Embed Size (px)
Citation preview

UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
INSTITUTO DE BIOLOGIA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA
ECOLOGIA E CONSERVAÇÃO DA BORBOLETA DA PRAIA,
PARIDES ASCANIUS (CRAMER, 1775)
(LEP: PAPILIONIDAE)
ELISA VIEIRA HERKENHOFF
ORIENTADOR: DR. RICARDO FERREIRA MONTEIRO
Dissertação de Mestrado apresentada ao Programa de Pós-Graduação em Ecologia, Instituto de Biologia, Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários à obtenção do título de Mestre em Ecologia.
RIO DE JANEIRO, RJ - BRASIL
MARÇO DE 2006

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

ii
FICHA CATALOGRÁFICA
HERKENHOFF, ELISA VIEIRA
Ecologia e conservação da borboleta-da-praia, Parides ascanius (Cramer, 1775)
(Lep: Papilionidae) [Rio de Janeiro] 2006
80 p. 29,7 cm (Instituto de Biologia/UFRJ, M.Sc., Ecologia, 2006)
Dissertação – Universidade Federal do Rio de Janeiro, PPGE
1. Poço das Antas 2. Biologia populacional 3. Parides ascanius
I. IB/UFRJ II. Título (série)

iii
Elisa Vieira Herkenhoff
Ecologia e conservação da borboleta-da-praia,
Parides ascanius (Cramer, 1775) (Lep: Papilionidae)
Rio de Janeiro, 31 de março de 2006
__________________________________________________
Ricardo Ferreira Monteiro, Doutor em Ecologia, UFRJ
__________________________________________________
Ricardo Iglesias Rios, Doutor em Ecologia, UFRJ
__________________________________________________
Onildo João Marini-Filho, Doutor em Ecologia, Conservação e Manejo da
Vida Silvestre, IBAMA

iv
AGRADECIMENTOS
Eu gostaria de agradecer, primeiramente, ao que muitos chamam de Deus; para
mim, o milagre da VIDA, aquele mistério que a ciência é incapaz de compreender.
Agradeço imensamente a minha mãe, meu pai e meu irmão, e à minha família pelo
incondicional amor, carinho e apoio ao longo de toda minha existência. Tenho que abusar
da redundância biológica, mas nada seria sem vocês.
Aos meus amigos, pelo carinho e amizade. Pela convivência e troca que me tornam
a cada dia uma pessoa melhor. Pela ajuda e compreensão nos momentos difíceis.
Ao meu orientador Ricardo e à Margarete, pelos anos de convívio e aprendizado. E
também pela paciência e boa vontade diante das minhas inconstâncias e devaneios.
A todos os amigos do Laboratório de Ecologia de Insetos: Abacaxi, Adriana, Carol,
Cristina, Diana, Jéssica, Jiraya, João, Karine, Luana, Luciano, Milena, Pedro, Pimenta,
Rafael, Raquel, Ronaldo, Sama, Thiago, Vagner, Vivian e Viviane. Ao longo de todos esses
anos o ambiente de trabalho não poderia ter sido mais agradável. Foram tantas ajudas,
colaborações e salvamentos que daria uma dissertação.
A todos aqueles que me ajudaram com as capturas em campo: Pimenta, Rafael,
Vagner e Ricardo Flores. A ajuda de vocês foi de fundamental importância para a
realização desse trabalho. E a companhia, valiosíssima. Muito obrigada!
Ao pessoal do Laboratório de Ecologia e Conservação de Populações pelas caronas
para dentro da área de estudo e pela ótima convivência em campo. Em especial, agradeço à

v
Camila Barros pela disposição em ajudar e por me passar os dados climáticos.
Ao Roberto Eizemberg, por sua incomensurável colaboração na realização do
documentário (fez quase sozinho), e por sua amizade. Não tenho palavras para agradecer e
nem descrever essa fantástica pessoa. A ciência agradece.
À Márcia Franco, pela gravação do áudio do documentário, pela viagem para campo
e pelas fotografias. Definitivamente, também se trata de uma pessoa rara no meio científico.
Ao Prof. Luiz Otero, por todos os anos de aprendizado e convivência, e imensa
receptividade. Por acender em mim a paixão pelas borboletas e, em especial, por Parides
ascanius. Agradeço também pelas primordiais sugestões para o trabalho e documentário.
A Eleonora Moura e Lívia Ferreira pela ajuda com a bibliografia, além de é claro,
pela amizade mencionada no terceiro parágrafo.
Ao Programa de Pós-Graduação em Ecologia (UFRJ), aos professores, alunos e
funcionários.
Ao Prof. Fernando Fernandez e ao Prof. Ricardo Iglesias pelas sugestões na tese e
por suas brilhantes aulas.
Ao CNPq pela bolsa.
Ao IBAMA/Reserva Biológica de Poço das Antas pela autorização para realização
do trabalho e pelo suporte em campo.

vi
“O que a lagarta
chama de fim do mundo o mestre chama de borboleta.”
Richard Bach

vii
RESUMO GERAL
Parides ascanius (Cramer, 1775), a borboleta da praia, foi a primeira espécie de
inseto a figurar como ameaçada de extinção no Brasil. Endêmica do Rio de Janeiro, ela
ocorre em pequenas manchas de vegetação brejosa ou pantanosa, e tem como única planta
hospedeira Aristolochia macroura (Aristolochiaceae). Até 1984 havia na Reserva Biológica
de Poços das Antas pelo menos 1000 hectares do habitat ideal dessa borboleta, mas com a
construção da barragem de Juturnaíba houve grande modificação dessa paisagem. O
capítulo I trata da avaliação e dinâmica do tamanho de uma população de P. ascanius na
ReBio. O capítulo II retrata a produção de um documentário sobre a biologia e ecologia
dessa borboleta, que é apresentado no CD em anexo. O método para avaliação da biologia
populacional foi o de captura,marcação e recaptura. Também eram anotados o sexo do
indivíduo, seu dano alar, faixa etária, horário e localização da captura. A população se
reproduziu o ano inteiro, porém o número de indivíduos capturados por dia variou muito ao
longo do ano. A razão sexual foi desviada para machos (3:1), porém houve grande variação
entre as amostras. As capturas não ocorreram de forma regular ao longo do transecto,
existindo claramente trechos de maior ocupação. A população de P. ascanius estudada
parece estar em boas condições na reserva, uma vez que é abundante ao longo de boa parte
do ano. Essa população está localizada em uma área bastante favorável, com uma grande
quantidade dos recursos alimentares das lagartas e dos adultos. A região ocupada fornece
ainda microclimas bastante variados em uma área relativamente pequena, facilitando a
permanência de indivíduos no local.

viii
GENERAL ABSTRACT
Parides ascanius (Cramer, 1775) was the first species of insect to appear as
threatened of becoming extinct in Brazil. Endemic of Rio de Janeiro, it occurs in small
spots of marshy vegetation, and has as only host plant Aristolochia macroura
(Aristolochiaceae). Until 1984 there were at least 1000 hectares of the ideal habitat of this
butterfly in the Biological Reserve of Poço das Antas, but with the construction of the
Juturnaíba dam, there was a great modification of this landscape, without no posterior
ecological study. Chapter I deals with the evaluation and dynamics of the size of a
population of P. ascanius in the Reserve. Chapter II portraits the production of a
documentary on the biology and ecology of this butterfly, that is presented in the CD in
annex. The method for evaluation of population biology was of capture, marking and
recapture. Also were noted the sex of the individual, its age class, wing damage, and the
time and place of capture. The studied population reproduced the entire year, however the
number of individuals captured per day varied a lot. The sexual reason of the population
was deviated for males (3:1), however had a great variation between the samples. The
captures did not occurr regularly along the transect, existing stretches of clearly higher
occupation. The population of P. ascanius seems to be in good condition in the Reserve,
since that it is abundant throughout the year. This population is located in a favorable area,
with a great amount of the feeding resources of the juvenile stages and also for the adults.
The area of occurrence supplies varied microclimates in a relatively small area, facilitating
the permanence of individuals in the place.

ix
LISTA DE ILUSTRAÇÕES
Figura 2.1. Esquema ilustrativo da localização da ReBio Poço das Antas....................8
Figura 2.2. Foto da região da lagoa e barragem de Juturnaíba, indicando a localização da
área de estudo..........................................................................................................10
Figura 3.1. Recorte de imagem de satélite (IKONOS) representando a área de estudo e seu
entorno....................................................................................................................20
Figura 3.2. Recorte de imagem de satélite (IKONOS) representando a área de estudo e o
transecto em que foram realizadas as capturas......................................................21
Figura 3.3. Fotografia da área de estudo, mostrando bem a separação das regiões de
“morrote” e “baixada”.................................................................................................22
Figura 3.4. Fotografia da região do “morrote”............................................................23
Figura 3.5. Fotografia da região da “baixada”.............................................................23
Figura 3.6. Adulto de Parides ascanius.......................................................................24
Figura 3.7. Marcação nas asas de P. ascanius.............................................................26
Figura 3.8. Média de indivíduos capturados por dia em cada mês estudado...............30
Figura 3.9. Razão entre número de machos para cada fêmea em cada um dos meses
estudados.....................................................................................................................32
Figura 3.10. Percentual de espécimens de cada faixa etária entre todos os indivíduos, entre
machos e entre fêmeas........................................................................................33

x
Figura 3.11. Percentual de indivíduos de cada faixa etária em cada um dos meses
estudados.....................................................................................................................34
Figura 3.12. Percentual de espécimens com nenhum, pouco ou muito dano alar entre todos
os indivíduos, entre machos e entre fêmeas.......................................................35
Figura 3.13. Número de indivíduos capturados em cada um dos trechos de 100 metros do
transecto..................................................................................................................36
Figura 3.14. Percentual de capturas de machos e fêmeas nos diferentes trechos do
transecto.......................................................................................................................37
Figura 3.15. Percentual de ocupação de trechos do transecto por machos e fêmeas de
diferentes faixas etárias................................................................................................38
Figura 3.16. Percentual de recapturas nos trechos do transecto..................................38
Figura 3.17. Número de recapturas nas diferentes classes de distância......................39
Figura 3.18. Percentual de cada faixa etária entre os indivíduos recapturados...........40
Figura 3.19. Percentual de cada faixa etária de indivíduos recapturados em cada uma das
classes de distância................................................................................................40
Figura 3.20. Percentual de indivíduos de cada faixa etária recapturado em cada uma das
principais classes de distância...............................................................................41
Figura 3.21. Percentual de espécimens de cada classe de dano alar entre os indivíduos
recapturados.................................................................................................................42
Figura 3.22. Percentual de cada classe de dano alar de indivíduos recapturados em cada

xi
uma das classes de distância................................................................................42
Figura 3.23. Percentual de indivíduos de cada classe de dano alar recapturado em cada uma
das principais classes de distância...............................................................43
Figura 3.24. Número de indivíduos recapturados em cada classe de dias após a captura
levando em consideração o esforço................................................................44
Figura 3.25. Recorte de imagem de satélite (IKONOS) representando a área de estudo, o
transecto e a mancha de Aristolochia macroura..........................................46
Figura 4.1. Márcia Franco e Roberto Eizemberg filmando uma cópula no sítio de Luiz
Otero............................................................................................................................63
Figura 4.2. Câmera, iluminação e cenário preparados para a filmagem da emergência do
adulto......................................................................................................................64

xii
1.3 LISTA DE TABELAS
Tabela 2.1. Dados climáticos dos meses estudados.......................................................9
Tabela 3.1. Regressões entre abundância de indivíduos em cada sessão de marcação e
recaptura e os fatores climáticos..................................................................................30
Tabela 3.2. Número de indivíduos por dia de captura (NICD), percentagem de indivíduos
recapturados, estimativa de tamanho populacional pelo método de Lincoln-Petersen e
intervalo de confiança da estimativa, em cada um dos meses
estudados......................................................................................................................31
Tabela 3.3. Dobro do número de machos, estimativa populacional pelo método de Lincoln-
Peterson (LP) e LP/NICD, em cada um dos meses estudados.........................32
Tabela 3.4. Ocorrência de outras espécies de papilionídeos na área estudada ao longo do
período de estudo....................................................................................................45

xiii
SUMÁRIO
1. INTRODUÇÃO GERAL................................................................................................1
2. ÁREA DE ESTUDO......................................................................................................7
3. BIOLOGIA POPULACIONAL DE PARIDES ASCANIUS.................................................12
3.1 INTRODUÇÃO..............................................................................................13
3.2 MATERIAIS E MÉTODOS..............................................................................20
3.3 RESULTADOS..............................................................................................29
3.3.1 Variação temporal e tamanho populacional................................29
3.3.2 Razão sexual................................................................................32
3.3.3 Distribuição etária e dano alar.....................................................33
3.3.4 Ocupação do habitat e movimentação.........................................35
3.3.5 Longevidade................................................................................43
3.3.6 Conservação................................................................................44
3.4 DISCUSSÃO.................................................................................................47
3.4.1 Variação temporal e tamanho populacional................................47
3.4.2 Razão sexual................................................................................48
3.4.3 Distribuição etária e dano alar.....................................................49
3.4.4 Ocupação do habitat e movimentação.........................................50
3.4.5 Longevidade................................................................................53
3.4.6 Conservação................................................................................54
3.5 CONCLUSÕES..............................................................................................56

xiv
4. DOCUMENTÁRIO “PARIDES ASCANIUS: O RESGATE DE UMA ESPÉCIE”.................58
4.1 INTRODUÇÃO..............................................................................................59
4.2 MATERIAS E MÉTODOS...............................................................................63
4.3 RESULTADOS..............................................................................................65
4.4 DISCUSSÃO.................................................................................................66
5. CONSIDERAÇÕES FINAIS.........................................................................................68
6. BIBLIOGRAFIA GERAL..............................................................................................73

xv

xvi
1. INTRODUÇÃO GERAL
Dentro de sua área geográfica, a distribuição de determinada espécie é irregular em
diversos graus devido à variação das características físicas, da disponibilidade de recursos e
das espécies competidoras, predadoras ou parasitas, e também a eventos históricos de
colonização e extinção. Muitas espécies, particularmente aquelas que possuem
necessidades bem restritas, consistem de algumas a muitas populações pequenas que estão,
com freqüência, amplamente isoladas geograficamente. O reconhecimento de que uma
espécie está dividida em populações locais que normalmente consistem de algumas dezenas
ou centenas de indivíduos, teve um impacto importante sobre o entendimento dos processos
evolutivos. Além do mais, populações pequenas são muito susceptíveis de extinção, e a
persistência de uma espécie em um local particular depende muitas vezes do influxo
contínuo de migrantes de outras populações (Futuyma, 1997).
A densidade e permanência da população de uma espécie em qualquer local
dependem de sua capacidade de aumentar numericamente e dos fatores que limitam sua
abundância. A compreensão dessa dinâmica é essencial para uma apreciação adequada da
teoria evolutiva (Ricklefs 1979).
Através de sucintas revisões da literatura sobre extinções, Simberloff (1986) e
Caughley (1994) distinguiram processos que tornam as populações raras em um primeiro
momento (o que Simberloff chama de “causas finais da extinção” e Caughley denomina
“paradigma das populações em declínio”) de processos que podem finalmente causar a
extinção (o que Simberloff denomina “causas próximas da extinção” e Caughley chama de
“paradigma das populações pequenas”).

xvii
As causas próximas da extinção são aquelas pelas quais populações pequenas se
extinguem, mesmo quando protegidas, incluindo a aleatoriedade demográfica,
aleatoriedade ambiental, deterioração genética e disfunção social. Entre as causas últimas
da extinção, que tornam as espécies raras, podem ser citadas a caça, destruição de habitat,
mudanças climáticas e poluição.
Como para grande parte dos outros organismos, a destruição e degradação de
hábitats estão entre os fatores de declínio mais importantes para comunidades de borboletas
(New, 1997). Há muito tempo, as atividades humanas vêm alterando os ambientes naturais,
e hoje a maior parte dos sistemas encontra-se transformada ou com diferentes graus de
fragmentação. Essa fragmentação apresenta como principais componentes a perda do
habitat original, a redução do tamanho e o aumento do isolamento das manchas desse
habitat (Wilcox & Murphy 1985; Saunders et al., 1991), todos exercendo efeitos diretos e
indiretos sobre a biota. Atualmente, a fragmentação florestal é um dos processos que mais
influencia a perda de diversidade e o aumento das taxas de extinção de espécies em várias
escalas (Pimm & Raven, 2000).
O entendimento de padrões e processos das mudanças das paisagens, assim como as
respostas de populações a essas modificações são de fundamental importância para que os
esforços de conservação biológica tenham êxito (Collinge, 2001). A extinção natural de
populações é apenas um dos aspectos das extinções de borboletas atualmente. O outro é a
tendência unidirecional e virtualmente global de perda de populações e espécies de
borboletas como um resultado da expansão da ocupação e atividade humana (Ehrlich et al.,
1980).
Algumas espécies podem ser naturalmente raras, embora estas também possam se
tornar ainda mais raras pela ação do homem, sendo incerto se as espécies naturalmente

xviii
raras e as antropogenicamente raras difiram de maneira fundamental em relação aos riscos
de extinção por causas próximas (Lawton, 1994).
Espécies filogeneticamente predispostas a baixas densidades populacionais e
distribuições geográficas pequenas são mais vulneráveis a causas últimas da extinção, como
exploração e destruição de hábitats. Paradoxalmente, estas podem também ser menos
ameaçadas pelas causas próximas da extinção, pois são pré-adaptadas à vida como
populações pequenas em reservas isoladas, comparadas com espécies outrora abundantes e
com amplamente distribuídas (Kunin & Gaston, 1993).
Mudanças nos hábitats confrontam muitas espécies especialistas com condições
apenas sub-ótimas para sua sobrevivência (Meyer-Hozak, 2000). O acesso a múltiplas
espécies de plantas hospedeiras, aumenta a exposição de espécies para a expansão de
estratégias, promovendo novas oportunidades evolutivas e saída da especialização. A perda
de variabilidade no uso de plantas hospedeiras com a perda de hábitats implica no
contrário: vulnerabilidade à extinção. Embora exista uma tendência de mudanças climáticas
atuais induzirem uma maior utilização de recursos em borboletas (Dennis et al., 1993;
Thomas et al., 2001), isto está sendo contrabalançado pela perda de hábitats.
No Brasil, das 55 espécies de borboletas ameaçadas de extinção, 26 pertencem à
família Nymphalidae, 8 são Hesperiidae, 2 Lycaenidae, 4 Pieridae, 6 Riodinidae e 9
Papilionidae (MMA, 2003). Estima-se que muitas espécies e subespécies de Lepidoptera
estejam ameaçadas de extinção devido à destruição e fragmentação que vem ocorrendo no
sudeste brasileiro (Brown Jr., 1992, 1997). Aproximadamente 2/3 das espécies de
borboletas que ocorrem no Brasil são representadas na região da Mata Atlântica, sendo que
uma grande parte dessas populações encontra-se hoje em fragmentos de menos de 1000
hectares (Brown & Freitas, 2000).

xix
Devido à sua elevada diversidade de espécies associada à alta taxa de endemismo e
a baixíssima proporção de florestas remanescentes, a Mata Atlântica é considerada um “hot
spot” que tem prioridade máxima para conservação (Myers et al., 2000). Esse bioma já
representou cerca de 12% da cobertura do território nacional e hoje apresenta-se reduzido a
aproximadamente 7,6% da sua ocupação original (MMA, 1999).
Segundo a lista do MMA (2003) de espécies ameaçadas de extinção, 57% (55) dos
insetos que se encontram nessa situação no Brasil são borboletas. O estado do Rio de
Janeiro conta com 56% (31) das borboletas que aparecem na lista, sendo muitas vezes o
único estado onde essas borboletas ocorrem. Isso provavelmente ocorre pelo alto grau de
endemismo que caracteriza os insetos da mata atlântica, assim como pelo elevado nível de
fragmentação em que se encontra esse ecossistema neste estado.
Parides ascanius (Cramer, 1775), a borboleta da praia, foi a primeira espécie de
inseto a figurar como ameaçada de extinção no Brasil. Em 31 de maio de 1973, a agência
brasileira de parques nacionais (através do IBDF, Instituto Brasileiro para o
Desenvolvimento Florestal) colocou Parides ascanius (usando o sinônimo anterior, Battus
orophobus D’Almeida) na lista oficial dos animais brasileiros ameaçados de extinção,
através da Portaria 3481-DN. Atualmente, P. ascanius aparece nas listas estadual na
categoria “criticamente em perigo” (Otero et al., 2000), nacional, sugerida “em perigo”
(MMA 2003), e internacional na categoria “vulnerável” (IUCN 2003).
Endêmica do Rio de Janeiro, ela ocorre em pequenas manchas de vegetação brejosa
ou pantanosa entre Atafona (São João da Barra) e Itaguaí (Rio de Janeiro), e tem como
única planta hospedeira Aristolochia macroura (Aristolochiaceae) (Otero, 1984; Otero &
Brown Jr., 1986; Otero & Marigo, 1990).

xx
Até 1984 havia na Reserva Biológica de Poços das Antas pelo menos 1000 hectares
do habitat original dessa espécie, em várias manchas conectadas (Otero & Brown, 1986).
Com a construção da barragem de Juturnaíba, em 1984, houve grande modificação de boa
parte das áreas de distribuição dessa borboleta na Reserva, sem que tenha havido nenhum
estudo ecológico posterior sobre o assunto.
O capítulo I trata do estudo do tamanho de uma população de P. ascanius na
Reserva Biológica de Poço das Antas, considerando além da variação temporal dos
indivíduos, sua proporção sexual e distribuição etária. Além disso, serão analisados também
a longevidade e o dano alar dos espécimens, a ocupação do habitat e a movimentação dos
indivíduos neste.
No capítulo II produziu-se um documentário sobre a biologia e ecologia de P.
ascanius, que é apresentado no CD em anexo. Nesse capítulo, haverá uma breve exposição
da situação da documentação científica no Brasil, da metodologia utilizada para o
desenvolvimento do audiovisual e dos objetivos pretendidos com a divulgação da mídia.

xxi

xxii
2. ÁREA DE ESTUDO
Localizada na parte central costeira do estado do Rio de Janeiro, a Reserva
Biológica de Poço das Antas estende-se por cerca de 5.500 hectares dentro do município de
Silva Jardim (22º30' e 22º33'S e 42º15' e 42º19'W Gr.), fazendo limite com Casimiro de
Abreu e Araruama (Figura 2.1).
O clima é do tipo quente e úmido com temperaturas médias anuais elevadas durante
quase todo o ano. A temperatura média anual é de 25,7ºC, sendo fevereiro geralmente o
mês mais quente e julho o mais frio. O índice pluviométrico anual é, em média, 2.000mm,
concentrados nos meses de outubro a abril, sendo julho e agosto os meses mais secos,
segundo os dados climáticos dos últimos 10 anos cedidos pela Associação Mico-Leão-
Figura 2.1. Esquema ilustrativo da localização da ReBio Poço das Antas (retirado de www.ibama.gov.br)

xxiii
Dourado. Durante os 12 meses estudados a precipitação foi de 2.815mm e a temperatura
média foi de 25,14°C, sendo um ano um pouco mais frio e chuvoso do que a média (Tabela
2.1).
O seu relevo é classificado em sete compartimentos geomorfológicos que são: faixa
meândrica (8,7%), várzea interna (8,7%), várzea externa (38,5%), alvéolos (2,6%),
morrotes baixos (6,3%), morrotes altos (25,9%) e morros (9,3%). Cerca de 60% de toda a
área de até 5% de declividade encontra-se sob saturação hídrica praticamente todo o ano
(IBAMA, 2004).
Uma das últimas manchas de floresta na baixada fluminense, a Reserva de Poço das
Antas abriga uma grande faixa de vegetação arbórea típica de Floresta Pluvial Atlântica
Baixo Montana ou também designada Floresta Ombrófila Densa Sub-montana em
diferentes estados de maturidade (Programa Mata Atlântica, 2005). Caracteriza-se por um
ecossistema de Mata Atlântica de baixada com forte interferência antrópica, cuja vegetação
Tabela 2.1. Dados climáticos do período estudado na Reserva Biológica de Poço das Antas (obtidos pela Associação Mico-Leão Dourado). Temp méd: temperatura média; temp min: temperatura mínima média; temp max: temperatura máxima média; temp MIN: temperatura mínima absoluta; temp MAX: temperatura máxima absoluta; prec: precipitação mensal (mm); prec MAX: precipitação diária máxima; dias de chuva: dias do mês com precipitação.
temp med temp min temp max temp MIN temp MAX prec prec MAX dias chuvaout/04 24,7 18,5 30,9 12 37 178,6 52,2 13nov/04 26,0 19,1 32,8 16 39 338,1 210,8 11dez/04 25,5 20,8 32,1 18 39 383,3 73,3 16jan/05 27,1 21,2 33,0 18 40 343,3 66,3 14fev/05 26,4 20,1 32,6 17 37 443,7 68,8 16mar/05 27,9 22,3 33,6 20 40 299,5 68,8 14abr/05 27,4 21,5 33,3 18 39 306,9 142,9 10mai/05 24,5 19,0 30,0 16 36 59,8 29,0 7jun/05 22,6 16,2 29,0 14 34 93,2 55,1 6jul/05 22,3 16,6 28,0 13 39 170,2 91,4 8
ago/05 23,8 16,4 31,3 12 39 13,7 6,8 3set/05 23,5 18,2 28,7 13 38 184,4 42,1 15

xxiv
compreende: Mata Atlântica em estágios inicial, médio e avançado, com cerca de 10% de
campos antrópicos e 26% de formação pioneira com influência fluvial, num total de 36%
de áreas degradadas (IBAMA, 2004). Acredita-se que essa fragmentação tenha se iniciado
na metade do século XX, determinada por desmatamento, construção da barragem de
Juturnaíba, obras de drenagem de baixadas e retificação de afluentes da lagoa de Juturnaíba
e dos rios São João e Aldeia Velha, e abertura de estradas (SEMADS, 2002).
O rio São João é o principal curso d' água da região e seus afluentes, os rios Capivari,
Bacaxá, Aldeia Velha, Preto e o Iguapé percorrem boa parte da reserva, inundando as terras
baixas dos seus vales. A lagoa de Juturnaíba (Figura 2.2) inunda uma boa parte da área,
recebendo as águas de alguns rios e descarregando no rio São João. O quadro hidrológico
sofreu profundas mudanças nos últimos anos com a construção da barragem de Juturnaíba
(Programa Mata Atlântica, 2005). A barragem alagou terras da unidade a montante e
promoveu uma radical drenagem a jusante, ocasionando a modificação de parte do
ecossistema, que era de matas inundáveis (área de cerca de 900 ha), e onde hoje ocorrem os
grandes incêndios da Reserva, aumentando a pressão antrópica sobre ela (IBAMA, 2004).
Figura 2.2.
Foto da região da lagoa (seta amarela) e barragem (seta verde)
de Juturnaíba, indicando a localização da área de estudo (seta vermelha).

xxv
A Reserva Biológica de Poços das Antas é a única unidade de conservação que
possui uma população abundante de Parides ascanius. Até 1984 havia nesta reserva pelo
menos 1000 hectares do habitat ideal dessa espécie, em várias manchas conectadas (Otero
& Brown, 1986). Com a construção da barragem de Juturnaíba, em 1984, houve grande
modificação de boa parte das áreas de distribuição dessa borboleta na Reserva, sem que
tenha havido nenhum estudo ecológico posterior sobre o assunto.
Por ocasião do plano de manejo, realizado na Rebio de Poço das Antas em 2003,
foram observados alguns indivíduos voando próximo a alagados decorrentes da construção
da barragem. Estas novas observações durante o ano de 2003 confirmaram a ocorrência de
indivíduos de P. ascanius na ReBio de Poço das Antas o que indicou que essa Reserva
constitui uma área de grande importância na preservação desta espécie. Por outro lado, é
preocupante o fato dessa população ter sido observada em uma área bem restrita, em
poucos hectares ao redor de áreas pantanosas do rio São João e da barragem de Juturnaíba.
Como a população de P. ascanius ocorre estritamente em uma área da ReBio que sofreu
grande transformação devido à construção da barragem, se faz necessário um
monitoramento de seus parâmetros populacionais. A investigação da abundância e
dinâmica dessa população nos possibilitou acessar o seu real estado populacional,
contribuindo dessa forma para ações de manejo dessa espécie.

xxvi

xxvii
3.1 INTRODUÇÃO
Em um trabalho clássico sobre a distribuição e abundância dos animais,
Andrewartha & Birch (1954) resumiram as questões básicas da ecologia de populações
animais: “Por quê um animal ocupa determinada área e não outras no planeta?”; “Por quê
este é abundante em algumas partes da sua distribuição e raro em outras?”; “Por quê é
algumas vezes abundante e outras raro?”. Tais questões, além de serem centrais para a
teoria ecológica e evolutiva, são de extrema importância em um mundo super-populoso
com recursos em declínio.
Geralmente é difícil determinar os limites da distribuição e o número de organismos
em uma dada localidade. Dificuldades técnicas e logísticas aparecem na investigação de
questões sobre estrutura e dinâmica de populações individuais, variando de acordo com o
grupo taxonômico. Organismos ideais para estudos populacionais devem ser bem
conhecidos taxonomicamente, fáceis de serem reconhecidos e marcados no campo, de
tranqüila manipulação em laboratório e com gerações curtas temporalmente. As borboletas
são organismos ideais para se investigar essas questões, sendo a estrutura e dinâmica de
suas populações mais conhecida do que de qualquer outro grupo de invertebrado,
possivelmente mais do que de qualquer grupo animal (Ehrlich, 1984), com a possível
exceção de aves.
Estudos populacionais de borboletas, utilizando métodos de marcação e recaptura,
são considerados um passo importante para a compreensão de sua ecologia e
comportamento, normalmente revelando muitas informações em poucos meses de trabalho
(Ehrlich, 1984). Espécies de Papilionidae são consideradas fáceis de serem estudadas; são

xxviii
fortemente atraídas por flores, de grande tamanho corporal, e fáceis de serem marcadas sem
causar trauma (Freitas & Ramos 2001).
Entretanto, bons estudos de marcação e recaptura de papilionídeos são escassos na
América do Norte e praticamente inexistentes com espécies neotropicais. Talvez, a raridade
da maioria das espécies, combinada com períodos de vôo relativamente curtos e a
dificuldade de estar periodicamente em campo para obter bons números de indivíduos
recapturados tenham frustrado muitos pesquisadores (Tyler et al., 1994).
A família Papilionidae apresenta cerca de 600 espécies distribuídas em 3
subfamílias: Baroniinae, Parnassiinae e Papilioninae. A subfamília Papilioninae contém
aproximadamente 550 espécies, divididas em 3 tribos (Leptocirciini, Troidini e Papilionini)
e 17 gêneros (Ackerey et al., 2001). Essa família, apesar de cosmopolita, concentra-se nos
sub-trópicos, sendo 143 espécies registradas para o continente americano (Feeny 1991;
Tyler et al., 1994).
Papilionidae são borboletas com vôo ágil e poderoso, mas nem sempre rápido. São
abundantes em ambientes úmidos, com grande abundância de árvores nativas e variedade
de microhábitats. Os machos possuem androcônias com cheiro bem forte e característico,
usados para acalmar as fêmeas durante os cortejamentos aéreos vigorosos e prolongados.
As fêmeas de muitas espécies são raramente vistas, até desconhecidas, tendo hábitos que
evitam encontros com coletores, como vôo no interior de matas densas, ou pendurando-se
como uma pipa a 50 m acima do chão, à procura de brotos novos em cima de plantas
hospedeiras. Esta é a família mais primitiva das borboletas “verdadeiras” (Papilionoidea), e
inclui muitas espécies antigas, relictuais, de distribuição muito restrita, à beira da extinção
devido à destruição, pelo homem, dos seus poucos hábitats remanescentes (Brown Jr.,
1992).

xxix
Durante o tempo que os Papilionidae têm estado nas Américas, eles se expandiram
para ocupar praticamente todos os hábitats possíveis, das gélidas tundras, e tórridos
desertos até rochedos, vegetações arbustivas, todos os tipos de florestas, encostas e topos de
montanhas, matas ciliares, planícies de baixada, até parques urbanos e jardins. A maioria
das espécies é típica de florestas tropicais, sendo apenas três características de vegetações
costeiras, Parides ascanius, Parides zacynthus e Heraclides aristodemus (Tyler et al.,
1994).
A maioria dos indivíduos de papilionídeos necessita de muito espaço. Entretanto,
uma população inteira pode estar comprimida em um espaço pequeno, como um parque
urbano, se os outros recursos forem sempre abundantes. Assim, o espaço é às vezes um
recurso limitante ou um eixo central do nicho determinando o tamanho das populações
quando o recurso alimentar é limitado, especialmente se machos forem territorialistas
(Tyler et al., 1994).
A maioria das dimensões do nicho se constitui de outros recursos e pressões
fundamentais do ambiente físico e biológico. O tempo é essencial para os adultos de
papilionídeos, que têm apenas alguns dias para amadurecer, preparar um pacote de
espermatóforos de alta qualidade (machos), acasalar, e encontrar a planta ideal para
oviposição (fêmeas). O uso eficiente do recurso “tempo” é crítico para o sucesso
reprodutivo do adulto. Recursos renováveis, incluindo luminosidade, temperatura e
umidade, podem limitar a atividade de adultos, e assim regular ou restringir o uso de todos
os outros recursos. Pouquíssimos estudos foram feitos sobre tolerâncias e limites destes
recursos para as espécies de Papilionidae (adultos ou jovens), variação entre indivíduos, ou
sobre o efeito desses fatores sobre o comportamento e fecundidade (Tyler et al., 1994).

xxx
A tribo Troidini é predominantemente tropical, com a maioria das espécies
concentrada nas florestas de planícies da América Central e do Sul e da região Indo
Australiana (Weintraub, 1995). Seus representantes oferecem um dos melhores exemplos
de oligofagia em borboletas, sendo exclusivos de uma pequena variedade de
Aristolochiaceae. Outros Papilioninae usam uma maior variedade de plantas hospedeiras,
como Annonaceae, Apiaceae, Canellaceae, Hernandiaceae, Lauraceae, Rutaceae,
Winteraceae, entre outras (Miller 1987; Weintraub, 1995).
Essa tribo inclui 130 espécies divididas em 12 gêneros, três dos quais ocorrem na
região neotropical: Battus (11 espécies), Euryades (2 espécies), e Parides (34 espécies)
(Tyler et al., 1994). O gênero Parides é exclusivamente neotropical, e inclui várias espécies
nas listas oficiais de espécies ameaçadas de extinção (IUCN, 2003; MMA, 2003). Os
Troidini parecem ser um grupo monofilético, com o gênero Battus sendo monofilético e
irmão das demais espécies da tribo, e Euryades sendo aparentado com o gênero
monofilético Parides (Silva-Brandão et al., 2005).
O comportamento sexual das espécies de Parides é bem complexo, com ambos os
sexos usando substâncias químicas pra comunicar virilidade ou receptividade, havendo uma
perseguição aérea seguida de uma dança conjunta. Os machos de Parides são mestres na
persuasão química e muitos fazem vôos acrobáticos ao redor de fêmeas estacionárias,
abrindo as asas posteriores e depositando a penugem branca cheia de ferormônios nas
antenas das fêmeas, a cada movimento de ida e vinda (Tyler et al., 1994).
Os machos de Parides, assim como os de outros papilionídeos, são comumente
vistos defendendo recursos florais contra outras borboletas. Esse comportamento não
parece ser sempre e exclusivamente para o propósito do direito às fêmeas. Ocasionalmente,

xxxi
fêmeas foram observadas perseguindo e expulsando outras da área da planta hospedeira
(Tyler et al., 1994).
Parides ascanius, a borboleta da praia, foi a primeira espécie de inseto brasileiro a
entrar na lista de espécies ameaçadas de extinção no Brasil. Endêmica do Rio de Janeiro,
ela ocorre em pequenas manchas de vegetação brejosa ou pantanosa entre Atafona (São
João da Barra) e Itaguaí (Rio de Janeiro), e tem como única planta hospedeira Aristolochia
macroura (Aristolochiaceae), conhecida como jarrinha (Otero, 1984; Otero & Brown Jr.,
1986; Otero & Marigo, 1990). Seu adulto, nectívoro, tem como flor favorita Lantana
camara (Verbenaceae) (Otero & Marigo, 1990). Sua lagarta armazena substâncias tóxicas
das folhas ou galhos de A. macroura, passando para os adultos, tornando-se impalatável
para alguns predadores (Otero, 1984; Otero & Brown Jr. 1986; Brown Jr., 1992). O adulto
voa praticamente o ano todo, podendo ter diapausa, na fase de crisálida, durante o inverno
(Otero, 1984). O hábito monófago da lagarta torna esta espécie ainda mais suscetível à
extinção, uma vez que depende da aptidão de A. macroura para manter populações viáveis.
A distribuição de Parides ascanius provavelmente sempre foi restrita e pontual.
Ocorre em partes alagadas da planície costeira do estado, especialmente com vegetação
arbustiva ao longo de vales de rios de baixada, mas pode ser encontrada até 30km da costa.
Seu habitat ocorre em manchas descontínuas dependendo de combinações estreitamente
definidas de clima, solo, drenagem, estrutura luminosa da vegetação e presença de certas
espécies de plantas. P. ascanius tem distribuição agregada no espaço; quando uma
população está presente, em certas épocas do ano podem ser avistados até 50 indivíduos em
uma manhã. Portanto, é improvável que populações importantes dessa espécie de borboleta,
bem aparente no tamanho e coloração, ainda sejam desconhecidas em regiões acessíveis e

xxxii
que tenham sido estudadas ou visitadas por especialistas em borboletas (Otero & Brown Jr.
1986).
A inacessibilidade e inospitalidade de seu habitat mantiveram muitas populações a
salvo da expansão urbana até recentemente. Com o crescimento populacional do estado do
Rio de Janeiro, os alagados costeiros e de baixada estão sendo drenados e transformados ao
longo de toda a distribuição de P. ascanius. Atualmente, populações de P. ascanius se
restringem a poucas regiões em áreas ou habitats específicos e sob forte impacto antrópico,
devendo existir, segundo Tyler et al. (2004), aproximadamente 20 colônias muito instáveis.
Existe um grande risco de restarem somente algumas populações isoladas em áreas
preservadas, que poderão enfrentar problemas genéticos devido a cruzamentos
consangüíneos (Campos Neto & Otero, 2001), sendo a Reserva Biológica de Poços das
Antas a única unidade de conservação considerada capaz de manter uma grande população
(Otero & Brown, 1986).
Desde a publicação do extenso trabalho de Otero & Brown (1986), existe um
domínio completo do ciclo de vida e comportamento alimentar e reprodutivo de P.
ascanius, na natureza e em cativeiro. Porém, nos últimos 20 anos, pouca informação nova
foi publicada sobre essa espécie, aparecendo esta somente marginalmente em alguns
artigos. Desde então, nenhuma população de P. ascanius foi estudada mais detalhadamente
quanto à sua estrutura, sendo ausentes informações ecológicas importantes para a
conservação dessa espécie.
A Reserva Biológica de Poços das Antas é a única unidade de conservação que
possui uma população abundante de Parides ascanius. Até 1984 havia nesta Reserva pelo
menos 1000 hectares do habitat ideal dessa espécie, em várias manchas conectadas (Otero
& Brown, 1986). Com a construção de barragem de Juturnaíba, em 1984, houve grande

xxxiii
modificação de boa parte das áreas de distribuição dessa borboleta na reserva, sem que
tenha havido nenhum estudo ecológico posterior sobre o assunto.
Portanto, nosso objetivo foi estimar o tamanho de uma população de P. ascanius e
sua dinâmica ao longo do tempo, com o intuito de confirmar a viabilidade dessa espécie na
ReBio Poço das Antas. Foram considerados também aspectos como a estrutura etária e
proporção sexual da população, além da longevidade e dano alar dos espécimens, a
ocupação do habitat e a movimentação dos indivíduos neste, com a finalidade de acessar
informações mais precisas que facilitem futuras ações de manejo dessa espécie.

xxxiv
3.2 MATERIAIS E MÉTODOS
O estudo foi realizado na Reserva Biológica de Poço das Antas, no município de
Silva Jardim, no estado do Rio de Janeiro, em uma área próxima à barragem de Juturnaíba
(Figura 3.1).
Figura 3.1. Recorte de imagem de satélite (IKONOS) representando a área de estudo e seu entorno. Em vermelho:
estrada principal da Reserva Biológica de Poço das Antas. Em amarelo: trilha. Em verde claro: área de baixada periodicamente inundada. Em verde escuro: mata. Em preto: alagado decorrente da Represa de Juturnaíba.

xxxv
Partindo de observações preliminares, foi demarcada a área de estudo, que consistiu
de aproximadamente um quilômetro na estrada da ReBio. Esse transecto (Figura 3.2) foi
marcado a cada 50 metros para facilitar a localização dos indivíduos, e foi feito na própria
estrada que atravessa a ReBio, o que, de certa maneira, facilitava a observação e captura
dos espécimens. Consistia de uma região de “morrote” e uma de “baixada” (Figura 3.3).
Figura 3.2. Recorte de imagem de satélite (IKONOS) representando a área de estudo. Em roxo: transecto em que foram realizadas as capturas. Seta azul: “morrote”. Seta branca: “baixada”. A marcação do transecto iniciou-se na estrada em vermelho (0m) e terminou na estrada em amarelo (1000m).

xxxvi
O “morrote” (Figura 3.4) é caracterizado por apresentar floresta ao longo das
margens da estrada, proporcionando diferentes áreas sombreadas ao longo do dia, e uma
área de “morrote” mais aberta, com alta luminosidade durante boa parte do dia, e cerca de
20 metros acima da baixada. Essa região tem uma fronteira abrupta com a baixada, com
poucos metros de faixa de transição, determinada basicamente pelo relevo.
Figura 3.3. Fotografia (R.F. Monteiro) da área de estudo, localizada a cerca de 1 km do rio São João, na parte sul da Reserva Biológica de Poço das Antas, mostrando bem a separação das regiões de “morrote” (seta azul) e “baixada” (seta branca).

xxxvii
A “baixada” (Figura 3.5) é caracterizada por uma região periodicamente alagada
dos dois lados da trilha, recebendo grande incidência solar ao longo da maior parte do dia.
Figura 3.4. Fotografia da região do “morrote”, caracterizada por uma área de grande sombreamento e uma faixa com intensa luminosidade.
Figura 3.5. Fotografia (R.F. Monteiro) da região da “baixada”, caracterizada por grande luminosidade e um terreno permanentemente inundado.

xxxviii
Durante o período de outubro de 2004 a setembro de 2005 foram realizadas 18
amostragens, com uma periodicidade mínima de uma vez por mês. Nos meses de
novembro, março, julho e setembro foram feitas amostras em dois dias consecutivos. Em
outubro e fevereiro foram feitas também duas amostras, porém separadas por intervalos de
cerca de 20 dias.
Em cada sessão, durante cinco horas, divididas em dois turnos (manhã e tarde),
foram capturados todos os indivíduos de P. ascanius (Figura 3.6) possíveis. Em cada um
desses turnos, dois pesquisadores equipados com rede entomológica percorriam lentamente
o transecto nas duas direções (ida e volta), procurando ativamente por indivíduos de P.
ascanius.
Como em 6 dos 12 meses estudados (outubro, novembro, fevereiro, março, julho e
setembro) foram realizadas sessões de marcação e recaptura durante dois dias, consecutivos
ou não, para uma melhor visualização da variação temporal do tamanho populacional e
comparação entre as amostras foram tiradas as médias correspondentes aos dois dias de
captura. Esse procedimento foi realizado para as análises de abundância, razão sexual e
Figura 3.6. Adulto de Parides ascanius.

xxxix
estrutura etária da população. Para as análises de recaptura, esse procedimento também foi
adotado, com exceção do mês de outubro, no qual foram feitas duas amostras com um
espaçamento de 20 dias. Em fevereiro também foram feitas amostras em dois dias distantes,
mas em um deles o mau tempo impediu que a amostragem fosse completa e, portanto, para
efeitos de recaptura, os dados desse mês foram somados.
Devido ao fato de mortalidade e emigração só poderem ser separadas em
populações abertas pelo método de Zhang & Brown (1989), que não pode ser utilizado
nesta dissertação, o termo ‘residência’ foi usado ao invés de ‘sobrevivência’, seguindo Watt
et al. (1977) e Tabashnik (1980).
Para estimar o tamanho populacional foi utilizado o estimador de Lincoln-Peterson
(1930) modificado por Bailey (1952). Esse índice é provavelmente o mais popular dos
modelos de abundância absoluta, como também o mais simples, sendo necessárias apenas
duas amostragens para obtê-lo: a marcação e a recaptura (Gall, 1985). É calculado da
seguinte maneira:
N = n1 (n2 + 1)/(m + 1)
sendo,
N = tamanho populacional total
n1 = número de animais capturados na primeira sessão de captura
n2 = número de indivíduos capturados na segunda sessão
m = número de indivíduos recapturados na segunda sessão
Esse estimador requer que sejam válidas algumas premissas, assim como todos os

xl
modelos de abundância absoluta, tais como: amostragens em intervalos separados, curtos
em relação ao tempo total de estudo; animais não têm marcas perdidas; animais marcados
podem ser distinguidos dos não marcados; ausência de alteração comportamental após a
marcação; “diluição” dos indivíduos marcados entre os não marcados; animais marcados e
não marcados com mesma probabilidade de captura; amostragem em proporções naturais
dos sexos, classes etárias, etc. Além dessas, o índice de Lincoln-Peterson/Bailey tem como
premissa adicional que a população não sofra nenhuma perda (morte ou emigração) ou
ganho (nascimento ou imigração) durante o intervalo entre as sessões de captura e recaptura
(Gall, 1985).
Os indivíduos capturados eram marcados com uma numeração na parte ventral das
quatro asas com uma caneta de marcação permanente (Figura 3.7), Sharpie-Stanford, não
tóxica, e depois soltos. A cor da caneta utilizada foi vermelha para que a marcação não
destoasse do padrão de coloração da espécie.
A marcação foi feita nas quatro asas para evitar que fosse perdida, uma vez que
partes das asas podem ser danificadas ou mesmo extirpadas, e garantir sua correta leitura,
Figura 3.7. Marcação nas asas de P. ascanius.

xli
visto que o cuidado para não danificar as borboletas muitas vezes impediu uma adequada
grafia da numeração.
O registro de indivíduos capturados, com os respectivos números, foi feito no
próprio local, em planilha própria. Além da marcação, eram anotados o sexo do indivíduo,
seu dano alar, faixa etária, horário e localização da captura. O dano alar e a faixa etária das
borboletas eram determinados em três classes: dano alar (0: nenhum dano, 1: pouco dano,
2: muito dano); faixa etária (0: novo, 1: intermediário, 2: velho). Em relação ao dano alar,
observavam-se os cortes (pedaços danificados) e rupturas (pedaços ausentes) nas asas. Se
cortes e rupturas eram inexistentes, os indivíduos eram classificados com nenhum dano
alar. Caso, houvesse de um a três cortes, ou um ou dois cortes e uma ruptura, era atribuída a
categoria pouco dano. Nas situações em que apareciam mais de três cortes ou 2 ou mais
rupturas, os espécimens eram classificados com muito dano. A faixa etária foi estimada a
partir do desgaste alar, sendo observada unicamente a situação das escamas das asas. Os
indivíduos considerados novos não possuíam nenhum desgaste alar, provavelmente sendo
incluídos nessa categoria apenas borboletas com um ou dois dias de vida. Os espécimens
classificados como intermediários apresentavam algum desgaste alar, mas o padrão de
cores das asas ainda era nítido. Já os indivíduos categorizados como velhos tinham a
coloração das asas já descaracterizada, com o preto e as manchas rosas já quase
imperceptíveis.
Não foram tomadas medidas do comprimento das asas para evitar manipulação dos
indivíduos, uma vez que se trata de uma espécie ameaçada de extinção e uma manipulação
excessiva pode acarretar estresse de indivíduos, com conseqüentes mudanças
comportamentais que poderiam acarretar em alterações biológicas, inclusive morte, e
metodológicas.

xlii
Idealmente, a captura e marcação devem afetar tão pouco a borboleta que depois de
solta ela volte a fazer o que estava fazendo. O “trauma de marcação” pode promover uma
dispersão para fora da área e com isso reduzir significativamente as taxas de recaptura
posteriores de papilionídeos (Lederhouse, 1982; Ehrlich 1984).
Durante as capturas de P. ascanius eram também observadas e registradas outras
espécies de borboletas na área (Nymphalidae, Papilionidae e Pieridae).
Além da amostragem de rotina, foram realizadas diversas expedições para a
localização de outras manchas ou populações, em diversos locais da ReBio, para permitir
um melhor delineamento da situação da espécie na ReBio e futuros trabalhos sobre
possíveis estruturas metapopulacionais e comparações entre manchas.
Para as análises estatísticas foi utilizado o programa GRAPHPAD INSTAT (1997),
versão 3.0. A normalidade dos dados foi testada por Kolmogorov-Smirnov. Foram feitas
regressões lineares entre a abundância de indivíduos e os fatores climáticos (temperatura
média, temperatura mínima média, temperatura máxima média, temperatura mínima
absoluta, temperatura máxima absoluta, precipitação total, precipitação máxima diária e
dias de chuva) do mês, da semana anterior e do dia da captura. Para testar a diferença entre
o número de machos e fêmeas foi usado o teste t não pareado com correção de Welch. A
distribuição dos indivíduos nas diferentes faixas etárias foi avaliada por repetidas ANOVAs
seguidas do teste de Tukey. A diferença entre a distribuição etária e o dano alar de machos
e fêmeas foi testada com o teste t não pareado com correção de Welch.

xliii
3.3 Resultados
Foram capturados e marcados 307 indivíduos de P. ascanius, 230 machos e 77
fêmeas (3:1). Tanto o número de indivíduos como as razões sexuais variaram muito ao
longo do ano.
3.3.1 Variação temporal e tamanho populacional
A população estudada se reproduziu o ano inteiro, porém o número de indivíduos
capturados por dia (NICD, incluindo marcados e recapturados) variou de 1 a 53. O NICD
de machos variou de 0 a 38 (x = 15,06 ± 12,85). O NICD de fêmeas variou de 1 a 16 (x
= 4,44 ± 4,03). O mês que registrou a menor abundância diária foi fevereiro, enquanto em
setembro foi detectado o maior número de indivíduos em um dia. Quando foi levada em
consideração a média dos meses, a abundância de P. ascanius na ReBio passou a variar de
5 a 39, sendo janeiro o mês com menor número de indivíduos e novembro o mês de maior
abundância.
Verificou-se um pico populacional na primavera, com uma posterior queda no verão
e a manutenção de níveis intermediários no outono e inverno (Figura 3.8). Apesar de
pequenas variações, tanto nas abundâncias mensais como nos meses de maior e menor
abundância, o padrão geral foi mantido. A primavera parece ser realmente a época de pico
populacional dessa espécie, sendo setembro, outubro e novembro os meses de maior
abundância. Em dezembro, já podemos começar a perceber um declínio populacional, que
chega ao seu limite no verão, durante os meses de janeiro, fevereiro e março.

xliv
A maior parte das regressões lineares entre a abundância de indivíduos e os fatores
climáticos foi negativa, sendo as únicas exceções com a temperatura máxima média e
absoluta de uma semana antes do dia de captura, e com a precipitação máxima diária do
mês. Entretanto, nenhuma dessas relações foi significativa (Tabela 3.1).
Foram recapturados 52 indivíduos de P. ascanius, 48 machos e 4 fêmeas, o que
corresponde a aproximadamente 17% das borboletas marcadas. O índice de recaptura dos
machos (21%) foi expressivamente maior do que das fêmeas (5%). Dos indivíduos
recapturados, 42 o foram uma única vez, enquanto 10 foram duas vezes recapturados.
Em metade das sessões de captura e recaptura não houve nenhuma recaptura,
enquanto nas demais essa taxa variou de 3% a 17%. Quando reunimos os dados dos meses
Figura 3.8. Média do número de indivíduos de Parides ascaniuscapturados por dia (NICD) em cada mês estudado.
0
510
1520
2530
3540
45
out nov dez jan fev mar abr mai jun jul ago set
meses de captura
NIC
D
Tabela 3.1. Regressões entre abundância de indivíduos em cada sessão de marcação e recaptura e os fatores climáticos: valores de R2
considerando os dados de clima do dia, da semana anterior e do mês da captura.
abundância temp med temp min temp max temp MIN temp MAX prec prec MAX dias chuvadia 0,03 0,13 0 0,15 0 0,18 0,18
semana 0,01 0,12 0,02 0,15 0 0,12 0,08 0,07mês 0,10 0,07 0,08 0,18 0,03 0,01 0,12 0,02

xlv
em que houve coletas em dois dias, os meses de novembro, fevereiro, julho e setembro, que
não apresentavam recaptura alguma em um dos dias de amostragem, passam a contar com
índices de recaptura razoáveis, com a exceção de setembro, que apresentou apenas 3% dos
indivíduos recapturados (Tabela 3.2).
As estimativas de tamanhos populacionais variaram de 8 a 475 indivíduos, tendo
sido janeiro o mês de menor abundância e outubro o de maior, porém devido aos baixos
índices de recaptura e grandes intervalos de confiança esses valores devem ser considerados
com cautela.
Em abril, o número de indivíduos capturados foi maior do que a estimativa
populacional para o mês. Nos demais meses, a proporção das estimativas que foram
representadas por indivíduos efetivamente capturados variou de 9 a 100%, o que indica
uma diferença grande na precisão das estimativas ao longo dos meses (Tabela 3.3). Em
alguns meses, as estimativas populacionais tiveram valores próximos do dobro do número
de machos, sendo este último sempre igual ou menor do que os valores estimados, sendo
abril a única exceção a esse padrão. As maiores diferenças entre os valores estimados e o
dobro do número de machos capturados ocorreram em outubro, março, junho e setembro,
justamente os meses que tiveram as menores proporções das estimativas representadas por
indivíduos capturados (NICD/LP).
Tabela 3.2. Número de indivíduos por dia de captura (NICD), % de indivíduos recapturados (recaptura), estimativa de tamanho populacional por Lincoln-Petersen (LP) e intervalo de confiança da estimativa (IC), em cada um dos meses estudados.
out out nov dez jan fev mar abr mai jun jul ago setNICD 22 43 68 23 5 13 13 12 19 16 27 10 65
recaptura 0 0 0,15 0,13 0 0,15 0 0,17 0,16 0,06 0,11 0 0,03LP 76 475 142 46 8 21 49 9 33 39 46 10 265IC 129 805 792 153 12 59 82 25 106 95 164 0 805

xlvi
3.3.2 Razão sexual
A razão sexual da população em geral foi desviada para machos (3:1), sendo essa
diferença significativa (Welch’s t = 2,44 g.l. 12, P=0,03), porém houve uma grande
variação entre as amostras. A razão sexual variou de 3 fêmeas para cada macho até 22
machos para cada fêmea. Na maior parte do ano os machos foram mais abundantes, sendo,
muitas vezes, essa proporção sexual bastante desviada (Figura 3.9).
Dezembro foi o mês com razão sexual mais desviada, seguido por abril, novembro e
outubro. No mês de agosto foi encontrada uma proporção sexual de 1:1, e valores próximos
Tabela 3.3. Dobro do número de machos (2M), estimativa populacional baseada em Lincoln-Peterson (LP) e LP/NICD, em cada um dos meses estudados.
out out nov dez jan fev mar abr mai jun jul ago set2M 36 76 118 44 8 18 20 22 28 8 32 10 96LP 76 475 142 46 8 21 49 9 33 39 46 10 265
LP/NICD 0,29 0,09 0,48 0,51 0,63 0,61 0,27 1,38 0,58 0,42 0,59 1,00 0,25
0
5
10
15
20
25
out nov dez jan fev mar abr mai jun jul ago set
meses de captura
mac
hos/
fêm
eas
Figura 3.9. Razão entre número de machos para cada fêmea em cada um dos meses estudados. Linha tracejada em vermelho corresponde a razão 1:1.

xlvii
a este em fevereiro e julho. Em junho, a razão sexual foi desviada para fêmeas, tendo sido
capturadas aproximadamente 3 fêmeas para cada macho.
3.3.3 Distribuição etária e dano alar
Aproximadamente 50% dos indivíduos capturados foram classificados na categoria
etária intermediária, 35% foram categorizados como novos e cerca de 15% como velhos
(Figura 3.10), sendo esta diferença significativa. Machos e fêmeas não diferiram
significativamente em relação à distribuição etária (Welch’s t = 1,84 g.l. 2, P = 0,2). O
padrão de distribuição da faixa etária foi determinado pelo padrão dos machos, já que estes
representaram aproximadamente 75% dos espécimens capturados. Cerca de 50% das
fêmeas também foram incluídas na categoria intermediária, porém foi constatada uma
maior proporção de indivíduos velhos e uma menor proporção de indivíduos novos em
relação aos machos.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
total machos fêmeas
velho
intermediário
novo
Figura 3.10. Percentual de espécimens de cada faixa etária entre todos os indivíduos (F = 25,85 g.l. 35, P 0,0001), entre machos e entre fêmeas.

xlviii
A distribuição etária da população não variou muito ao longo do ano, tendo o
padrão diferido mais nos meses em que a abundância foi menor (Figura 3.11). A proporção
de indivíduos novos foi em média 32%, variando de 20% em janeiro a 50% em agosto e
mantendo-se próxima a 30% nos demais meses. Os indivíduos com idade intermediária
representaram em média 53% dos indivíduos capturados, constituindo 40% das borboletas
de dezembro e agosto e 80% em janeiro. A média de indivíduos velhos foi de 15%, mas
esse valor variou de 0% em janeiro e março a 25% em outubro, abril e junho.
Considerando a primeira captura de cada indivíduo, um pouco mais da metade dos
espécimens não apresentou nenhum dano alar, enquanto que aproximadamente 15% dos
indivíduos apresentaram muito dano nas asas (Figura 3.12). Comparando machos e fêmeas,
pode-se perceber algumas diferenças com relação à incidência de danos nas asas, porém
não significativas. O padrão dos machos foi bastante semelhante ao padrão geral, o qual
deve ter sido regido justamente pelos machos, que representaram um número três vezes
0%
20%
40%
60%
80%
100%
ou
t
no
v
de
z
jan
fev
ma
r
ab
r
ma
i
jun jul
ag
o
set
velho
médio
novo
Figura 3.11. Percentual de indivíduos de cada faixa etária em cada um dos meses estudados.

xlix
maior do que as fêmeas. Entre as fêmeas, houve uma maior proporção de indivíduos com
muito dano.
Entre os indivíduos novos foi detectado nenhum ou pouco dano alar. As borboletas
incluídas na faixa etária intermediária apresentaram nenhum, pouco ou muito dano nas
asas. Já os espécimens categorizados como velhos mostraram pouco ou muito dano alar. As
combinações encontradas foram as mesmas em ambos os sexos, e desprezível a diferença
da regressão entre a faixa etária e o dano alar entre os sexos (R2 = 0,49 e F = 263,5 para
machos; R2 = 0, 42 e F = 59,21 para fêmeas), sendo ambas significativas (P 0,0001).
3.3.4 Ocupação do habitat e movimentação
As capturas não ocorreram de forma homogênea ao longo do transecto, existindo
claramente trechos de maior probabilidade de captura. Um trecho de 100 metros (500-
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
total machos fêmeas
muito
pouco
nenhum
Figura 3.12. Percentual de espécimens com nenhum, pouco ou muito dano alar entre todos os indivíduos (total), entre machos e entre fêmeas.

l
600m) não teve nenhum indivíduo capturado, enquanto que três trechos (0-100m, 100-
200m e 800-900m) foram responsáveis por cerca de 73% das capturas (Figura 3.13).
As distribuições espaciais de machos e fêmeas foram um pouco diferentes,
principalmente no que se refere aos três trechos mais utilizados por P. ascanius. Ambos os
sexos tiveram grande parte dos seus indivíduos capturados nesses trechos (79% fêmeas e
76% dos machos), porém, as fêmeas foram proporcionalmente mais capturadas no trecho
de 0 a 100 metros do transecto, enquanto que os machos foram encontrados mais
freqüentemente no trecho entre 800 e 900 metros (Figura 3.14).
0 20 40 60 80 100 120
0-100
101-200
201-300
301-400
401-500
501-600
601-700
701-800
801-900
901-1000
trec
hos
do t
rans
ecto
número de indivíduos
Figura 3.13. Número de indivíduos capturado em cada um dos trechos de 100 metros do transecto.

li
Foram obtidos padrões diferentes no uso do espaço dentro de cada sexo,
dependendo da faixa etária. As fêmeas novas se distribuíram semelhantemente nos três
trechos mais ocupados do transecto, e fêmeas intermediárias e velhas foram capturadas
majoritariamente no primeiro trecho. Os machos novos foram mais comuns no trecho
correspondente ao trecho 0-100 metros do transecto. Os machos classificados como
intermediários ocuparam semelhantemente as áreas de 0-100 e 800-900 metros. Já os
machos velhos foram raros nos primeiros duzentos metros do transecto, tendo sido 56%
destes capturados no trecho de 800-900 metros (Figura 3.15).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
machos fêmeas
outros
800-900
100-200
0-100
Figura 3.14. Percentual de capturas de machos (N=286) e fêmeas (N=82) nos diferentes trechos do transecto.

lii
Levando em consideração as recapturas, os três trechos do transecto responderam
por cerca de 71% dos indivíduos. A proporção de espéciemens recapturados foi semelhante
nesses três trechos, sendo um pouco maior na região correspondente a 800-900 metros do
transecto (Figura 3.16).
3241
2
514
11
1831
4
5
9
521 43
23
4
6
724 3512 6 10
0
0%
20%
40%
60%
80%
100%
machos 0 machos 1 machos 2 fêmeas 0 fêmeas 1 fêmeas 2
outros
800-900
100-200
0-100
Figura 3.15. Percentual de ocupação de trechos do transecto por machos e fêmeas de diferentes faixas etárias. Faixas etárias: 0, nova; 1, intermediária; 2, velha.
23%
21%
27%
29%
0-100
100-200
800-900
outros
Figura 3.16. Percentual de recapturas nos trechos do transecto (N=62).

liii
A distância entre o ponto de captura e o ponto de recaptura dos indivíduos variou de
0 a 800 metros. Com uma maior freqüência, as distâncias entre captura e recaptura ficaram
nas faixas de 0-100 metros e 301-400 metros (Figura 3.17).
Não foi encontrada nenhuma relação entre a distância da captura e a recaptura e o
número de dias decorridos da captura. Indivíduos recapturados no mesmo dia percorreram
distâncias de até 500 metros. Um espécimen recapturado após 27 dias o foi exatamente no
mesmo local de sua captura. A maior distância percorrida registrada foi de 800 metros.
A distribuição etária dos indivíduos que foram recapturados foi desviada para
espécimens considerados intermediários, sendo apenas 13% correspondentes a indivíduos
velhos (Figura 3.18). As proporções de recapturados novos e velhos foram menores do que
as representadas por essas categorias entre o total de indivíduos capturados.
0 5 10 15 20 25 30
0-100
101-200
201-300
301-400
401-500
501-600
601-700
701-800
clas
ses
de d
istâ
ncia
número de indivíduos
Figura 3.17. Número de indivíduos recapturados nas diferentes classes de distância (N=62).

liv
Figura 3.18. Percentual de espécim ns de cada faixa etária entre os indivíduos recapturados (N=62).
Borboletas classificadas como intermediárias foram recapturadas em todas as
classes de distância, sendo maioria em quase todas elas. Os espécimens novos foram
capturados novamente até uma distância de 500 metros, sendo essa faixa etária maioria na
classe de distância de 401-500 metros. Os indivíduos velhos foram recapturados no
máximo a 350 metros de distância de seu local de captura (Figura 3.19).
26%
61%
13%
novo
intermediário
velho
0% 20% 40% 60% 80% 100%
0-100
101-200
201-300
301-400
401-500
501-600
601-700
701-800
clas
ses
de d
istâ
ncia
novo
intermediário
velho
Figura 3.19. Percentual de cada faixa etária de indivíduos recapturados em cada uma das classes de distância.

lv
Os indivíduos de todas as faixas etárias foram mais recapturados nas distâncias de
0-100 metros e menos na faixa de 101-200 metros. Entre as borboletas novas também
houve grande proporção de indivíduos recapturados a 301-400 metros do local da captura.
Aproximadamente 80% dos espécimens velhos foram capturados novamente muito
próximos do ponto de captura (0-100 metros), tendo essa proporção ficado próxima a 50%
entre indivíduos novos e intermediários (Figura 3.20).
Considerando o dano alar dos indivíduos que foram recapturados, pode-se notar
uma maior proporção de espécimens com nenhum dano nas asas, seguida de perto por
borboletas com pouco dano alar. Os indivíduos com muito dano nas asas representaram
cerca de 20% das borboletas recapturadas (Figura 3.21).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
novo intermediário velho
outras
301-400
101-200
0-100
Figura 3.20. Percentual de indivíduos de cada faixa etária recapturado em cada uma das principais classes de distância (N=62).

lvi
Espécimens classificados com nenhum ou pouco dano alar foram recapturados em
todas as classes de distância. Na classe de 0-100 metros os indivíduos com muito dano nas
asas foram tão representados como aqueles com muito dano. A uma distância de 101-200
metros do local de captura, as borboletas com pouco dano alar foram a maioria. Na classe
de 301-400 metros as proporções de indivíduos com nenhum ou pouco dano foram
semelhantes (Figura 3.22).
42%
37%
21%
nenhum
pouco
muito
Figura 3.21. Percentual de espécimens de cada classe de dano alar entre os indivíduos recapturados (N=62).
0% 20% 40% 60% 80% 100%
0-100
101-200
201-300
301-400
401-500
501-600
601-700
701-800
clas
ses
de d
istâ
ncia
nenhum
pouco
muito
Figura 3.22. Percentual de cada classe de dano alar de indivíduos recapturados em cada uma das classes de distância (N=62).
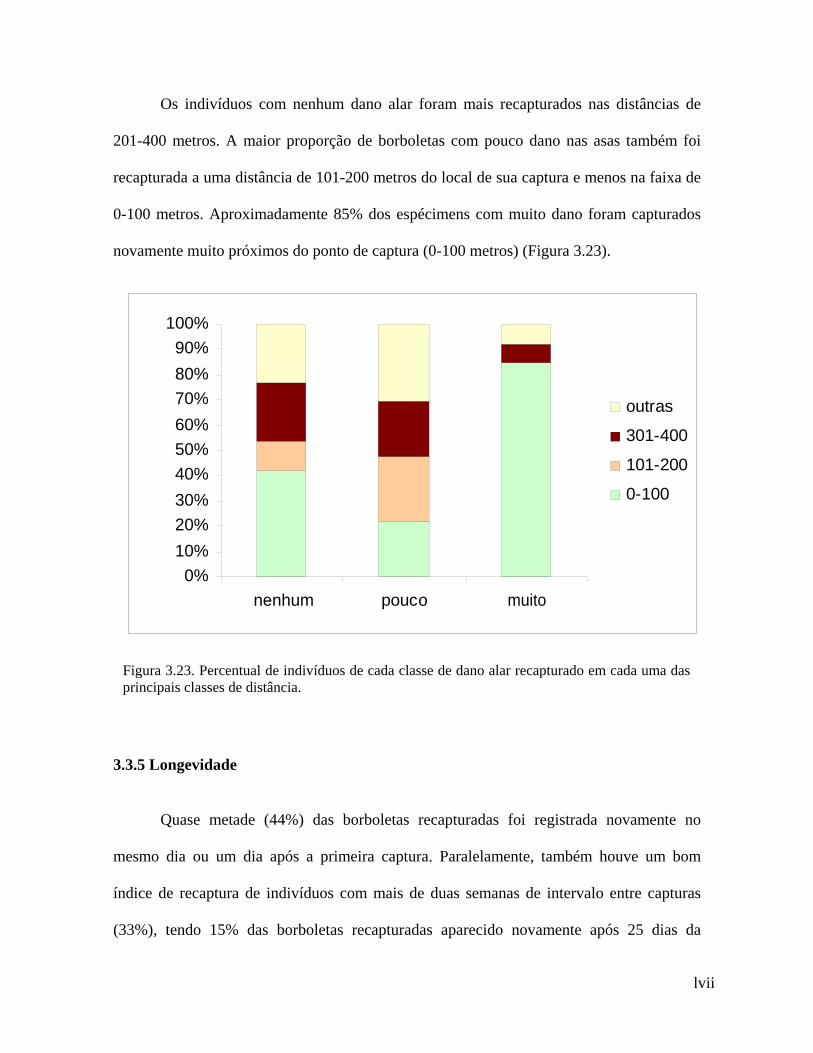
lvii
Os indivíduos com nenhum dano alar foram mais recapturados nas distâncias de
201-400 metros. A maior proporção de borboletas com pouco dano nas asas também foi
recapturada a uma distância de 101-200 metros do local de sua captura e menos na faixa de
0-100 metros. Aproximadamente 85% dos espécimens com muito dano foram capturados
novamente muito próximos do ponto de captura (0-100 metros) (Figura 3.23).
3.3.5 Longevidade
Quase metade (44%) das borboletas recapturadas foi registrada novamente no
mesmo dia ou um dia após a primeira captura. Paralelamente, também houve um bom
índice de recaptura de indivíduos com mais de duas semanas de intervalo entre capturas
(33%), tendo 15% das borboletas recapturadas aparecido novamente após 25 dias da
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
nenhum pouco muito
outras
301-400
101-200
0-100
Figura 3.23. Percentual de indivíduos de cada classe de dano alar recapturado em cada uma das principais classes de distância.

lviii
captura inicial. A fêmea mais longeva foi recapturada após 17 dias e a maior permanência
de um macho foi de 28 dias. Porém, quando o esforço de recaptura nas diferentes classes de
dias foi levado em consideração, uma maior proporção de recapturas foi encontrada na
classe de 7-12 dias, sendo as proporções nas outras classes semelhantes entre si (Figura
3.24).
3.3.6 Conservação
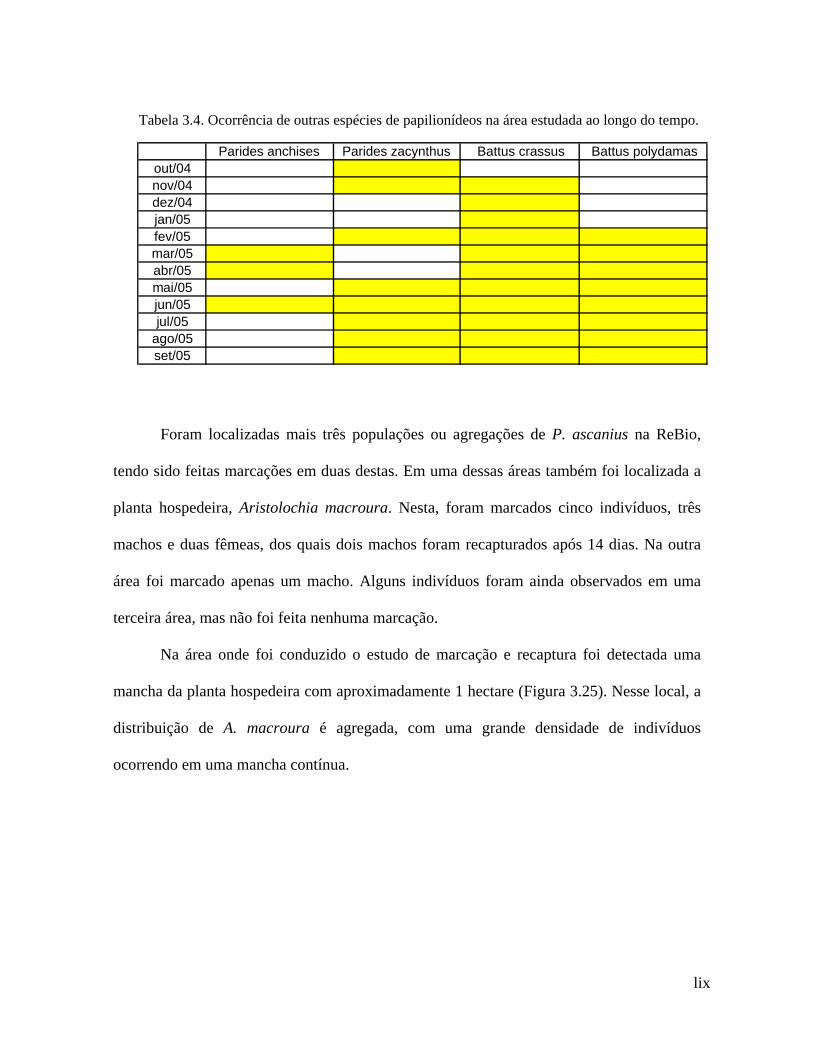
As espécies Parides zacynthus, Parides anchises, Battus crassus e Battus
polydamas, cujas larvas também se alimentam de A. macroura, embora não
exclusivamente, constituindo-se assim potenciais competidoras de P. ascanius, foram vistas
voando durante praticamente todo o ano na Reserva Biológica de Poço das Antas, na
mesma área de amostragem de P. ascanius (Tabela 3.4).
0
2
4
6
8
10
1--6 7--12 13--18 19--24 25--30
classes de dias
abun
dânc
ia/e
sfor
ço
Figura 3.24. Número de indivíduos recapturados em cada classe de dias após a captura levando em consideração o esforço.
lix
Foram localizadas mais três populações ou agregações de P. ascanius na ReBio,
tendo sido feitas marcações em duas destas. Em uma dessas áreas também foi localizada a
planta hospedeira, Aristolochia macroura. Nesta, foram marcados cinco indivíduos, três
machos e duas fêmeas, dos quais dois machos foram recapturados após 14 dias. Na outra
área foi marcado apenas um macho. Alguns indivíduos foram ainda observados em uma
terceira área, mas não foi feita nenhuma marcação.
Na área onde foi conduzido o estudo de marcação e recaptura foi detectada uma
mancha da planta hospedeira com aproximadamente 1 hectare (Figura 3.25). Nesse local, a
distribuição de A. macroura é agregada, com uma grande densidade de indivíduos
ocorrendo em uma mancha contínua.
Tabela 3.4. Ocorrência de outras espécies de papilionídeos na área estudada ao longo do tempo.
Parides anchises Parides zacynthus Battus crassus Battus polydamasout/04nov/04dez/04jan/05fev/05mar/05abr/05mai/05jun/05jul/05
ago/05set/05

lx
Figura 3.25. Recorte de imagem de satélite (IKONOS) representando a área de estudo. Em roxo: transecto em que foram realizadas as capturas. Em verde claro: mancha de Aristolochia macroura.

lxi
3.4 Discussão
3.4.1 Variação temporal e tamanho populacional
O número de indivíduos de Parides ascanius capturados por dia (NICD: 1 a 53) foi
de forma geral bastante alto ao longo de todo o ano, se comparado com outras espécies de
Parides. Estudando uma população de Parides anchises nephalion na cidade de São
Vicente (SP), Freitas & Ramos (2001) registraram de 1 a 11 indivíduos por dia, e
encontraram um ligeiro aumento populacional em dezembro com posterior declínio da
população em janeiro e fevereiro, sendo as maiores abundâncias encontradas nos meses
mais quentes e úmidos. Valores próximos a 10 indivíduos foram registrados nos meses de
menor abundância de P. ascanius no presente estudo.
O único outro estudo de marcação e recaptura com essa espécie, realizado no ano de
1987 em Barra de São João (RJ), por Otero & Brown, foi responsável pela marcação de 100
indivíduos entre os meses de agosto e dezembro, com o máximo de 19 indivíduos
registrados por dia, o que ocorreu no mês de outubro (ver Tyler et al., 1994). Há evidências
de que a agregação/população na ReBio Poço das Antas é maior, porém, como não foi
mencionada a metodologia utilizada naquele estudo, não é possível estabelecer
comparações com precisão.
Os dados da variação temporal de P. ascanius na ReBio sugerem que esta espécie
apresenta um pico populacional na primavera, tendo outro pico no final do outono. Não
houve nenhuma relação significativa entre os fatores climáticos e a abundância de
indivíduos. Entretanto, a influência dos fatores climáticos sobre o tamanho populacional de
P. ascanius pode ser indireta, através da relação da temperatura e pluviosidade com a

lxii
abundância dos recursos alimentares e com as taxas de parasitismo.
O índice de recaptura, de forma geral, foi baixo, já que apenas 17% dos indivíduos
marcados foram recapturados, e em metade das amostragens não houve recapturas. Sendo
assim, as estimativas populacionais calculadas com base em recapturas devem ser tratadas
com cautela. Esse índice poderia ter sido mais alto se o intervalo entre as coletas tivesse
sido menor. Em vários meses, as estimativas populacionais tiveram valores próximos do
dobro do número de machos capturados. Devido à dificuldade para se obter bons índices de
recaptura para troidíneos, e uma maior capturabilidade de machos, sugere-se o uso do
dobro do número destes como uma boa medida para estudos de dinâmica de populações.
Talvez seja meio imprudente discutir algum padrão sazonal para essa espécie na
ReBio, visto que o estudo foi conduzido por apenas um ano, e que o pequeno número de
amostras por mês, um ou dois, o que pode fazer com que determinado mês seja
caracterizado por uma amostragem muito pontual e que pode não representar o conjunto
das condições climáticas daquele mês. Outra dificuldade imposta ao estabelecimento de
algum padrão sazonal é a grande oscilação populacional e intensa atividade das borboletas.
Entretanto, o fato de adultos jovens de ambos os sexos de P. ascanius terem voado durante
todo o ano na ReBio, indica que esta espécie está se reproduzindo em todas as estações.
Essa é uma importante informação, uma vez que não é incomum que essa borboleta entre
em diapausa na fase de pupa, especialmente em épocas frias (Otero & Brown, 1986).
3.4.2 Razão sexual
A razão sexual desviada para machos, salientada pelas diferentes taxas de recaptura
dos sexos, encontradas para P. ascanius na ReBio, corroboram a hipótese de
capturabilidade diferencial entre sexos para espécies de borboletas. Segundo tal hipótese, a

lxiii
diferença de captura entre os sexos ocorreria por causa de diferenças comportamentais dos
adultos, com machos voando nos mesmos lugares mais freqüentados por pesquisadores
(trilhas abertas, com maior incidência da luz solar e espaço para vôo), e fêmeas mais
dispersas pelo habitat, à procura de plantas hospedeiras (Ehrlich, 1984). Pode-se especular
ainda que os machos de P. ascanius sejam mais ativos do que as fêmeas, o que poderia
estar relacionado com a poligamia destes, e uma conseqüente intensa procura por fêmeas.
No entanto, o registro em uma amostra de razão sexual desviada para fêmea, aliado
ao fato de uma grande variação das proporções de machos e fêmeas entre as coletas,
também pode sugerir um efeito da aleatoriedade demográfica.
Em 1987, em Barra de São João (RJ), foi registrada uma razão sexual para P.
ascanius de aproximadamente 1,86 machos para cada fêmea (Tyler et al., 1994). Freitas &
Ramos (2001) encontraram uma razão sexual de 2,5 machos para cada fêmea para P.
anchises nephalion, proporção semelhante à encontrada por nosso estudo (3:1), tendo essa
razão variado de 57% a 100% dos machos ao longo dos diferentes meses.
Parece provável que as razões sexuais nas populações de borboletas mudem
freqüentemente com condições ecológicas de geração para geração (Ehrlich, 1984).
3.4.3 Distribuição etária e dano alar
A maior proporção de indivíduos na faixa etária intermediária pode significar,
simplesmente, que essa faixa de idade seja mais ampla, isto é, contemple um intervalo de
idades maior. É mais recomendável, por isso, analisar a distribuição etária ao longo do
tempo. No caso de P. ascanius, a proporção de indivíduos nas diferentes classes etárias não
variou muito ao longo do ano, tendo o padrão diferido mais nos meses em que a abundância

lxiv
foi menor, o que possivelmente pode ser explicado por variação estocástica.
A maior porcentagem encontrada de espécimens de P. ascanius sem nenhum dano
nas asas parece estar de acordo com o aposematismo e a impalatabilidade dessa espécie, o
que reduziria a pressão de predação sobre esta.
Analisando machos e fêmeas separadamente, foi encontrada uma proporção maior
de fêmeas do que machos velhos enquanto que machos novos foram mais comuns que
fêmeas novas. Além disso, registrou-se também uma maior porcentagem de fêmeas com
muito dano nas asas e machos com nenhum dano alar. Esse padrão portanto, poderia indicar
que os horários e/ou locais de amostragem favoreceram a captura de machos de P.
ascanius.
A correlação entre a faixa etária e o dano alar, tanto para machos como para fêmeas
de P. ascanius na ReBio, pode sugerir que ambos os sexos são igualmente ativos ou mesmo
predados. Por se tratar de uma espécie aposemática e impalatável, acredita-se que a maior
parte dos danos nas asas corresponda a injúrias provocadas pela movimentação por entre a
vegetação.
3.4.4 Ocupação do habitat e movimentação
As capturas não ocorreram de forma regular ao longo do transecto, existindo
claramente áreas de maior probabilidade de captura do que outras. Da mesma maneira, uma
grande parte das recapturas também ocorreu nestas regiões. Agregando-se ainda os dados
de movimentação, inferidos a partir das recapturas, há fortes indícios de se tratar de uma
unidade demográfica claramente definida, com os indivíduos de P. ascanius ocupando

lxv
determinados locais do habitat.
As distribuições espaciais de machos e fêmeas foram um pouco diferentes,
principalmente no que se refere aos trechos mais utilizados por P. ascanius, sugerindo um
padrão distinto no uso do espaço. As fêmeas, de todas as idades, foram mais capturadas na
região do “morrote”, o que poderia estar ligado a um maior sombreamento e,
conseqüentemente, maior umidade, o que pode estar associado a sítios de reprodução. Já os
machos, foram mais capturados na região da “baixada”, porém foram detectadas, nesse
caso, algumas diferenças quando se consideraram as faixas etárias. A maioria dos machos
novos foi encontrada na região do “morrote”, enquanto que os machos velhos foram
capturados principalmente na área de “baixada”. Isso poderia ser explicado por diferenças
comportamentais entre machos e fêmeas, ou diferentes requisitos de microhabitat.
A importância da área do “morrote” fica ainda mais evidente quando se considera
que lá as borboletas voam a uma altura maior, tornando a capturabilidade menor nesse
local. Levando isso em consideração, poderia se especular que essa área tem uma ocupação
maior ainda. Nessa região, encontra-se uma grande quantidade de recursos alimentares dos
adultos.
Na ReBio, a distância de vôo máxima encontrada para P. ascanius foi de 800
metros, porém essa distância só foi registrada para um indivíduo, sendo a média de
movimentação muito menor. Para P. anchises a distância de vôo máxima registrada variou
de 140 a 1500 metros, porém as médias devem ser um parâmetro mais representativo da
população do que valores máximos (Brown Jr et al., 1994).
Outro dado que reforça ainda mais a proposição de uma unidade demográfica
claramente definida para P. ascanius na ReBio é a ausência de correlação entre a distância
percorrida e os dias decorridos da primeira captura. Grande parte dos movimentos de

lxvi
maior distância (maiores que 200 metros) ocorreu justamente entre as duas principais áreas
de captura (“morrote” e “baixada”). Além desse resultado, essa movimentação também
pode ser largamente observada no campo, ficando bastante claro que os indivíduos de P.
ascanius na ReBio se locomovem através de rotas de movimentação.
A menor proporção de indivíduos novos entre os recapturados, em relação ao total
de espécimens capturados levanta algumas questões. Uma menor proporção de indivíduos
velhos entre os recapturados seria naturalmente esperada, uma vez que o intervalo entre as
amostras foi, em geral, semelhante à expectativa de vida dessa espécie. A menor proporção
de machos novos entre os indivíduos recapturados pode indicar uma maior dispersão desses
indivíduos. Foi demonstrado que o comportamento agressivo de machos de papilionídeos
territorialistas pode aumentar a dispersão de machos novos (Lederhouse, 1983).
A maior proporção de borboletas com nenhum dano nas asas entre as recapturadas,
assim como a maior proporção dessa classe na maioria das faixas de distância, sugere que a
ocorrência de danos nas asas influencia negativamente a movimentação de P. ascanius.
Pouca quantidade de dano alar, por sua vez, parece não contribuir para uma menor
locomoção dessa espécie, visto que uma maior proporção de indivíduos com pouco dano
foi recapturada a grandes distâncias. Outro fato que chama a atenção é a menor proporção
de borboletas recapturadas com nenhum dano nas asas em relação a essa porcentagem no
total das capturas, o que de certa forma é contraditório. Uma possível explicação é que
indivíduos “geneticamente sedentários” teriam naturalmente uma menor incidência de
danos nas asas devido à baixa mobilidade. Já espécimens “geneticamente mais móveis”, ou
seja, mais propensos a se locomover, teriam danos nas asas pela própria movimentação. E
quando fosse atingido um nível grande de dano alar sua locomoção seria reduzida. Uma
outra hipótese é que os machos com nenhum dano teriam uma grande mobilidade e

lxvii
capacidade dispersora, de forma que uma parte destes dispersasse em busca de novas áreas,
principalmente nas épocas de altas densidades populacionais. Tal explicação pode fazer
mais sentido ainda quando se considera que as fêmeas de P. ascanius são monogâmicas, só
acasalando uma única vez, e os machos poligâmicos. Assim, em situações de altas
densidades, poderia ser vantajoso para alguns machos dispersar para novas regiões,
principalmente os com emergência retardada.
3.4.5 Longevidade
O tempo de permanência médio dos indivíduos de P. ascanius, com maior
proporção de recapturas de 7 a 12 dias após a captura, foi semelhante ao reportado na
literatura (Otero & Brown, 1986), porém a longevidade máxima registrada foi bem
diferente. Esses autores encontraram uma longevidade média de 10 dias, com um máximo
de 14 dias, porém em uma situação artificial. Na ReBio, a longevidade máxima registrada
foi de 28 dias, sendo essa provavelmente maior se considerado que o indivíduo já estava
vivo e que assim permaneceu após a segunda captura.
A maior longevidade encontrada para machos, em relação às fêmeas, de P. ascanius
na ReBio pode não representar a realidade, uma vez que a taxa de recaptura das fêmeas foi
bem menor do que de machos. Portanto, a maior longevidade de machos poderia ser apenas
um artefato de coleta.
O tempo máximo de permanência registrado para a maioria das espécies de Parides
é por volta de 6 a 8 semanas em ambiente natural, exceto para P. anchises, P. ascanius e P.
neophilus, que apresentam valores máximos de permanência de 2 a 4 semanas (Tyler et al.,
1994). Cook et al. (1971) encontraram uma expectativa de vida de 5,7 dias para P.

lxviii
neophilus, enquanto Freitas & Ramos (2001) registraram uma expectativa de vida de 5,6
dias para P. anchises.
O tempo médio de residência para as espécies de Parides pode ser considerado
baixo se comparado com o de outras borboletas tropicais miméticas como os Heliconini
(Ehrlich & Gilbert, 1973; Ramos & Freitas, 1999) e Ithomiinae (Freitas 1993, 1996).
Brown Jr et al. (1981) citaram a baixa expectativa de vida dos adultos associada a
amostragem semanais (resultando em baixas taxas de recaptura) como a razão principal
para a não utilização de estimativas no estudo de Troidini (Freitas & Ramos, 2001).
Segundo esses autores, futuros estudos com populações de Troidini devem considerar isso,
devendo a população ser estudada em intervalos menores do que uma semana.
A obtenção de informações precisas sobre a longevidade de adultos em populações
naturais de borboletas é extremamente difícil. Devido à dispersão, o tempo de residência
baseado em estudos de marcação e recaptura devem ser uma subestimativa do tempo de
vida de adultos (Komonen et al., 2004).
3.4.6 Conservação
O conhecimento da distribuição da planta hospedeira na área de estudo é uma
informação de grande valor, pois possibilita a integração do recurso em possíveis ações de
manejo da P. ascanius. A presença de espécies competidoras parece não limitar muito a
população de P. ascanius na ReBio, visto que esta voa o ano inteiro. Entretanto, para que se
tenha maior clareza quanto ao papel da competição pelo recurso alimentar da lagarta seria
desejável um acompanhamento no campo dos estágios imaturos dessa espécie e das
potenciais competidoras, levando-se em conta ainda a fenologia e estrutura espacial de A.

lxix
macroura, no intuito de avaliar também a competição intra-específica. O monitoramento
dos estágios imaturos também é de fundamental importância para a compreensão dos
mecanismos que afetam a mortalidade dessa borboleta, o que facilitaria enormemente o
entendimento da estrutura espacial e variação temporal dessa espécie.
Para um maior enriquecimento das informações sobre a viabilidade de P. ascanius
na Rebio seria interessante ainda uma quantificação e qualificação mais apurada da
distribuição espacial e temporal dos recursos alimentares dos adultos e da competição, intra
e interespecífica, pelo uso do espaço.
A localização de outras populações ou manchas foi um registro importante, uma vez
que permite o monitoramento destas, o que pode fornecer dados complementares, inclusive
sobre a provável dinâmica metapopulacional dessa borboleta na ReBio.

lxx
3.5 CONCLUSÕES
A população de P. ascanius estudada parece estar em boas condições nessa parte da
reserva, uma vez que é abundante ao longo de boa parte do ano, mesmo na presença de
espécies potencialmente competidoras. Essa população está localizada em uma área
bastante favorável, com uma grande quantidade dos recursos alimentares das lagartas e dos
adultos. A região ocupada fornece ainda microclimas bastante variados em uma área
relativamente curta, facilitando a permanência de indivíduos no local.
Provavelmente esta é a maior população remanescente de P. ascanius, podendo os
estudos realizados nessa área servir de base para comparações com outras populações e
para o estabelecimento de estratégias de manejo para essa espécie. Trata-se de um núcleo
importante de indivíduos com potencial para fornecer espécimens colonizadores para áreas
satélites da reserva. Representa também um banco genético importante para troca ou
“reforço genético” da variabilidade de outras populações pequenas e isoladas próximas ao
Rio de Janeiro.
Quanto ao efeito da represa de Juturnaíba sobre P. ascanius pouco se pode dizer,
pois faltam dados anteriores sobre a distribuição espacial dessa espécie na ReBio. Um
efeito positivo poderia ser sugerido na área onde o estudo foi conduzido, uma vez que
sofreu alagamento com a construção da represa. Porém, esse empreendimento promoveu
uma radical drenagem à jusante, ocasionando a modificação de parte do ecossistema, que
era de matas inundáveis (área de cerca de 900 ha), onde hoje ocorrem os grandes incêndios
da Reserva. Nessas áreas é esperado um efeito negativo sobre P. ascanius através da
redução do habitat de sua planta hospedeira.

lxxi
A verificação do tamanho da população de P. ascanius na Reserva Biológica de
Poço das Antas não é suficiente para dizer alguma coisa em relação ao status de
conservação dessa espécie. Para isso, seria desejável não só um monitoramento dessa
população na ReBio junto à investigação de uma provável estrutura metapopulacional, mas
como uma avaliação da estrutura e dinâmica do maior número possível de manchas
populacionais no estado do Rio de Janeiro.

lxxii

lxxiii
4.1 INTRODUÇÃO
Desde muito cedo, a possibilidade de registrar em filme processos naturais,
procedimentos técnicos e/ou científicos, aspectos corriqueiros ou mesmo raros, instigou os
educadores para a sua utilização como ferramenta auxiliar no processo educativo. O filme
científico strictu sensu é produzido no âmbito da pesquisa científica e cumpre papéis
específicos, tais como: documentar o trabalho ou objeto da pesquisa, para arquivamento ou
para o uso na divulgação e ensino. Pode ainda, ter uma função investigativa, mormente,
quando se utilizam técnicas especiais, como a microfilmagem ou a filmagem em alta
velocidade (Santos, 2005).
O aprimoramento das técnicas de filmagem e a crescente produção de filmes
científicos alavancaram a indústria de produção de filmes para o uso no ensino. Alguns
países, inclusive o Brasil, criaram órgãos próprios para a produção de filmes educativos
(Schvarzman, 2001).
O objetivo do montador de documentários é criar uma atmosfera – dramatizar
acontecimentos. Para o montador de filmes educativos isto é, em grande parte, irrelevante,
devendo ser seus objetivos a clareza, a exposição lógica e a avaliação correta da
receptividade da platéia (Reisz & Millar, 1978).
Todo o filme nasce de uma idéia inicial, sendo esta melhorada e ampliada até que se
chegue a um argumento que contenha os elementos básicos do filme: do que trata a história,
onde serão feitas as filmagens, o tempo de duração do filme, como será contada a história e
para quem. Em seguida, vem a fase em que esses elementos serão trabalhados do ponto de
vista técnico e de linguagem, procedimentos que chamamos de roteiro. O próximo passo é a
obtenção das imagens e dos sons, que deve seguir da melhor forma possível o roteiro,

lxxiv
vindo, na seqüência, a fase de montagem (Santos, 2005). Segundo Aumont et al. (1995), a
montagem é “o princípio que rege a organização de elementos fílmicos visuais e sonoros ou
de agrupamentos de tais elementos, justapondo-os, encadeando-os e/ou organizando sua
duração”. A montagem é de grande importância na confecção de um filme, uma vez que
influencia na linguagem, pois ela introduz códigos que irão facilitar ou, em alguns casos,
viabilizar o entendimento do filme. Seguindo ou não um roteiro, é através da montagem
que se vai criando a narrativa. Com ela podemos direcionar a percepção e influenciar os
elementos psicológicos que nos dão a sensação de temporalidade (Santos, 2005).
Segundo Sluys (1922, apud Serrano e Venâncio Filho,1930) a duração máxima das
projeções deve ser de 20 minutos para crianças de menos de 12 anos e 30 minutos para
pessoas com mais de 12 anos. Isto estaria relacionado com o fato do núcleo da formação
reticular, que tem papel na manutenção da atenção, só se tornar inteiramente mielinizado na
puberdade ou depois dela (Carter, 2003).
Santos (2005) analisou 3.725 audiovisuais científicos, obtidos de 10 diferentes
acervos, potencialmente utilizáveis no contexto educacional, produzidos nas últimas seis
décadas, veiculados em mídias e linguagens distintas. Apesar da maior parte (n = 1.809 ou
57,16%) dos audiovisuais de acervos acessíveis aos professores ser de cunho didático, a
grande maioria (n=2.497 ou 78,89%) desses mesmos acervos é de origem internacional, o
que sugere que os professores tenham ao seu dispor um material descontextualizado,
distante da realidade dos alunos.
Entre os professores do Colégio Pedro II (n = 33) que fazem uso de audiovisuais em
suas aulas, a maioria declarou preferir o gênero documentário. Considerando que
documentários, filmes e reportagens têm o caráter de divulgação, os resultados indicam que
66,66% desses professores não fazem uso de audiovisuais didáticos (Santos, 2005).

lxxv
A divulgação de audiovisuais científicos tem tomado força nos últimos 50 anos,
sendo que nos últimos 10 esse processo tem se intensificado, mormente, com a colocação
dos canais especializados em ciência que são veiculados pelas televisões por assinatura.
Esses canais fechados têm investido em um nicho que era anteriormente ocupado por
horários específicos em redes de televisão aberta e têm compromisso, como os próprios
alegam, de veicular uma programação de excelência. E trabalham com materiais
desenvolvidos por produtoras internacionais de grande experiência e qualidade.
No Brasil, um recorte dessa trajetória pode ser visto com a veiculação em horários
definidos de uma programação cientifica em vários canais de televisão aberta (TV Globo,
Rede Record, TV Cultura SP, RedeTV, TVE RJ), e, mais intensamente, na televisão
fechada (TV Sesc/Senac, TV Universitária, TV Escola). O fenômeno da exploração
desses nichos de divulgação visa alcançar uma fatia definida de público.
Nos últimos 10 anos um novo veículo vem ameaçando a divulgação televisiva. A
internet, ou rede mundial de computadores, permite a disponibilização e acesso a arquivos
de vídeo cada vez com melhor qualidade, de forma rápida e barata, não só para quem
acessa mas também para quem produz. Considerando as facilidades proporcionadas pelas
novas tecnologias audiovisuais, hoje em dia, qualquer instituição ou mesmo pessoa pode
produzir material de divulgação.
A divulgação de um documentário sobre uma espécie ameaçada de extinção deve se
constituir como uma valiosa ferramenta para a conservação desta, levando um público
maior a entrar em contato com noções de sua biologia e ecologia. Em um contexto mais
amplo, isto poderia levar também a uma maior conscientização ambiental e uma maior
percepção da problemática da destruição de habitats e extinção de espécies.

lxxvi
Muitas espécies ameaçadas de extinção são de difícil visualização, o que torna mais
complicada a realização de um audiovisual. Em relação a insetos esta situação é ainda mais
acentuada, tanto pelas dificuldades da macrofilmagem como pela falta de apelo estético do
grupo (Gruzman, 2003). Desta maneira, qualquer contribuição neste sentido é muito bem
vinda, aumentando o conhecimento popular sobre eles e desmistificando os preconceitos
que rondam os insetos há anos.
Nesse sentido, objetivou-se produzir um documentário sobre Parides ascanius para
divulgação científica nas unidades de conservação do estado do Rio de Janeiro,
principalmente naquelas nas quais P. ascanius ocorre, e nos principais veículos de mídia
audiovisual (televisão e internet). Este documentário também deve ser usado para educação
ambiental de alunos do ensino fundamental e médio, podendo ainda ser utilizado em aulas
de cursos de graduação de biologia e áreas afins.

lxxvii
4.2 MATERIAIS E METÓDOS
A obtenção das imagens foi feita principalmente pelo documentarista científico
Roberto Eizemberg dos Santos, mas também contou com a colaboração da fotógrafa
Márcia Franco e de integrantes do Laboratório de Ecologia de Insetos do Departamento de
Ecologia da UFRJ. As filmagens foram realizadas tanto em ambiente natural (Reserva
Biológica de Poço das Antas, Parque Ecológico de Marapendi, beira de estrada em Magé,
sítio do Dr. Luiz Otero) como em ambientes controlados (borboletário do Dr. Luiz Otero,
laboratório, estúdio do Roberto Eizemberg) (Figuras 4.1 e 4.2).
Figura 4.1. Márcia Franco e Roberto Eizemberg filmando uma cópula de Parides ascanius no sítio do Dr. Luiz Otero.

lxxviii
A produção deste documentário contou não apenas com a colaboração científica do
Dr. Luiz Otero, mas também com a sua ajuda para a filmagem do ciclo de vida da espécie.
O tempo total de imagens capturadas foi de aproximadamente quatro horas.
Os equipamentos utilizados foram três filmadoras digitais padrão miniDV e duas
câmeras fotográficas digitais. A edição das imagens foi feita em duas ilhas de edição não
linear, com o programa Premiere 6.5 e o documentário ficou com oito minutos e vinte
segundos.
Figura 4.2. Câmera, iluminação e cenário preparados para a filmagem da emergência do adulto de P. ascanius, no laboratório de Ecologia de Insetos da UFRJ.

lxxix
4.3 RESULTADOS
O documentário foi produzido em DVD e CD (Anexo 1).

lxxx
4.4 DISCUSSÃO
O resultado obtido com o documentário ainda está para ser submetido à apreciação
do público, mas espera-se que este seja uma importante ferramenta para a conservação de
Parides ascanius, através da divulgação de informações científicas e conscientização de um
maior número de pessoas.
Apesar da proteção legal dessa espécie, a execução das leis é ineficiente, pois os
oficiais responsáveis pela implementação da legislação são, na maioria das vezes, incapazes
de identificar as espécies de borboletas, e a maioria das pessoas não sabe que as borboletas
são protegidas. Além disso, a falta de uma real consciência ambiental faz, muitas vezes, os
próprios órgãos públicos passarem por cima das questões ambientais em favor de questões
econômicas. A divulgação do documentário pode se inserir muito bem dentro deste
contexto, facilitando o treinamento de oficiais responsáveis pelo cumprimento da
legislação, e estimulando esses oficiais a realizar estudos com essa espécie, facilitando o
trabalho de monitoramento das populações.
A divulgação do documentário em todas as unidades de conservação do estado e
para estudantes de ensino fundamental, médio e superior, além da população em geral, via
televisão e internet, pode orientar e educar os visitantes e moradores de áreas onde P.
ascanius ocorre, e alertar os cidadãos para a problemática dessa espécie e de todo o seu
habitat.
O material fotográfico produzido durante a realização do documentário pode ser
distribuído e utilizado para facilitar a correta identificação das espécies por parte dos
oficiais responsáveis pelo cumprimento legislação ambiental, e alertar comunidades

lxxxi
humanas localizadas em áreas de possível ocorrência da espécie para a situação dessa
borboleta, com o intuito de ganhar novos parceiros para sua conservação.

lxxxii

lxxxiii
5. CONSIDERAÇÕES FINAIS
É difícil prever precisamente o risco de extinção que Parides ascanius sofre. Na
Reserva Biológica de Poço das Antas, a população parece apresentar uma boa condição.
Entretanto, essa pode ser a maior população desta espécie, havendo várias pequenas
populações seriamente ameaçadas pela destruição do habitat ao longo de toda sua
distribuição.
No caso de P. ascanius, a principal causa última de extinção é a destruição de
habitat, o qual vem sendo sistematicamente descaracterizado ao longo de toda distribuição
dessa borboleta. Quanto às causas próximas de extinção de P. ascanius, nenhum estudo
ainda foi feito especificamente neste sentido. Este trabalho sugere que tanto a aleatoriedade
ambiental como a demográfica poderiam oferecer algum risco para essa espécie, mas novos
estudos precisam ainda ser feitos nessa área, assim como enfocando também a deterioração
genética e a disfunção social.
Uma espécie naturalmente rara, como P. ascanius, pode ser mais vulnerável à
extinção por causas últimas, ou declínio de suas populações, porém de alguma maneira
estar adaptada para lidar melhor com as causas próximas da extinção, ou seja, persistir
melhor em pequenas populações isoladas. Entretanto, vale lembrar que nenhum estudo
relacionado às causas próximas da extinção foram feitos com essa espécie, e que a
crescente degradação do seu habitat continua a ameaçar também as pequenas populações.
No caso dessa borboleta, parece não haver tendência para a utilização de mais
recursos, uma vez que já foram feitos experimentos com outras espécies de Aristolochia e
verificou-se que P. ascanius é mesmo monófaga em A. macroura. Assim, tanto a restrita

lxxxiv
distribuição geográfica como a monofagia tornam essa borboleta propensa
à extinção.
A indiscriminada destruição de habitat pode deixar populações isoladas de grande
importância para conservação em hábitats marginais. Tais populações podem ter taxas de
crescimento muito baixas, tornando sua sobrevivência ainda mais precária. Em casos
extremos, algumas populações podem persistir somente devido à imigração da fonte; a
destruição dessa fonte leva à extinção final das populações satélites, tornando sua
conservação impossível. Se, os números populacionais decaem devido à queda da taxa de
natalidade ou aumento da taxa de mortalidade, espera-se que a distribuição se contraia de
qualquer maneira, mesmo não havendo destruição de habitat. Se a distribuição original
tinha um centro único e bem definido, as distribuições devem colapsar na direção do centro
(Lawton, 1994).
Esse pode ser o caso de algumas populações de P. ascanius, que com a intensa
destruição de seu habitat podem ter ficado isoladas em áreas marginais e ainda sujeitas a
fortes impactos antrópicos. Pode-se imaginar que sua distribuição esteja se contraindo na
direção da ReBio de Poço das Antas, o que evidenciaria ainda mais a importância desta
reserva para a conservação de P. ascanius. A população estudada poderia estar servindo de
fonte de indivíduos para outras manchas dentro da ReBio e, quem sabe, para outras
populações isoladas em áreas sem nenhuma proteção legal. Nesse contexto, a população de
P. ascanius estudada na ReBio de Poço das Antas apresenta importância central, devendo
ser monitorada continuamente, juntamente com a investigação de uma possível estrutura
metapopulacional. Isso poderia fornecer informações de grande relevância para a
conservação dessa espécie, inclusive sobre sua resposta à fragmentação e sobre a
vulnerabilidade às causas próximas da extinção.

lxxxv
Mohanraj & Veenakumari (1996) propuseram 7 recomendações para a conservação
de papilionídeos na Índia: controle do crescimento populacional; proteção mais efetiva de
florestas em unidades de conservação; identificação e proteção legal de manchas de
vegetação fora de unidades de conservação que abriguem espécies raras, endêmicas ou
ameaçadas; monitoramento para acessar status das espécies, permitindo a formulação de
medidas de conservação mais apropriadas; treinamento de oficiais responsáveis pelo
cumprimento da legislação; estimulação desses oficiais para a realização de estudos com as
borboletas, o que pode ser feito em parceria com instituições de pesquisa; criação de
parques de borboletas, permitindo a educação ambiental de crianças e adultos. Muitas
destas propostas podem ser aplicadas à conservação de muitas espécies, não só de
papilionídeos, mas também de borboletas e insetos em geral. Para a proteção efetiva de
determinada espécie, são desejáveis propostas mais específicas.
Apresento como propostas para a conservação de Parides ascanius: (1) proteção
mais efetiva das restingas e florestas de baixada em unidades de conservação federais,
estaduais e municipais do estado do Rio de Janeiro; (2) identificação e proteção legal de
manchas de vegetação fora de unidades de conservação que abriguem essa espécie; (3)
monitoramento detalhado da borboleta e sua planta hospedeira na ReBio Poço das Antas;
(4) busca por novas populações da borboleta ao longo e nos limites de sua distribuição; (5)
monitoramento do maior número possível de populações para acessar status de conservação
da espécie, permitindo a formulação de medidas de manejo mais apropriadas; (6)
treinamento de oficiais responsáveis pelo cumprimento da legislação, inclusive com a
utilização do documentário; (7) estimulação desses oficiais para a realização de estudos
com essa espécie, o que pode ser feito em parceria com instituições de pesquisa, facilitando
o trabalho de monitoramento das populações; (8) divulgação do documentário em todas as

lxxxvi
unidades de conservação do estado, orientando e educando os visitantes e moradores dessas
áreas; (9) divulgação do documentário para estudantes de ensino fundamental, médio e
superior; (10) divulgação do documentário e material escrito para a população em geral, via
televisão, jornal e internet, alertando para a problemática dessa espécie e de todo o seu
habitat; (11) criação de uma rede de pessoas interessadas na conservação dessa espécie,
criando uma base de dados conjunta; (12) visitas a comunidades humanas localizadas em
áreas de possível ocorrência da espécie, divulgando material fotográfico e alertando para a
situação dessa borboleta, com o intuito de ganhar novos parceiros para sua conservação;
(13) criação de parques de borboletas ou borboletários, permitindo a educação ambiental de
crianças e adultos, podendo servir de fonte para possíveis reintroduções; (14) cultivo de A.
macroura para possíveis replantios.

lxxxvii

lxxxviii
6. BIBLIOGRAFIA GERAL
ACKEREY, P.R.; DE JONG, R. & VANE-WRIGHT, R.I. 1999. The Butterflies: Hedyloidea,
Hesperioidea and Papilionoidea. In: Handbook of Zoology Volume IV Arthropoda:
Insecta. Kristensen, N.P (ed.) Lepidoptera, Moths and Butterflies. Volume 1: Evolution,
Systematics and Biogeography. Part 35. Walter de Gruyter, Berlin, New York.
ANDREWARTHA, H. G. & BIRCH, L. C. 1954. The distribution and abundance of animals.
University of Chicago Press.
BROWN JR., K.S. 1992. Borboletas da serra do Japi: diversidade, hábitats, recursos
alimentares e variação temporal. Pgs.142-187, In Morellato, L.P. (org.) História natural
da Serra do Japi. Ecologia e preservação de uma área florestal no Sudeste do Brasil.
Ed. UNICAMP, SP, 321p.
BROWN JR., K.S. 1997. Diversity, disturbance, and sustainable use of Neotropical forests:
insects as indicators for conservation monitoring. Journal of Insect Conservation 1: 25-
42.
BROWN-JR, K. S.; DAMMAN, A. J. & FEENY, P. 1981. Troidine swallowtails (Lepidoptera:
Papilionidae) in southeastern Brazil: natural history and foodplant relationships. Journal
of Research on the Lepidoptera 19: 199-226.
BROWN-JR, K. S.; KLITZKE, C. F.; BERLINGERI, C. & ROBBO DOS SANTOS, P. E. 1994.
Neotropical swallowtails: chemistry of foodplant relationships, population ecology, and
biosystematics, pp.405-445. In: J. M. Scriber, Y. Tsubaki & R. C. Lederhouse (eds.),
Swallowtail butterflies: Ecology and Evolutionary Biology. Scientific Publishers,
Gainville.
BROWN JR., K.S. & FREITAS, A.V.L. 2000. Atlantic Forest butterflies: indicators for

lxxxix
landscape conservation. Biotropica 32 (special issue): 934-956.
CAMPOS NETO, F.C. & OTERO, L.S. 2001. Parides ascanius. Ecologia e Desenvolvimento
96: 32-35.
CARTER, R. 2003. O livro de ouro da mente (título original: mapping the mind). Rio de
Janeiro: Ediouro.
CAUGHLEY, G. 1994. Directions in conservation biology. Journal of Animal Ecology 63:
COLLINGE, S.K. 2001. Spatial ecology and biological conservation. Biological
Conservation 100: 1-2.
COOK, L. M.; FRANK, K. & BROWER, L. P. 1971. Experiments on the demography of
tropical butterflies. I. Survival rate and density in two species of Parides. Biotropica 3:
17-20.
DENNIS, R. L. H.; SHREEVE, T. G. & VAN DYCK, H. 2003. Towards a functional resource-
based concept for habitat: a butterfly biology viewpoint. Oikos 102: 417-426
EHRLICH, P. R. 1984. The structure and dynamics of butterfly populations, pp.25-40. In: R.
I. Vane-Wright & P. R. Ackery (eds.), The Biology of Butterflies. Academic Press,
London.
EHRLICH, P. R. & GILBERT, L. E. 1973. Population structure and dynamics of the tropical
butterfly Heliconius ethilla. Biotropica 5: 69-82.
EHRLICH, P. R.; MURPHY, D. D.; SINGER, M. C.; SHERWOOD, C. B.; WHITE, R. R. &
BROWN, I. L. 1980. Extinction, reduction, stability and increase: the responses of
checkerspot butterfly (Euphydryas) populations to California drought. Oecologia. 46:
101-105.
FEENY, P. 1991. Chemical constraints on the evolution of swallowtail butterflies Pp 315-
340. In: Price, PW, Lewinsohn TM, Fernandes GW, Benson WW (eds). Plant-animal

xc
interactions: evolutionary ecology in tropical and temperate regions. New York, John
Wiley & Sons.
FREITAS, A. V. L. 1993. Biology and population dynamics of Placidula euryanassa, a relict
ithomiine butterfly (Nymphalidae: Ithomiinae). J. Lepid. Soc. 47: 87-105.
FREITAS, A. V. L. 1996. Population biology of Heterosais edessa (Nymphalidae) and its
associated Atlantic Forest Ithomiinae community. J. Lepid. Soc. 50: 273-289.
FREITAS, A. V. L. & RAMOS R. R. 2001. Population biology of Parides anchises nephalion
(Papilionidae) in a coastal site in southeast Brazil. Braz. J. Biol. 61(4): 623-630.
FUTUYMA, D.J. 1997. Biologia evolutiva. 2 ed. Sociedade Brasileira de Genética/CNPq,
Ribeirão Preto, 646pp.
GALL, L. F. 1985. Measuring the size of Lepidopteran populations. J. Res. Lepid. 24: 97-
116.
GRAPHPAD INSTAT. 1997. Version 3.0. GraphPad Software Inc. San Diego, Califórnia.
GRUZMAN, E. 2003. Representações dos insetos através da imagem: uma investigação
teórico-prática para a realização de um vídeo educativo em ecoentomologia. Dissertação
(mestrado) - UFRJ/NUTES. Programa de Pós-graduação em Tecnologia Educacional
nas Ciências da Saúde. Rio de Janeiro.
IBAMA. 2004 (www.ibama.gov.br)
IUCN 2003. The red list of threatened species. Gland, IUCN
KNIGHT, A.; BROWER, L. P. & WILLIAMS, E. H. 1999. Spring remigration of the monarch
butterfly Danaus plexippus (Lepidoptera: Nymphalidae) in north-central Florida:
estimating population parameters using mark-recapture. Biological Journal of the
Linnean Society 68: 531-556.
KOMONEN, A.; GRAPPUTO, A.; KAITALA, V.; KOTIAHO, J. S. & PÄIVINEN, J. 2004. The role

xci
of niche breadth, resource availability and range position on the life history of
butterflies. Oikos 105: 41-54.
KUNIN, W.E. & GASTON, K.J. 1993. The biology of rarity: patterns, causes and
consequences. Trends in Ecology and Evolution 8: 289-301.
LAWTON, J.H. 1994. Population dynamic principles. Philosophical Transactions of the
Royal Society of London B 344: 61-68.
LEDERHOUSE, R.C. 1982. Factors affecting equal catchability in two swallowtail butterflies,
Papilio polyxenes and P. glaucus. Ecological Entomology 7: 379-383.
LEDERHOUSE, R.C. 1983. Population structure, residency and weather related mortality in
the black swallowtail butterfly, Papilio polyxenes. Oecologia 59: 307-311.
LINO, C.F. 1992. Reserva da Biosfera da Mata Atlântica. Plano de Ação. Campinas:
Consórcio Mata Atlântica/UNICAMP.
MEYER-HOZAK, C. 2000. Population biology of Maculinea rebeli (Lepidoptera:
Lycaenidae) on the chalk grassland of Eastern Westphalia (Germany) and implications
for conservation. Journal of Insect Conservation 4: 63-72.
MILLER, J. S. 1987. Host-plant relationships in the Papilionidae (Lepidoptera): Parallel
cladogenesis or colonization? Cladistics 3: 105-120.
MMA 1999. Diretrizes para a política de conservação e desenvolvimento sustentável da
Mata Atlântica. Caderno n 13. Brasília, DF, Brasil.
MMA 2003. Lista nacional das espécies da fauna brasileira ameaçadas de extinção.
Disponível em www.mma.gov.br/port/sbf /fauna em 22/05/2003.
MOHANRAJ, P. & VEENAKUMARI, K. 1996. Host plants, phonologies and status of
swallowtails (Papilionidae), Lepidoptera, in the Andaman and Nicobar islands, Bay of
Bengal, Indian ocean. Biological Conservation 78: 215-221.

xcii
MYERS N.; MITTERMEIER R.A.; MITTERMEIER, C.G.; FONSECA, G.A.B. & KENT, J. 2000.
Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
NEW, T.R. 1997. Are Lepidoptera an effective “umbrella group” for biodiversity
conservation? Journal of Insect Conservation 1: 5-12.
OTERO, L.S. 1984. Parides ascanius (Cramer, 1775), borboleta ameaçada de extinção. In:
Lacerda, L.D.; Araújo, D.S.D.; Cerqueira, R.; Turcq, B. (eds.). Restingas - Origem,
Estrutura, Processos. Pp. 369-371. CEUFF, Niterói.
OTERO, L.S. & BROWN JR, K.S. 1986. Biology and ecology of Parides ascanius (Cramer,
1775) (Lep.: Papilionidae), a primitive butterfly threatened with extinction. Atala 10-12:
2-16.
OTERO, L.S. & MARIGO, L.C. 1990. Butterflies: Beauty and Behavior of Brazilian Species,
128pp. Marigo Comun. Visual, Rio de Janeiro.
OTERO, L.S.; BROWN JR., K.S.; MIELKE, O.H.H.; MONTEIRO, R.F.; COSTA, J.M.; MACÊDO,
M.V.; MACIEL, N.C.; BECKER, J.; SALGADO, N.C.; SANTOS, S.B.; MOYA, G.E.;
ALMEIDA, J.M. & SILVA, M.D. 2000. Invertebrados terrestres. In: A fauna ameaçada de
extinção do Estado do Rio de Janeiro. Bergallo, H.F.; Rocha, C.F.D.; Alves, M.A.S.;
Sluys, M.V. (eds.) Ed. UERJ, FAPERJ, 166 pp.
PIMM, S.L. & RAVEN, P. 2000. Extinction by numbers. Nature 403: 843-845.
PROGRAMA MATA ATLÂNTICA, 2005 (http://www.jbrj.gov.br/pesquisa/pma/), acessado em
15/01/2006.
RAMOS, R. R. & FREITAS, A. V. L. 1999. Population biology, wing color variation and
ecological plasticity in Heliconius erato phyllis (Nymphalidae). J. Lepid. Soc. 53: 11-21.
REISZ, K. & MILLAR, G. 1978. A Técnica da Montagem Cinematográfica. Rio de Janeiro:
Editora Civilização Brasileira S. A.

xciii
RICKLEFS, R.E. 1979. Ecology. 2 ed. Chiron Press, New York.
SANTOS, R.E. 2005. Perfil de Tempo de Audiovisuais Científicos e um Estudo de Caso da
Utilização de Audiovisuais no Ensino de Biologia e Ciências por professores das
Unidades do Colégio Pedro II. Dissertação (mestrado) - Programa de Pós- Graduação
em Química Biológica (Educação, Gestão e Difusão em Biociências), Instituto de
Bioquímica Médica, Universidade Federal do Rio de Janeiro.
SAUNDERS, D.A., Hobbs, R.J. & Margules, C.R. 1991. Biological consequences of
ecosystem fragmentation: a review. Conservation Biology 5: 18-32.
SCHVARZMAN, S. 2001. Humberto Mauro e a constituição da memória do cinema brasileiro.
p 119 – 129. In Ramos, Fernão Pessoa (Org). Estudos de Cinema 2000 – SOCINE. Porto
Alegre: Sulina.
SEMADS. 2002. Atlas das Unidades de Conservação da Natureza do Estado do Rio de
Janeiro. Metalivros, São Paulo.
SERRANO, J. & VENÂNCIO FILHO, F. 1930. Cinema e Educação. São Paulo, Cayeiras, Rio:
Melhoramentos, vol. XIV.
SILVA-BRANDÃO, K.L.; FREITAS, A.V.L.; BROWER, A.V.Z & SOLFERINI, V.N. 2005.
Phylogenetic relationships of the New World Troidini swallowtails (Lepidoptera:
Papilionidae) based on COI, COII and EF-1a genes. Molecular Phylogenetics and
Evolution 36: 468-483.
SIMBERLOFF, D. 1986. The proximate causes of extinction, pp. 259-276. In: D. M. Raup &
D. Jablonski (eds.), Patterns and processes in the history of life. Springer-Verlag, Berlin.
SLUYS, A. La Cinématographie Scolaire et post-scolaire. Bruxelas. 1922.
TABASHNIK, B. E. 1980. Population structure of Pierid butterflies. III. Pest population of
Colias philodice eriphyle. Oecologia 47: 175-183.

xciv
THOMAS, C. D., BODSWORTH, E. J., WILSON, R. J., SIMMONS, A. D., DAVIES, Z. G. &
MUSCHE. 2001. Ecological and evolutionary processes at expanding range margins.
Nature 411: 577-581.
TYLER, H. A.; BROWN-JR, K. S. & WILSON, K. H. 1994. Swallowtail butterflies of the
Americas: a study in biological dynamics, ecological diversity, biosystematics and
conservation. Gainesville, Scientific Publishers, 376 p.
WATT, W. B., CHEW, F. S., SNYDE, R. L. G., WATT, A. G. & ROTHSCHILD, D. E. 1977.
Population structure of pierid butterflies. I. Numbers and movements of some montane
Colias species. Oecologia 27: 1-22.
WEINTRAUB, J.D. 1995. Host plant association patterns and phylogeny in the tribe Troidini
(Lepidoptera: Papilionidae). In: Scriber, J.M., Tsubaki, Y., Lederhouse, R.C. (Eds.),
Swallowtail Butterflies: Their Ecology and Evolutionary Biology. Scientific Publishers,
Gainesville, FL, pp. 307-316.
WILCOX, D.S. & MURPHY, D.D. 1985. Conservation strategy: The effects of fragmentation
on extinction. American Naturalist 125: 879-887.

This document was created with Win2PDF available at http://www.win2pdf.com.The unregistered version of Win2PDF is for evaluation or non-commercial use only.This page will not be added after purchasing Win2PDF.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo





























