorischen Epithels (Shipley et al., 1995).
Die Riechschleimhaut besitzt ein mehrreihiges Epithel. Es setzt sich aus folgenden
Zelltypen zusammen: Stützzellen (SZ), Sinneszellen (olfaktorische Neurone, ON),
Basalzellen (BZ), Mikrovillizellen (MV) sowie Zellen des Ausführungsganges der
Bowmanschen Drüsen (AZ). Eine Basalmembran bildet die Grenze zur Lamina propria.
In deren Bindegewebe sind zahlreiche Blutgefäße, die tubuloazinösen Bowmanschen
Drüsen sowie die von olfaktorischen Schwannzellen umgebenen Fila olfactoria (=
gebündelte Axone der Sinneszellen) enthalten. Letztere bilden zusammengenommen
den Nervus olfactorius (Mendoza, 1993).
Die Zellkörper der verschiedenen Zelltypen der Riechschleimhaut liegen in Schichten
übereinander. Von basal nach apikal findet sich die Basalzellschicht, die
Sinneszellschicht und die Stützzellschicht. Zellkörper der Mikrovillizellen liegen in der
oberflächlichen Sinneszellschicht und der Stützzellschicht. Die Zellkörper der Zellen
des Ausführungsganges der Bowmanschen Drüsen durchziehen das olfaktorische
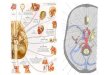
Epithel vertikal (Abb. 1.1.1).

Einleitung 3
Abb. 1.1.1: Das olfaktorische Epithel.
Links ein mit Methylenblau gefärbter Semidünnschnitt, rechts ein schematischer
Longitudinalschnitt durch das olfaktorische Epithel der Ratte (aus Asan, E. in:
Benninghoff-Drenckhahn, 2004). MV: Mikrovillizellen; SC: Stützzellen; ON:
olfaktorische Neurone; B: Basalzellen; AZ: Ausführungsgangszellen; LP: Lamina
propria; Maßstab linkes Bild: 20 µm.
Aufgrund ihrer typischen Lage und Morphologie können die einzelnen Zelltypen der
Riechschleimhaut meist in licht- und elektronenmikroskopischen Gewebepräparaten
erkannt werden. Darüberhinaus ist ihre Identifizierung durch charakteristische
Zytoplasma- und Plasmamembranproteine mit immunhistochemischen Methoden
möglich.
Stützzellen sind säulenförmige Zellen. Basal erreichen sie die Basalmembran, apikal
mit ihrem Mikrovillisaum das Lumen der Nasenhöhle. Ihr Kernniveau liegt über dem
der Sinneszellen. Das apikale Zytoplasma ist reich an glattem endoplasmatischen
Retikulum, das in parallel liegenden Lamellen angeordnet ist (Mendoza, 1993). Basal
finden sich zahlreiche lipofuszinähnliche lamellierte dichte Körperchen lysosomaler
Natur, welche die Riechschleimhaut makroskopisch gelbbräunlich erscheinen lassen
(Drenckhahn et al., 1994). Apikal sind die Stützzellen untereinander und mit
benachbarten Dendriten der Sinneszellen durch Zellkontakte verbunden, die
elektronenmikroskopische Charakteristika von Zonulae occludentes, Zonulae

Einleitung 4
adherentes und Desmosomen aufweisen (Mendoza, 1993). Stützzellen besitzen das für
sie typische Intermediärfilament Zytokeratin 18 (CK-18) (Abb. 1.1.2; Drenckhahn et al.,
1994).
Basalzellen finden sich dicht über der Basalmembran. Es werden zwei Gruppen
unterschieden, rundliche und horizontale (Mendoza, 1993). Rundliche Basalzellen
haben einen euchromatinreichen Kern und, im Gegensatz zu horizontalen Basalzellen,
keine Tonofilamente oder Ribosomen. Rundliche Basalzellen unterliegen ständiger
mitotischer Aktivität. Erst vor kurzem konnte gezeigt werden, daß rundliche Basalzellen
neuronale Stammzellen sind (Jang et al., 2003). Während der Differenzierung ihrer
Tochterzellen zu olfaktorischen Neuronen wandert der Zellkörper nach apikal, der
Zellkern wird euchromatinreicher, der Dendrit erstreckt sich entlang der Stützzellen zur
Oberfläche des olfaktorischen Epithels und das Axon bahnt sich seinen Weg durch die
Basalmembran in die Lamina propria (Morrison et al., 1992). Horizontale Basalzellen
sind polygonal, haben einen heterochromatinreichen Kern und zahlreiche
Tonofilamente (Drenckhahn et al., 1994). Sie umgeben die zur Lamina propria
ziehenden intraepithelialen kleinen Faszikel der Sinneszell-Axone (Mendoza, 1993).
Ihre Funktion ist letztlich nicht geklärt.
Die Zellen des Ausführungsganges der Bowmanschen Drüsen haben einen
kuboidalen Zellkörper. Dessen Längsachse verläuft parallel zur Richtung des
Ausführungsganges. Seine luminale Plasmamembran enthält zahlreiche Mikrovilli (aus
Asan, E. in: Benninghoff-Drenckhahn, 2004) Die Ausführungsgangzellen sind
epithelialer Herkunft und CK-18 positiv (Abb. 1.1.2). Ihre basolaterale Plasmamembran
enthält Na+/K+-ATPase. Die Bowmanschen Drüsen produzieren den Riechschleim. Es
wird vermutet, daß Zellen der Drüsen oder des Ausführungsganges Stammzellen der
epithelialen Zellen des olfaktorischen und des respiratorischen Epithels sind (Jang et al.,
2003).

Einleitung 5
Abb. 1.1.2: Stützzellen, Mikrovillizellen und Zellen des Ausführungsgangs der Bowmanschen
Drüsen.
Inkubation eines longitudinalen Semidünnschnitts durch das olfaktorische Epithel mit
CK-18-Antikörper und Gam-Cy3. AZ: Ausführungsgangszelle; MV: Mikrovillizelle;
SZ: Stützzelle; Maßstab: 20 µm; (Abbildung: E. Asan).
Die Sinneszellen der Riechschleimhaut sind bipolare Neurone. Ihr Dendrit, der reich an
Mitochondrien und Miktrotubuli ist, zieht geradewegs oder gewunden nach apikal. Im
Bereich des Epithelsaumes ist er kolbenförmig zum 1 bis 2 µm im Durchmesser großen
dendritischen Kolben („olfactory knob“) aufgetrieben. Darin befinden sich
Basalkörperchen, aus denen ca. 10 bis 25 Riechgeißeln hervorgehen. Diese besitzen in
ihrem Anfagsteil ein typisches 9 x 2 + 2-Mikrotubulusskelett. Mit zunehmender Länge
nimmt die Anzahl der Mikrotubuli jedoch ab (Moran et al., 1982). Zwischen
benachbarten Sinneszellen und Stützzellen sind auf Höhe der Riechkolben Zellkontakte
bandförmig angeordnet. Der Zellkörper der Neurone liegt unter dem der Stützzellen.
Basal entspringt das 0,1 bis 0,7 µm im Durchmesser große Axon. Es wird oftmals von
Fortsätzen der Stützzellen umgeben. Intraepithelial bilden mehrere Axone kleine
Faszikel, die, von Ausläufern der horizontalen Basalzellen umhüllt, die Basalmembran
durchbrechen und sich in der Lamina propria zu größeren Bündeln sammeln. Diese
erreichen über die Lamina cribrosa des Os ethmoidale die Schädelhöhle, wo sie als N.
olfactorius zum Bulbus olfactorius ziehen (Morrison et al., 1992). Alle Sinneszellen

Einleitung 6
sind reich an β-Tubulin, da sie ein Mikrotubulussystem haben, welches den Dendriten,
den Zellkörper und das Axon durchzieht. Zudem findet sich in ihrer basolateralen
Memban Na+/K+-ATPase, die unter anderem nach Aktionspotentialen das
Ruhemembranpotential wieder herstellt (Abb. 1.1.3).
Eine Besonderheit olfaktorischer Sinneszellen ist das olfaktorische Markerprotein OMP
(Margolis, 1972; Abb. 1.1.3). Es findet sich ausschließlich in Dendrit, Zellkörper und
Axon reifer olfaktorischer Neurone (Morrison et al., 1992). Dabei handelt es sich um
ein niedrigmolares saures Protein, das möglicherweise trophische Funktionen hat
(Drenckhahn et al., 1994).
Abb. 1.1.3: Sinneszellen des olfaktorischen Epithels.
Inkubation von longitudinalen semidünnen Folgeschnitten durch das olfaktorische
Epithel mit OMP-Antikörper (links) und β-Tubulin-Antikörper (rechts). OMP-
Antikörper markieren Zilien, Dendriten, Zellkörper und Axone der olfaktorischen
Neurone, β-Tubulin-Antikörper hauptsächlich Zilien, Dendriten und Axone; Maßstab:
10 µm; (Abbildung: E. Asan).
Mikrovillizellen sind regelmäßig vorkommende Bestandteile des olfaktorischen
Epithels. Ihr Verhältnis zu Sinneszellen beträgt 1:10 bis 1:20. Ihre birnenförmigen
Zellkörper sind apikal lokalisiert. Sie besitzen einen apikalen Fortsatz, von dem
ausgehend die namensgebenden Mikrovilli in das Lumen der Nasenhöhle ragen. Ihr

Einleitung 7
Zytopolasma ist relativ arm an Organellen. Meist werden zwei Typen unterschieden.
Viele Mikrovillizellen haben einen ca. 0,4 bis 0,6 µm dünnen basalen axon-ähnlichen
Fortsatz (Morrison et al., 1992).
Abb. 1.1.4: Mikrovillizelle.
Ultradünner Longitudinalschnitt durch das apikale olfaktorische Epithel. Pfeil: Apikaler
Zellkontakt. MV: Mikrovilluszelle; SZ: Stützzelle; Maßstab: 1 µm.
Mikrovillizellen sind epithelialer Herkunft und synthetisieren das Intermediärfilament
Zytokeratin 18. Sie zeigen deutliche Immunreaktivität gegenüber Ankyrin in ihrer
basolateralen Plasmamembran (Abb. 1.1.5). Bei Mikrovillizellen vom Typ 1 findet sich
Villin im Bereich der apikalen Mikrovilli sowie der basolateralen Plasmamembran.
Mikrovillizellen vom Typ 2 sind reich an Na+/K+-ATPase, die in der basolateralen
Plasmamembran lokalisiert ist. Morphologische und immunhistochemische Befunde
legen eine Identifizierung von Mikrovillizellen vom Typ 1 als Bürstenzellen nahe,
welche möglicherweise chemosensitive Funktion haben (Hofer et al., 1999). Die
Aufgabe der Mikrovillizellen vom Typ 2 ist nicht geklärt. Mikrovillizellen vom Typ I
und II unterscheiden sich in ihrem Gehalt an einzelnen Proteinen (Tab. 1.1.1; Asan et
al., 1999).

Einleitung 8
Abb. 1.1.5: Mikrovillizelle.
Inkubation eines longitudinalen Semidünnschnitts durch das olfaktorische Epithel mit
Ankyrin-Antikörper und Gam-Cy3; Maßstab: 10 µm.
Mikrovillizellen Typ I
(Bürstenzellen)
Mikrovillizellen Typ II
Ankyrin + ++
β-Tubulin (+) (+)
Cytokeratin 18 (CK 18) ++ ++
E-Cadherin ++ ++
Na+/K+-ATPase (+) ++
Villin ++ -
Tab. 1.1.1: Immunreaktivität der Mikrovillizellen vom Typ I und II.
Erscheinungsbild der Zellgrenzen und Interzellularkontakte im histologischen
Schnittbild
Im Longitudinalschnitt stellen sich die Zellgrenzen zwischen allen Zelltypen als mehr
oder weniger longitudinale Streifen dar (Abb. 1.1.6). Das Schema der Zellgrenzen im
Tangentialschnitt beruht zunächst auf theoretischen Überlegungen und Literaturangaben
(Abb. 1.1.6). Es basiert auf der Beobachtung, dass Dendriten im Bereich ihres apikalen
Endes durch Stützzellen teilweise oder vollständig eingehüllt werden (Mendoza, 1993;
Miragall et al., 1994; Hussar et al., 2002). Sollte dies auch der Fall für die apikalen
Enden der Mikrovillizellen sein, deren Durchmesser im Vergleich zu dem der Dendriten
größer ist, könnte sich im apikalen Tangentialschnitt ein Bild für die Zellgrenzen wie

Einleitung 9
unten gezeigt ergeben: Zellgrenzen zwischen Stützzellen hätten eine polygonale Form,
Zellgrenzen zwischen Stützzellen und Mikrovillizellen wären in Form größerer Kreise
zu erwarten, Zellgrenzen zwischen Stützzellen und Dendriten in Form kleinerer Kreise.
Da die Ausführungsgangszellen das Epithel longitudinal durchziehen, wären im
Tangentialschnitt die Kontakte zwischen Stützzellen und Ausführungsgangszellen als
sehr große Kreise, die Kontakte zwischen Ausführungsgangszellen als radiäre Linien
innerhalb dieser Kreise zu erwarten (siehe auch Abb 4.2.11).
Abb. 1.1.6: Schematische Darstellung der Markerproteine und der möglichen Topographie
der Zellgrenzen für die Zelltypen des olfaktorischen Epithels.
Schema eines Longitudinalschnittes und eines apikalen Tangentialschnittes durch das
Riechepithel. Gelb: Cytokeratin 18; rot: Na+/K+-ATPase; grün: β-Tubulin oder OMP;
hellblau: Villin; dunkelblau: Ankyrin; (Abbildung: E. Asan).
1.2 Interzellularkontakte der Riechschleimhaut
Das elektronenmikroskopische Bild der Riechschleimhaut zeigt im apikalen Bereich
typische epitheliale Interzellularkontakte. Dabei können der Barrierenkontakt Zonula
occludens („tight junction“) sowie die Adhäsionskontakte Zonula adherens und Macula
adherens (Desmosom) identifiziert werden. Die Kombination aus Zonula occludens,

Einleitung 10
Zonula adherens und Macula adherens wird Haft- oder Schlußleistenkomplex genannt
(Drenckhahn et al., 1994). Lateral finden sich zudem Puncta adherentia.
Alle Interzellularkontakte weisen vergleichbare Strukturen auf. Transmembranproteine
bilden extrazellulär homo- oder heterotypische Zell-Zell-Kontakte aus. Intrazellulär sind
sie mit zytoplasmatischen Plaqueproteinen verbunden, welche den Kontakt zum
Zytoskelett herstellen, regulatorische Funktionen haben und an der Signaltransduktion
beteiligt sind (Balda et al., 2003).
Die Zonula occludens ist der am meisten apikal gelegene epitheliale
Interzellularkontakt, der die Zellmembran in eine apikale und eine basolaterale Domäne
unterteilt. Dies ist Voraussetzung sowohl für die Entstehung als auch für die
Aufrechterhaltung der physiologischen Funktionen eines epithelialen Gewebes. Hierzu
gehören die Zellpolarität, die intramembranäre Diffusionsbarriere, die semipermeable
parazelluläre Diffusionsbarriere und der selektive transzelluläre Transport (Creijido,
1999; Balda et al., 2000). Erst vor kurzem wurde die Zonula occludens mit Vorgängen
wie der Regulierung von Zelldifferenzierung und Zellproliferation in Zusammenhang
gebracht. Dies scheint über die Modulierung bekannter Signaltransduktionsvorgänge zu
erfolgen (Li et al., 2000; Reichert et al., 2000; Vietor et al., 2001).
Elektronenmikroskopische Aufnahmen von Gefrierbrüchen zeigen, daß die Zonula
occludens aus einem Netzwerk an Leisten besteht (Staehelin, 1973). Deren
Molekularstruktur war bis vor kurzem kaum bekannt. Mittlerweile sind drei
Transmembranproteine identifiziert: Occludin (Abb. 1.2.1), die Claudine und die JAMs
(„junctional adhesion molecules“) (Van Itallie et al., 1997; Martin-Padura et al., 1998;
Kubota et al., 1999). Alle drei Proteine vermitteln Interzellularkontakte. Occludin und
die Claudine sind funktionelle Bestandteile der semipermeablen parazellulären
Diffusionsbarriere (Balda et al., 2003). Die Transmembranproteine stehen im
Zytoplasma mit Adaptorproteinen der MAGUK-Familie („membrane-associated
guanylate kinases“), den Proteinen mit PDZ-Domäne („PSD-95, Discs large A, ZO-1
homology“; Abb. 1.2.1) und zahlreichen Regulatorproteinen und Proteinen der
Signaltransduktion wie Proteinkinasen, Proteinphosphatasen, kleinen und
heterotrimeren GTP-bindenden Proteinen und Regulatoren der Transkription in Kontakt

Einleitung 11
(Tsukita et al., 2001; Gonzalez-Mariscal et al., 2003). Die Zonula occludens hat
Beziehung zum Aktin-Zytoskelett (Balda et al., 2003).
Abb. 1.2.1: Zonula occludens und Zonula adherens, vereinfachtes Schema (Abbildung: D.
Drenckhahn).
Die Zonula adherens ist einer der bedeutendsten Interzellularkontakte (Balda et al.,
2003). Sie ist basal der Zonula occludens gelegen und umgibt gürtelförmig die
apikolaterale Plasmamembran. Die Transmembranproteine der Zonula adherens sind die
klassischen Cadherine E-Cadherin, N-Cadherin, P-Cadherin, R-Cadherin, und B-
Cadherin (Gumbiner, 1996; Petruzzelli et al., 1999). Sie sind unter anderem über β-
Catenin oder das nahe verwandte Plakoglobin (γ-Catenin) mit α-Catenin verbunden.
Zum Zytoskelettprotein F-Aktin stellt α-Catenin entweder einen direkten oder einen
indirekten Kontakt über die Aktin-bindenden Proteine α-Actinin oder Vinculin her
(Abb. 1.2.1). Sowohl die klassischen Cadherine als auch die Aktin-bindenden Proteine
sind zelltypspezifisch (Itoh et al., 1999; Nagafuchi, 2001; Pokutta et al., 2002). Vor
kurzem wurde auch Nektin als Transmembranprotein der Zonula adherens identifiziert.
Es steht über Afadin/AF-6 mit dem Aktin-Zytoskelett in Verbindung (Takai et al.,
2003).
In epithelialen Geweben bilden Zonulae adherentes erste Interzellularkontakte aus, in
deren Folge Zonulae occludentes und Maculae adherentes entstehen (Gumbiner et al.,
1988; Adams et al., 1996). Sie stabilisieren und verändern Gewebestrukturen (Mooseker
et al., 1984), regulieren die parazelluläre Permeabilität (Madara et al., 1987; Madara,

Einleitung 12
1987; Madara et al., 1988) und können kleinere epitheliale Defekte schließen (Bement
et al., 1993). Proteine, die mit der Zonula adherens assoziiert sind, sind an der
Regulation der Genexpression und an der Signaltransduktion beteiligt (Balda et al.,
2003).
Puncta adherentia sind kleine, fleckförmige Adherenskontakte. Ihre molekulare
Struktur entspricht der der Zonula adherens.
Im Gegensatz zur Zonula adherens, die in jeder Zelle vorkommt, findet sich die Macula
adherens (Desmosom) in Epithelzellen, den Glanzstreifen der Herzmuskelzellen,
Arachnoidalzellen und follikulär dendritischen Retikulumzellen des Lymphknotens. Sie
ist nicht in Form eines Gürtels, sondern einer im Durchmesser 0,1 bis 0,5 µm2 großen
Fläche basal der Zonula adherens angeordnet (Drenckhahn et al., 1994). Als
Haftproteine werden die Transmembranproteine der Desmogleine (Dsg1, Dsg2, Dsg3)
und Desmocolline (Dsc1, Dsc2, Dsc3) unterschieden, die zu den desmosomalen
Cadherinen gehören. Zytoplasmatisch sind sie über Plakoglobin und Plakophiline mit
Desmoplakin und wahrscheinlich weiteren Proteinen der Plakin-Familie (Plektin,
Envoplakin, Periplakin) verbunden. In diese zytoplasmatischen Plaques strahlen Bündel
von Intermediärfilamenten ein (Ishii, 2003).
Desmosomale Cadherine können antigene Ziele von Autoimmunerkrankungen sein,
deren Symptome die Bedeutung der Proteine bezüglich der Ausbildung von Zell-Zell-
Kontakten und der Aufrechterhaltung epithelialer Integrität deutlich machen.
Autoantikörper gegen Dsg1 rufen Pemphigus foliaceus hervor, Autoantikörper gegen
Dsg3 verursachen Pemphigus vulgaris (Amagai, 1999), Dsc1 ist Ziel des IgA-
Pemphigus (Hashimoto et al., 1997), und der paraneoplastische Pemphigus wird durch
Autoantikörper gegen Proteine der Plakin-Familie und Dsg3 verursacht (Amagai et al.,
1998). Zudem ist Dsg1 Ziel des Exfoliatin A, ein von Staphylococcus-aureus-Stämmen
der Phagengruppe II gebildetes Toxin, das die Impetigo contagiosa und das
staphylogene Lyell-Syndrom auslöst (Amagai et al., 2000).

Einleitung 13
1.3 Die Familie der Cadherine
Die Familie der Cadherine besteht aus der Gruppe der klassischen Cadherine, der
Desmogleine, der Desmocolline, der Protocadherine, der CNR („cadherin-related
neuronal receptor“)-Proteine, der Fat-Proteine, der Cadherine mit sieben
Transmembrandomänen und der Ret-Tyrosinkinasen. Die ca. 80 bekannten Cadherine
sind Transmembranproteine. Die einzelnen Gruppen unterscheiden sich zum einen in
der Anzahl der extrazellulären Domänen, zum anderen in der Aminosäuresequenz der
transmembranären und zytoplasmatischen Domäne (Yagi et al., 2000).
Am längsten bekannt und am intensivsten untersucht ist die Gruppe der klassischen
Cadherine. Dabei handelt es sich um transmembranäre Glykoproteine, die
kalziumabhängig Zell-Zell-Kontakte herstellen (Magee et al., 1991; Grunwald, 1993).
Gewöhnlich treten gleiche (homophile Kontakte) und nur ausnahmsweise ungleiche
(heterophile Kontakte) Vertreter der Proteingruppe miteinander in Kontakt. Die
klassischen Cadherine setzen sich aus fünf extrazellulären Domänen, einer
transmembranären Domäne und einer zytoplasmatischen Domäne zusammen
(Grunwald, 1993). Im Bereich der ersten extrazellulären Domäne befindet sich bei
verschiedenen Cadherinen (u.a. E-Cadherin, N-Cadherin) die Aminosäuresequenz HAV
(Histidin-Alanin-Valin), welche möglicherweise an der Ausbildung der
Transinteraktion der Cadherine beteiligt ist. Der C-Terminus jeder extrazellulären
Domäne enthält eine Bindungsstelle für Kalziumionen. Diese scheinen durch
Stabilisierung der Struktur des Cadherins den Zell-Zell-Kontakt indirekt zu beeinflussen
(Vaughn et al., 1996). Interzellularkontakte werden wahrscheinlich durch Cadherin-
Dimere vermittelt. Der Interzellularkontakt wird nach derzeitiger Vorstellung durch
einen „linear zipper“ hergestellt, in dem sich die Cadherin-Dimere wie ein
Reißverschluß ineinanderfügen (Shapiro et al., 1995a; Shapiro et al., 1995b).
Desmogleine und Desmocolline besitzen eine den klassischen Cadherinen sehr ähnliche
extrazelluläre Domäne, unterscheiden sich aber deutlich in der Aminosäuresequenz
ihrer zytoplasmatischen Domäne (Ishii, 2003).
Auf der Suche nach neuen Gruppen der Cadherine wurden aus dem Gehirn von Ratten
Proteine isoliert, die aus sechs bis sieben extrazellulären Domänen, einer
transmembranären Domäne und einer zytoplasmatischen Domäne aufgebaut sind (Sano

Einleitung 14
et al., 1993; Yagi et al., 2000). Da die extrazellulären Domänen Ähnlichkeiten mit den
klassischen Cadherinen aufwiesen, wurde die Gruppe „Protocadherine“ genannt. Die
zytoplasmatische Domäne unterscheidet sich jedoch deutlich (Sano et al., 1993).
Protocadherine bilden homotypische Interzellularkontakte aus. Bislang sind keine
Proteine bekannt, die intrazellulär mit ihnen in Verbindung stehen. Protocadherine sind
an der Embryogenese beteiligt (Yagi et al., 2000).
Die Gruppe der mit den Cadherinen verwandten neuronalen Rezeptoren CNR
(„cadherin-related neuronal receptor“) hat wie die Protocadherine sechs extrazelluläre
Domänen, eine transmembranäre Domäne und eine zytoplasmatische Domäne, die mit
der Tyrosinkinase Fyn interagiert (Kohmura et al., 1998). Fyn ist an der Regulation von
Struktur und Funktion des Gehirns beteiligt (Yagi et al., 2000).
1.4 Zielsetzung der vorliegenden Arbeit
Ziel der vorliegenden Arbeit war die Lokalisierung und Charakterisierung von
Proteinen der Barrieren- und Adhäsionskontakte im olfaktorischen Epithel. Hierzu
wurde die Riechschleimhaut der Ratte für immunhistochemische, licht- und
elektronenmikroskopische sowie biochemische Experimente aufbereitet. Das Gewebe
wurde mit Antikörpern inkubiert, die gegen Transmembranproteine und
zytoplasmatische Plaqueproteine der Zellkontakte gerichtet waren. Durch die
gleichzeitige Detektion spezifischer Markermoleküle der verschiedenen Zelltypen des
olfaktorischen Epithels wurde die genaue zelltypspezifische Zusammensetzung der
Zellkontakte dargestellt.

Material 15
2. Material
2.1 Geräte und übrige Materialien
Elektronenmikroskop EM 109 Zeiss, Jena
Elektronenmikroskop LEO AB912 Omega LEO, Oberkochen
Fluoreszenzmikroskop BH2-RFCA Olympus, Hamburg
Fotolaborchemikalien Whatman, Göttingen
Hyperfilm ECL Amersham Biosciences, Freiburg
Nitrozellulosemembran Amersham, Braunshweig
Plastikreaktionsgefäß Eppendorf, Hamburg
Schneidemaschine für Semidünnschnitte Reichert-Jung, Nussloch
Schwarz-Weiß-Filme TMAX 400 Kodak, Rochester, USA
Semi-Dry-Blot-Gerät Kreuz, Reiskirchen
Spektralphotometer Beckmann, Palo Alto, USA
Tischzentrifuge 5415 C Eppendorf, Hamburg
Ultracut E Reichert-Jung, Nußloch
Vakuumblot-Gerät Typ V jr. Schleicher und Schuell, Dassel
2.2 Chemikalien
Amidoschwarz Serva, Heidelberg
Ampuwa Fresenius Kabi, Bad Homburg
Blei (II)-zitrat-3-hydrat Merck, Darmstadt
BSA (Bovine Serum Albumine) 10% Serva, Heidelberg
CWFS (cold water fish skin) Sigma, Deisenhofen
Epon Serva, Heidelberg
GLA (Glutaraldehyd) 25% Roth, Karlsruhe
Glyzerin Roth, Karlsruhe
KLH Calbiochem-Novabiochem,
Nottingham
LR-White Agar Scientific, Berkshire

Material 16
Magermilchpulver Molkerei Feldental
Methylenblau Chroma, Stuttgart
Natriummethanolat Fluka, Steinheim
N-Propylgallat Serva, Heidelberg
(3,4,5-Trihydroxybenzoesäurepropylester)
Ponceau S Sigma, Deisenhofen
SDS (Natriumdodecylsulfat) Applichem, Darmstadt
Serva-Blau G 250 (Coomassie) Serva, Heidelberg
Tween 20 Roth, Karlsruhe
Uranylazetatdihydrat MERCK, Darmstadt
2.3 Puffer
PBS (phosphate buffered saline) 137 mM NaCl
2,7 mM KCl
8,1 mM Na2HPO4
1,5 mM KH2PO4
2.4 Antiseren
Erstantikörper:
α-Actinin-Antikörper, polyklonal Institut für Anatomie und
Zellbiologie, Würzburg
α-Catenin-Antikörper, monoklonal BDTL, Heidelberg
Ankyrin-Antikörper, polyklonal Institut für Anatomie und
Zellbiologie, Würzburg
β-Tubulin-Antikörper, monoklonal Sigma, Deisenhofen
β-Catenin-Antikörper, monoklonal BDTL, Heidelberg
CK-18-Antikörper, monoklonal Progen, Heidelberg
E-Cadherin-Antikörper, monoklonal STL

Material 17
Na+/K+-ATPase-Antikörper, polyklonal Institut für Anatomie und
Zellbiologie, Würzburg
N-Cadherin-Peptidantikörper, polyklonal Sigma, Deisenhofen
N-Cadherin-Fusionsprotein-Antikörper, polyklonal s. Fannon et al., 1996
Occludin-Antikörper, polyklonal Zymed, San Francisco, USA
Pan-Cadherin-Antikörper, polyklonal Sigma, Deisenhofen
ZO-1-Antikörper, monoklonal Biotrend, Köln
Zweitantikörper:
Garat-Cy3 (= goat-anti-rat Cy3): Jackson ImmunoResearch
Cy3 (Cyanine)-konjugiertes Ziege-anti-Ratte-IgG Laboratories, West Grove,
USA
Garb-Cy2 (= goat-anti-rabbit Cy2): Jackson ImmunoResearch
Cy2 (Cyanine)-konjugiertes Ziege-anti-Kaninchen IgG Laboratories, West Grove,
USA
Garb-Cy 3(= goat-anti-rabbit Cy3): Jackson ImmunoResearch
Cy3 (Indocarbocyanine)-konjugiertes Ziege-anti- Laboratories, West Grove,
Kaninchen IgG USA
Gam-Cy 2(= goat-anti-mouse Cy2): Jackson ImmunoResearch
Cy2 (Cyanine)-konjugiertes Ziege-anti-Maus IgG Laboratories, West Grove,
USA
Gam-Cy 3(= goat-anti-mouse Cy3): Jackson ImmunoResearch
Cy3 (Indocarocyanine)-konjugiertes Ziege-anti- Laboratories, West Grove,
Maus IgG USA

Material 18
Garb-Au 10 nm (goat-anti-rabbit Au 10 nm): Sigma, Deisenhofen
mit einem 10 nm-Goldpartikel konjugiertes Ziege-anti-
Kaninchen IgG
Garb-Au 5 nm (goat-anit-rabbit Au 5 nm) Sigma, Deisenhofen
mit einem 5 nm-Goldpartikel konjugiertes Ziege-anti-
Kaninchen IgG
Garb-Au US (goat-anti-rabbit Au ultrasmall): Aurion, Wageningen
mit einem 0,8 nm ultrasmall-Goldpartikel konjugiertes
Ziege-anti-Kaninchen IgG
Gam-Au 10 nm (goat-anti-mouse Au 10 nm): Sigma, Deisenhofen
mit einem 10 nm-Goldpartikel konjugiertes Ziege-anti-
Maus IgG
Gam-Au 5 nm (goat-anti-mouse Au 5 nm): Sigma, Deisenhofen
mit einem 5 nm-Goldpartikel konjugiertes Ziege-anti-
Maus IgG
Gam-Au US (goat-anti-mouse Au ultrasmall): Aurion, Wageningen
mit einem 0,8 nm ultrasmall-Goldpartikel konjugiertes
Ziege-anti-Maus IgG
Garb-HRP-POX (goat-anti-rabbit HRP-POX): Sigma, Deisenhofen
mit Meerettich-Peroxidase konjugiertes Ziege-anti-
Kaninchen-IgG
Gam-HRP-POX (goat-anti-mouse HRP-POX): Sigma, Deisenhofen
mit Meerettich-Peroxidase konjugiertes Ziege-anti-
Maus-IgG

Material 19
2.5 Größenstandard
Eichproteine, biotinyliert (EPH-Marker) Roche, Mannheim
(200 kDA; 116,3 kDa; 97,4 kDa; 66,2 kDa; 45 kDa)
2.6 Peptid 86-2
Aminosäuren 4 bis 22 der ersten extrazellulären Domäne des N-Cadherin der Maus
Aminosäuresequenz: KNLSLRYSVTGPGADQPPTG
Peptidsynthese durch das Theodor-Boveri-Institut für Biowissenschaften, Abteilung für
Physiologische Chemie I

Methoden 20
3. Methoden
3.1 Perfusion von Geweben
Fixiertes Gewebe: 4 erwachsene männliche Wistar-Ratten aus eigener Zucht wurden in
Ätheranästhesie über die Aorta ascendens zunächst kurz mit Vorspüllösung perfundiert.
Daraufhin wurden sie 14 min lang perfusionsfixiert. Die dafür verwendete Lösung
setzte sich für eine anschließende Epon-Einbettung (2 Ratten) aus 2 % Glutaraldehyd
und 0,6 % Paraformaldehyd, für eine anschließende LR-White-Einbettung (2 Ratten)
aus 0,1 % Glutaraldehyd und 2 % Paraformaldehyd in PBS zusammen.
Nun wurde möglichst viel des knöchernen Nasenskeletts entfernt, die Riechschleimhaut
zusammen mit unterliegendem Gewebe en bloc herauspräpariert und in ca. 5 mm dicke
Gewebeblöcke unterteilt. Diese wurden für 2 h bei 4°C in der entsprechenden
Perfusionslösung postfixiert.
Vorspüllösung:
0,5 % (w/v) Procainhydrochlorid, 6250 IE Heparin, in PBS
Gefriergetrocknetes Gewebe: Vier erwachsene männliche Wistar-Ratten wurden in
Äthernarkose dekapitiert, die Riechschleimhaut wie oben beschrieben präpariert, sofort
in mit flüssigem Stickstoff gekühltes Isopentan eingefroren und in flüssigen Stickstoff
überführt. Die gefrorenen Präparate wurden für 12 h bei -90°C, -40°C, -20°C, 0°C und
20°C im Vakuum bei 10-5 Torr gefriergetrocknet, danach bei 20°C für 48 h bei 10-3 Torr
in Epon eingebettet und anschliessend in Gelatinekapseln bei 60°C für 24 h-48 h
ausgehärtet.
3.2 LR-White-Einbettung
Das postfixierte Material für die LR-White-Einbettung (3.1) wurde dreimal 10 min lang
in PBS (2.3) gewaschen und für jeweils 5 min in 30 %, 40 %, 50 %, 60 %, 70 %, 80 %,
90 % und 96 % Ethanol entwässert. Nach dreimal 10 min 100 % Ethanol wurde 60 min
lang in LR-White inkubiert. Die Gewebeblöcke wurden in frischem LR-White auf dem
Schüttler über Nacht bei 4°C gelagert. Nun wurde erneut LR-White gewechselt. Die

Methoden 21
Präparate wurden in Gelatinecups eingebettet und zwei bis drei Tage lang bei 4°C mit
UV-Licht bestrahlt. Sie wurden für die Immun-Elektronenmikroskopie (3.11) benötigt.
3.3 Epon-Einbettung
Fixiertes Gewebe: Die Gewebeblöcke wurden aus der Perfusionslösung genommen
(3.1) und 1 h lang in 0,01 M PBS (2.3) gespült. Nach Osmierung mit 1 % (w/v) OsO4 in
0,1 M PBS für 1 h wurde mit destilliertem Wasser gespült und jeweils 15 min lang mit
30 %, 50 % und 70 % Ethanol entwässert. Nun wurde für maximal 3 h in 2 % (v/v)
Uranylacetatdihydrat in 70% Ethanol blockkontrastiert. Es folgte eine Behandlung mit
70 %, 80 %, 90 % und 96 % Ethanol für jeweils 15 min. Mit 100 % Ethanol wurde
zweimal 30 min inkubiert. Anschließend wurde zweimal 30 min mit Propylenoxid
behandelt. Über Nacht (12 h) wurden die Gewebeblöcke in ein Gemisch aus zwei Teilen
Propylenoxid und einem Teil Epon gelegt. Nachdem sie zweimal 2 h lang mit Epon
inkubiert worden waren, wurden sie eingebettet und polymerisierten bei 55°C - 65°C 48
h lang aus. So behandeltes Gewebe wurde für die Lichtmikroskopie an
Semidünnschnitten (3.7) und für die elekronenmikroskopische Beurteilung der
Ultrastruktur des Riechepithels (3.10) verwendet.
Gefriergetrocknetes Gewebe: Das gefriergetrocknete Gewebe wurde mit Epon
infiltriert und wie oben beschrieben polymerisiert. Gefriergetrocknetes und in Epon
eingebettetes Gewebe wurde für die Immunfluoreszenzuntersuchungen an
Semidünnschnitten verwendet.
3.4 Trimmen
Bevor von dem eingebetteten Gewebe (3.2; 3.3) Semi- (3.5) oder Ultradünnschnitte
(3.6) angefertigt werden konnten, mußte das quader- oder zylinderförmige
Harzblöckchen an einer seiner kleinen Flächen unter einer Lupenlampe mit einer
Rasierklinge eben angeschnitten werden, bis Gewebe auf der Oberfläche erschien.

Methoden 22
3.5 Semidünnschnitte
Das getrimmte Gewebeblöckchen wurde in eine Schneidemaschine für
Semidünnschnitte (2.1) eingespannt. Es wurden 1 µm dicke Schnitte angefertigt. Diese
wurden in einen Tropfen destillierten Wassers auf einem Deckglas gegeben und auf
einer Heizplatte 3 h lang getrocknet.
3.6 Lichtmikroskopie
Um einen schnellen Überblick über die Struktur eines Gewebeschnittes zu bekommen,
wurden Semidünnschnitte (3.5) mit Methylenblau überschichtet und ca. 30 s auf einer
Heizplatte angefärbt. Nachdem der Farbstoff mit PBS (2.3) abgewaschen worden war,
wurden die Präparate getrocknet und mit Entellan eingedeckt.
3.7 Ultradünnschnitte
Das getrimmte Gewebeblöckchen (3.4) wurde in eine Schneidemaschine für
Ultradünnschnitte (2.1) eingespannt. Die ca. 100 nm dicken Schnitte wurden mit Hilfe
eines Wimpernhaares auf ein mit Formvar beschichtetes Nickelnetzchen aufgespannt.
Anschließend wurden sie luftgetrocknet.
3.8 Kontrastierung von Ultradünnschnitten
Ultradünnschnitte (3.7) von in LR-White (3.2) oder Epon (3.3) eingebetteten Präparaten
werden kontrastiert, um Gewebestrukturen sichtbar zu machen. Dazu wurde eine
Glasplatte mit Parafilm bezogen, worauf entsprechend der Anzahl der Netzchen ein
Tropfen 2 % (w/v) Uranylacetatdihydrat in H2O gegeben wurde. Jedes Präparat wurde
mit der Schnittseite nach unten 20 min lang auf einen dieser Tropfen gelegt.
Anschließen wurden die Netzchen mit destilliertem Wasser gewaschen und mit
Filterpapier vom Rand her vorsichtig getrocknet. Nun wurden sie für 7 min in einem
Tropfen 0,2 % (w/v) Bleizitrat inkubiert. Nach erneutem Spülen mit destilliertem

Methoden 23
Wasser und Absaugen der Flüssigkeit mit Filterpapier wurden die Ultradünnschnitte
luftgetrocknet.
3.9 Immun-Elektronenmikroskopie
Ultradünnschnitte (3.7) von in LR-White eingebettetem Gewebe (3.2) wurden 15 min
lang in je einem Tropfen 0,05 M Glycin-Puffer in PBS inkubiert. Daraufhin wurden sie
für 30 min in einen Blockpuffer gegeben. Dreimal 5 min lang wurden sie in
Inkubationspuffer gewaschen. Die Netzchen wurden mit 20 µl in PBS verdünnten
Primär-Antikörpers (2.4) in einer feuchten Kammer bei 4°C über Nacht gelagert.
Nachdem sie für sechsmal 5 min mit Inkubationspuffer gespült worden waren, wurden
sie in 20 µl des in PBS verdünnten Sekundär-Antikörpers (2.4) in einer feuchten
Kammer 2 h lang bei Raumtemperatur inkubiert. Die Präparate wurden erneut für
sechsmal 5 min mit Inkubationspuffer gewaschen. Sie wurden dreimal 5 min lang in
PBS (2.3) gelegt, bevor sie für 5 min mit 2 % (v/v) Glutaraldehyd in PBS postfixiert
wurden. Sie wurden 5 min lang in PBS und für fünfmal 2 min in Ampuwa gespült.
Die verwendeten Sekundär-Antikörper waren mit Goldpartikeln gekoppelt, welche
unterschiedlich groß waren, 10 nm, 5 nm und 0,8 nm. 0,8 nm große sogenannte
Ultrasmall-2.-Antikörper mußten versilbert werden (3.10), um unter dem
Elektronenmikroskop sichtbar zu werden. Dieses wurde durchgeführt, bevor alle
Präparate kontrastiert (3.8) wurden.
Blockpuffer:
5 % (v/v) BSA-c, 0,1 % (v/v) CWFS, 5 % (v/v) NGS, in PBS
Inkubationspuffer:
1 % (v/v) BSA-c, 0,05 % (v/v) NaN3 in PBS
3.10 Silberverstärkung nach Danscher
Die wie unter 3.7 beschrieben vorbereiteten Ultradünnschnitte wurden lichtgeschützt in
je einen Tropfen der Gebrauchslösung zur Silberverstärkung gelegt. Nach sechsmal 5
min waschen in Ampuwa wurden die Präparate 45 min lang in Ampuwa belassen.

Methoden 24
Anschließend wurden sie mit destilliertem Wasser gespült und getrocknet. Sie konnten
nun kontrastiert (3.8) werden.
Gebrauchslösung:
60 % (v/v) Lösung A, 10 % (v/v) Lösung B, 15 % (v/v) Lösung C, 15 % (v/v) Lösung D
Lösung A = Schutzkolloid :
1 % (w/v) Gummi Arabicum in Aqua dest.
Lösung B = Puffer:
25 % (w/v) Zitronensäure, 23,6 % (w/v) Natriumzitrat in Aqua dest.
Lösung C = Entwickler:
5,7 % (w/v) Hydrochinon in Aqua dest.
Lösung D = Silbersalzlösung:
7,2 % (w/v) Silberlaktat in Aqua dest.
3.11 Herauslösen von Epon aus Semidünnschnitten von gefriergetrocknetem Gewebe
Semidünnschnitte (3.5) von gefriergetrocknetem, in Epon eingebettetem Gewebe
wurden der Reihe nach mit Na-Methanolat (6 min), Methanol/Toluol (5 min), Aceton
(zweimal 5 min), destilliertem Wasser (5 min) und PBS (5 min) (2.3) behandelt
(Drenckhahn et al., 1986). Nun war das nachfolgende Inkubationsschritte der
Fluoreszenzimmunhistochemie (3.12) störende Epon-Harz (3.3) herausgelöst. Die
Präparate wurden bis zur Weiterbehandlung in PBS belassen.
3.12 Fluoreszenzimmunhistochemie
Bei der Fluoreszenzimmunhistochemie wird ein bestimmtes Protein mit Hilfe eines
spezifischen 1.-Antikörpers (2.4) detektiert. An dessen FC-Abschnitt bindet ein 2.-
Antikörper (2.4), der mit einem fluoreszierenden Agens gekoppelt ist. Dieses leuchtet
bei Bestrahlung mit Licht einer entsprechenden Wellenlänge auf. Somit können unter
dem Fluoreszenzmikroskop die gesuchten Strukturen auf dunklem Grund sichtbar
gemacht werden.

Methoden 25
Nachdem das Epon aus den Semidünnschnitten herausgelöst worden war (3.11), wurden
die Deckgläser kurz in destilliertem Wasser gespült und mit ca. 20 µl des mit PBS (2.3)
verdünnten 1.-Antikörpers bedeckt. Die Präparate wurden bei 4°C über Nacht in einer
feuchten Kammer inkubiert (Goldman, 1963). Anschließend wurde sechsmal 5 min lang
mit PBS gespült und 20 µl des mit PBS verdünnten 2.-Antikörpers aufgetragen. Die
Gewebeschnitte wurden so in einer feuchten Kammer für 3 h bei Raumtemperatur
gelagert. Erneut wurde sechsmal 5 min lang mit PBS gewaschen. Die Deckgläser
wurden mit einem Papiertuch getrocknet, ohne das Gewebe zu beschädigen. Nun wurde
ein Tropfen 1,5 % (w/v) N-Propylgallat in 60 % Glyzerin in H2O auf einen Objektträger
gegeben und das Präparat mit der leeren Seite nach außen luftblasenfrei darauf gelegt.
3.13 Herstellung eines Peptidantikörpers
Injiziert man Fremdprotein in die Blutbahn eines Tieres, produziert es bei intaktem
Immunsystem dagegen gerichtete wirtsspezifische Antikörper. Diese können nach
Aufreinigung des entnommenen Blutes für immunhistochemische Versuche verwendet
werden.
3.13.1 Koppelung von Peptid an KLH
Da anzunehmen ist, dass das eingesetzte Peptid (2.6) aufgrund seiner geringen Größe
im Wirtstier nicht immunogen wirkt, wurde es als Hapten an KLH gekoppelt.
Die folgenden Angaben beziehen sich auf 1 mg Peptid. 1 mg Peptid wurde mit 1 mg
KLH in 967,6 µl PBS (2.3) auf Eis gelöst. 1 ml 2,5 % (v/v) Glutaraldehyd wurde
langsam hinzugetropft. Der Ansatz wurde 1 h lang bei 4°C inkubiert. Anschließend
wurden 10 mg Na-Borhydrid dazugegeben und erneut für 1 h inkubiert. Um
Verunreinigungen zu entfernen,) wurde die Lösung gegen PBS dialysiert (3.13.3).
3.13.2 Aufbereitung von Dialyseschläuchen
Ein Dialyseschlauch wurde nach Bedarf zugeschnitten und in einem Erlenmeyerkolben
in destilliertem Wasser mit einer Spatelspitze EDTA 3 min lang aufgekocht. Dieses

Methoden 26
wurde zweimal mit frischem Medium wiederholt. Die Schläuche konnten nun eingesetzt
werden. Sie konnten auch nach Zugabe von 0,05 % (w/v) NaN3 in PBS (2.3) bei 4°C
aufbewahrt werden.
3.13.3 Dialyse der Lösung von KLH-gekoppeltem Peptid gegen PBS
Ein entsprechend vorbereiteter (3.13.2), mit der Lösung von KLH-gekoppeltem Peptid
(3.13.1) gefüllter und an seinen Enden mit Klemmchen verschlossener Dialyseschlauch
wurde bei 4°C gegen drei Liter PBS (2.3) dialysiert. Dies erfolgte über 48 h, wobei PBS
am ersten Tag viermal, am zweiten Tag zweimal gewechselt wurde. Die dialysierte
Lösung wurde nun in ein Plastikreaktionsgefäß überführt und bis zu ihrer Verwendung
bei –20°C gelagert.
3.13.4 Immunisierung eines Kaninchens
Um die Antikörperproduktion und –gewinnung so effektiv wie möglich zu gestalten,
erfolgte die Immunisierung des Wirtstieres nach einem genau vorgegebenen Zeitplan.
Zunächst wurde dem unbehandelten Kaninchen Blut aus der Ohrvene abgenommen, um
sogenanntes Präimmunserum zu erhalten. Damit prüfte man unspezifische Antigen-
Antikörper-Reaktionen mit dem Zielgewebe, die nicht mit der spezifischen
Immunisierung zusammenhingen.
Drei Wochen nach der subkutanen Injektion einer Immunisierungslösung wurde eine
Boosterlösung subkutan injiziert. Zwei Wochen später wurde aus einer Ohrvene Blut
abgenommen, das Serum wurde analysiert (3.13.5). Es schloß sich ein Rhythmus aus
einer Boosterung nach einer Woche und einer Blutentnahme zwei Wochen später an,
der mehrmals wiederholt wurde.
Immunisierungslösung:
20 % (v/v) PAO compl. (Fluka, Hannover), 30 % (v/v) PBS, 50 % (v/v) Peptidlösung
Boosterlösung:
20 % (v/v) PAO incompl. (Fluka, Hannover), 80 % (v/v) Peptidlösung

Methoden 27
3.13.5 Aufbereitung und Testung des Immunserums
Das entnommene Blut wurde zentrifugiert (Tischzentrifuge 5415 C, 14000 g, 10 min,
Raumtemperatur) und das Serum abpipettiert. In einer Verdünnung von 1:10 in PBS
(2.3) wurde es auf Test- und Kontrollgewebe aufgetragen (3.8) und somit der Erfolg der
Immunisierung überprüft. Positives Serum wurde in Plastikreaktionsgefäße gefüllt und
bei –20°C aufbewahrt.
3.13.6 Affinitätsreinigung des Immunserums
Die Affinitätsreinigung diente der Extraktion hochaffiner Peptid-Antikörper aus dem
Serum des Kaninchen. Dazu wurde natives Peptid (2.8) mit Hilfe des Dot-Blots auf
einer Nitrozellulosemembran (2.1) immobilisiert und mit Immunserum (3.13.5)
inkubiert, das im Verhältnis 1:30 mit PBS (2.3) verdünnt worden war. Durch Waschen
mit PBS wurden verunreinigende Bestandteile entfernt, so daß nur mehr Antigen-
Antikörper-Komplexe (= natives Peptid-Peptidantikörper-Komplexe), die an der
Nitrozellulosemembran hafteten, verblieben. Nach Hitzeablösung wurde der
Peptidantikörper ausgetestet (3.12) und bei –20°C gelagert.
Zunächst wurde eine Nitrozellulosemembran (2.1) mit PBS (2.3) befeuchtet und in ein
Vakuumblot-Gerät (2.1) eingespannt, dessen Vertiefungen mit PBS und Peptid-PBS-
Lösung in einer Konzentration von 1 mg/ ml beschickt wurden. Nach Anschließen einer
Wasserstrahlpumpe floß das Gemisch über die Nitrozellulosemembran ab, wobei das
Peptid haften bliebt. Es wurde zweimal mit PBS nachgespült. Anschließend wurde die
Nitrozellulosemembran in Ponceau S angefärbt. Der Farbstoff, der Peptid markiert,
wurde mit PBS entfernt. Unspezifische Proteinbindungsstellen wurden über Nacht in
einer Magermilchlösung abgesättigt. Es wurde dreimal 5 min lang mit 0,05 % (v/v)
Tween 20 in PBS gewaschen. Die so präparierte Nitrozellulosemembran konnte in 0,05
% (w/v) NaN3 in PBS bei 4°C gelagert werden.
Ponceau S-Lösung:
0,5 % (w/v) Ponceau S; 3 % (w/v) Trichloressigsäure in H2O

Methoden 28
Magermilchlösung:
5 % (w/v) Magermilch; 0,05 % (v/v) Tween 20 in PBS
Die vorbereitete Nitrozellulosemembran wurde in ein sialinisiertes Glasgefäß gegeben
und zweimal mit PBS (2.3) gewaschen. Daraufhin wurde sie mit Immunserum (3.13.5),
das im Verhältnis 1:30 mit PBS verdünnt worden war, bei 4°C über Nacht auf dem
Schüttler inkubiert. Es wurde erneut zweimal mit PBS gespült. 450 µl PBS wurden
hinzugefügt. Im 55,6°C-Wasserbad lösten sich nach 10 min die Peptidantikörper von
den auf der Nitrozellulosemembran fixierten nativen Peptiden. Die Flüssigkeit wurde
sofort in ein frisches silaniertes Plastikreaktionsgefäß (2.1) abgehoben, um die erneute
Ausbildung von Antigen-Antikörper-Komplexen bei niedrigeren Temperaturen zu
verhindern. Die Konzentration des Peptidantikörpers konnte photometrisch gegen den
Leerwert PBS bei einer Wellenlänge λ=280 nm bestimmt werden (E 1,35 = 1 mg/ ml
IgG). In einer Verdünnung von 1:10 mit PBS wurde Test- und Kontrollgewebe
inkubiert (3.12). Der gereinigte Peptidantikörper wurde bei –20°C aufbewahrt.
3.14 Amidoschwarz-Proteinbestimmung (Dieckmann-Schuppert et al., 1997)
Eine Zellulose-Azetat-Folie (2.1) wurde in ca. 1 cm2 große Felder eingeteilt. Maximal
10 µl einer Verdünnung definierter Mengen von BSA (Standardproben) und der zu
analysierenden Proteinproben, welche in SDS-Probenpuffer gelöst waren, wurden auf
jeweils zwei Bereiche aufgetragen und in einem kalten Luftstrom getrocknet. Die Folie
wurde auf einem Schüttler zunächst 10 min lang mit Färbelösung, anschließend mit
Entfärber behandelt, bis der Hintergrund weiß erschien. Nachdem sie in einem kalten
Luftstrom getrocknet worden war, wurden die einzelnen Felder ausgeschnitten und in
beschriftete Plastikreationsgefäße (2.1) gegeben. Unter Zugabe einer Auflöselösung
löste sich die Folie im 50°C-Wasserbad nach 30 min auf. Die durchmischten Proben
wurden in Plastikküvetten umgefüllt und bei einer Wellenlänge von 620 nm gegen den
Leerwert Probenpuffer gemessen. Mit Hilfe der Standardproben wurde eine Eichkurve
erstellt, aus der der Gehalt der Proteinproben abgelesen werden konnte.
Auflöselösung:
80 % (v/v) Ameisensäure; 10 % (v/v) Essigsäure; 10 % (v/v) Trichloressigsäure

Methoden 29
Entfärbelösung:
47,5 % (v/v) Methanol; 47,5 % (v/v) Aqua dest.; 5 % (v/v) Eisessig
Färbelösung:
0,5 % (w/v) Amidoschwarz; % (v/v) Methanol; 45 % (v/v) Aqua dest.; 10 % (v/v) Eisessig
2-fach SDS-Probenpuffer:
250 mM Tris/HCl (pH 6,8); 4 % (w/v) SDS; 20 % (v/v) Glycerin; 100 mM Dithiothreitol; 0,16 % (w/v)
Bromphenolblau in H2O
3.15 Bradford-Proteinbestimmung
Jeweils 20 bis 100 µl der Leerwert-, BSA-Standard- und Proteinproben wurden in einer
Plastikküvette mit 1 ml des Bradford-Reagenz 10 min lang bei Raumtemperatur
inkubiert. Bei einer Wellenlänge von 595 nm erfolgte die photometrische Auswertung
in der Reihenfolge Leerwertprobe – BSA-Standard-Proben in absteigender
Konzentration – Proteinproben. Mit Hilfe der Leerwert- und BSA-Standard-Proben
wurde eine Eichgerade erstellt, mit der der Gehalt der Proteinproben ermittelt werden
konnte.
Bradford-Reagenz:
20 % (v/v) Protein Assay Cat. 500-0006 in Aqua dest.
BSA-Standard-Proben:
1 %/ 0,75 %/ 0,5 %/ 0,25 %/ 0,1 % (w/v) BSA in PBS
Leerwertprobe:
PBS (2.3)
Proteinproben:
1 % (v/v) Proteinlösung in PBS und 0,25 % (v/v) Proteinlösung in PBS
3.16 SDS-Polyacrylamid-Gelelektrophorese (SDS-Page) (Laemmli, 1970)
Die SDS-Polyacrylamid-Gelelektrophorese wurde nach dem Verfahren von Laemmli
(1970) durchgeführt. Dabei erfolgt aufgrund der denaturierenden Eigenschaften von

Methoden 30
Natrium-Dodecylsulfat (SDS) die Auftrennung der Proteine im elektrischen Feld allein
nach ihrem Molekulargewicht.
Das SDS-Polyacrylamidgel wurde zwischen zwei senkrechte Glasplatten gegossen (9
cm x 6 cm x 0,8 cm). Es setzte sich aus einem Trenngel und einem Sammelgel
zusammen. Die Polyacrylamidkonzentration des Trenngels betrug 7,5 % (w/v), die des
Sammelgels 5 % (w/v).
Die Proteinproben (Proteinmenge 80 µg) wurden 5 min bei 95°C erhitzt und auf das Gel
aufgetragen. Die Proteinauftrennung erfolgte bei einer Spannung von 80 V im
Sammelgel und von 200 V im Trenngel. Die Elektrophorese wurde beendet, wenn die
Lauffront des Bromphenolblaus (SDS-Probenpuffer) das Ende des Gels erreicht hatte.
Anschließend wurden die Proteine auf Nitrozellulose elektrotransferiert (3.17).
7,5 % Trenngel:
7,275 % (w/v) Acrylamidlösung (30 % {w/v}[Roth, Karlsruhe]); 0,225 % (w/v) Bisacrylamidlösung (2 %
{w/v}[Roth, Karlsruhe]); 6,75 % (v/v) Glycerin; 375 mM Tris/HCl (pH 8,8); 0,1 % (w/v) SDS; 0,05 %
(v/v) TEMED; 0,05 % (w/v) Ammoniumperoxodisulfat (APS)
5 % Sammelgel:
4,85 % (w/v) Acrylamidlösung (30 % {w/v}[Roth, Karlsruhe]); 0,15 % (w/v) Bisacrylamidlösung (2 %
{w/v}[Roth, Karlsruhe]); 125 mM Tris/HCl (pH 6,8); 0,1 % (w/v) SDS; 0,2 % (v/v) TEMED; 0,1 %
(w/v) Ammoniumperoxodisulfat (APS)
2-fach SDS-Probenpuffer:
250 mM Tris/HCl (pH 6,8); 4 % (w/v) SDS; 20 % (v/v) Glycerin; 100 mM Dithiothreitol; 0,16 % (w/v)
Bromphenolblau
Elektrophoresepuffer, pH 8,3:
25 mM Tris; 172 mM Glycin; 0,1 % (w/v) SDS
3.17 Immunreplikanalyse (Western-Blot)
Nach der Auftrennung der Proteine in der SDS-Polyacrylamid-Gelelektrophorese
wurden die Proteine mit Hilfe des Semi-dry-Verfahrens auf eine Nitrozellulosemembran
(2.1) übertragen (Burnette, 1981). Dazu wurden Gel und Nitrozellulosemembran

Methoden 31
zwischen puffergetränkte Filterpapiere geschichtet und luftblasenfrei auf die
Graphitelektroden einer Laufkammer gelegt.
Der Transfer der Proteine erfolgte 90 min lang bei einer Stromstärke von 0,8 mA pro 1
cm2 SDS-Polyacrylamidgel. Bei einer Größe des Minigels von 9 cm x 6 cm entsprach
dies einer Stromstärke von 43 mA. Der Erfolg des Transfers wurde überprüft, indem die
Proteinbanden reversibel durch Ponceau S-Lösung rot angefärbt wurden (Salinovich et
al., 1986). Die spätere Immunfärbung der Proteinproben wurde dadurch nicht
beeinträchtigt. Die mit Ponceau S angefärbte Laufspur der Eichproteine (EPH-Marker,
2.5) wurde abgetrennt und getrocknet.
Anodenpuffer I:
0,3 M Tris; 20 % (v/v) Ethanol in H2O
Anodenpuffer II:
25 mM Tris; 20 % (v/v) Ethanol in H2O
Kathodenpuffer:
25 mM Tris; 0,725 % (w/v) Aminocapronsäure; 20 % (v/v) Ethanol in H2O
Ponceau S-Lösung:
0,5 % (w/v) Ponceau S; 3 % (w/v) Trichloressigsäure in H2O
Die Nitrozellulosestreifen wurden zunächst in einer Magermilchlösung bei 4°C über
Nacht inkubiert. Hierdurch wurden unspezifische Proteinbindungsstellen abgesättigt.
Nach Entfernen der Magermilchlösung und Waschen mit 0,05 % (v/v) Tween 20 in
PBS wurden die Nitrozellulosemembranen bei 4°C über Nacht mit einem Erstantikörper
(2.4) auf einem Schüttler inkubiert. Dieser wurde in Magermilchlösung nach Angaben
des Herstellers verdünnt. Nach Entfernung des ungebundenen Primärantikörpers mit
0,05 % (v/v) Tween 20 in PBS (dreimal 5 min) folgte die Inkubation mit dem
jeweiligen Zweitantikörper (2.4) in einer Verdünnung entsprechend den Angaben des
Herstellers in Magermilchlösung bei Raumtemperatur für 90 min. Die
Nitrozellulosestreifen wurden anschließend wie oben beschrieben gewaschen und für
die ECL-Entwicklung eingesetzt.

Methoden 32
Magermilchlösung:
5 % (w/v) Magermilch; 0,05 % (v/v) Tween 20 in PBS
Die an den Zweitantikörper gekoppelte Meerrettichperoxidase (POX) katalysiert unter
Reduktion von H2O2 die Redox-Reaktion des ECL-Systems, in deren Verlauf Luminol
zur Chemolumineszenz angeregt wird. Luminol kehrt direkt nach seiner Oxidation aus
seinem angeregten Zustand in den Grundzustand zurück und emittiert dabei Photonen,
die mit einem Röntgenfilm (2.1) nachgewiesen werden können.
Die Nitrozellulosemembranen wurden für 1 min in einer frisch angesetzten ECL-
Lösung (ECL I und II im Verhältnis 1:1) bei Raumtemperatur inkubiert und
luftblasenfrei in eine Klarsichtfolie eingeschlagen. Anschließend wurden sie in einer
Filmkassette auf einem Röntgenfilm (Hyperfilm ECL) je nach Intensität des Signals 10
s bis 60 s exponiert. Nach Sichtbarwerden der Banden (Schwarzfärbung) wurde der
Röntgenfilm zur Dokumentation entwickelt, fixiert und getrocknet.
ECL I-Lösung:
2,5 mM Luminol (Fluka, Deisenhofen); 0,04 mM p-Coumaric-Acid (Sigma, Deisenhofen); 0,1 M
Tris/HCL (pH 8,5)
ECL II-Lösung:
0,02 % (w/v) H2O2; 0,1 M Tris/HCl (pH 8,5)

Ergebnisse 33
4. Ergebnisse
Die Zellen des olfaktorischen Epithels bilden typische epitheliale Kontakte aus. Hierzu
gehören Zonula occludens, Zonula adherens, Macula adherens und Punctum adherens.
Zonula occludens, Zonula adherens und Macula adherens bilden als Haft- oder
Schlußleistenkomplex die Grenzstruktur zwischen apikaler und basolateraler
Plasmamembran. Puncta adherentia sind lateral lokalisiert.
4.1 Darstellung von Proteinen der Zonula occludens im olfaktorischen Epithel
Durch Darstellung von ZO-1 in einem tangentialen Semidünnschnitt durch das apikale
olfaktorische Epithel wurde ein typisches Muster der apikalen Zonulae occludentes
sichtbar (Abb. 4.1.1). Es entsprach der in der Einleitung beschriebenen vermuteten
Topographie der apikalen Zellgrenzen. Danach markieren polygonale Formen Stützzell-
Stützzell-Kontakte, größere Kreise Stützzell-Mikrovillizell-Kontakte und kleinere
Kreise Stützzell-Neuron-Kontakte. Die Intensität der Immunfluoreszenz für ZO-1 war
in allen Zellkontakten gleich stark.
Abb. 4.1.1: ZO-1 Immunreaktivität in einem apikalen Tangentialschnitt durch das
olfaktorische Epithel.
Die ZO-1-Immunreaktivität stellt die apikalen Zonulae occludentes dar.
Polygonale Formen: Stützzell-Stützzell-Kontakte (Pfeilspitze); größere Kreise:

Ergebnisse 34
Stützzell-Mikrovillizell-Kontakte (großer Pfeil); kleinere Kreise: Stützzell-
Neuron-Kontakte (kleiner Pfeil); Maßstab: 10 µm.
Durch Behandlung von Folgeschnitten mit Antikörpern gegen Occludin und ZO-1
wurde deutlich, daß die Lokalisation von Occludin und ZO-1 sehr ähnlich war.
Allerdings kamen die verschiedenen Zellkontakte nicht in der gleichen
Immunfluoreszenzintensität zur Darstellung (Abb. 4.1.2 und 4.1.3): Die
Immunfluoreszenz für Occludin in Stützzell-Neuron-Kontakten (kleine Kreise) war
geringer als in Stützzell-Stützzell-Kontakten (polygonale Formen) (Abb. 4.1.2).
Abb. 4.1.2: Occludin (links) und ZO-1 (rechts) in tangentialen Folgeschnitten durch das
olfaktorische Epithel. Maßstab: 5 µm.
In mit Occludin (grün) und ZO-1 (rot) doppelmarkierten longitudinalen
Semidünnschnitten war die relativ intensivste Occludin- und ZO-1-Immunreaktivität in
den gelb-orange fluoreszierenden Zellkontakten zwischen Ausführungsgangszellen zu
erkennen (Abb. 4.1.3).

Ergebnisse 35
Abb. 4.1.3: Occludin- (grün) und ZO-1-Immunreaktivität (rot) in einem
Longitudinalschnitt durch das olfaktorische Epithel. Maßstab: 10 µm.
Tabelle 4.1.1 zeigt zusammenfassend die Immunreaktivitätsintensität der beiden
Proteine in Zonulae occludentes zwischen verschiedenen Zellen des olfaktorischen
Epithels.
Kontakt Occludin ZO-1
Stützzelle-Stützzelle ++ ++
Stützzelle-Mikrovillizelle ++ ++
Ausführungsgangszelle-
Ausführungsgangszelle
+++ ++
Stützzelle-Neuron + ++
Tab. 4.1.1: Vergleich der Immunfluoreszenzintensität von Occludin und ZO-1 in
Zonulae occludentes zwischen verschiedenen Zelltypen des olfaktorischen
Epithels.
4.2 Darstellung von Proteinen der Zonula adherens im olfaktorischen Epithel
Das typische Transmembranprotein epithelialer Adhärenskontakte ist E-Cadherin. Ein
typisches neuronales klassisches Cadherin ist N-Cadherin, das auch das Cadherin der

Ergebnisse 36
Adhärenskontakte des Herzmuskels ist. Adhärenskontakte enthalten neben
Transmembranproteinen unter anderem die zytoplasmatischen Plaqueproteine α-
Catenin, β-Catenin und α-Actinin.
Während der Herzmuskel kein E-Cadherin enthält, besitzen Epithelien wie
beispielsweise das Dünndarmepithel kein N-Cadherin.
Lokalisation von E-Cadherin im olfaktorischen Epithel
Um zu prüfen, ob E-Cadherin im olfaktorischen Epithel vorkommt, wurde eine
Immunreplikanalyse durchgeführt (3.17). Dabei wurden Gewebeproben des
olfaktorischen Epithels (Positivprobe) und des Herzmuskels (Negativprobe) mit dem
Primär-Antikörper E-Cadherin und dem Sekundär-Antikörper Gam-HRP-POX
IgG/IGM inkubiert (Abb. 4.2.1).
Abb. 4.2.1: Nachweis von E-Cadherin im olfaktorischen Epithel.
Herzmuskel-Gewebe (HM), das kein E-Cadherin exprimiert, diente als Negativprobe.
Die Bande in der Spur des olfaktorischen Epithels bei 120 kDa entsprach der
molekularen Masse von E-Cadherin. Im Herzmuskel (Negativprobe) wurde kein Protein
detektiert. Damit waren die Spezifität des Antikörpers für E-Cadherin in der
Immunreplikanalyse sowie die Anwesenheit von E-Cadherin im olfaktorischen Epithel
nachgewiesen.

Ergebnisse 37
Ein longitudinaler (Abb. 4.2.2) und ein tangentialer (Abb. 4.2.3) Semidünnschnitt der
Riechschleimhaut wurde mit dem Primär-Antikörper E-Cadherin und dem Sekundär-
Antikörper Gam-Cy3 inkubiert.
Abb. 4.2.2: Einfachmarkierung (oben) für E-Cadherin (grün) und Doppelmarkierung (unten)
für E-Cadherin (grün) und Na+/K+-ATPase (rot) in einem Longitudinalschnitt
durch das olfaktorische Epithel. Maßstab: 5 µm.
In Longitudinalschnitten (Abb. 4.2.2) waren im gesamten olfaktorischen Epithel kräftig
fluoreszierende längsverlaufende Streifen zu erkennen, die im Bereich der luminalen
Oberfläche in einem noch intensiver fluoreszierenden Punkt endeten.
Doppelmarkierungen für Na+/K+-ATPase, dem Markerprotein für Membranen von
olfaktorischen Neuronen und Mikrovillizellen, zeigten, daß die Immunreaktivität von E-
Cadherin im Bereich der apikalen und lateralen Stützzell-Stützzell-, Stützzell-
Mikrovillizell- und Stützzell-Ausführungsgangszell-Kontakte lokalisiert waren und
damit wahrscheinlich die Zonulae adherentes sowie die Puncta adherentia zwischen
diesen Zellen markierten.
Zellkontakte zwischen Stützzellen und Sinneszellen, deren Membranen relativ stark
Na+/K+-ATPase-reaktiv waren, zeigten eine außerordentlich schwache E-Cadherin-
Imunfluoreszenz (Abb. 4.2.2).

Ergebnisse 38
Tangentialschnitte bestätigten die kräftige E-Cadherin-Immunreaktivität der Stützzell-
Stüzzell-Kontakte (polygonale Formen).
Abb. 4.2.3: E-Cadherin-Immunreaktivität im olfaktorischen Epithel.
Schräger Tangentialschnitt, der den Bereich der Zonulae adherentes (Pfeil) und
der Puncta adherentia (Pfeilspitze) zwischen Stützzellen zeigt. Maßstab: 5 µm.
Um eine Kreuzreaktivität des E-Cadherin-Antikörpers mit anderen eventuell im
olfaktorischen Epithel vorhandenen Cadherinen auszuschließen, wurden ein
Semidünnschnitt des Dünndarmepithels (Positivprobe) und des Herzmuskels
(Negativprobe) mit dem Primär-Antikörper gegen E-Cadherin und dem Sekundär-
Antikörper Gam-Cy3 inkubiert (keine Abbildung; 3.12). Das Bild des für E-Cadherin
positiven und N-Cadherin negativen Dünndarmepithels zeigte deutlich markierte
Zellgrenzen, während im E-Cadherin negativen und N-Cadherin positiven Herzmuskel
keine immunhistochemische Reaktion nachzuweisen war. Auch für diese Methode war
damit die Spezifität des Antikörpers für E-Cadherin nachgewiesen.
Zur detaillierten ultrastrukturellen Lokalisation wurde ein Ultradünnschnitt der
Riechschleimhaut mit dem Primär-Antikörper E-Cadherin und dem Sekundär-
Antikörper Gam-US inkubiert und durch Versilberung verstärkt (Abb. 4.2.4; 3.10). Es
fand sich eine Lokalisation der Goldkonjugate im Bereich von Zonulae adherentes
ausschließlich zwischen Stützzellen und Stützzellen, nicht zwischen Stützzellen und
Dendriten der Sinneszellen.

Ergebnisse 39
Abb. 4.2.4: E-Cadherin–Immunogoldlokalisierung im olfaktorischen Epithel.
Inkubation eines longitudinalen (links) und eines tangentialen (rechts) Ultradünnschnitts
des olfaktorischen Epithels mit E-Cadherin-Antikörper und Gam-US. D: Dendrit; E-
cad: E-Cadherin; SC: Stützzelle (supporting cell); Maßstab: 200 nm.
Die Ergebnisse der Untersuchungen zur Lokalisation von E-Cadherin im olfaktorischen
Epithel belegen, dass E-Cadherin das Transmembranprotein der Adherenskontakte
zwischen den epithelialen Zellen ist.
Das Cadherin der Neurone im olfaktorischen Epithel
Da E-Cadherin offensichtlich nicht das Cadherin der Zonulae adherentes der Stützzell-
Sinneszell-Kontakte ist, wurde im nächsten Schritt untersucht, ob eventuell N-Cadherin
das neuronale Cadherin sein könnte. Zur Lokalisierung von N-Cadherin wurde zunächst
ein handelsüblicher Peptidantikörper eingesetzt (2.4). Dieser zeigte jedoch mehrfach in
immunhistochemischen Reaktionen weder im olfaktorischen Epithel noch im
Herzmuskel (Positivprobe) eine Reaktion. Zum Nachweis des Transmembranproteins
neuronaler Zellen wurden deshalb Pan-Cadherin-Antikörper (2.4), Antikörper gegen ein
N-Cadherin-Fusionsprotein (Fannon et al., 1996) und selbsthergestellte Antikörper
gegen ein spezifisches Peptid aus der Sequenz von N-Cadherin (2.6) eingesetzt.
Der Pan-Cadherin-Antikörper war gegen 24 Aminosäuren des C-Terminus von N-
Cadherin gerichtet. Da die Sequenz dieses Abschnitts hoch konserviert ist, sollte der
Antikörper mit verschiedenen Cadherinen unterschiedlicher Spezies reagieren.

Ergebnisse 40
Das Ergebnis einer Immunreplikanalyse mit dem Pan-Cadherin-Primär-Antikörper und
dem Sekundär-Antikörper Garb-HRP-Pox (3.17) zeigte Abb. 4.2.5. Die Spur des
Herzmuskels (Positivprobe) zeigte eine Bande bei 130 kDa, die Spur des olfaktorischen
Epithels wies eine etwas breitere und geringfügig schneller laufende Bande auf.
Abb. 4.2.5: Pan-Cadherin- und E-Cadherin-Immunreaktivität im olfaktorischen
Epithel.
Immunreplikanalyse des olfaktorischen Epithels (OE) und des Herzmuskels
(HM) mit Pan-Cadherin-Antikörper und Garb-HRP-Pox sowie E-Cadherin-
Antikörper und Gam-HRP-POX.
N-Cadherin hat eine molekulare Masse von 130 kDa. Im Herzmuskel wurde
offensichtlich N-Cadherin mit Pan-Cadherin-Antikörper nachgewiesen. Im
olfaktorischen Epithel wurde durch Pan-Cadherin-Antikörper eine etwa gleich große
Bande markiert. Eine zusätzliche Bande mit der apparenten molekularen Masse von E-
Cadherin (120 kDa) war unter den gewählten Bedingungen nicht eindeutig abgrenzbar
(Abb. 4.2.5).
Ein longitudinaler (Abb. 4.2.6) und ein tangentialer (Abb. 4.2.7) Semidünnschnitt der
Riechschleimhaut wurde mit Primär-Antikörpern Pan-Cadherin/ β-Tubulin und den
Sekundär-Antikörpern Garb-Cy3/Gam-Cy2 inkubiert (3.12). Die Darstellung von β-
Tubulin diente der eindeutigen Identifizierung der Dendriten der Sinneszellen.

Ergebnisse 41
Abb. 4.2.6: Pan-Cadherin- (rot) und β-Tubulin-Immunreaktivtät (grün) in einem
Longitudinalschnitt durch das olfaktorische Epithel.
Maßstab: 5 µm.
Abbildung 4.2.6 zeigt apikal kräftig bzw. schwach fluoreszierende Punkte. Die β-
Tubulin-Immunreaktivität der Dendriten wurde apikal von der Pan-Cadherin-
Immunreaktivität begrenzt, dies deutete darauf hin, daß Pan-Cadherin-Reaktionen
Stützzell-Neuron-Kontakte markierten. Die polygonale Form der Pan-Cadherin-
Immunreaktivität zeigte, dass das durch Pan-Cadherin-Antikörper markierte Cadherin
auch in Stützzell-Stützzell-Kontakten lokalisiert war. Schwache Punkte im Bereich der
lateralen Zellgrenzen stellten Puncta adherentia dar. Basal nahm die Dichte der
Markierungen zu. Hier sind die Basalzellen gelegen.
Aufgrund der typischen Formen der unterschiedlichen Zonulae adherentes und der β-
Tubulin-Immunreaktivtät der Dendriten ließen sich im doppelimmunreagierten
Tangentialschnitt (Abb. 4.2.7) die Pan-Cadherin-Immunreaktivitäten den Stützzell-
Stützzell- und den Stützzell-Neuron-Kontakten zuordnen. Im Gegensatz zu
fluoreszenzimmunhistochemischen Reaktionen des olfaktorischen Epithels mit einem
Antikörper gegen E-Cadherin wurden Zellgrenzen zwischen Stützzellen und

Ergebnisse 42
Mikrovillizellen (größere Kreise im Tangentialschnitt), Kontakte im Bereich der
Ausführungsgänge der Bowmanschen Drüsen (longitudinale Linien im
Longitudinalschnitt) sowie Kontakte des respiratorischen Epithels nicht dargestellt.
Abb. 4.2.7: Pan-Cadherin- (rot) und β-Tubulin-Immunreaktivität (grün) in einem
Tangentialschnitt durch das olfaktorische Epithel.
Pfeil: Stützzell-Stützzell-Kontakt (polygonale Form); Pfeilspitze: Stützzell-Neuron-
Kontakt (kleiner Kreis); die β-Tubulin-Immunreaktivität markiert die Dendriten der
Sinneszellen. Maßstab: 5 µm.
Semidünnschnitte des Herzmuskelgewebes (Positivprobe) und Dünndarmepithels
(Negativprobe) wurden mit Pan-Cadherin-Antikörper inkubiert, um dessen relative
Spezifität gegenüber E-Cadherin in immunhistochemischen Untersuchungen zu
überprüfen (keine Abbildung; 3.12). Die Glanzstreifen des Herzmuskels stellten sich
deutlich dar, während im Dünndarmepithel, das große Mengen an E-Cadherin
exprimiert, keine Immunreaktivität nachzuweisen war. Unter den gewählten
Gewebebehandlungs- und Immunreaktionsbedinungen wurde E-Cadherin offenbar
durch Pan-Cadherin-Antikörper in situ praktisch nicht detektiert.
Um das durch Pan-Cadherin-Antikörper dargestellte Cadherin in den Zellen der
Riechschleimhaut genauer lokalisieren zu können, wurde ein Ultradünnschnitt mit dem
Pan-Cadherin-Antikörper und dem Sekundär-Antikörper Gam-US inkubiert und die
Goldpartikel durch Silberintensivierung verstärkt (Abb. 4.2.8; 3.10). Markierungen

Ergebnisse 43
fanden sich in den Stützzell-Dendrit- und, weniger dicht, in den Stützzell-Stützzell-
Kontakten.
Abb. 4.2.8: Pan-Cadherin-Immunreaktivität im olfaktorischen Epithel.
Inkubation von tangentialen Ultradünnschnitten des olfaktorischen Epithels mit Pan-
Cadherin-Antikörper (Pancad) und Gam-US. D: Dendrit; SC: Stützzelle (supporting
cell). Pfeile: Goldpartikel im Kontaktbereich zwischen Dendrit und Stützzelle;
Pfeilspitze: Goldpartikel im Kontaktbereich zwischen Stützzelle und Stützzelle.
Maßstab: 400 nm.
Das spezifische Lokalisationsmuster der Adherensproteine im olfaktorischen Epithel
wurde durch zusätzliche Untersuchungen von Doppelimmunreaktivitäten weiter
dokumentiert.
Longitudinale (Abb. 4.2.9) und schräge Semidünnschnitte (Abb. 4.2.10) wurden mit den
Primär-Antikörpern E-Cadherin und Pan-Cadherin sowie den Sekundär-Antikörpern
Gam-Cy3 und Garb-Cy2 inkubiert (3.12).

Ergebnisse 44
Abb. 4.2.9: E-Cadherin- (rot) und Pan-Cadherin-Immunreaktivität (grün) in einem
Longitudinalschnitt durch das olfaktorische Epithel.
Doppelinkubation mit E-Cadherin-/ Pan-Cadherin-Antikörper und Gam-Cy3/ Garb-Cy2
(rot/ grün). Maßstab: 10 µm.
Abbildung 4.2.9 zeigte apikal gelb fluoreszierende (doppelmarkierte) Streifen und
kleinere grüne (Pan-Cadherin-reaktive) bzw. größere rote (E-Cadherin-reaktive) Kreise,
welche den Zonulae adherentes zwischen Stützzellen und Stützzellen, Stützzellen und
Neuronen sowie Stützzellen und Mikrovillizellen entsprachen.
Abb. 4.2.10: E-Cadherin- (rot) und Pan-Cadherin-Immunreaktiviät (grün) in einem
Schrägschnitt durch das olfaktorische Epithel.

Ergebnisse 45
Doppelinkubation mit E-Cadherin-/ Pan-Cadherin-Antikörper und Gam-Cy3/ Garb-Cy2
(rot/ grün). Maßstab: 5µm.
Im Schrägschnitt (Abb. 4.2.10) waren apikal grün bzw. gelb und rot fluoreszierende
Halbkreise und Kreise zu sehen. Es handelte sich hierbei um die Zonulae adherentes
zwischen Stützzellen und Neuronen bzw. zwischen Stützzellen und Stützzellen sowie
zwischen Stützzellen und Mikrovillizellen.
Sowohl im Longitudinal- als auch im Schrägschnitt waren die E-Cadherin-reaktiven
lateralen Puncta adherentia zwischen Stützzellen und Mikrovillizellen deutlich als rote
Punkte erkennbar. Zudem waren grüne und gelbe punktförmige
Immunfluoreszenzmarkierungen zu erkennen, die den Puncta adherentia zwischen
Stützzellen und Neuronen bzw. zwischen Stützzellen entsprachen.
Ein wichtiger Nebenbefund dieser Untersuchungen war, dass apikal keine direkten
Kontakte zwischen einzelnen Neuronen, zwischen einzelnen Mikrovillizellen, zwischen
Neuronen und Mikrovillizellen, zwischen Neuronen und Ausführungsgangszellen oder
zwischen Mikrovillizellen und Ausführungsgangszellen zu beobachten waren. Sowohl
Neurone als auch Mikrovillizellen waren offensichtlich, zumindest in ihrem apikalen
Bereich, immer vollständig von Stützzellen eingehüllt. Dieser Befund wurde durch
elektronenmikroskopische Untersuchungen bestätigt (Abb. 1.1.4).
Abb. 4.2.11: E-Cadherin- (rot) und Pan-Cadherin-Immunreaktiviät (grün) in einem
Tangentialschnitt durch das olfaktorische Epithel.
Doppelinkubation mit E-Cadherin-/ Pan-Cadherin-Antikörper und Gam-Cy3/ Garb-Cy2
(rot/ grün). Pfeil: Ausführungsgang. Maßstab: 10 µm.

Ergebnisse 46
Intraepitheliale Schrägschnitte von Ausführungsgängen zeigen die erwartete
Immunfluoreszenz von E-Cadherin in Form größerer Kreise (Stützzell-
Ausführungsgangszell-Kontakt) und radiärer Streifen (Ausführungsgangszell-
Ausführungsgangszell-Kontakt)
Um die Zuordnung der Immunreaktivitäten zu den unterschiedlichen Zellkontakten zu
belegen, wurden Doppelinkubationen mit den Antikörpern gegen ZO-1 und Pan-
Cadherin durchgeführt. In Abb. 4.2.12 ist die differentielle Lokalisation der beiden
Proteine deutlich zu erkennen und belegt, daß die ZO-1-Immunfluoreszenz die apikalen
Zonulae occludentes, die Pan-Cadherin-Immunfluoreszenz die direkt darunter
gelegenen Zonulae adherentes markiert.
Abb. 4.2.12: ZO-1- (grün) und Pan-Cadherin-Immunreaktivität (rot) in einem
Longitudinalschnitt durch das olfaktorische Epithel.
Doppelinkubation mit ZO-1-/ Pan-Cadherin-Antikörper und Garat-Cy2/ Garb-Cy3
(grün/ rot). Maßstab: 5 µm.
Sowohl das apparente Molekulargewicht (130 kDa) als auch die Lokalisation
vorwiegend in neuronalen Kontakten deuteten darauf hin, dass es sich bei dem durch
Pan-Cadherin-Antikörper detektierten Cadherin um N-Cadherin handeln könnte. Dazu
wurden mit spezifischen Antikörpern weitere Untersuchungen durchgeführt.

Ergebnisse 47
In der Immunreplikanalyse (3.17) von Proteinen des olfaktorischen Epithels mit Hilfe
eines Antikörpers gegen ein Fusionsprotein aus der ersten extrazellulären Domäne von
N-Cadherin (Aminosäure 1-108; Dr. D.R. Colman) wurde ein Bandenmuster gefunden,
das in etwa dem der Immunreplikanalyse mit Pan-Cadherin-Antikörper entsprach.
Ein Schrägschnitt durch die Riechschleimhaut wurde mit dem Primär-Antikörper gegen
das N-Cadherin-Fusionsprotein und dem Sekundär-Antiköper Garb-Cy3 inkubiert (Abb.
4.2.13; 3.12). Die apikal gelegenen schwach fluoreszierenden Streifen bzw. Halbkreise
entsprachen den Zonulae adherentes zwischen Stützzellen und Stützzellen bzw.
zwischen Stützzellen und Sinneszellen. Puncta adherentia wurden nicht dargestellt.
Abb. 4.2.13: N-Cadherin-Immunreaktivität in einem Tangentialschnitt durch das
olfaktorische Epithel.
Inkubation mit N-Cadherin-Fusionsprotein-Antikörper und Garb-Cy3.
Maßstab: 10 µm.
Zum Ausschluß einer Kreuzreaktivität des Antikörpers gegen das N-Cadherin-
Fusionsprotein mit E-Cadherin wurden Semidünnschnitte des Herzmuskels
(Positivprobe; Abb. 4.2.14) und Vomeronasalorgans (Positivprobe; keine Abbildung)
sowie des Dünndarmepithels (Negativprobe; keine Abbildung) mit N-Cadherin-
Fusionsprotein-Antikörper und dem Sekundär-Antikörper Garb-Cy3 inkubiert (keine
Abbildung; 3.12). Im Herzmuskel und Vomeronasalorgan zeigten sich spezifische

Ergebnisse 48
Antigen-Antikörper-Reaktionen, im Dünndarmepithel wurden keine Strukturen
markiert. Der N-Cadherin-Fusionsprotein-Antikörper zeigte also unter unseren
Untersuchungsbedingungen keine Kreuzreaktivität mit E-Cadherin.
Abb. 4.2.14: N-Cadherin-Immunreaktivität in einem Semidünnschnitt durch das Herzmuskel-
Gewebe. Beachte die intensive Immunreaktivität der Glanzstreifen.
Inkubation mit N-Cadherin-Fusionsprotein-Antikörper und Garb-Cy3. Maßstab: 10 µm.
Um weiter zu untersuchen, ob N-Cadherin im olfaktorischen Epithel lokalisiert war,
wurde zudem ein Peptidantikörper (2.8) hergestellt (3.13). Er war gegen die
Aminosäuren 4 bis 22 der ersten extrazellulären Domäne des N-Cadherin der Ratte
gerichtet, die nahe der Bindungsdomäne lokalisiert sind. Diese Peptidsequenz erfüllte
folgende Kriterien:
a) Sie war 19 Aminosäuren lang und damit groß genug, um als Antigen erkannt zu
werden.
b) Sie hatte mit den Aminosäuren Arginin (R), Asparagin (N), Asparaginsäure (D),
Glutamin (Q), Serin (S), Threonin(T) und Tyrosin (Y) hydrophilen Charakter,
war also wasserlöslich.
c) Sie hatte einen Lysin-Rest (K), über den sie als Hapten an ein Trägermolekül
(z.B. KLH) gekoppelt werden konnte.
d) Sie wies keine Glykosylierungsstellen auf, welche die Aminosäurenstruktur
gestört hätten.
e) Sie enthielt keine Bindungsstellen für Cadherine oder Kalziumionen, so daß sie
einer Antigen-Antiköper-Reaktion frei zugänglich war.
f) Sie wies kaum Homologien auf, die Kreuzreaktivität zeigen konnten.

Ergebnisse 49
Das Peptid wurde über Glutaraldehyd an KLH gekoppelt (3.13.1) und gegen PBS
dialysiert (3.13.3).
Als Wirtstiere dienten zwei Kaninchen. Nachdem das Präimmunserum an
Semidünnschnitten des olfaktorischen Epithels und des Herzmuskels keine
unspezifischen Antigen-Antikörper-Reaktionen zeigte (keine Abbildung; 3.13.4), wurde
die Immunisierung wie unter 3.13.4 beschrieben durchgeführt. Das durch regelmäßige
Blutentnahmen gewonnene Immunserum wurde an Semidünnschnitten der
Riechschleimhaut (Testprobe) und des Herzmuskels (Positivprobe) untersucht (3.13.5)
und bei Nachweis einer spezifischen Antigen-Antikörper-Reaktion affinitätsgereinigt
(3.13.6). Als die optimale Antikörperkonzentration erreicht war, wurden die Wirtstiere
geschlachtet und deren gesamtes Blut verarbeitet (3.13.5; 3.13.6).
Nach Inkubation einer Immunreplikanalyse (3.17) mit dem N-Cadherin-
Peptidantikörper und dem Sekundär-Antikörper Garb-HRP-Pox zeigte sich in der Spur
des olfaktorischen Epithels keine Bande, in der Spur des Herzmuskel-Gewebes eine
Bande bei 130 kDa (Abb. 4.2.15). Diese war schwächer als die Banden, die durch den
Antiköper gegen Pan-Cadherin und den Antikörper gegen das N-Cadherin-
Fusionsprotein im Herzmuskel markiert wurden.
Abb. 4.2.15: Immunreaktivität des N-Cadherin-Peptidantikörpers im olfaktorischen
Epithel und im Herzmuskel.

Ergebnisse 50
Inkubation einer Immunreplikanalyse des olfaktorischen Epithels (OE) und des
Herzmuskels (HM) mit N-Cadherin-Peptidantikörper und Garb-HRP-Pox.
Trotz zahlreicher Inkubationen (3.12) wurde mit dem N-Cadherin-Peptidantikörper
keine Immunreaktivität im olfaktorischen Epithel beobachtet (keine Abbildung). Im
Herzmuskel konnten Glanzstreifen regelmäßig dargestellt werden (Abb. 4.2.16). Auch
im Vomeronasalorgan konnten Interzellularkontakte nachgewiesen werden (Abb.
4.2.17).
Abb. 4.2.16: Immunreaktivität des N-Cadherin-Peptidantikörpers in einem Semidünnschnitt
durch den Herzmuskel.
Inkubation mit N-Cadherin-Peptidantikörper und Garb-Cy3. Maßstab: 10 µm.
Abb. 4.2.17: Immunreaktivität des N-Cadherin-Peptidantikörpers in einem
Longitudinalschnitt durch das Vomeronasalorgan.
Inkubation mit N-Cadherin-Peptidantikörper und Garb-Cy3. Maßstab: 5 µm.

Ergebnisse 51
Der N-Cadherin-Peptidantikörper detektierte somit spezifisch N-Cadherin. Entweder
war die Sensitivität des Peptidantikörpers für eine Detektion von N-Cadherin in der
Riechschleimhaut zu gering, oder das N-Cadherin-Epitop, gegen das der
Peptidantikörper gerichtet war, war dem Peptidantikörper nicht zugänglich. Alternativ
könnte das durch Pan-Cadherin- und den N-Cadherin-Fusionsprotein-Antikörper
detektierte Cadherin der olfaktorischen Neurone in der durch den Peptidantikörper
detektierten Sequenz Unterschiede zu N-Cadherin aufweisen.
Lokalisation von Cateninen
Das zytoplasmatische Plaqueprotein α-Catenin besteht aus 906 Aminosäuren. Drei
Sequenzabschnitte sind dem zytoplasmatischen Plaqueprotein Vinculin sehr ähnlich. α-
Catenin liegt als Homodimer in Lösung vor. Es dissoziiert und bildet ein Heterodimer
mit β-Catenin (Pokutta et al., 2002).
Längs- (Abb. 4.2.18) und Schrägschnitte (Abb. 4.2.19) wurden mit dem α-Catenin-
Antikörper und dem Sekundär-Antikörper Gam-Cy3 inkubiert (3.12).
Abb. 4.2.18: α-Catenin-Immunreaktivität in einem Längsschnitt durch das
olfaktorische Epithel. Maßstab: 10 µm.

Ergebnisse 52
Apikal zeigten sich kräftig und schwach fluoreszierende Streifen, welche den Zonulae
adherentes zwischen Stützzellen und Stützzellen sowie zwischen Stützzellen und
Neuronen entsprachen. Schwache fluoreszierende Punkte im Bereich der lateralen
Zellgrenzen markierten Puncta adherentia. Von basal nach apikal zogen schwach
fluoreszierende tubuläre Strukturen, welche den Ausführungsgängen der Bowmanschen
Drüsen entsprachen.
Abb. 4.2.19: α-Catenin-Immunreaktivität in einem Schrägschnitt durch das
olfaktorische Epithel.
Großer Pfeil: Apikale Zellkontakte; mittelgroßer Pfeil: Intraepitheliale
Stützzell-Dendrit-Kontakte; kleiner Pfeil: Stützzell-Stützzell-Kontakte.
Maßstab: 10 µm.
In Abbildung 4.2.19 erkannte man apikal lokalisierte kräftig fluoreszierende Kreise, die
aufgrund ihres Durchmessers in aufsteigender Reihenfolge den Zonulae adherentes
zwischen Stützzellen und Neuronen, zwischen Stützzellen und Mikrovillizellen sowie
zwischen Stützzellen und Stützzellen zugeordnet werden können.
β-Catenin besteht aus einem 149 Aminosäuren langen N-Terminus, der die
Bindungsstelle für α-Catenin enthält, einer 515 Aminosäuren langen Zwischendomäne,
in der die Bindungsstelle für Cadherine und Transkriptionsfaktoren der Familie der T-
Zell-Faktoren gelegen ist, und einem 119 Aminosäuren langen C-Terminus. β-Catenin
ist Teil der Zonula und des Punctum adhaerens und Aktivator der Transkription der
wnt-Signaltransduktionskette (Pokutta et al., 2002; Gumbiner, 1993).

Ergebnisse 53
Eine Längs- (Abb. 4.2.20) und ein Schrägschnitt (Abb. 4.2.21) wurde mit dem Primär-
Antikörper gegen β-Catenin und dem Sekundär-Antikörper Gam-Cy3 inkubiert (3.12).
Abb. 4.2.20: β-Catenin-Immunreaktivität in einem Schrägschnitt durch das
olfaktorische Epithel. Maßstab: 10 µm.
Abb. 4.2.21: β-Catenin-Immunreaktivität in einem Schrägschnitt durch das olfaktorische
Epithel.
Großer Pfeil: Apikale Zellkontakte; mittelgroßer Pfeil: Intraepitheliale Stützzell-
Dendrit-Kontakte; kleiner Pfeil: Stützzell-Stützzell-Kontakte. Maßstab: 10 µm.

Ergebnisse 54
Die Verteilung des β-Catenins entsprach der des α-Catenins (siehe oben).
Die Doppelinkubation eines Tangentialschnitts durch das olfaktorische Epithel mit den
Pan-Cadherin- und β-Catenin-Antikörpern belegt die Kolokalisation der beiden Zonula-
adherens-Proteine (gelbe Fluoreszenz) und bestätigt damit die Identifizierung der durch
den Pan-Cadherin-Antikörper markierten Strukturen als Zonulae adherentes bzw.
Puncta adherentia (Abb. 4.2.22).
Abb. 4.2.22: Pan-Cadherin- (grün) und β-Catenin-Immunreaktivität (rot) in einem
Tangentialschnitt durch das olfaktorische Epithel.
Doppelinkubation mit Pan-Cadherin-/ β-Catenin-Antikörper und Garb-Cy3/ Gam-Cy2
(rot/ grün). Maßstab: 5 µm.
Lokalisation von α-Actinin
α-Actinin vermittelt den Kontakt der zytoplasmatischen Plaqueproteine mit dem Aktin-
Zytoskelett sowie die Verbindung zwischen Aktinfilamenten (Gumbiner, 1993).
Ein tangentialer Semidünnschnitt der Riechschleimhaut wurde mit dem Primär-
Antikörper gegen α-Actinin und dem Sekundär-Antikörper Garb-Cy3 inkubiert (Abb.
4.2.23; 3.12).

Ergebnisse 55
Abb. 4.2.23: α-Actinin-Immunreaktiviät in einem Tangentialschnitt durch das
olfaktorische Epithel. Maßstab: 5 µm.
Apikal gelegene kräftig fluoreszierende Kreise entsprachen nach ihrem Durchmesser
geordnet in absteigender Reihenfolge den Zonulae adherentes zwischen Stützzellen und
Stützzellen, Stützzellen Mikrovillizellen sowie Stützzellen und Neuronen.

Diskussion 56
5. Diskussion
Im Rahmen der vorliegenden Arbeit wurden die Zellkontaktproteine des olfaktorischen
Epithels lokalisiert und charakterisiert. Hierzu wurden Zellen und Zellkontakte der
Riechschleimhaut immunhistochemisch, licht- und elektronenmikroskopisch sowie
biochemisch identifiziert und einander zugeordnet.
Das olfaktorische Epithel ist ein einzigartiger Abschnitt des Nervensystems. Es hat eine
Fläche von ungefähr 62 mm2 und kleidet beidseits das obere Drittel der
Nasenscheidewand aus. Das Neuroepithel enthält neben den epithelialen Stützzellen,
Mikrovillizellen vom Typ 1 und Typ 2, Zellen des Ausführungsganges der
Bowmanschen Drüsen und horizontalen Basalzellen die neuronalen Sinneszellen und
rundlichen Basalzellen. Die Zellkörper der verschiedenen Zellen liegen in Schichten
übereinander. Von basal nach apikal finden sich Basalzellen, Sinneszellen, Stützzellen
und Mikrovillizellen vom Typ 1 und 2. Die Zellkörper der Zellen des
Ausführungsganges der Bowmanschen Drüsen durchziehen die Riechschleimhaut
vertikal (Mendoza, 1993).
Die neuronalen Sinneszellen sind bipolare Neurone. Ihr Dendrit reicht mit jeweils 5 bis
20 Zilien in das Lumen der Nasenhöhle. Die olfaktorischen Sinneszellen sind damit die
einzigen Neurone, die in direktem Kontakt mit der Außenwelt stehen (Drenckhahn et
al., 1994; Mori et al., 1995). Zudem sind sie die einzigen sensorischen Neurone, die in
Form eines Epithels und nicht eines Ganglions organisiert (Miragall et al., 1994) und
somit an der Ausbildung einer Epithelbarriere beteiligt sind.
Elektronenmiskroskopische Untersuchungen haben gezeigt, dass die Epithelbarriere aus
Zonulae occludentes, Zonulae adherentes und Desmosomen zwischen den
unterschiedlichen Zelltypen des olfaktorischen Epithels aufgebaut wird. Ein Ziel der
vorliegenden Arbeit war es, die Lokalisation von Zellkontakt-assoziierten Proteinen in
den für Neurone ungewöhnlichen Zonulae occludentes (Occludin, ZO1) sowie in
Zonulae adherentes (Cadherine, Catenine, α-Actinin) zu untersuchen. Voraussetzung
dafür war, dass die verschiedenen Zelltypen sowie ihre Zellgrenzen zueinander
erkennbar waren und dass homotypische (Stützzelle-Stützzelle) sowie die verschiedenen
möglichen heterotypischen Zellkontakte (Stützzelle-Neuron, Stützzelle-Mikrovillizelle,
Stützzelle-Ausführungsgangszelle) eindeutig identifiziert werden konnten. Durch

Diskussion 57
Anwendung unterschiedlicher Kombinationen von Antikörpern gegen Markerproteine
und Zellkontaktproteine gelang es in der vorliegenden Arbeit, das morphologische
Erscheinungsbild der Zellkontakte der verschiedenen Zelltypen zu dokumentieren. Im
apikalen Transversalschnitt konnten polygonale Formen als Kontakte zwischen
Stützzellen, kleinere Kreise als Kontakte zwischen Stützzellen und Neuronen
(Dendriten) und größere Kreise als Kontakte zwischen Stützzellen und Mikrovillizellen
identifiziert werden. Kontakte zwischen Stützzellen und Ausführungsgangszellen sowie
zwischen Ausführungsgangszellen waren als longitudinale Linien im
Longitudinalschnitt bzw. als intraepitheliale Punkte im Tangentialschnitt zu erkennen.
Diese Morphologie der Zellkontakte fand sich sowohl in der immunhistochemischen
Darstellung der Zonula-occludens- als auch der Zonula-adherens-Proteine. Eine
wichtige Beobachtung war, dass Stützzellen alle anderen Zelltypen kontaktieren und
umhüllen, während direkte apikale Kontakte zwischen einzelnen Neuronen oder
einzelnen Mikrovillizellen nicht beobachtet wurden. Stützzellen scheinen also sowohl
Neurone als auch Mikrovillizellen zu isolieren und damit zumindest bezüglich der
Neurone ähnliche Eigenschaften zu haben wie Gliazellen. Die Tatsache, dass
Mikrovillizellen wie die Neurone komplett eingehüllt werden, könnte auf eine rezeptive
Funktion dieser Zellen hindeuten. Zumindest bei Mikrovillizellen vom Typ II
unterstützt die starke Na+/K+-ATPase-Reaktivität der basolateralen Membranen diese
Hypothese.
Durch Toxine in der Atemluft oder Infektionen kann es zu einem massiven Untergang
der olfaktorischen Neurone kommen. Ein Verlust des Riechsinnes wird vermieden,
indem olfaktorische Neurone zeitlebens durch Proliferation von rundlichen Basalzellen
regeneriert werden (Doucette, 1990). Während ihrer Differenzierung zu reifen
Sinneszellen wandert der Zellkörper nach apikal, der Zellkern wird euchromatinreich.
Der Dendrit erstreckt sich entlang der Stützzellen zur Oberfläche des olfaktorischen
Epithels und bildet neue Interzellularkontakte zur Aufrechterhaltung der Epithelbarriere
aus. Das Axon bahnt sich seinen Weg durch die Basalmembran in die Lamina propria
(Morrison et al., 1992) und wächst in den Fila olfactoria zu spezifischen Glomeruli des
Bulbus olfactorius aus, wo neue synaptische Verbindungen zu den Zielneuronen
ausgebildet werden.

Diskussion 58
Es ist unklar, über welche Mechanismen olfaktorische Neurone das zielgerichtete
Wachstum ihrer Axone steuern. Auch die zonale Projektion wird durch Faktoren
bestimmt, deren molekulare Struktur kaum bekannt ist (Levai et al., 2003). Hierzu
gehören der Geruchsrezeptor (Mombaerts et al., 1996; Wang et al., 1998; Rodriguez et
al., 1999) und Zellkontaktproteine, z.B. das olfaktorische Zell-Adhäsions-Molekül
OCAM (cell adhesion molecule) (Yoshihara et al., 1997). OCAM wurde erstmals von
Yoshihara et al. (1997) beschrieben. Es handelt sich um ein Zellkontaktprotein aus der
Familie der Immunglobuline. Sinneszellen, die OCAM exprimierten, projizieren auf
Mitral- und Büschelzellen, die ebenfalls OCAM enthielten. Die Struktur des
Zellkontaktproteins ähnelt am ehesten NCAM. Im Gegensatz zu OCAM wird jedoch
NCAM im Bereich des gesamten zentralen Nervensystems exprimiert. Eine
Immunglobulin-ähnliche Struktur wurde auch für die Zellkontaktproteine apCAM und
Fasciclin II gefunden, die der Familie der NCAMs von Aplysia und Drosophila
angehören (Yoshihara et al., 1997). apCAM und Fasciclin II werden von bestimmten
Neuronen exprimiert und kontrollieren das Auswachsen von Axonen. Lang anhaltende
Stimulation der Neurone führt zu einer Verminderung der Anzahl von apCAM und
Fasciclin II durch Internalisierung oder Konformationsänderung (Bailey et al., 1992;
Mayford et al., 1992; Yoshihara et al., 1993). Zugleich nimmt die Anzahl synaptischer
Verbindungen deutlich zu (Yoshihara et al., 1997). Die Tatsache, daß OCAM nach
Bindung eines Geruchsmoleküls an einen Geruchsrezeptor nicht mehr nachweisbar ist,
legt die Vermutung nahe, daß es nicht nur bezüglich seiner Struktur, sondern auch
bezüglich seiner Funktion apCAM und Fasciclin II ähnelt (Yoshihara et al., 1997).
Neuere Untersuchungen haben ergeben, daß die Expression von OCAM in den
Dendriten der Sinneszellen Informationen über das Auswachsen der Axone vermittelt
(Treloar et al., 2003). OCAM-ähnliche Eigenschaften scheint das von Alenius et al.
(1997) beschriebene Immunglobulin RNCAM (Rb-8 neural cell adhesion molecule)zu
haben. Desweiteren scheinen die Zellkontaktproteine N-Cadherin, NCAM und L1 am
Auswachsen von Axonen beteiligt zu sein (Doherty et al., 1995; Doherty et al., 2000;
Kamiguchi et al., 2000; Utton et al., 2001). Cadherine könnten also neben ihrer
Bedeutung für die olfaktorische Epithelbarriere auch wichtige Funktionen für die
axonale Zielfindung im olfaktorische Epithel haben.

Diskussion 59
Der Kontakt zwischen der präsynaptischen Membran der Sinneszelle und der
postsynaptischen Membran der Mitral- oder Büschelzelle wird über klassische
Cadherine und Catenine vermittelt (Yamagata et al., 1995; Fannon et al., 1996; Uchida
et al., 1996). Die Transmembranproteine wurden zusammen mit den zytoplasmatischen
Plaqueproteinen symmetrisch angeordnet und direkt an das aktive Zentrum der Synapse
angrenzend lokalisiert (Uchida et al., 1996). Aufgrund ihrer besonderen
Bindungseigenschaften wird vermutet, daß Cadherine nach dem Schlüssel-Schloß-
Prinzip zielgerichtete interneuronale Verbindungen herstellen (Yagi et al., 2000). Im
Bereich des aktiven Zentrums kleiner und unreifer Synapsen wurden Proteine der CNR-
Familie identifiziert (Kohmura et al., 1998). Diese Zellkontaktproteine sind mit der
Tyrosinkinase Fyn assoziiert. Neben der Bildung synaptischer Bindungen scheinen sie
an der Signaltransduktion beteiligt zu sein (Yagi et al., 2000). Auch Proteine der
Protocadherin-Familie finden sich im Bereich von Synapsen. Arcadlin wurde mit
Synaptophysin, einem präsynaptischen Markerprotein, kolokalisiert. OL-Pc scheint in
funktionellen Einheiten wie zum Beispiel den Kernen des olfaktorischen Systems
exprimiert zu werden. Die Vielfalt synaptischer Bindungen beruht auf der Kombination
dieser einzelnen Zellkontaktproteine. N-Cadherin stellt nicht nur eine synaptische
Bindung her. Das Transmembranprotein ist an Vorgängen der synaptischen Plastizität
beteiligt. Tanaka et al. (2000) beschrieben, daß nach Depolarisation der präsynaptischen
Plasmamembran und Fusion der präsynaptischen Vesikel N-Cadherin lateralwärts
diffundierte. Zudem war eine Fraktion von N-Cadherin bis zwei Stunden nach der
Depolarisation gegen Proteasen resistent und lag als Monomer vor (Yagi et al., 2000).
Zellkontaktproteine, unter anderem Cadherine, scheinen somit nicht nur an der
Ausbildung epitheltypischer Kontakte, sondern im olfaktorischen System wie in
anderen Abschnitten des Nervensystems auch an axonalem Wachstum, axonaler
Zielfindung sowie an Synaptogenese und synaptischer Plastizität beteiligt zu sein.
Aufgrund seiner hohen Plastizität ist das olfaktorische Epithel ein ideales Modell zur
Untersuchung der Rolle von Zellkontaktproteinen bei den beschriebenen Vorgängen
(Miragall et al., 1994; Yoshihara et al., 1997). Voraussetzung für derartige
Untersuchungen ist zunächst die Analyse der Expression solcher Zellkontaktproteine,
z.B. der Cadherine, insbesondere in olfaktorischen Neuronen. Neben der Darstellung
der Lokalisation von Kontaktproteinen war es daher ein weiteres Ziel der vorliegenden

Diskussion 60
Arbeit, die von den verschiedenen Zelltypen des olfaktorischen Epithels exprimierten
Cadherine zu bestimmen.
Proteine der Zonulae occludentes im olfaktorischen Epithel
Die Zonula occludens besteht unter anderem aus den Transmembranproteinen Occludin,
verschiedenen Claudinen und dem zytoplasmatischen Plaqueprotein ZO-1 (1.2). Im
Rahmen der vorliegenden Arbeit wurden Semidünnschnitte des Riechepithels mit
Antikörpern sowohl gegen Occludin als auch gegen ZO-1 inkubiert (Abb. 4.1.1 – 4.1.3).
Beide Proteine waren in allen Zonulae occludentes des olfaktorischen Epithels
lokalisiert. Das Vorkommen von Occludin in Neuronen war bereits beschrieben worden
(Hussar et al., 2002), das Verteilungsmuster des Proteins konnte jedoch hier erstmals
gezeigt werden. Während die ZO1-Immunfluoreszenz in allen Zellkontakten gleich
stark erschien, war die relative Occludin-Immunreaktivität in epithelialen
Interzellularkontakten (Stützzelle-Stützzelle, Stützzelle-Mikrovillizelle, Stützzelle-
Ausführungsgangszelle) stärker ausgeprägt als in epithelial-neuronalen
Interzellularkontakten (Stützzelle-Sinneszelle). Wahrscheinlich ist Occludin in
epithelial-neuronalen Interzellularkontakten geringer konzentriert ist als in epithelialen
Interzellularkontakten, was auf eine unterschiedliche molekulare Komposition der
Zonulae occludentes zwischen den verschiedenen Zelltypen und auf unterschiedliche
Barriereeigenschaften der jeweiligen Interzellularkontakte hinweist (McCarthy et al.,
1996; Hirase et al., 1997; Balda et al., 2000). Eine Untersuchung der verschiedenen
Claudine des olfaktorischen Epithels muss zur genaueren Charakterisierung der
molekularen Zusammensetzung der verschiedenen Zonulae occludentes noch
durchgeführt werden.
Proteine der Zonulae adherentes im olfaktorischen Epithel
Die Zonula adherens epithelialer Zellen enthält das Transmembranprotein E-Cadherin.
Ein typisches Transmembranprotein der Zonula adherens neuronaler Zellen ist das N-
Cadherin. Die zytoplasmatischen Plaqueproteine und Proteine des Zytoskeletts
unterscheiden sich nicht.
Im Rahmen der vorliegenden Arbeit wurden Semidünnschnitte des olfaktorischen
Epithels mit E-Cadherin- und Pan-Cadherin-Antikörpern inkubiert. Durch den E-

Diskussion 61
Cadherin-Antikörper stellten sich Zonulae adherentes und Puncta adherentia zwischen
epithelialen Stützzellen, Mikrovillizellen und Zellen des Ausführungsganges der
Bowmanschen Drüsen dar (Abb. 4.2.2; Abb. 4.2.3). Pan-Cadherin-Antikörper zeigten in
den Zonulae adherentes zwischen Stütz- und Sinneszellen stärkere, in den Zonulae
adherentes und Puncta adherentia zwischen Stützzellen schwächere Immunreaktivität.
Dies bedeutet, dass epitheliale Stützzellen und Neurone einen Interzellularkontakt auf
der Basis von Cadherinen ausbilden. Da Pan-Cadherin-Antikörper gegen einen Teil der
intrazellulären Domäne von N-Cadherin gerichtet sind, in unseren Gewebepräparaten E-
Cadherin nicht detektieren und in Immunreplikanalysen eine Bande detektieren, die
ähnliche Laufeigenschaften wie N-Cadherin aufweist, ist anzunehmen, dass das
Cadherin der Stützzell-Sinneszell-Kontakte N-Cadherin oder ein N-Cadherin-ähnliches
Cadherin ist.
Mit Hilfe der Doppelinkubation konnte gezeigt werden, daß sich die Immunreaktivität
für E-Cadherin- und Pan-Cadherin-Antikörper im Bereich der Zonulae adherentes
zwischen Stützzellen überlagerten (Abb. 4.2.9; Abb. 4.2.10). Ein heterophiler Kontakt
zwischen E-Cadherin und anderen Cadherinen, insbesondere N-Cadherin, wurde bislang
nicht beschrieben, sondern im Gegenteil durch in vitro Experimente wiederlegt
(Takeichi, 1995; Gumbiner, 1996; Yap et al., 1997). Eine Erklärung für unsere Befunde
ist, dass Stützzellen sowohl E-Cadherin als auch N-Cadherin bzw. ein N-Cadherin-
ähnliches Cadherin exprimieren, während die übrigen epithelialen Zellen (Mikrovilli-
und Ausführungsgangszellen) ausschließlich E-Cadherin und die Neurone
ausschließlich N-Cadherin oder das N-Cadherin-ähnliche Cadherin bilden. Je nach
Bindungspartner wird in Stützzellen offensichtlich E-Cadherin in die Membrandomänen
lokalisiert, die in Kontakt mit anderen epithelialen Zellen stehen, N-Cadherin oder das
N-Cadherin-ähnliche Cadherin in die Membrandomänen, die in Kontakt zu Neuronen
stehen, und möglicherweise beide Cadherine in die Membrandomänen zwischen
Stützzellen. Als Mechanismus für die Lokalisierung der verschiedenen Cadherine in den
spezifischen Membrandomänen ist vorstellbar, dass die in der basolateralen Membran
der Stützzellen zunächst frei beweglichen Cadherine durch homophile Bindung an die
jeweiligen Cadherine der Kontaktzelle immobilisiert, konzentriert, und damit
immunhistochemisch detektierbar werden. Eine Stabilisierung dieser Lokalisierung
erfolgt anschließend sekundär über eine Koppelung an das Aktin-Zytoskelett durch die

Diskussion 62
peripheren Membranproteine der Zonulae adherentes (s.o.). Ähnliche spezifische
Lokalisationsmuster in Membrandomänen wurden für E-Cadherin bzw. N-Cadherin in
hippocampalen Neuronen gezeigt (Fannon et al., 1996). Die stabilisierende Wirkung des
Aktin-Zytoskeletts durch Cadherine vermittelte Zellkontakte konnte kürzlich
nachgewiesen werden (Baumgartner et al., 2003).
Das Cadherin der olfaktorischen Neurone
Um das Cadherin der Neurone genauer zu charakterisieren, führten wir Untersuchungen
mit N-Cadherin-spezifischen Antikörpern durch. In der Immunreplikanalyse detektierte
der N-Cadherin-Fusionsprotein-Antikörper im olfaktorischen Epithel eine etwa gleich
laufende Bande wie im Herzmuskel. In situ war mit diesem Antikörper in Zonulae
adherentes zwischen Stütz- und Sinneszellen sowie zwischen Stützzellen eine relativ
schwache Immunfluoreszenz zu beobachten. Der N-Cadherin-Peptidantikörper, den wir
gegen eine in der unmittelbaren Nähe der putativen HAV-Bindungsdomäne von N-
Cadherin gelegene Aminosäuresequenz generiert hatten und der sowohl im Herzmuskel
als auch im Vomeronasalorgan Zonulae adherentes markierte, zeigte im olfaktorischen
Epithel zum einen in der Immunhistochemie keine Reaktion und detektierte zum
anderen in der Immunreplikanalyse kein Protein. Eine mögliche Erklärung für diese
Beobachtungen wäre, dass Stützzellen und Sinneszellen nicht N-Cadherin, sondern ein
mit N-Cadherin eng verwandtes Cadherin exprimieren. Ein relativ hoher Grad an N-
Cadherin-Homologie, besonders in der intrazellulären Domäne dieses Cadherins,
könnte erklären, warum die epithelial-neuronalen Zonulae adherentes durch den gering
spezifischen Pan-Cadherin-Antikörper gut, durch den gegen die gesamte erste
extrazelluläre Domäne des N-Cadherin gerichteten N-Cadherin-Fusionsprotein-
Antikörper weniger gut, und durch den hochspezifischen N-Cadherin-Peptidantikörper
gar nicht detektiert wurde. Eine weitere Möglichkeit wäre, dass die olfaktorischen
Neurone und Stützzellen zwar N-Cadherin synthetisieren, dass aber die erste
extrazelluläre Domäne bzw. die Aminosäuresequenz in der Nähe des putativen HAV-
Bindungsmotivs durch die ausgebildete Zonula adherens den Antikörpern im
Riechepithel nicht zugänglich ist. Ein ähnliches Phänomen wurde bei einer
Tumorzelllinie beobachtet (Schmelz et al., 1998).

Diskussion 63
Eine genaue Identifizierung des Cadherins der olfaktorischen Neurone ist nur durch In-
situ-Hybridisierung der Cadherin-mRNA möglich. Diese Untersuchungen müssen in
zukünftigen Studien durchgeführt werden.
Zytoplasmatische Proteine der Zonulae adherentes
α-Catenin und β-Catenin konnte in allen Zonulae adherentes dargestellt werden, die
durch Cadherin-Immunreaktivitäten detektierbar waren. Die Anbindung an das Aktin-
Zytoskelett scheint daher keine Unterschiede zu anderen epithelialen Zellkontakten
aufzuweisen.
Schlußfolgerungen
Die Untersuchungen der vorliegenden Arbeit haben eine Reihe neuer Erkenntnisse
bezüglich der Zellkontakte des olfaktorischen Epithels der Ratte erbracht. Die genaue
Charakterisierung des Erscheinungsbildes der verschiedenen Interzellularkontakte im
histologischen Schnitt ermöglichte die spezifische Zuordnung der Immunreaktivitäten
für die einzelnen Zellkontaktproteine. Es wurde nachgewiesen, dass Stützzellen alle
übrigen Zelltypen des olfaktorischen Epithels umhüllen. Dies bedeutet, dass zumindest
im Bereich des apikalen Epithels sowohl Neurone als auch Mikrovillizellen
untereinander und voneinander isoliert sind. Es konnte erstmals die Verteilung des
Zonula-occludens-Proteins Occludin im Riechepithel dargestellt werden. Bei der
Charakterisierung der molekularen Zusammensetzung der Zonulae adherentes zwischen
unterschiedlichen Zelltypen wurde beobachtet, dass Stützzellen zwei verschiedene
Cadherine synthetisieren: E-Cadherin wird für Zellkontakte zu epithelialen Zellen
benötigt (Mikrovillizellen, Ausführungsgangszellen), ein neuronales Cadherin für
Zellkontakte zu neuronalen Zellen (Sinneszellen) und beide Cadherine für Zellkontakte
zwischen Stützzellen. Es gibt Hinweise, dass es sich bei dem neuronalen Cadherin um
N-Cadherin handeln könnte, eine genaue Identifizierung muss in zukünftigen
Untersuchungen erfolgen.

Zusammenfassung 64
5. Zusammenfassung
Ziel der vorliegenden Arbeit war die Lokalisierung und Charakterisierung von
Zellkontaktproteinen im olfaktorischen Epithel. Hierzu wurde die Riechschleimhaut der
Ratte für immunhistochemische, elektronenmikroskopische und biochemische
Experimente aufbereitet. Nachdem das Erscheinungsbild der verschiedenen
Interzellularkontakte im histologischen Schnitt charaktersisiert worden war, wurde
durch die Detektion von Transmembranproteinen, zytoplasmatischen Plaqueproteinen
und spezifischen Markermolekülen die genaue zelltypspezifische Zusammensetzung der
Zellkontakte dargestellt.
Die Untersuchungen haben eine Reihe neuer Erkenntnisse erbracht. Erstmals wurde
nicht nur das Vorkommen, sondern auch die Verteilung des Zonula-occludens-Proteins
Occludin dargestellt. Es wurde nachgewiesen, dass alle Zelltypen in der apikalen
Riechschleimhaut von Stützzellen umhüllt werden. Sowohl Neurone als auch
Mikrovillizellen sind somit untereinander und voneinander getrennt. Es wurde
beobachtet, dass Stützzellen zwei verschiedene Cadherine synthetisieren: E-Cadherin
für Zellkontakte zu epithelialen Zellen (Mikrovillizellen, Ausführungsgangszellen), ein
neuronales Cadherin, möglicherweise N-Cadherin, für Zellkontakte zu neuronalen
Zellen (Sinneszellen) und beide Cadherine für Zellkontakte zwischen Stützzellen.

Literaturverzeichnis 65
7. Literaturverzeichnis
Adams, C. L., Nelson, W. J., and Smith, S. J. (1996): Quantitative analysis of cadherin-
catenin-actin reorganization during development of cell-cell adhesion. J Cell Biol 135,
1899-911.
Alenius, M., and Bohm, S. (1997): Identification of a novel neural cell adhesion
molecule-related gene with a potential role in selective axonal projection. J Biol Chem
272, 26083-6.
Amagai, M., Nishikawa, T., Nousari, H. C., Anhalt, G. J., and Hashimoto, T. (1998):
Antibodies against desmoglein 3 (pemphigus vulgaris antigen) are present in sera from
patients with paraneoplastic pemphigus and cause acantholysis in vivo in neonatal mice.
J Clin Invest 102, 775-82.
Amagai, M. (1999): Autoimmunity against desmosomal cadherins in pemphigus. J
Dermatol Sci 20, 92-102.
Amagai, M., Matsuyoshi, N., Wang, Z. H., Andl, C., and Stanley, J. R. (2000): Toxin in
bullous impetigo and staphylococcal scalded-skin syndrome targets desmoglein 1. Nat
Med 6, 1275-7.
Asan, E. (2004): Geruchssystem. In: Benninghoff-Drenckhahn, Anatomie, 16. Auflage,
Band 2, D. Drenckhahn (Herausgeber), Elsevier, Urban und Fischer, S. 746-754.
Asan, E., Höfer, and Drenckhahn, D. (1999): Immunzytochemische Charakterisierung
von Mikrovillizellen des olfaktorischen Epithels. Ann Ant, 67.
Bailey, C. H., Chen, M., Keller, F., and Kandel, E. R. (1992): Serotonin-mediated
endocytosis of apCAM: an early step of learning-related synaptic growth in Aplysia.
Science 256, 645-9.

Literaturverzeichnis 66
Balda, M. S., and Matter, K. (2000): Transmembrane proteins of tight junctions. Semin
Cell Dev Biol 11, 281-9.
Balda, M. S., Flores-Maldonado, C., Cereijido, M., Matter, K. (2000): Multiple domains
of occludin are involved in the regulation of paracellular permeability. Journal of
Cellular Biochemistry 78, 85-96
Balda, M. S., and Matter, K. (2003): Epithelial cell adhesion and the regulation of gene
expression. Trends Cell Biol 13, 310-8.
Baumgartner, W., Golenhofen, N., Grundhofer, N., Wiegand, J., Drenckhahn, D.
(2003): Ca2+ dependency of N-cadherin function probed by laser tweezer and atomic
force microscopy. J Neurosci 23, 11008-14
Bement, W. M., Forscher, P., and Mooseker, M. S. (1993): A novel cytoskeletal
structure involved in purse string wound closure and cell polarity maintenance. J Cell
Biol 121, 565-78.
Borisy, F. F., Ronnett, G. V., Cunningham, A. M., Juilfs, D., Beavo, J., and Snyder, S.
H. (1992): Calcium/calmodulin-activated phosphodiesterase expressed in olfactory
receptor neurons. J Neurosci 12, 915-23.
Breer, H., Wanner, I., and Strotmann, J. (1996): Molecular genetics of mammalian
olfaction. Behav Genet 26, 209-19.
Burnette, W. N. (1981): "Western blotting": electrophoretic transfer of proteins from
sodium dodecyl sulfate--polyacrylamide gels to unmodified nitrocellulose and
radiographic detection with antibody and radioiodinated protein A. Anal Biochem 112,
195-203.

Literaturverzeichnis 67
Cepek, K. L., Shaw, S. K., Parker, C. M., Russell, G. J., Morrow, J. S., Rimm, D. L.,
and Brenner, M. B. (1994): Adhesion between epithelial cells and T lymphocytes
mediated by E-cadherin and the alpha E beta 7 integrin. Nature 372, 190-3.
Chen, T. Y., and Yau, K. W. (1994): Direct modulation by Ca(2+)-calmodulin of cyclic
nucleotide-activated channel of rat olfactory receptor neurons. Nature 368, 545-8.
Creifido, M (1991): Tight junctions, CRC Press Inc., Boca Raton, FL
Dear, T. N., Boehm, T., Keverne, E. B., and Rabbitts, T. H. (1991): Novel genes for
potential ligand-binding proteins in subregions of the olfactory mucosa. Embo J 10,
2813-9.
Dieckmann-Schuppert, A., and Schnittler, H. J. (1997): A simple assay for
quantification of protein in tissue sections, cell cultures, and cell homogenates, and of
protein immobilized on solid surfaces. Cell Tissue Res 288, 119-26.
Doherty, P., Williams, E., and Walsh, F. S. (1995): A soluble chimeric form of the L1
glycoprotein stimulates neurite outgrowth. Neuron 14, 57-66.
Doherty, P., Williams, G., and Williams, E. J. (2000): CAMs and axonal growth: a
critical evaluation of the role of calcium and the MAPK cascade. Mol Cell Neurosci 16,
283-95.
Doucette, R. (1990): Glial influences on axonal growth in the primary olfactory system.
Glia 3, 433-49.
Drenckhahn, D., and Franz, H. (1986): Identification of actin-, alpha-actinin-, and
vinculin-containing plaques at the lateral membrane of epithelial cells. J Cell Biol 102,
1843-52.

Literaturverzeichnis 68
Drenckhahn, D., Zenker (1994): Benninghoff, Anatomie. Urban und Schwarzenberg,
15. Auflage, S. 784-790.
Drenckhahn, D. (Herausgeber): Benninghoff-Drenckhahn, Anatomie, 16. Auflage, Band
1 und 2, Elsevier, Urban und Fischer, München.
Fannon, A. M., Sherman, D. L., Ilyina-Gragerova, G., Brophy, P. J., Friedrich, V. L.,
Jr., and Colman, D. R. (1995): Novel E-cadherin-mediated adhesion in peripheral nerve:
Schwann cell architecture is stabilized by autotypic adherens junctions. J Cell Biol 129,
189-202.
Fannon, A. M., and Colman, D. R. (1996): A model for central synaptic junctional
complex formation based on the differential adhesive specificities of the cadherins.
Neuron 17, 423-34.
Gonzalez-Mariscal, L., Betanzos, A., Nava, P., and Jaramillo, B. E. (2003): Tight
junction proteins. Prog Biophys Mol Biol 81, 1-44.
Grunwald, G. B. (1993): The structural and functional analysis of cadherin calcium-
dependent cell adhesion molecules. Curr Opin Cell Biol 5, 797-805.
Gumbiner, B., Stevenson, B., and Grimaldi, A. (1988): The role of the cell adhesion
molecule uvomorulin in the formation and maintenance of the epithelial junctional
complex. J Cell Biol 107, 1575-87.
Gumbiner, B. M. (1993): Proteins associated with the cytoplasmic surface of adhesion
molecules. Neuron 11, 551-64.
Gumbiner, B. M. (1996): Cell adhesion: the molecular basis of tissue architecture and
morphogenesis. Cell 84, 345-57.

Literaturverzeichnis 69
Hashimoto, T., Kiyokawa, C., Mori, O., Miyasato, M., Chidgey, M. A., Garrod, D. R.,
Kobayashi, Y., Komori, K., Ishii, K., Amagai, M., and Nishikawa, T. (1997): Human
desmocollin 1 (Dsc1) is an autoantigen for the subcorneal pustular dermatosis type of
IgA pemphigus. J Invest Dermatol 109, 127-31.
Hirase, T., Staddon, J. M., Saitou, M., Ando-Akatsuka, Y., Itoh, M., Furuse, M.,
Fujimoto, K., Tsukita, S., Rubin, L. L. (1997): Occludin as a possible determinant of
tight junction permeability in endothelial cells. Journal of Cell Science 110, 1603-1613
Hofer, D., Asan, E., and Drenckhahn, D. (1999): Chemosensory Perception in the Gut.
News Physiol Sci 14, 18-23.
Holbrook, E. H., Szumowski, K. E., and Schwob, J. E. (1995): An immunochemical,
ultrastructural, and developmental characterization of the horizontal basal cells of rat
olfactory epithelium. J Comp Neurol 363, 129-46.
Hussar, P., Tserentsoodol, N., Koyama, H., Yokoo-Sugawara, M., Matsuzaki, T.,
Takami, S., Takata, K. (2002): The glucose transporter GLUT1 and the tight junction
protein occludin in nasal olfactory mucosa. Chem Senses 27, 7-11
Ishii, K. (2003): Greater diversity of desmosomal cadherins. J Invest Dermatol 120, ix-
x.
Itoh, M., Furuse, M., Morita, K., Kubota, K., Saitou, M., and Tsukita, S. (1999): Direct
binding of three tight junction-associated MAGUKs, ZO-1, ZO-2, and ZO-3, with the
COOH termini of claudins. J Cell Biol 147, 1351-63.
Jang, W., Youngentob, S. L., and Schwob, J. E. (2003): Globose basal cells are required
for reconstitution of olfactory epithelium after methyl bromide lesion. J Comp Neurol
460, 123-40.

Literaturverzeichnis 70
Kamiguchi, H., and Lemmon, V. (2000): IgCAMs: bidirectional signals underlying
neurite growth. Curr Opin Cell Biol 12, 598-605.
Kohmura, N., Senzaki, K., Hamada, S., Kai, N., Yasuda, R., Watanabe, M., Ishii, H.,
Yasuda, M., Mishina, M., and Yagi, T. (1998): Diversity revealed by a novel family of
cadherins expressed in neurons at a synaptic complex. Neuron 20, 1137-51.
Kramer, R. H., and Siegelbaum, S. A. (1992): Intracellular Ca2+ regulates the
sensitivity of cyclic nucleotide-gated channels in olfactory receptor neurons. Neuron 9,
897-906.
Kubota, K., Furuse, M., Sasaki, H., Sonoda, N., Fujita, K., Nagafuchi, A., and Tsukita,
S. (1999): Ca(2+)-independent cell-adhesion activity of claudins, a family of integral
membrane proteins localized at tight junctions. Curr Biol 9, 1035-8.
Laemmli, U. K. (1970): Cleavage of structural proteins during the assembly of the head
of bacteriophage T4. Nature 227, 680-5.
Lazard, D., Zupko, K., Poria, Y., Nef, P., Lazarovits, J., Horn, S., Khen, M., and Lancet,
D. (1991): Odorant signal termination by olfactory UDP glucuronosyl transferase.
Nature 349, 790-3.
Lee, K. H., Wells, R. G., and Reed, R. R. (1987): Isolation of an olfactory cDNA:
similarity to retinol-binding protein suggests a role in olfaction. Science 235, 1053-6.
Levai, O., Breer, H., and Strotmann, J. (2003): Subzonal organization of olfactory
sensory neurons projecting to distinct glomeruli within the mouse olfactory bulb. J
Comp Neurol 458, 209-20.
Li, D., and Mrsny, R. J. (2000): Oncogenic Raf-1 disrupts epithelial tight junctions via
downregulation of occludin. J Cell Biol 148, 791-800.

Literaturverzeichnis 71
Madara, J. L. (1987): Intestinal absorptive cell tight junctions are linked to cytoskeleton.
Am J Physiol 253, C171-5.
Madara, J. L., Moore, R., and Carlson, S. (1987): Alteration of intestinal tight junction
structure and permeability by cytoskeletal contraction. Am J Physiol 253, C854-61.
Madara, J. L., Stafford, J., Barenberg, D., and Carlson, S. (1988): Functional coupling
of tight junctions and microfilaments in T84 monolayers. Am J Physiol 254, G416-23.
Magee, A. I., and Buxton, R. S. (1991): Transmembrane molecular assemblies regulated
by the greater cadherin family. Curr Opin Cell Biol 3, 854-61.
Manabe, T., Togashi, H., Uchida, N., Suzuki, S. C., Hayakawa, Y., Yamamoto, M.,
Yoda, H., Miyakawa, T., Takeichi, M., and Chisaka, O. (2000): Loss of cadherin-11
adhesion receptor enhances plastic changes in hippocampal synapses and modifies
behavioral responses. Mol Cell Neurosci 15, 534-46.
Margolis, F. L. (1972): A brain protein unique to the olfactory bulb. Proc Natl Acad Sci
U S A 69, 1221-4.
Martin-Padura, I., Lostaglio, S., Schneemann, M., Williams, L., Romano, M., Fruscella,
P., Panzeri, C., Stoppacciaro, A., Ruco, L., Villa, A., Simmons, D., and Dejana, E.
(1998): Junctional adhesion molecule, a novel member of the immunoglobulin
superfamily that distributes at intercellular junctions and modulates monocyte
transmigration. J Cell Biol 142, 117-27.
Matsunami, H., Miyatani, S., Inoue, T., Copeland, N. G., Gilbert, D. J., Jenkins, N. A.,
and Takeichi, M. (1993): Cell binding specificity of mouse R-cadherin and
chromosomal mapping of the gene. J Cell Sci 106 ( Pt 1), 401-9.

Literaturverzeichnis 72
Mayford, M., Barzilai, A., Keller, F., Schacher, S., and Kandel, E. R. (1992):
Modulation of an NCAM-related adhesion molecule with long-term synaptic plasticity
in Aplysia. Science 256, 638-44.
McCarthy, K. M., Skare, I. B., Stankewich, M. C., Furuse, M., Tsukita, S., Rogers, R.
A., Lynch, R. D., Schneeberger, E. E. (1996): Occludin is a functional component of the
tight junction. Journal of Cell Science 109, 2287-2298
Mendoza, A. S., and Kuhnel, W. (1991): Postnatal changes in the ultrastructure of the
rat olfactory epithelium: the supranuclear region of the supporting cells. Cell Tissue Res
265, 193-6.
Mendoza, A. S. (1993): Morphological studies on the rodent main and accessory
olfactory systems: the regio olfactoria and vomeronasal organ. Anat Anz 175, 425-46.
Miragall, F., Krause, D., de Vries, U., and Dermietzel, R. (1994): Expression of the
tight junction protein ZO-1 in the olfactory system: presence of ZO-1 on olfactory
sensory neurons and glial cells. J Comp Neurol 341, 433-48.
Mombaerts, P., Wang, F., Dulac, C., Chao, S. K., Nemes, A., Mendelsohn, M.,
Edmondson, J., and Axel, R. (1996): Visualizing an olfactory sensory map. Cell 87,
675-86.
Mooseker, M. S., Bonder, E. M., Conzelman, K. A., Fishkind, D. J., Howe, C. L., and
Keller, T. C., 3rd (1984): Brush border cytoskeleton and integration of cellular
functions. J Cell Biol 99, 104s-112s.
Moran, D. T., Rowley, J. C., 3rd, and Jafek, B. W. (1982): Electron microscopy of
human olfactory epithelium reveals a new cell type: the microvillar cell. Brain Res 253,
39-46.

Literaturverzeichnis 73
Morrison, E. E., and Costanzo, R. M. (1992): Morphology of olfactory epithelium in
humans and other vertebrates. Microsc Res Tech 23, 49-61.
Nagafuchi, A. (2001): Molecular architecture of adherens junctions. Curr Opin Cell
Biol 13, 600-3.
Nef, P., Heldman, J., Lazard, D., Margalit, T., Jaye, M., Hanukoglu, I., and Lancet, D.
(1989): Olfactory-specific cytochrome P-450. cDNA cloning of a novel neuroepithelial
enzyme possibly involved in chemoreception. J Biol Chem 264, 6780-5.
Petruzzelli, L., Takami, M., and Humes, H. D. (1999): Structure and function of cell
adhesion molecules. Am J Med 106, 467-76.
Pokutta, S., and Weis, W. I. (2002): The cytoplasmic face of cell contact sites. Curr
Opin Struct Biol 12, 255-62.
Redies, C., and Takeichi, M. (1993): Expression of N-cadherin mRNA during
development of the mouse brain. Dev Dyn 197, 26-39.
Reichert, M., Muller, T., and Hunziker, W. (2000): The PDZ domains of zonula
occludens-1 induce an epithelial to mesenchymal transition of Madin-Darby canine
kidney I cells. Evidence for a role of beta-catenin/Tcf/Lef signaling. J Biol Chem 275,
9492-500.
Ressler, K. J., Sullivan, S. L., and Buck, L. B. (1994): Information coding in the
olfactory system: evidence for a stereotyped and highly organized epitope map in the
olfactory bulb. Cell 79, 1245-55.
Rodriguez, I., Feinstein, P., and Mombaerts, P. (1999): Variable patterns of axonal
projections of sensory neurons in the mouse vomeronasal system. Cell 97, 199-208.

Literaturverzeichnis 74
Salinovich, O., and Montelaro, R. C. (1986): Reversible staining and peptide mapping
of proteins transferred to nitrocellulose after separation by sodium dodecylsulfate-
polyacrylamide gel electrophoresis. Anal Biochem 156, 341-7.
Sano, K., Tanihara, H., Heimark, R. L., Obata, S., Davidson, M., St John, T., Taketani,
S., and Suzuki, S. (1993): Protocadherins: a large family of cadherin-related molecules
in central nervous system. Embo J 12, 2249-56.
Schleicher, S., Boekhoff, I., Arriza, J., Lefkowitz, R. J., and Breer, H. (1993): A beta-
adrenergic receptor kinase-like enzyme is involved in olfactory signal termination. Proc
Natl Acad Sci U S A 90, 1420-4.
Shapiro, L., Fannon, A. M., Kwong, P. D., Thompson, A., Lehmann, M. S., Grubel, G.,
Legrand, J. F., Als-Nielsen, J., Colman, D. R., and Hendrickson, W. A. (1995a):
Structural basis of cell-cell adhesion by cadherins. Nature 374, 327-37.
Shapiro, L., Kwong, P. D., Fannon, A. M., Colman, D. R., and Hendrickson, W. A.
(1995b): Considerations on the folding topology and evolutionary origin of cadherin
domains. Proc Natl Acad Sci U S A 92, 6793-7.
Staehelin, L. A. (1973): Further observations on the fine structure of freeze-cleaved
tight junctions. J Cell Sci 13, 763-86.
Suzuki, S., Sano, K., and Tanihara, H. (1991): Diversity of the cadherin family:
evidence for eight new cadherins in nervous tissue. Cell Regul 2, 261-70.
Takai, Y., and Nakanishi, H. (2003): Nectin and afadin: novel organizers of intercellular
junctions. J Cell Sci 116, 17-27.
Takeichi, M. (1995): Morphogenetic roles of classic cadherins. Curr Opin Cell Biol 7,
619-27.

Literaturverzeichnis 75
Tanaka, H., Shan, W., Phillips, G. R., Arndt, K., Bozdagi, O., Shapiro, L., Huntley, G.
W., Benson, D. L., and Colman, D. R. (2000): Molecular modification of N-cadherin in
response to synaptic activity. Neuron 25, 93-107.
Tang, L., Hung, C. P., and Schuman, E. M. (1998): A role for the cadherin family of
cell adhesion molecules in hippocampal long-term potentiation. Neuron 20, 1165-75.
Treloar, H. B., Gabeau, D., Yoshihara, Y., Mori, K., and Greer, C. A. (2003): Inverse
expression of olfactory cell adhesion molecule in a subset of olfactory axons and a
subset of mitral/tufted cells in the developing rat main olfactory bulb. J Comp Neurol
458, 389-403.
Tsukita, S., Furuse, M., and Itoh, M. (2001): Multifunctional strands in tight junctions.
Nat Rev Mol Cell Biol 2, 285-93.
Uchida, N., Honjo, Y., Johnson, K. R., Wheelock, M. J., and Takeichi, M. (1996): The
catenin/cadherin adhesion system is localized in synaptic junctions bordering
transmitter release zones. J Cell Biol 135, 767-79.
Utton, M. A., Eickholt, B., Howell, F. V., Wallis, J., and Doherty, P. (2001): Soluble N-
cadherin stimulates fibroblast growth factor receptor dependent neurite outgrowth and
N-cadherin and the fibroblast growth factor receptor co-cluster in cells. J Neurochem
76, 1421-30.
Van Itallie, C. M., and Anderson, J. M. (1997): Occludin confers adhesiveness when
expressed in fibroblasts. J Cell Sci 110 ( Pt 9), 1113-21.
Vassar, R., Chao, S. K., Sitcheran, R., Nunez, J. M., Vosshall, L. B., and Axel, R.
(1994): Topographic organization of sensory projections to the olfactory bulb. Cell 79,
981-91.

Literaturverzeichnis 76
Vaughn, D. E., and Bjorkman, P. J. (1996): The (Greek) key to structures of neural
adhesion molecules. Neuron 16, 261-73.
Vietor, I., Bader, T., Paiha, K., and Huber, L. A. (2001): Perturbation of the tight
junction permeability barrier by occludin loop peptides activates beta-
catenin/TCF/LEF-mediated transcription. EMBO Rep 2, 306-12.
Wang, F., Nemes, A., Mendelsohn, M., and Axel, R. (1998): Odorant receptors govern
the formation of a precise topographic map. Cell 93, 47-60.
Yagi, T., and Takeichi, M. (2000): Cadherin superfamily genes: functions, genomic
organization, and neurologic diversity. Genes Dev 14, 1169-80.
Yamagata, M., and Sanes, J. R. (1995): Lamina-specific cues guide outgrowth and
arborization of retinal axons in the optic tectum. Development 121, 189-200.
Yap, A. S., Brieher, W. M., and Gumbiner, B. M. (1997): Molecular and functional
analysis of cadherin-based adherens junctions. Annu Rev Cell Dev Biol 13, 119-46.
Yoshihara, Y., Katoh, K., and Mori, K. (1993): Odor stimulation causes disappearance
of R4B12 epitope on axonal surface molecule of olfactory sensory neurons.
Neuroscience 53, 101-10.
Yoshihara, Y., Kawasaki, M., Tamada, A., Fujita, H., Hayashi, H., Kagamiyama, H.,
and Mori, K. (1997): OCAM: A new member of the neural cell adhesion molecule
family related to zone-to-zone projection of olfactory and vomeronasal axons. J
Neurosci 17, 5830-42.


Danksagung
Frau Prof. Dr. Asan danke ich für die Überlassung des Themas und für ihre engagierte
Betreuung sowie ihre ständige Bereitschaft zur Diskussion und Unterstützung der
Arbeit. Besonders bedanken möchte ich mich auch bei Prof. Dr. Drenckhahn für seine
freundliche Unterstützung.
Weiterhin möchte ich mich bei allen Mitarbeitern des Instituts für Anatomie und
Zellbiologie für ihre Hilfsbereitschaft bedanken.

Lebenslauf Name Anna Sofia Meier-Stiegen Geburtsdatum 25. Januar 1975 Geburtsort Hamburg Konfession evangelisch-lutherisch Schulbildung 1981-1985 Grundschule Wasserburg am Inn 1985-1989 Luitpold-Gymnasium Wasserburg am Inn 1989-1991 Johanneum Lüneburg 1991-1994 Luitpold-Gymnasium Wasserburg am Inn 01. Juli 1994 Abiturprüfung Stipendium für Hochbegabte nach dem
Bayerischen Begabtenförderungsgesetz Praktika 10/1994 Archäologisches Praktikum Germanisches
Nationalmuseum Nürnberg 11/1994–03/1995 Pflegepraktikum Kreiskrankenhaus Wasserburg am Inn Universitäre Ausbildung 04/1995-10/2001 Studium der Humanmedizin (Universität Würzburg) 18. März 1997 Ärztliche Vorprüfung 24. März 1998 Erster Abschnitt der ärztlichen Prüfung 04. April 2000 Zweiter Abschnitt der ärztlichen Prüfung 17. Oktober 2001 Dritter Abschnitt der ärztlichen Prüfung Gesamtnote gut Famulaturen 04/1998 Chirurgie (Missionsärztliche Klink Würzburg) 09/1998 Pädiatrie (Kinderklinik am Mönchberg Würzburg) 10/1998 Dermatologie (Dres. Schubert, Frank, Beck-Bohlen,
Kristen, Würzburg) 03/1999 Pädiatrie (Universität Köln) 09/1999 Innere Medizin (Medizinische Poliklinik der Universität
Würzburg) Praktisches Jahr 04/2000-08/2000 Medizinische Poliklinik der Universität Würzburg 08/2000-12/2000 Chirurgische Universitätsklinik Würzburg 12/2000-03/2001 Pädiatrische Universitätsklinik Würzburg Ärztin im Praktikum Innere Medizin 11/2001-04/2003 Medizinische Poliklinik der Universität Würzburg Assistenzärztin Innere Medizin 08/2003-03/2004 Klinikum Straubing seit 04/2004 Krankenhaus Barmherzige Brüder Regensburg