Embed Size (px)
Citation preview

Brain Research 966 (2003) 194–205www.elsevier.com/ locate/brainres
Research report
L ocalization of the O-GlcNAc transferase and O-GlcNAc-modifiedproteins in rat cerebellar cortex
a b b aYoshihiro Akimoto , Frank I. Comer , Robert N. Cole , Akihiko Kudo ,a a,c b ,*Hayato Kawakami , Hiroshi Hirano , Gerald W. Hart
aDepartment of Anatomy, Kyorin University School of Medicine, Mitaka, Tokyo 181-8611, JapanbDepartment of Biological Chemistry, The Johns Hopkins University School of Medicine, Baltimore, MA 21205,USA
cNittai Jusei Medical College for Judo Therapeutics, Yoga, Setagaya, Tokyo 158-0097, Japan
Accepted 26 November 2002
Abstract
O-linked N-acetylglucosamine (O-GlcNAc) is a ubiquitous nucleocytoplasmic protein modification that has a complex interplay withphosphorylation on cytoskeletal proteins, signaling proteins and transcription factors. O-GlcNAc is essential for life at the single celllevel, and much indirect evidence suggests it plays an important role in nerve cell biology and neurodegenerative disease. Here we showthe localization of O-GlcNAc Transferase (OGTase) mRNA, OGTase protein, and O-GlcNAc-modified proteins in the rat cerebellarcortex. The sites of OGTase mRNA expression were determined by in situ hybridization histochemistry. Intense hybridization signalswere present in neurons, especially in the Purkinje cells. Fluorescent-tagged antibody against OGTase stained almost all of the neuronswith especially intense reactivity in Purkinje cells, within which the nucleus, perikaryon, and dendrites were most intensely stained. Usingimmuno-electron microscopic labeling, OGTase was seen to be enriched in euchromatin, in the cytoplasmic matrix, at the nerve terminal,and around microtubules in dendrites. In nerve terminals, immuno-gold labeling was observed around synaptic vesicles, with the enzymemore densely localized in the presynaptic terminals than in the postsynaptic ones. Using an antibody to O-GlcNAc, we found the sugarlocalizations reflected results seen for OGTase. Collectively, these data support hypothesized roles for O-GlcNAc in key processes ofbrain cells, including the regulation of transcription, synaptic vesicle secretion, transport, and signal transduction. Thus, by modulating thephosphorylation or protein associations of key regulatory and cytoskeletal proteins, O-GlcNAc is likely important to many functions ofthe cerebellum. 2002 Elsevier Science B.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporter, and receptors
Topic: Signal transduction
Keywords: O-GlcNAc transferase; O-GlcNAcylation; Purkinje cell; Synapse; Post-translational modification; Cerebellum; Phosphorylation; Signaling
1 . Introduction cytoplasmic proteins [31,66]. Phosphorylation and O-GlcNAc modification are often reciprocal, occurring at the
Phosphorylation of nerve terminal proteins and post- same or adjacent hydroxyl moieties [10,41]. O-GlcNAcsynaptic proteins plays an important role in neuronal modification, which is termed O-GlcNAcylation, is afunction, where it is involved in such processes as growth regulatory modification that has a complex dynamic inter-cone migration, synapse formation, neurotransmission, and play with O-phosphorylation (Fig. 1) [15,72].synaptic plasticity [39,65,67].O-linked N-acetylglucos- O-GlcNAc transferase (OGTase: EC 2-4-1) is a uniqueamine moieties (O-GlcNAc) are also dynamically attached nuclear and cytosolic glycosyltransferase that containsto serine or threonine residues of many nuclear and multiple tetratricopeptide repeats [30,42]. The liver en-
zyme contains two immunologically related subunits ofMr
110 kDa (a-subunit) and 78 kDa (b-subunit). Other*Corresponding author. Tel.:11-410-614-5993; fax:11-410-614-tissues, such as brain, contain only thea-subunit, which8804.
E-mail address: [email protected](G.W. Hart). contains the active site. Thea-subunit forms homo and
0006-8993/02/$ – see front matter 2002 Elsevier Science B.V. All rights reserved.doi:10.1016/S0006-8993(02)04158-6

195Y. Akimoto et al. / Brain Research 966 (2003) 194–205
cortex. These studies show that OGTase and O-GlcNAc-modified proteins are abundant in all neurons, but areparticularly enriched in the nucleus, perikaryon, dendrite,and synapses of the Purkinje cells. The high concentrationof O-GlcNAc-modified proteins in neurons may reflect animportant functional plasticity of neurons in the cere-bellum.
2 . Materials and methods
2 .1. Rat cerebellum
Cerebellum was extirpated from Wistar rats (male, 9–11weeks,n55). All experimental procedures using labora-tory animals were approved by the Animal Care and UseCommittee of Kyorin University School of Medicine. Allefforts were made to minimize their suffering.
2 .2. Antibodies
Fig. 1. Complex and dynamic interplay between O-GlcNAc and phos-Rabbit polyclonal anti-OGTase antibody (AL-25,phate modifications. This model depicts some of the possible combina-
purified IgG) was raised against a purified recombinanttions of O-GlcNAc and phosphate in the simple examples of a proteinwith two modification sites. Most of the O-GlcNAc-modified proteins 110-kDa subunit of OGTase expressed inEscherichia coliidentified to date have multiple sites of O-GlcNAc and phosphate either at [42]. AL-25 recognizes both 110-kDa and 78-kDa subunitsthe same site or at adjacent sites. The potential complexity of the
of OGTase. Moreover, OGTase enzymatic activity isinterplay between O-GlcNAc and phosphate may exert multiple levels ofprecipitated by AL-25 [42]. The generation of the mousecontrol.monoclonal anti-O-GlcNAc antibody (Mab CTD 110.6,
heterotrimers that appear to have different binding af- isotype: IgM) was previously described [16]. CTD 110.6finities for UDP-GlcNAc over the entire physiological specifically recognizes O-GlcNAc inb-O-glycosidic link-range [43]. The cDNA encoding the OGTasea-subunit has age to either serine or threonine and has no cross-reactivitybeen cloned from rat,Caenorhabditis elegans, and humans toward similar carbohydrate antigens or toward peptide[42,49]. OGT is highly conserved fromC. elegans to determinants [16]. Cy3-conjugated donkey anti-rabbit IgGhumans, and maps to the locus for X-linked Parkinsonia antibody and FITC-conjugated donkey anti-mouse IgG1
dystonia [62]. OGTase is unlike any glycosyltransferase IgM were obtained from Jackson Immunoresearch (Westpreviously described [35,57]. The brain has among the Grove, PA). Secondary antibodies (ultra small gold-conju-highest mRNA and protein levels of OGTase [42,58]. gated F(ab9) fragments of goat anti-rabbit IgG and goat2
O-GlcNAc is particularly abundant in the nerve terminal, anti-mouse IgG1IgM), acetylated bovine serum albuminand several of the hundreds of O-GlcNAc-modified pro- (BSA-c), coldwater fish skin gelatin (CWFS gelatin),teins in neurons have been recently identified, e.g., neuro- Enhancement Conditioning Solution (ECS), and R-gentfilaments, tau, synapsins, collapsin response mediator SE-EM electron microscopy grade silver enhancementprotein-2 (CRMP-2),b-synuclein, and ubiquitin-carboxyl reagent were purchased from Aurion (Wageningen, Thehydrolase-L1, (UCH-L1),b-amyloid precursor protein; Netherlands).synapse-specific clathrin assembly protein (AP-3), andankyrin G [5,13,14,18,19,28,52,80]. All of these proteins 2 .3. Western blot analysisare also phosphoproteins. Addition of O-GlcNAc is re-quired for life at the single cell level and for development Crude protein extracts were prepared from the dissected[62]. Collectively, the data strongly suggest that O- cerebellum of Wistar rats (males, 9–11 weeks). CerebellarGlcNAc is important to numerous neuronal functions. tissue was homogenized in a homogenization buffer (20
Here, we have extended these biochemical studies to the mM Tris–HCl, pH 7.4, containing 5 mM EDTA, 5 mMmorphological level, using DIG-labeled anti-sense cRNA EGTA, 1 mM DTT, 2 mM phenylmethylsulfonyl fluorideprobe, anti-OGTase and anti-O-GlcNAc antibodies for in [PMSF], and protease inhibitor cocktail [Pic]-1 and -2situ hybridization analysis, immunofluorescence and im- [1/1000 dilution]; [59]). The proteins were separated bymunoelectron microscopic experiments to localize OGTase 7.5% sodium dodecyl sulfate-polyacrylamide gel electro-mRNA, OGTase protein, and O-GlcNAc-modified pro- phoresis (SDS-PAGE) and transferred to a polyvinylideneteins, respectively, in sections of intact fixed rat cerebellar difluoride (PVDF) membrane. Purified rabbit polyclonal

Y. Akimoto et al. / Brain Research 966 (2003) 194–205196
IgG AL-25 (dilution, 1:5000) or mouse monoclonal IgM with 10 mM PBS, pH 7.4, and then with 4% form-CTD 110.6 (dilution, 1:5000) was used as the primary aldehyde-PBS at room temperature for 20 min. Theantibody; and goat anti-rabbit IgG or goat anti-mouse IgM cerebellum was immediately dissected out and placed incoupled to horseradish peroxidase (Amersham-Pharmacia 4% formaldehyde-PBS at 48C for 2 h. After having beenBiotech, Piscotaway, NJ), as the secondary antibody washed with PBS, it was cut into sections (40mm in(dilution, 1:20 000). For carbohydrate competition experi- thickness) with a microslicer DTD-1500 (DSK; Kyoto,ments 10 mMN-acetylglucosamine was included during Japan). After quenching of excess formaldehyde for 10the incubation with the primary antibody (CTD 110.6). min with 10 mM glycine in PBS, the sections were washedDetection of the horseradish peroxidase activity was three times with PBS and treated with 5% BSA-PBS for 1enhanced by using chemiluminescence (ECL), as described h. They were then incubated with AL25 (1:250) and CTDby the manufacturer (Amersham-Pharmacia Biotech). (1:500) in 0.1% BSA-PBS for 24 h at 48C, washed three
times for 20 min each with PBS, and then incubated with2 .4. Preparation of cRNA probes Cy3-conjugated donkey anti-rabbit IgG (1:500, Jackson
Immunoresearch) and FITC-conjugated donkey anti-mouseAntisense and sense RNA probes were prepared by in IgM (1:500, Jackson Immunoresearch). The nuclei were
vitro transcription from a fragment of OGTase cDNA stained with SYBR Green I or TOPRO-3 (Molecular(from nt 1193 to nt 1942 of rat OGTase cDNA sequence in Probes, Eugene, OR). After a wash with PBS, the speci-the GenBankTM/EBI Data Bank [accession number mens were mounted in 90% (v/v) glycerol–0.1 M Tris–U76557]) by use of T3 or T7 RNA polymerase in the HCl buffer (pH 8.5) containing 0.5 mMp-phenylenepresence of digoxigenin (DIG)-linked UTP in the reaction diamine and observed under a laser confocal scanningmixture as described previously [1]. microscope equipped with Ar /Kr ion lasers (MRC-1024,
BioRad).2 .5. In situ hybridization
2 .7. Double immunogold-silver particle labeling forIn situ hybridization was performed with digoxigenin electron microscopy
(DIG)-cRNA on cryosections as described earlier [1].Cryosections of unfixed rat cerebellum were cut at a 4-mm Double immunogold labeling was carried out accordingthickness, thaw-mounted onto silane-coated glass slides, to Yi et al. [78]. Fixation and sectioning were performedallowed to air dry, fixed in 4% paraformaldehyde-PBS for by the same method as described for the light microscopic1 h, incubated twice in 0.1% (w/v) DEPC-PBS (15 min observation. Sections were placed in 0.1 M phosphateeach time), and rinsed with DEPC-treated 43SSC. The buffer (PB, pH 7.4)–0.1% sodium borohydride for 15 minsections were then prehybridized in prehybridization buffer to inactivate aldehyde groups in the tissue sections, washed(50% formamide, 13Denhardt solution, 0.6 M NaCl, 10 with 0.1 M PB, treated with 0.1 M PB containing 0.05%mM Tris–HCl, 1 mM EDTA, 100 mg/ml salmon sperm Triton X-100 for 30 min, and then incubated in blockingDNA, 100 mg/ml yeast tRNA) at 458C for 3 h, and then solution (PBS, pH 7.4, containing 5% normal goat serumhybridized in hybridization buffer (50% [v/v] formamide, [NGS], 5% BSA, and 0.1% CWFS gelatin) for 1 h at 48C.13Denhardt solution, 0.6 M NaCl, 10 mM Tris–HCl [pH After blocking, the sections were incubated for 24 h at7.4], 1 mM EDTA, 100 mg/ml salmon sperm DNA, 0.4 48C in a mixture of rabbit anti-OGTase (AL25, dilutionmg/ml DIG-labeled probe RNA) at 508C for 15 h. The 1:250) and mouse anti-O-GlcNAc (CTD110.6, dilutionsections were next sequentially rinsed with DEPC-treated 1:250) antibodies diluted with PBS containing 0.2% BSA-43SSC at 458C for 15 min and with 50% formamide-23 c. After a wash with PBS-0.2% BSA-c, they were incu-SSC at 458C for 15 min, and then incubated in 1 mg/ml bated for 24 h at 48C in the first secondary antibody, i.e.RNase A solution at 378C for 15 min. Next, they were ultra small gold-conjugated F (ab9) fragments of goat2
rinsed with DEPC-treated 23SSC at 458C for 15 min, and anti-mouse IgG1IgM (dilution, 1:100) in PBS–0.2%then twice with DEPC-treated 0.53SSC at 458C for 15 BSA-c. Next, the sections were washed with PBS–0.2%min. The sections were incubated in a solution of polyclon- BSA-c, then with 0.1 M PB, and further washed with ECS.al sheep anti-DIG Fab antibody conjugated with alkaline They were thereafter incubated with R-gent SE-EM silver-phosphatase [1:500 diluted in DIG buffer (0.1 M Tris–HCl enhancement solution at room temperature for 90 min for[pH 9.5], 0.1 M NaCl, 50 mM MgCl )]. The signal was the first silver enhancement. After the enhancement had2
detected with nitro blue tetrazolium chloride (NBT) and been stopped by incubation with 0.03 M sodium thiosul-5-bromo-4-chloro-3-indolyl phosphate (BCIP, Boehringer- fate in ECS for 10 min, the sections were washed withMannheim). ECS, and then with 0.1 M PB. After the first silver
enhancement, the sections were incubated for 24 h at 48C2 .6. Immunostaining for light microscopic observation with the second secondary antibody, i.e. ultra small gold-
conjugated F(ab9) fragments of goat anti-rabbit IgG2
Under pentobarbital anesthesia, the rats were perfused (1:100) in PBS–0.2% BSA-c. Subsequently they were

197Y. Akimoto et al. / Brain Research 966 (2003) 194–205
washed with PBS–0.2% BSA-c, followed by 0.1 M PB,and then fixed with 2.5% glutaraldehyde in 0.1 M PB for 2h to prevent the loss of labeling. After the second silverenhancement had been carried out for 60 min, the sectionswere washed with 0.1 M PB, fixed with 0.5% O O for 1s 4
h, dehydrated by passage through a series of gradedalcohol, and flat-embedded in Epon 812. Ultrathin sectionswere cut, stained with lead citrate, and observed under atransmission electron microscope (JEM-1010; JEOL,Tokyo, Japan).
As cytochemical controls for double labeling of OGTaseand O-GlcNAc, the sections were incubated with eitherrabbit anti-OGTase antibody only, mouse anti-O-GlcNAcantibody only, or with preimmune rabbit IgG and preim-mune mouse IgM instead of primary antibodies. After theprimary antibody incubations, these cytochemical controlgroups were processed identically.
2 .8. Quantification and statistical analysis
The subcellular distribution of silver-enhanced goldparticles was analyzed quantitatively in sections. Surface
Fig. 2. Immunoblot analysis of cerebellum lysate by use of anti-OGTasesections, where the labeling was optimal, from differentantibody (AL25) and anti-O-GlcNAc antibody (CTD110.6). Normal
blocks were scanned with the electron microscope. Addi- cerebellar tissue was lysed, and the lysate was then electrophoresed on ational sections from tissues incubated in the absence of7.5% polyacrylamide gel and immunoblotted with AL25 (A) or with
CTD110.6 in the presence or absence of 10 mM GlcNAc (B). Theprimary antibodies were also scanned to assess the non-molecular masses of the marker proteins are indicated at the left margin.specific background labeling.
The negatives of EM photographs were digitized, andthe particle diameters were measured using NIH imagesoftware (National Institute of Health, Bethesda, MD). Atotal of .3000 particles were measured. The size N-GlcNAc, all immunoreactivity was specifically lost due(mean6S.D.) of once-enhanced particles representing OG- to the competition. Using these antibodies, we performedTase localization was 10.6063.51 nm (n52888), and the imunofluorescence staining and immunoelectron micro-size of twice-enhanced particles representing O-GlcNAc scopy on OGTase and O-GlcNAc.localization was 26.0167.56 nm (n5948). The differencein the size was significant by Student’st-test (P,0.001). 3 .2. In situ hybridization
2Areas of 1mm were selected from the nucleus (n525),perikaryon (n517), and dendrites (n519) of Purkinje cells, The localization of OGT mRNA in sections of ratand from the axons (n584), presynaptic nerve terminals cerebellar cortex was examined by in situ hybridization(n594), and postsynaptic nerve terminals (n590) in the histochemistry using the DIG-labeled antisense RNA probemolecular layer. The numbers of the gold-silver particles (Fig. 3). The rat cerebellar cortex contains three dis-were measured by using NIH image software. Statistical tinguishable layers. The outermost layer is molecular layeranalysis was performed with Student’st-test. (indicated as M in Fig. 3) that is composed of axons of
granule cells, neurons (stellate and basket cells) and thedendrites of the underlying Purkinje cells. Beneath the
3 . Results molecular layer is the Purkinje cell layer, which consists ofa single cell layer of the large cell bodies of Purkinje cells
3 .1. Characterization of the antibodies (AL25 and (indicated as P in Fig. 3). The innermost layer is granularCTD110.6) layer that contains a large number of small neurons, called
granule cells (indicated as G in Fig. 3). The hybridizationThe specificity of the antibodies was examined by signals were observed in the neurons of all these three
Western-blot analysis of crude cerebellum lysate with anti- layers. Strong staining was observed in the Purkinje cells.OGTase antibody (AL25) and anti-O-GlcNAc antibody Neurons in the molecular layer and Golgi cells in the(CTD110.6, Fig. 2). The AL-25 antibody detected a single granular layer were moderately stained. Granule cells were110-kDa band, whereas CTD110.6 detected many proteins. weakly stained. When an equal concentration of the senseWhen CTD110.6 was reacted in the presence of 10 mM RNA probe was used instead of the antisense riboprobe, no

Y. Akimoto et al. / Brain Research 966 (2003) 194–205198
Fig. 3. OGTase mRNA is expressed in the neurons in the rat cerebellar cortex. Nomarski differential interference images. Distribution of OGTase mRNAexamined by in situ hybridization histochemistry using DIG-labeled anti-sense cRNA probe (A, B). (A) Low magnification. (B) Enlargement of rectanglein A. Intense hybridization signals are seen in Purkinje cells (red arrows in ‘B’). Moderate signals are seen in neurons (blue arrowheads) in the molecularlayer (M) and in Golgi cells (green arrowheads) in the granular cell layer (G). Weak hybridization signals are seen in granule cells. (C) No hybridizationsignals are observed with DIG-labeled sense cRNA probe. Red arrows in ‘C’ indicate the Purkinje cells. Scale bars, 100mm.
positive signals were observed. These results indicate that3 .4. Immuno-electron microscopic localization ofOGT mRNA is expressed in all neurons, but is especially OGTase in the cerebellar cortexenriched in the Purkinje cells.
To further observe the precise subcellular co-localizationof OGTase and O-GlcNAc in Purkinje cells, we carried out
3 .3. Light microscopic localization of OGTase and O- the new procedure for double immunogold-silver particleGlcNAc-modified proteins in the cerebellar cortex labeling described by Yi et al. [78]. Different-size particles
(diameter, 11 and 26 nm) could be used to distinguishVarious anti-O-GlcNAc antibodies have been generated, between the localization of OGTase and that of O-GlcNAc
such as HGAC85, RL2 [64,68], but there are some (Fig. 5). The localization of the OGTase was almost theproblems with the specificity of the antibodies [16]. We same as that of O-GlcNAc. In the Purkinje cells, bothraised a monoclonal antibody (CTD110.6) that reacts small and large particles were densely localized in thespecifically with O-GlcNAc [16]. We have now used this euchromatin of the nucleus (Fig. 5A). Particle labels wereanti-O-GlcNAc antibody and anti-OGTase antibody to also seen in the nuclear pores. The cytoplasmic matrix waslocalize the major O-GlcNAc-modified proteins and OG- less densely labeled with particles than the nucleus. ThereTase in cerebellar cortex. Reactivities for O-GlcNAc were few particles in the endoplasmic reticulum (Fig. 5A)showed a similar distribution patterns as that for OGTase and Golgi body (data not shown). In the dendrites of(Fig. 4). Reactivities for both OGTase and O-GlcNAc Purkinje cells, many particles were observed along thewere observed in almost all of the neurons. Especially microtubules (Fig. 5B). Some particles were seen in theintense reactivity was observed in the Purkinje cells, in presynaptic terminal of a climbing fiber (Fig. 5B). In thewhich the nucleus, perikaryon, and dendrites were stained synaptic region where the basket cell axon makes contact(Fig. 4B–D). Neurons in the molecular layer and Golgi with the soma of the Purkinje cell, particles were seen incells in the granular layer were moderately stained (Fig. the basket cell axon terminal (Fig. 5C). In some synapses4B–D) Granule cells showed a weak staining pattern. in the molecular layer many particles were seen aroundThese results of the immunofluorescence study were synaptic vesicles in the presynaptic nerve terminals (Fig.similar to those of the in situ hybridization. In the control 5D). Particles were also seen in the postsynaptic nerveexperiment, in which the primary antibody (AL25 and terminals (Fig. 5D) and in the axons (data not shown) inCTD110.6) was omitted or replaced with the pre-immune the molecular layer. In control samples from which one ofrabbit IgG or mouse IgM, no positive staining was the primary antibodies had been omitted, there were noobserved (data not shown). particles of relevant size (data not shown). In other control
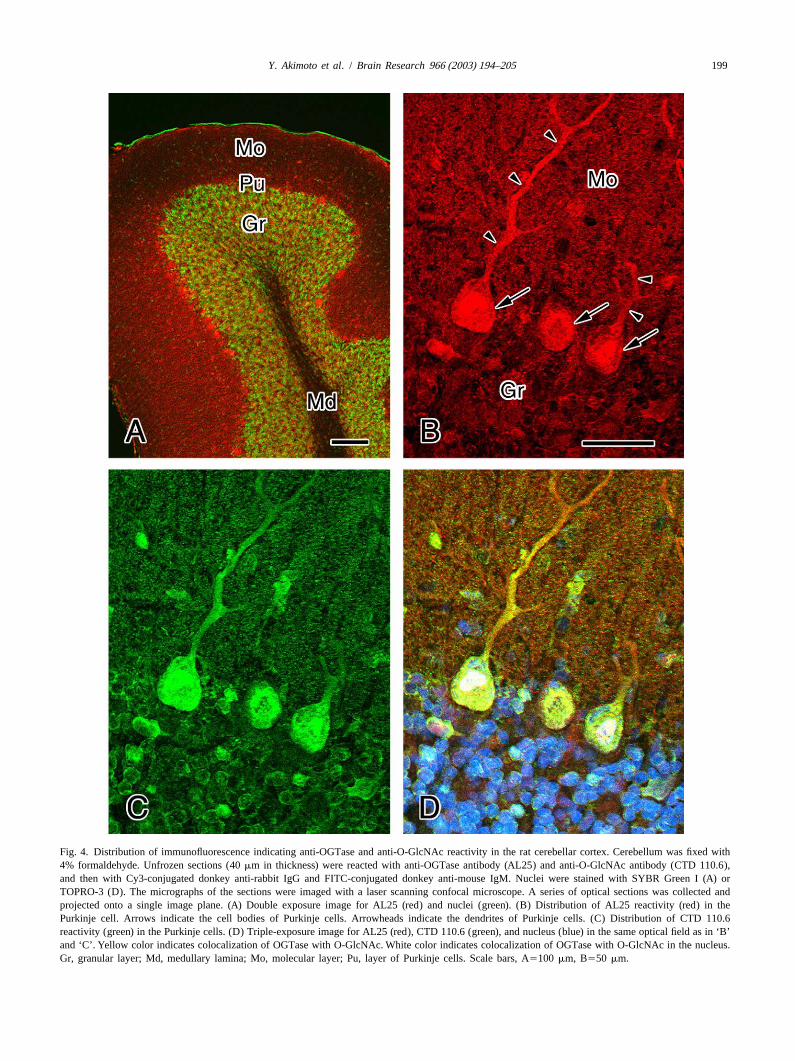
199Y. Akimoto et al. / Brain Research 966 (2003) 194–205
Fig. 4. Distribution of immunofluorescence indicating anti-OGTase and anti-O-GlcNAc reactivity in the rat cerebellar cortex. Cerebellum was fixed with4% formaldehyde. Unfrozen sections (40mm in thickness) were reacted with anti-OGTase antibody (AL25) and anti-O-GlcNAc antibody (CTD 110.6),and then with Cy3-conjugated donkey anti-rabbit IgG and FITC-conjugated donkey anti-mouse IgM. Nuclei were stained with SYBR Green I (A) orTOPRO-3 (D). The micrographs of the sections were imaged with a laser scanning confocal microscope. A series of optical sections was collected andprojected onto a single image plane. (A) Double exposure image for AL25 (red) and nuclei (green). (B) Distribution of AL25 reactivity (red) in thePurkinje cell. Arrows indicate the cell bodies of Purkinje cells. Arrowheads indicate the dendrites of Purkinje cells. (C) Distribution of CTD 110.6reactivity (green) in the Purkinje cells. (D) Triple-exposure image for AL25 (red), CTD 110.6 (green), and nucleus (blue) in the same optical field as in ‘B’and ‘C’. Yellow color indicates colocalization of OGTase with O-GlcNAc. White color indicates colocalization of OGTase with O-GlcNAc in the nucleus.Gr, granular layer; Md, medullary lamina; Mo, molecular layer; Pu, layer of Purkinje cells. Scale bars, A5100 mm, B550 mm.

Y. Akimoto et al. / Brain Research 966 (2003) 194–205200
Fig. 5. Subcellular distribution of OGTase and O-GlcNAc immunoreactivities in the rat cerebellar cortex detected by using pre-embedding double-immunogold-silver particle-labeling. Small particles represent OGTase localization, while large particles represent O-GlcNAc localization. (A) Enlargedview of nucleus and cytoplasm in the Purkinje cell. Both small and large gold particles are densely colocalized in the euchromatin of the nucleus (N). Somenuclear pores (arrowheads) are labeled with particles. Particles are also colocalized in the cytoplasmic matrix, but less densely than in the nucleus. (B)Numerous particles are found along microtubules in the dendrites (D) of this Purkinje cell. Some gold particles are seen in the presynaptic terminal(asterisk) of a climbing fiber. (C) Synaptic region where a basket cell axon contacts the soma of a Purkinje cell (P). Particles are seen around synapticvesicles (arrowheads) in the axon terminal of the basket cell (asterisk). (D) Synapses in the molecular layer. Many particles are seen in the presynapticterminals (T). Particles are also seen in the postsynaptic terminal (asterisk). E, endoplasmic reticulum, M, mitochondrion. Scale bars, A–D, 100 nm.
samples from which both primary antibodies had been GlcNAc particle labeling in subcellular locations, weomitted, only a few background particles were randomly counted particles visible in electron micrographs of nu-distributed (data not shown). cleus, perikaryon, and dendrites of Purkinje cells and
To provide a quantitative comparison of OGT and O- axons, presynaptic terminals, and postsynaptic terminals in

201Y. Akimoto et al. / Brain Research 966 (2003) 194–205
where most O-GlcNAcylated proteins are found, stainedmore intensely than the cytoplasm. Furthermore, the resultsof immunoelectron microscopy confirmed the existence ofOGTase in the nucleus, especially in the euchromatin.
Several post-translational modifications in the nucleus,such as phosphorylation, methylation, acetylation, andubiquitination on specific amino acid residues have beenwell studied [24,39,46,54–56,71,75,79]. These modifica-tions have been shown to be directly involved in manyimportant regulatory processes. It is now clear that dy-namic O-GlcNAcylation of nuclear and cytosolic proteinsis as abundant and widespread as phosphorylation [26,72].To date only a small fraction of the O-GlcNAc modifiedproteins have been identified [13,14,69], including RNApolymerase II, its associated transcription factors, chro-
Fig. 6. Quantitative analysis of the subcellular distribution of OGTasematin-associated proteins, nuclear oncogene and tumor(A) and O-GlcNAc (B) particle labeling. Density of particles in thesuppressor proteins, and steroid receptors [14]. We earliernucleus (number of observations,n525), perikaryon (n517), dendrite
(n519) of Purkinje cells and in axons (n584), presynaptic terminals proposed that O-GlcNAcylation of nuclear proteins is(n594), and postsynaptic terminals (n590) in the molecular layer. More involved in various aspects of gene expression [14]. Wethan 3000 particles counted were counted. Results are shown as thealso showed that OGTase was predominantly localized in2particle number (mean6S.E.M.) in the unit area (1mm ) examined after
the euchromatin in pancreatic cells and aortic smooththe subtraction of the background labeling. The values for nucleus andmuscle cells [1,3], and is involved in the regulation ofpresynaptic terminal are significantly larger than all other values (P,
0.001). transcription [14,15]. The present immunohistochemicalstudy indicates that O-GlcNAcylation may also correlatewith the transcriptionally active state of genes in the
the molecular layer (Fig. 6). Both OGT- and O-GlcNAc- neuron.labeling particles were more densely localized in nucleus The present observation that O-GlcNAc is abundant inof Purkinje cell and in the presynaptic terminals than in the the nuclear pore is consistent with previous biochemicalperikaryon, dendrites, axons, and postsynaptic terminals [33] and WGA-immunohistochemical [63] findings. The(P,0.001). nucleoporins are the major structural proteins of the
nuclear pore complex and are involved in the assembly ofthe nuclear pore and nuclear transport through the pore
4 . Discussion [22]. The fact that nucleoporins p62, p58, and p54, whichare the components of a multisubunit complex, are exten-
Nucleocytoplasmic O-GlcNAc modification is dynamic sively modified by O-GlcNAc [33] suggests that O-and abundant, exhibiting characteristics more like those of GlcNAc modification could be involved in the nuclear porephosphorylation than those of typicalN- and O-linked assembly /disassembly or import of molecules into theglycosylation [8,9,40,60]. Although O-GlcNAc modifica- nucleus [44]. More interestingly, our present result that nottion of proteins is potentially involved in many cellular only O-GlcNAc but also OGTase is localized in theprocesses and several disease states, including Alzheimer’s nuclear pores suggests that O-GlcNAc modification ofdisease and diabetes [2,3,15,47,70,72] and several key nucleoporins may occur at the nuclear pore complex orproteins important in neurodegenerative disease, including that proteins that are synthesized in the cytoplasm andtau, AP-3, andb-amyloid precursor proteins have been transported into the nucleus could be O-GlcNAc modifieddemonstrated to be modified by O-GlcNAc [5,28,76,77], at the nuclear pore during transit.the molecular functions of O-GlcNAc are only beginning We also detected OGTase enriched around the micro-to be understood. Renewed interest in O-GlcNAc’s po- tubules in the dendrites. Cytoskeletal proteins such astential roles in neurodegenerative diseases has also been MAPs, neurofilaments, tau, ankyrin G, talin, synapsins,stimulated by the finding that the gene for O-GlcNAcase etc. are O-GlcNAcylated and also phosphorylated[26], the enzyme which removes O-GlcNAc, maps to the [5,6,12,15,17–19,29,50,80]. Thus, we hypothesize thatlocus for late-onset Alzheimer’s Disease [7]. OGTase may be involved in the modulation of neuro-
transmitter transport by regulating the O-GlcNAcylation/4 .1. OGTase and O-GlcNAc-modified proteins in O-phosphorylation of cytoskeletal proteins in conjunction
21nucleus, perikaryon, and dendrites of neurons with specific kinases such as Ca /calmodulin kinase II(Cam K II) or protein kinase C (PKC; [12]). In Alzheim-
The present study shows that OGTase is localized in er’s disease, the highly phosphorylated forms of tau, inboth the nucleus and cytoplasm of neurons. The nucleus, which O-GlcNAc modification is reduced, accumulate and
Y. Akimoto et al. / Brain Research 966 (2003) 194–205202
tend to precipitate as insoluble filamentous inclusions multiply O-GlcNAcylated near neighboring phosphoryla-(neurofibrillar tangle), which leads to block of the axonal tion sites within domains involved in synaptic vesicletransport and cell death. binding and in nucleating actin. O-GlcNAcylation of
synapsin I in the presynaptic terminal may regulate the4 .2. OGTase and O-GlcNAc-modified proteins in nerve binding and anchoring of synaptic vesicles to actin.terminals The localization and possible function of the OGTase in
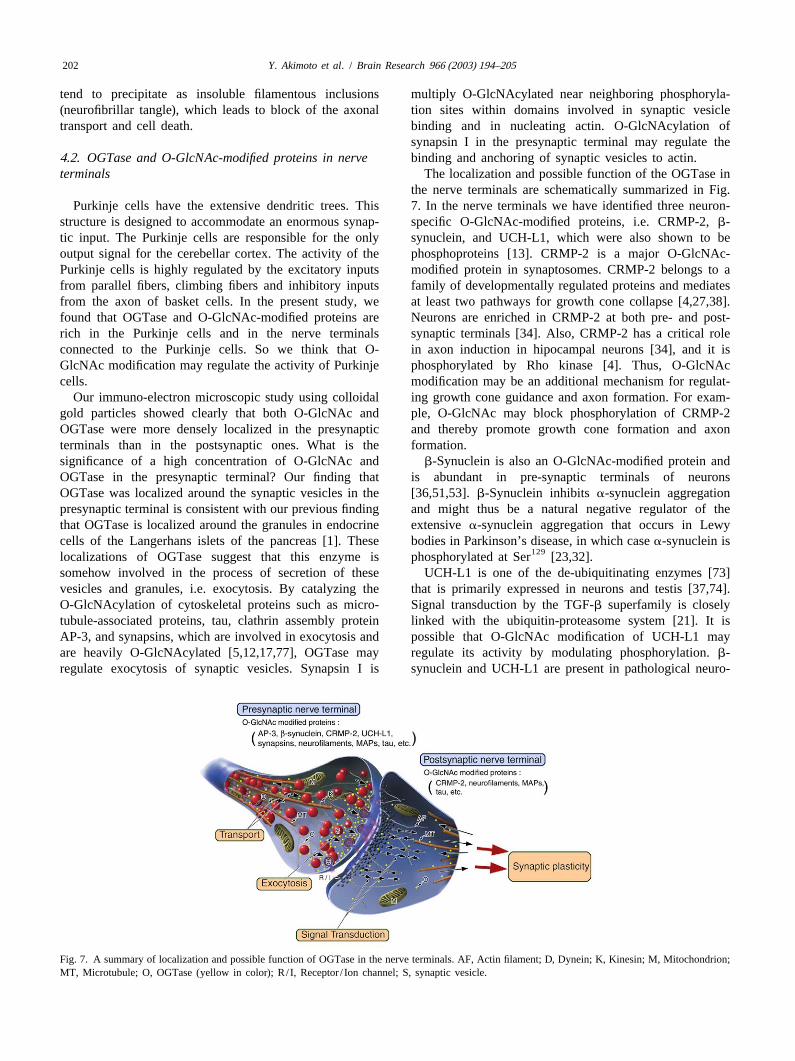
the nerve terminals are schematically summarized in Fig.Purkinje cells have the extensive dendritic trees. This 7. In the nerve terminals we have identified three neuron-
structure is designed to accommodate an enormous synap- specific O-GlcNAc-modified proteins, i.e. CRMP-2,b-tic input. The Purkinje cells are responsible for the only synuclein, and UCH-L1, which were also shown to beoutput signal for the cerebellar cortex. The activity of the phosphoproteins [13]. CRMP-2 is a major O-GlcNAc-Purkinje cells is highly regulated by the excitatory inputs modified protein in synaptosomes. CRMP-2 belongs to afrom parallel fibers, climbing fibers and inhibitory inputs family of developmentally regulated proteins and mediatesfrom the axon of basket cells. In the present study, we at least two pathways for growth cone collapse [4,27,38].found that OGTase and O-GlcNAc-modified proteins are Neurons are enriched in CRMP-2 at both pre- and post-rich in the Purkinje cells and in the nerve terminals synaptic terminals [34]. Also, CRMP-2 has a critical roleconnected to the Purkinje cells. So we think that O- in axon induction in hipocampal neurons [34], and it isGlcNAc modification may regulate the activity of Purkinje phosphorylated by Rho kinase [4]. Thus, O-GlcNAccells. modification may be an additional mechanism for regulat-
Our immuno-electron microscopic study using colloidal ing growth cone guidance and axon formation. For exam-gold particles showed clearly that both O-GlcNAc and ple, O-GlcNAc may block phosphorylation of CRMP-2OGTase were more densely localized in the presynaptic and thereby promote growth cone formation and axonterminals than in the postsynaptic ones. What is the formation.significance of a high concentration of O-GlcNAc and b-Synuclein is also an O-GlcNAc-modified protein andOGTase in the presynaptic terminal? Our finding that is abundant in pre-synaptic terminals of neuronsOGTase was localized around the synaptic vesicles in the [36,51,53].b-Synuclein inhibitsa-synuclein aggregationpresynaptic terminal is consistent with our previous finding and might thus be a natural negative regulator of thethat OGTase is localized around the granules in endocrine extensivea-synuclein aggregation that occurs in Lewycells of the Langerhans islets of the pancreas [1]. These bodies in Parkinson’s disease, in which casea-synuclein is
129localizations of OGTase suggest that this enzyme is phosphorylated at Ser [23,32].somehow involved in the process of secretion of these UCH-L1 is one of the de-ubiquitinating enzymes [73]vesicles and granules, i.e. exocytosis. By catalyzing the that is primarily expressed in neurons and testis [37,74].O-GlcNAcylation of cytoskeletal proteins such as micro- Signal transduction by the TGF-b superfamily is closelytubule-associated proteins, tau, clathrin assembly protein linked with the ubiquitin-proteasome system [21]. It isAP-3, and synapsins, which are involved in exocytosis and possible that O-GlcNAc modification of UCH-L1 mayare heavily O-GlcNAcylated [5,12,17,77], OGTase may regulate its activity by modulating phosphorylation.b-regulate exocytosis of synaptic vesicles. Synapsin I is synuclein and UCH-L1 are present in pathological neuro-
Fig. 7. A summary of localization and possible function of OGTase in the nerve terminals. AF, Actin filament; D, Dynein; K, Kinesin; M, Mitochondrion;MT, Microtubule; O, OGTase (yellow in color); R/ I, Receptor / Ion channel; S, synaptic vesicle.

203Y. Akimoto et al. / Brain Research 966 (2003) 194–205
G.W. Hart, The microtubule-associated protein tau is extensivelynal structures characteristic of neurodegenerative diseasemodified with O-linked N-acetylglucosamine, J. Biol. Chem. 271[11,20,25,48]. Missense mutations in the UCH-L1 gene(1996) 28741–28744.
cause Parkinson’s disease [45]. In-frame deletion in the [6] C.S. Arnold, G.W. Hart, Dynamic cytoskeletal glycosylation andUCH-L1 gene causes gracile axonal dystrophy [61]. These neurodegenerative disease, Trends Glycosci. Glycotech. 11 (1999)O-GlcNAc-modified proteins are probably correlated with 355–370.
[7] L. Bertram, D. Blacker, K. Mullin, D. Keeney, J. Jones, S. Basu, S.neurodegenerative disorders. The abundance and localiza-Yhu, M.G. Mcinnis, R.C.P. Go, K. Vekrellis, D.J. Selkoe, A.J.tion of OGTase and O-GlcNAc in neurons, especially inSaunders, R.E. Tanzi, Evidence for Genetic Linkage of Alzheimer’sPurkinje cells, suggest that O-GlcNAc modification, likeDisease to Chromosome 10q, Science 290 (2000) 2302–2303.
phosphorylation, plays an important role in neurons. By [8] C.-F. Chou, M.B. Omary, Mitotic arrest-associated enhancement ofregulating phosphorylation of various synaptic proteins, O-linked glycosylation and phosphorylation of human keratins 8 and
18, J. Biol. Chem. 268 (1993) 4465–4472.OGTase may be involved in the signal transduction that is[9] C.-F. Chou, A.J. Smith, M.B. Omary, Characterization and dy-important for neuronal function such as synaptic plasticity,
namics ofO-linked glycosylation of human cytokeratin 8 and 18, J.long-term depression, etc. (Fig. 7). O-GlcNAc modifica-Biol. Chem. 267 (1992) 3901–3906.
tion will be a subject of continued studies to further our [10] T.-Y. Chou, C.V. Dang, G.W. Hart, Glycosylation of the c-Mycknowledge about the biology of the neuron and to under- transactivation domain, Proc. Natl. Acad. Sci. USA 92 (1995)
4417–4421.stand its relationship to neurodegenerative diseases.[11] D.F. Clayton, J.M. George, Synucleins in synaptic plasticity and
neurodegenerative disorders, J. Neurosci. Res. 58 (1999) 120–129.[12] R.N. Cole, G.W. Hart, Glycosylation sites flank phosphorylation
A cknowledgements sites on synapsin I: O-linkedN-acetylglucosamine residues arelocalized within domains mediating synapsin I interactions, J.Neurochem. 73 (1999) 418–428.This work was supported in part by a National Institute
[13] R.N. Cole, G.W. Hart, Cytosolic O-glycosylation is abundant inof Health Grant HD13563 awarded to G.W. Hart. Thisnerve terminals, J. Neurochem. 79 (2001) 1080–1089.
work was also supported in part by Grants-in Aid for [14] F.I. Comer, G.W. Hart, O-GlcNAc and the control of gene expres-Scientific Research (10877004 awarded to Y. Akimoto) sion, Biochim. Biophys. Acta 1473 (1999) 161–171.
[15] F.I. Comer, G.W. Hart,O-glycosylation of nuclear and cytosolicfrom the Ministry of Education, Culture, Sports, Science,proteins. Dynamic interplay betweenO-GlcNAc and O-phosphate,and Technology, Japan.J. Biol. Chem. 275 (2000) 29179–29182.We wish to express our appreciation to Mr M. Fukuda,
[16] F.I. Comer, K. Vosseller, L. Wells, M.A. Accavitti, G.W. Hart,Ms S. Matsubara, Ms M. Kanai, and Ms S. Shibata Characterization of a mouse monoclonal antibody specific for O-(Laboratory for Electron Microscopy and Department of linked N-acetylglucosamine, Anal. Biochem. 293 (2001) 169–177.
´[17] M. Ding, D.D. Vandre, High molecular weight microtubule-associ-Anatomy, Kyorin University School of Medicine) for theirated proteins contain O-linkedN-acetylglucosamine, J. Biol. Chem.technical assistance.271 (1996) 12555–12561.Under a licensing agreement between Covance Research
[18] D.L.-Y. Dong, Z.-S. Xu, M.R. Chevrier, R.J. Cotter, D.W. Cleveland,Products and Hoffman-La Roche and The Johns Hopkins G.W. Hart, Glycosylation of mammalian neurofilaments. Localiza-University, G.W. Hart and F.I. Comer receive a share of tion of multiple O-linkedN-acetylglucosamine moieties on neuro-
filament polypeptides L and M, J. Biol. Chem. 268 (1993) 16679–royalty received by the university on sales of the CTD16687.110.6 antibody. The terms of this arrangement are being
[19] D.L.-Y. Dong, Z.-S. Xu, G.W. Hart, D.W. Cleveland, Cytoplasmicmanaged by The Johns Hopkins University in accordanceO-GlcNAc modification of the head domain and the KSP repeat
with its conflict of interest policies. motif of the neurofilament protein neurofilament-H, J. Biol. Chem.271 (1996) 20845–20852.
[20] J.E. Duda, V.M.-Y. Lee, J.Q. Trojanowski, Neuropathology ofsynuclein aggregates: New insights into mechanisms of neurode-
R eferences generative disease, J. Neurosci. Res. 61 (2000) 121–127.[21] T. Ebisawa, M. Fukuchi, G. Murakami, T. Chiba, K. Tanaka, T.
[1] Y. Akimoto, L.K. Kreppel, H. Hirano, G.W. Hart, Localization of the Imamura, K. Miyazono, Smurf1 interacts with transforming growthO-linked N-acetylglucosamine transferase in rat pancreas, Diabetes factor-b type I receptor through Smad7 and induces receptor48 (1999) 2407–2413. degradation, J. Biol. Chem. 276 (2001) 12477–12480.
[2] Y. Akimoto, L.K. Kreppel, H. Hirano, G.W. Hart, Increased O- [22] D.R. Finlay, E. Meier, P. Bradley, J. Horecka, D.J. Forbes, AGlcNAc transferase in pancreas of rats with streptozotocin-induced complex of nuclear pore proteins required for pore function, J. Celldiabetes, Diabetologia 43 (2000) 1239–1247. Biol. 114 (1991) 169–183.
[3] Y. Akimoto, L.K. Kreppel, H. Hirano, G.W. Hart, Hyperglycemia [23] H. Fujiwara, M. Hasegawa, N. Dohmae, A. Kawashima, E. Masliah,and the O-GlcNAc transferase in rat aortic smooth muscle cells: M.S. Goldberg, J. Shen, K. Takio, T. Iwatsubo, Alpha-synuclein isElevated expression and altered patterns of O-GlcNAcylation, Arch. phosphorylated in synucleinopathy lesions, Nat. Cell Biol. 4 (2002)Biochem. Biophys. 389 (2001) 166–175. 160–164.
´[4] N. Arimura, N. Inagaki, K. Chihara, C. Menager, N. Nakamura, M. [24] M. Fukuchi, T. Imamura, T. Chiba, T. Ebisawa, M. Kawabata, K.Amano, A. Iwamatsu, Y. Goshima, K. Kaibuchi, Phosphorylation of Tanaka, K. Miyazono, Ligand-dependent degradation of Smad3 by acollapsin response mediator protein-2 by Rho-kinase. Evidence for ubiquitin ligase complex of ROC1 and associated proteins, Mol.two separate signalling pathways for growth cone collapse, J. Biol. Biol. Cell 12 (2001) 1431–1443.Chem. 275 (2000) 23973–23980. [25] J.E. Galvin, K. Uryu, V.M.-Y. Lee, J.Q. Trojanowski, Axon pathol-
[5] C.S. Arnold, G.V.W. Johnson, R.N. Cole, D.L.-Y. Dong, M. Lee, ogy in Parkinson’s disease and Lewy body dementia hippocampus

Y. Akimoto et al. / Brain Research 966 (2003) 194–205204
containsa-, b-, and g-synuclein, Proc. Natl. Acad. Sci. USA 96 Hindsgaul (Eds.), Molecular and Cellular Glycobiology, OxfordUniversity Press, New York, 2000, pp. 199–219.(1999) 13450–13455.
[45] E. Leroy, R. Boyer, G. Auburger, B. Leube, G. Ulm, E. Mezey, G.[26] Y. Gao, L. Wells, F.I. Comer, G.J. Parker, G.W. Hart, DynamicHarta, M.J. Brownstein, S. Jonnalagada, T. Chernova, A. Dehejia, C.O-glycosylation of nuclear and cytosolic proteins. Cloning andLavedan, T. Gasser, P.J. Steinbach, K.D. Wilkinson, M.H. Poly-characterization of neutral, cytosolicb-N-acetylglucosaminidasemeropoulos, The ubiquitin pathway in Parkinson’s disease [letter],from human brain, J. Biol. Chem. 276 (2001) 9838–9845.Nature 395 (1998) 451–452.[27] Y. Goshima, F. Nakamura, P. Strittmatter, S.M. Strittmatter, Collap-
[46] M.D. Litt, M. Simpson, M. Gaszner, C.D. Allis, G. Felsenfeld,sin-induced growth cone collapse mediated by an intracellularCorrelation between histone lysine methylation and developmentalprotein related to UNC-33, Nature 376 (1995) 509–514.changes at the chickenb-globin locus, Science 293 (2001) 2453–[28] L.S. Griffith, M. Mathes, B. Schmitz,b-amyloid precursor protein is2455.modified with O-linkedN-acetylglucosamine, J. Neurosci. Res. 41
[47] K. Liu, A.J. Paterson, E. Chin, J.E. Kudlow, Glucose stimulates(1995) 270–278.protein modification byO-linked GlcNAc in pancreaticb cells:[29] J. Hagmann, M. Grob, M.M. Burger, The cytoskeletal protein talinLinkage ofO-linked GlcNAc tob cell death, Proc. Natl. Acad. Sci.is O-glycosylated, J. Biol. Chem. 267 (1992) 14424–14428.USA 97 (2000) 2820–2825.
[30] R.S. Haltiwanger, M.A. Blomberg, G.W. Hart, Glycosylation of[48] J. Lowe, H. McDermott, M. Landon, R.J. Mayer, K.D. Wilkinson,
nuclear and cytoplasmic proteins, J. Biol. Chem. 267 (1992) 9005– .Ubiquitin carboxyl-terminal hydrolase (PGP 9 5) is selectively9013.
present in ubiquitinated inclusion bodies characteristic of human[31] G.W. Hart, Dynamic O-linked glycosylation of nuclear and cyto- neurodegenerative diseases, J. Pathol. 161 (1990) 153–160.
skeletal proteins, Annu. Rev. Biochem. 66 (1997) 315–335. [49] W.A. Lubas, D.W. Frank, M. Krause, J.A. Hanover,O-linked[32] M. Hashimoto, E. Rockenstein, M. Mante, M. Mallory, E. Masliah, GlcNAc transferase is a conserved nucleocytoplasmic protein con-
b-synuclein inhibitsa-synuclein aggregation: A possible role as an taining tetratricopeptide repeats, J. Biol. Chem. 272 (1997) 9316–anti-Parkinsonian factor, Neuron 32 (2001) 213–223. 9324.
[33] G.D. Holt, C.M. Snow, A. Senior, R.S. Haltiwanger, L. Gerace, ¨ ¨[50] T. Luthi, R.S. Haltiwanger, P. Greengard, M. Bahler, SynapsinsG.W. Hart, Nuclear pore complex glycoproteins contain cytoplasmi- contain O-linked N-acetylglucosamine, J. Neurochem. 56 (1991)cally disposed O-linkedN-acetyl-glucosamine, J. Cell Biol. 104 1493–1498.(1987) 1157–1164. [51] L. Maroteaux, J.T. Campanelli, R.H. Scheller, Synuclein: A neuron-
´[34] N. Inagaki, K. Chihara, N. Arimura, C. Menager, Y. Kawano, N. specific protein localized to the nucleus and presynaptic nerveMatsuo, T. Nishimura, M. Amano, K. Kaibuchi, CRMP-2 induces terminal, J. Neurosci. 8 (1988) 2804–2815.axons in cultured hippocampal neurons, Nat. Neurosci. 4 (2001) [52] J.-E. Murphy, J.A. Hanover, M. Froehlich, G. DuBois, J.H. Keen,781–782. Clathrin assembly protein AP-3 is phosphorylated and glycosylated
[35] D.H. Joziasse, Mammalian glycosyltransferases: genomic organiza- on the 50-kDa structural domain, J. Biol. Chem. 269 (1994) 21346–tion and protein structure, Glycobiology 2 (1992) 271–277. 21352.
¨[36] P.J. Kahle, M. Neumann, L. Ozmen, V. Muller, H. Jacobsen, A. [53] D.D. Murphy, S.M. Rueter, J.Q. Trojanowski, V.M.-Y. Lee, Synu-Schindzielorz, M. Okochi, U. Leimer, H. van der Putten, A. Probst, cleins are developmentally expressed, anda-synuclein regulates theE. Kremmer, H.A. Kretzschmar, C. Haass, Subcellular localization size of the presynaptic vesicular pool in primary hippocampalof wild-type and Parkinson’s disease-associated mutanta-synuclein neurons, J. Neurosci. 20 (2000) 3214–3220.in human and transgenic mouse brain, J. Neurosci. 20 (2000) [54] J. Nakayama, J.C. Rice, B.D. Strahl, C.D. Allis, S.I.S. Grewal, Role6365–6373. of histone H3 lysine 9 methylation in epigenetic control of hetero-
[37] Y. Kajimoto, T. Hashimoto, Y. Shirai, N. Nishino, T. Kuno, C. chromatin assembly, Science 292 (2001) 110–113.Tanaka, c DNA cloning and tissue distribution of a rat ubiquitin [55] S.J. Nielsen, R. Schneider, U.-M. Bauer, A.J. Bannister, A. Mor-carboxyl-terminal hydrolase PGP9.5, J. Biochem. 112 (1992) 28– rison, D. O’Carroll, R. Firestein, M. Cleary, T. Jenuwein, R.E.32. Herrera, T. Kouzarides, Rb targets histone H3 methylation and HP1
[38] T. Kamata, M. Subleski, Y. Hara, N. Yuhki, H. Kung, N.G. to promoters, Nature 412 (2001) 561–565.Copeland, N.A. Jenkins, T. Yoshimura, W. Modi, T.D. Copeland, [56] K. Noma, C.D. Allis, S.I.S. Grewal, Transitions in distinct histoneIsolation and characterization of a bovine neural specific protein H3 methylation patterns at the heterochromatin domain boundaries,(CRMP-2) cDNA homologous tounc-33, a C. elegans gene Science 293 (2001) 1150–1155.implicated in axonal outgrowth and guidance, Mol. Brain Res. 54 [57] J.C. Paulson, K.J. Colley, Glycosyltransferases. Structure, localiza-(1998) 219–236. tion, and control of cell type-specific glycosylation, J. Biol. Chem.
[39] H. Kang, L.D. Sun, C.M. Atkins, T.R. Soderling, M.A. Wilson, S. 264 (1989) 17615–17618.Tonegawa, An important role of neural activity-dependent CaMKIV [58] M. Rex-Mathes, S. Werner, D. Strutas, L.S. Griffith, C. Viebahn, K.signaling in the consolidation of long-term memory, Cell 106 (2001) Thelen, B. Schmitz, O-GlcNAc expression in developing and ageing771–783. mouse brain, Biochimie 83 (2001) 583–590.
[40] K.P. Kearse, G.W. Hart, Lymphocyte activation induces rapid [59] E.P. Roquemore, T.-Y. Chou, G.W. Hart, Detection ofO-linkedchanges in nuclear and cytoplasmic glycoproteins, Proc. Natl. Acad. N-acetylglucosamine (O-GlcNAc) on cytoplasmic and nuclear pro-Sci. USA 88 (1991) 1701–1705. teins, Methods Enzymol. 230 (1994) 443–460.
[41] W.G. Kelly, M.E. Dahmus, G.W. Hart, RNA polymerase II is [60] E.P. Roquemore, M.R. Chevrier, R.J. Cotter, G.W. Hart, Dynamicglycoprotein: modification of the COOH-terminal domain byO- O-GlcNAcylation of the small heat shock proteinaB-crystallin,GlcNAc, J. Biol. Chem. 268 (1993) 10416–10424. Biochemistry 35 (1996) 3578–3586.
[42] L.K. Kreppel, M.A. Blomberg, G.W. Hart, Dynamic glycosylation of [61] K. Saigoh, Y.L. Wang, J.G. Suh, T. Yamanishi, Y. Sakai, H.nuclear and cytosolic proteins. Cloning and characterization of a Kiyosawa, T. Harada, N. Ichihara, S. Wakana, T. Kikuchi, K. Wada,unique O-GlcNAc transferase with multiple tetratricopeptide re- Intragenic deletion in the gene encoding ubiquitin carboxy-terminalpeats, J. Biol. Chem. 272 (1997) 9308–9315. hydrolase in gad mice, Nat. Genet. 23 (1999) 47–51.
[43] L.K. Kreppel, G.W. Hart, Regulation of a cytosolic and nuclear [62] R. Shafi, S.P.N. Iyer, L.G. Ellies, N. O’Donnell, K.W. Marek, D.O-GlcNAc transferase. Role of the tetratricopeptide repeats, J. Biol. Chui, G.W. Hart, J.D. Marth, The O-GlcNAc transferase geneChem. 274 (1999) 32015–32022. resides on the X chromosome and is essential for embronic stem cell
[44] L.K. Kreppel, G.W. Hart, Functional significance of O-GlcNAc viability and mouse ontogeny, Proc. Natl. Acad. Sci. 97 (2000)glycoproteins in the nucleus and cytoplasm, in: M. Fukuda, O. 5735–5739.

205Y. Akimoto et al. / Brain Research 966 (2003) 194–205
[63] D.M. Snow, G.W. Hart, Nuclear and cytoplasmic glycosylation, Int. [72] L. Wells, K. Vosseller, G.W. Hart, Glycosylation of nucleocytoplas-Rev. Cytol. 181 (1998) 43–74. mic proteins: Signal transduction and O-GlcNAc, Science 291
[64] C.M. Snow, A. Senior, L. Gerace, Monoclonal antibodies identify a (2001) 2376–2378.group of nuclear pore complex glycoproteins, J. Cell Biol. 104 [73] K.D. Wilkinson, K. Lee, S. Deshpande, P. Duerksen-Hughes, J.M.(1987) 1143–1156. Boss, J. Pohl, The neuron-specific protein PGP 9.5 is a ubiquitin
[65] T.R. Soderling, V.A. Derkach, Postsynaptic protein phosphorylation carboxyl-terminal hydrolase, Science 246 (1989) 670–673.and LTP, Trends Neurosci. 23 (2000) 75–80. [74] K.D. Wilkinson, S. Deshpande, C.N. Larsen, Comparisons of
[66] C.-R. Torres, G.W. Hart, Topography and polypeptide distribution of neuronal (PGP 9.5) and non-neuronal ubiquitin C-terminal hydro-terminal N-acetylglucosamine residues on the surfaces of intact lases, Biochem. Soc. Trans. 20 (1992) 631–637.lymphocytes. Evidence forO-linked GlcNAc, J. Biol. Chem. 259 [75] W. Xu, H. Chen, K. Du, H. Asahara, M. Tini, B.M. Emerson, M.(1984) 3308–3317. Montminy, R.M. Evans, A transcriptional switch mediated by
[67] K.M. Turner, R.D. Burgoyne, A. Morgan, Protein phosphorylation cofactor methylation, Science 294 (2001) 2507–2511.and the regulation of synaptic membrane traffic, Trends Neurosci. [76] P.J. Yao, P.D. Coleman, ReducedO-glycosylated clathrin assembly22 (1999) 459–464. protein AP180: implication for synaptic vesicle recycling dysfunc-
[68] J.R. Turner, A.M. Tartakoff, N.S. Greenspan, Cytologic assessment tion in Alzheimer’s disease, Neurosci. Lett. 252 (1998) 33–36.of nuclear and cytoplasmicO-linked N-acetylglucosamine distribu- [77] P.J. Yao, P.D. Coleman, Reduction of O-linkedN-acetylglucosamine-tion by using anti-streptococcal monoclonal antibodies, Proc. Natl. modified assembly protein-3 in Alzheimer’s disease, J. Neurosci. 18Acad. Sci. USA 87 (1990) 5608–5612. (1998) 2399–2411.
[69] K. Vosseller, L. Wells, G.W. Hart, NucleocytoplasmicO-glycosyla- [78] H. Yi, J.L.M. Leunissen, G.M. Shi, C.A. Gutekunst, S.M. Hersch, Ation: O-GlcNAc and functional proteomics, Biochimie 83 (2001) novel procedure for pre-embedding double-immunogold-silver label-575–581. ing at the ultrastructural level, J. Histochem. Cytochem. 49 (2001)
[70] K. Vosseller, L. Wells, M.D. Lane, G.W. Hart, Elevated nucleocytop- 279–283.lasmic glycosylation by O-GlcNAc results in insulin resistance [79] K. Zanger, S. Radovick, F.E. Wondisford, CREB binding proteinassociated with defects in Akt activation in 3T3-L1 adipocytes, recruitment to the transcription complex requires growth factor-Proc. Natl. Acad. Sci. USA 99 (2002) 5313–5318. dependent phosphorylation of its GF box, Mol. Cell 7 (2001)
[71] H. Wang, Z.-Q. Huang, X. Li, Q. Feng, H. Erdjument-Bromage, 551–558.B.D. Strahl, S.D. Briggs, C.D. Allis, J. Wong, P. Tempst, Y. Zhang, [80] X. Zhang, V. Bennett, Identification ofO-linked N-Methylation of histone H4 at arginine 3 facilitating transcriptional acetylglucosamine modification of ankirin isoforms targeted toG
activation by nuclear hormone receptor, Science 293 (2001) 853– nodes of Ranvier, J. Biol. Chem. 271 (1996) 31391–31398.857.





























