Embed Size (px)
Citation preview

Lipid Peroxide-Derived Short-Chain Carbonyls MediateHydrogen Peroxide-Induced and Salt-InducedProgrammed Cell Death in Plants1[OPEN]
Md. Sanaullah Biswas2 and Jun’ichi Mano*
United Graduate School of Agriculture, Tottori University, Tottori 680–8550, Japan (M.S.B., J.M.); and ScienceResearch Center (J.M.) and Graduate School of Agriculture (J.M.), Yamaguchi University, Yamaguchi 753–8515,Japan
Lipid peroxide-derived toxic carbonyl compounds (oxylipin carbonyls), produced downstream of reactive oxygen species (ROS),were recently revealed to mediate abiotic stress-induced damage of plants. Here, we investigated how oxylipin carbonyls causecell death. When tobacco (Nicotiana tabacum) Bright Yellow-2 (BY-2) cells were exposed to hydrogen peroxide, several species ofshort-chain oxylipin carbonyls [i.e. 4-hydroxy-(E)-2-nonenal and acrolein] accumulated and the cells underwent programmedcell death (PCD), as judged based on DNA fragmentation, an increase in terminal deoxynucleotidyl transferase dUTP nick endlabeling-positive nuclei, and cytoplasm retraction. These oxylipin carbonyls caused PCD in BY-2 cells and roots of tobacco andArabidopsis (Arabidopsis thaliana). To test the possibility that oxylipin carbonyls mediate an oxidative signal to cause PCD, weperformed pharmacological and genetic experiments. Carnosine and hydralazine, having distinct chemistry for scavengingcarbonyls, significantly suppressed the increase in oxylipin carbonyls and blocked PCD in BY-2 cells and Arabidopsis roots,but they did not affect the levels of ROS and lipid peroxides. A transgenic tobacco line that overproduces 2-alkenal reductase, anArabidopsis enzyme to detoxify a,b-unsaturated carbonyls, suffered less PCD in root epidermis after hydrogen peroxide or salttreatment than did the wild type, whereas the ROS level increases due to the stress treatments were not different between thelines. From these results, we conclude that oxylipin carbonyls are involved in the PCD process in oxidatively stressed cells. Ourcomparison of the ability of distinct carbonyls to induce PCD in BY-2 cells revealed that acrolein and 4-hydroxy-(E)-2-nonenalare the most potent carbonyls. The physiological relevance and possible mechanisms of the carbonyl-induced PCD are discussed.
In plants, environmental stressors such as extremetemperatures, drought, intense UV-B radiation, and soilsalinity can cause tissue damage, growth inhibition, andeven death. These detrimental effects are often ascribedto the action of reactive oxygen species (ROS) producedin the stressed plants for the following reasons: (1)various environmental stressors commonly cause theoxidation of biomolecules in plants; and (2) transgenicplants with enhanced antioxidant capacities show im-proved tolerance to environmental stressors (Suzukiet al., 2014). The production of ROS such as superoxideanion radical and hydrogen peroxide (H2O2) is intrin-sically associated with photosynthesis and respiration(Foyer and Noctor, 2003; Asada, 2006).
Plant cells are equipped with abundant antioxidantmolecules such as a-tocopherol, b-carotene, and ascorbicacid and an array of ROS-scavenging enzymes such assuperoxide dismutase and ascorbate peroxidase tomaintain low intracellular ROS levels. When plants areexposed to severe and prolonged environmental stress,the balance between the production and scavenging ofROS is disrupted and the cellular metabolism reaches anew state of higher ROS production and lower anti-oxidant capacity. Then, the oxidation of vital biomol-ecules such as proteins and DNA proceeds, and as aconsequence, cells undergo oxidative injury (Mano,2002). The cause-effect relationship between ROS andtissue injury in plants is thus widely accepted, but thebiochemical processes between the generation of ROSand cell death are poorly understood.
Increasing evidence shows that oxylipin carbonylsmediate the oxidative injury of plants (Yamauchi et al.,2012; for review, see Mano, 2012; Farmer and Mueller,2013). Oxylipin carbonyls are a group of carbonylcompounds derived from oxygenated lipids and fattyacids. The production of oxylipin carbonyls in livingcells is explained as follows. Lipids in the membranesare constitutively oxidized by ROS to form lipid per-oxides (LOOHs; Mène-Saffrané et al., 2007) becausethey are the most immediate and abundant targetsnear the ROS production sites. There are two types ofLOOH formation reaction from ROS (Halliwell and
1 This work was supported by the Japan Society for the Promotionof Science (KAKENHI grant no. 26440149).
2 Present address: Science Research Center, Yamaguchi Univer-sity, Yoshida 1677–1, Yamaguchi 753–8515, Japan.
* Address correspondence to [email protected] author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de-scribed in the Instructions for Authors (www.plantphysiol.org) is:Jun’ichi Mano ([email protected]).
M.S.B. performed most of the experiments; J.M. supervised theexperiments.
[OPEN] Articles can be viewed without a subscription.www.plantphysiol.org/cgi/doi/10.1104/pp.115.256834
Plant Physiology�, July 2015, Vol. 168, pp. 885–898, www.plantphysiol.org � 2015 American Society of Plant Biologists. All Rights Reserved. 885 www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from
Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Gutteridge, 2007). One is the radical-dependent reac-tion. Highly oxidizing radicals, such as hydroxyl rad-ical (standard reduction potential of the HOc/H2Opair, +2.31 V) and the protonated form of superoxideradical (HO2/H2O2, +1.06 V), can abstract a hydrogenatom from a lipid molecule, especially at the centralcarbon of a pentadiene structure in a polyunsaturatedfatty acid, to form a radical. This organic radical rap-idly reacts with molecular oxygen, forming a lipidhydroperoxyl radical, which then abstracts a hydro-gen atom from a neighboring molecule and becomesa LOOH. The other reaction is the addition of singletoxygen to a double bond of an unsaturated fattyacid to form an endoperoxide or a hydroperoxide(both are LOOHs). A variety of LOOH species areformed, depending on the source fatty acid and alsoby the oxygenation mechanism (Montillet et al., 2004).LOOH molecules are unstable, and in the presence ofredox catalysts such as transition metal ions or freeradicals, they decompose to form various aldehydesand ketones (i.e. oxylipin carbonyls; Farmer andMueller, 2013). The chemical species of oxylipin car-bonyl formed in the cells differ according to the fattyacids and the type of ROS involved (Grosch, 1987;Mano et al., 2014a).
More than a dozen species of oxylipin carbonyls areformed in plants (for review, see Mano et al., 2009).Oxylipin carbonyls are constitutively formed in plantsunder normal physiological conditions, and the levelsof certain types of oxylipin carbonyls rise severalfoldunder stress conditions, detected as increases in thefree carbonyl content (Mano et al., 2010; Yin et al.,2010; Kai et al., 2012) and by the extent of the carbonylmodification of target proteins (Winger et al., 2007;Mano et al., 2014b). Among the oxylipin carbonyls,the a,b-unsaturated carbonyls, such as acrolein and4-hydroxy-(E)-2-nonenal (HNE), have high reactivityand cytotoxicity (Esterbauer et al., 1991; Alméras et al.,2003). They strongly inactivate lipoate enzymes inmitochondria (Taylor et al., 2002) and thiol-regulatedenzymes in chloroplasts (Mano et al., 2009) in vitroand cause tissue injury in leaves when they are fumi-gated (Matsui et al., 2012).
The physiological relevance of oxylipin carbonylshas been shown by the observation that the over-expression of different carbonyl-scavenging enzymescommonly confers stress tolerance to transgenic plants(for review, see Mano, 2012). For example, 2-alkenalreductase (AER)-overproducing tobacco (Nicotianatabacum) showed tolerance to aluminum (Yin et al., 2010),aldehyde dehydrogenase-overproducing Arabidopsis(Arabidopsis thaliana) showed tolerance to osmotic andoxidative stress (Sunkar et al., 2003), and aldehydereductase-overproducing tobacco showed tolerance tochemical and drought stress (Oberschall et al., 2000). Inaddition, the genetic suppression of a carbonyl-scavengingenzyme made plants susceptible to stressors (Kotchoniet al., 2006; Shin et al., 2009; Yamauchi et al., 2012; Tanget al., 2014). Under stress conditions, there are positivecorrelations between the levels of certain carbonyls and
the extent of tissue injury (Mano et al., 2010; Yin et al.,2010; Yamauchi et al., 2012). Thus, it is evident thatoxylipin carbonyls, downstream products of ROS, arecauses of oxidative damage in plant cells.
To investigate how oxylipin carbonyls damage cellsin oxidatively stressed plants, we here examined themode of cell death that is induced by oxylipin car-bonyls and identified the carbonyl species responsiblefor the cell death. We observed that oxylipin carbonylscause programmed cell death (PCD), and our resultsdemonstrated that the oxylipin carbonyls mediate theoxidative stress-induced PCD in tobacco Bright Yellow-2(BY-2) cultured cells and in roots of tobacco andArabidopsis plants. We then estimated the relativestrengths of distinct carbonyl species to initiate the PCDprogram. Our findings demonstrate a critical role of thelipid metabolites in ROS signaling.
RESULTS
Oxylipin Carbonyls Formed in H2O2-Stressed Cells CanCause PCD
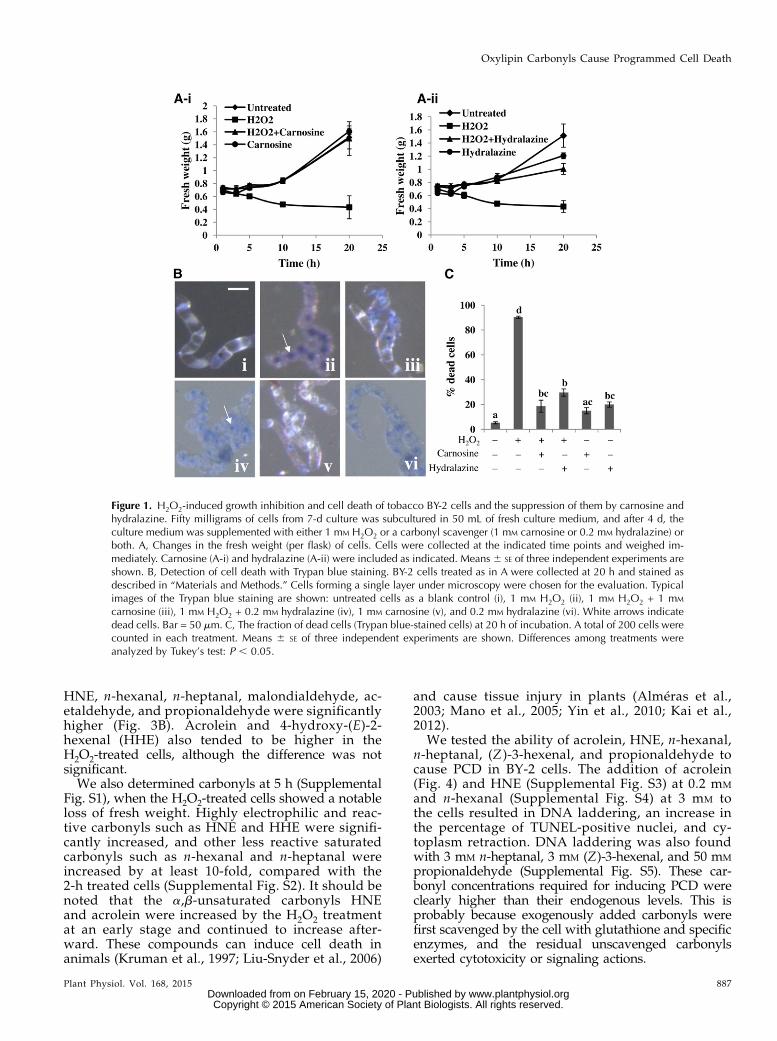
To investigate the process of oxidative injury inplant cells, we first used tobacco BY-2 cells and gavethem an oxidative stimulus with H2O2. Cells that un-derwent a 4-d culture (0.7 g fresh weight per flask)propagated to double weight in approximately 20 hunder the normal culture conditions (Fig. 1A, untreated).When H2O2 was added to 1 mM, the cells stoppedpropagation and their fresh weight started to decrease.At 20 h, the fresh weight was reduced to 0.4 g (Fig. 1A)and the cells were apparently dead, as detected byTrypan blue staining (Fig. 1, B and C).
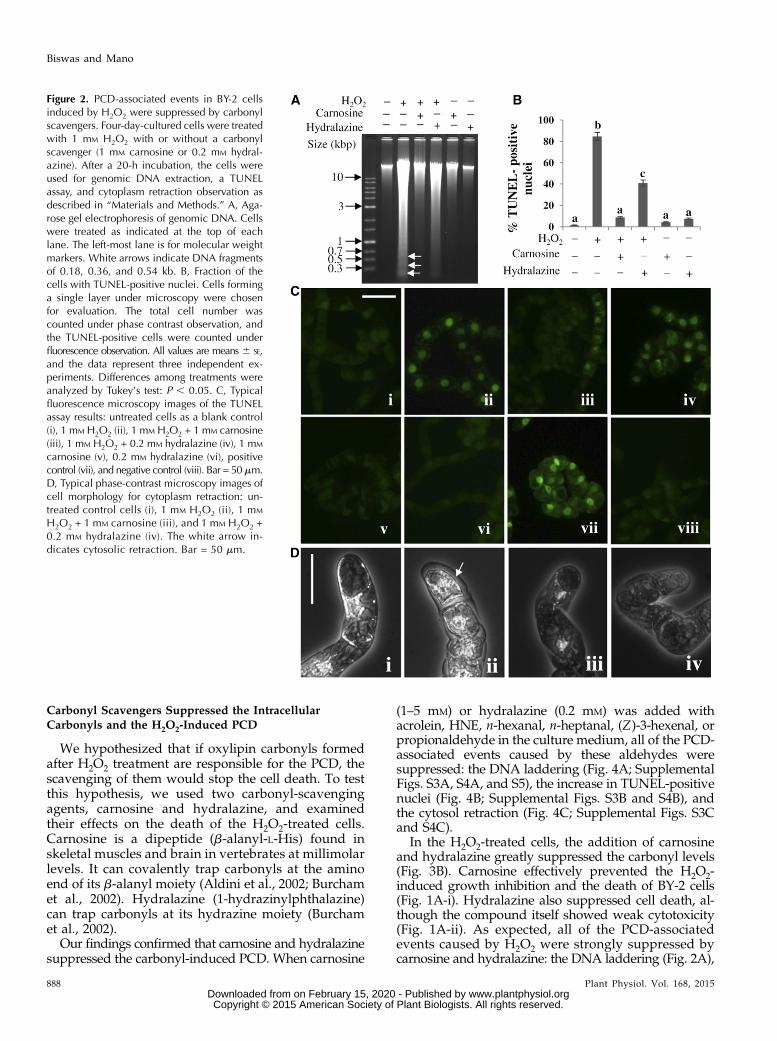
H2O2 induces PCD in BY-2 cells, appearing as mor-phological changes and nuclear DNA fragmentation(Houot et al., 2001). We also detected PCD-associatedevents in the BY-2 cells in the 20-h H2O2 treatmentgroup, as follows. (1) The fragmentation of genomicDNA into 0.18 kb and its multiples was detected asDNA laddering (Fig. 2A). (2) More than 80% of the cellshad positive nuclei in the terminal deoxynucleotidyltransferase dUTP nick end labeling (TUNEL) assay,which represents the fragmentation of DNA, whereasless than 2% of the untreated control cells were positive(Fig. 2B). (3) Retraction of cytosol from the cell wall(Reape and McCabe, 2008) was observed (Fig. 2D).Thus, under our experimental conditions, too, H2O2induced PCD in BY-2 cells.
According to our previous observations of to-bacco leaves and roots (Mano et al., 2010; Yin et al.,2010), oxidative stress treatment will increase thelevels of oxylipin carbonyls before apparent celldeath is observed. At 2 h after treatment, whenthe H2O2-treated cells had just stopped growth(Fig. 1A), we extracted carbonyls from the cells,derivatized them with 2,4-dinitrophenylhydrazine,and analyzed them by HPLC. Eight species of car-bonyls of C1 to C9 were detected in the untreatedcells (Fig. 3A). In the H2O2-treated cells, the levels of
886 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
HNE, n-hexanal, n-heptanal, malondialdehyde, ac-etaldehyde, and propionaldehyde were significantlyhigher (Fig. 3B). Acrolein and 4-hydroxy-(E)-2-hexenal (HHE) also tended to be higher in theH2O2-treated cells, although the difference was notsignificant.We also determined carbonyls at 5 h (Supplemental
Fig. S1), when the H2O2-treated cells showed a notableloss of fresh weight. Highly electrophilic and reac-tive carbonyls such as HNE and HHE were signifi-cantly increased, and other less reactive saturatedcarbonyls such as n-hexanal and n-heptanal wereincreased by at least 10-fold, compared with the2-h treated cells (Supplemental Fig. S2). It should benoted that the a,b-unsaturated carbonyls HNEand acrolein were increased by the H2O2 treatmentat an early stage and continued to increase after-ward. These compounds can induce cell death inanimals (Kruman et al., 1997; Liu-Snyder et al., 2006)
and cause tissue injury in plants (Alméras et al.,2003; Mano et al., 2005; Yin et al., 2010; Kai et al.,2012).
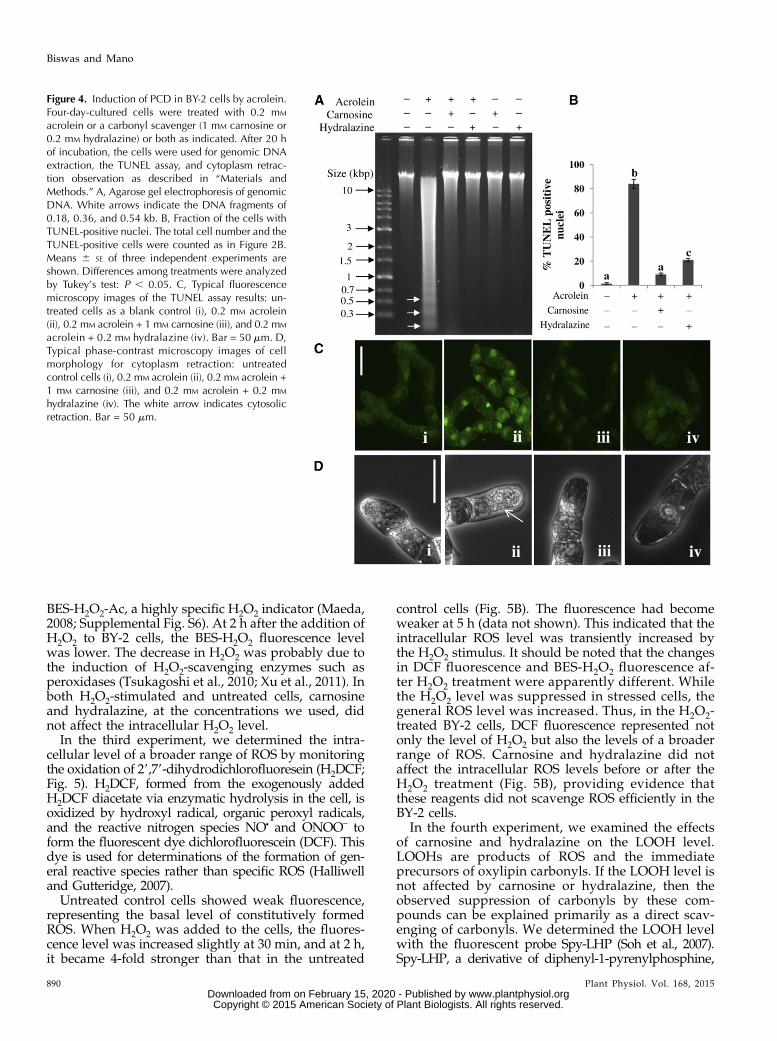
We tested the ability of acrolein, HNE, n-hexanal,n-heptanal, (Z)-3-hexenal, and propionaldehyde tocause PCD in BY-2 cells. The addition of acrolein(Fig. 4) and HNE (Supplemental Fig. S3) at 0.2 mM
and n-hexanal (Supplemental Fig. S4) at 3 mM tothe cells resulted in DNA laddering, an increase inthe percentage of TUNEL-positive nuclei, and cy-toplasm retraction. DNA laddering was also foundwith 3 mM n-heptanal, 3 mM (Z)-3-hexenal, and 50 mM
propionaldehyde (Supplemental Fig. S5). These car-bonyl concentrations required for inducing PCD wereclearly higher than their endogenous levels. This isprobably because exogenously added carbonyls werefirst scavenged by the cell with glutathione and specificenzymes, and the residual unscavenged carbonylsexerted cytotoxicity or signaling actions.
Figure 1. H2O2-induced growth inhibition and cell death of tobacco BY-2 cells and the suppression of them by carnosine andhydralazine. Fifty milligrams of cells from 7-d culture was subcultured in 50 mL of fresh culture medium, and after 4 d, theculture medium was supplemented with either 1 mM H2O2 or a carbonyl scavenger (1 mM carnosine or 0.2 mM hydralazine) orboth. A, Changes in the fresh weight (per flask) of cells. Cells were collected at the indicated time points and weighed im-mediately. Carnosine (A-i) and hydralazine (A-ii) were included as indicated. Means 6 SE of three independent experiments areshown. B, Detection of cell death with Trypan blue staining. BY-2 cells treated as in A were collected at 20 h and stained asdescribed in “Materials and Methods.” Cells forming a single layer under microscopy were chosen for the evaluation. Typicalimages of the Trypan blue staining are shown: untreated cells as a blank control (i), 1 mM H2O2 (ii), 1 mM H2O2 + 1 mM
carnosine (iii), 1 mM H2O2 + 0.2 mM hydralazine (iv), 1 mM carnosine (v), and 0.2 mM hydralazine (vi). White arrows indicatedead cells. Bar = 50 mm. C, The fraction of dead cells (Trypan blue-stained cells) at 20 h of incubation. A total of 200 cells werecounted in each treatment. Means 6 SE of three independent experiments are shown. Differences among treatments wereanalyzed by Tukey’s test: P , 0.05.
Plant Physiol. Vol. 168, 2015 887
Oxylipin Carbonyls Cause Programmed Cell Death
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
Carbonyl Scavengers Suppressed the IntracellularCarbonyls and the H2O2-Induced PCD
We hypothesized that if oxylipin carbonyls formedafter H2O2 treatment are responsible for the PCD, thescavenging of them would stop the cell death. To testthis hypothesis, we used two carbonyl-scavengingagents, carnosine and hydralazine, and examinedtheir effects on the death of the H2O2-treated cells.Carnosine is a dipeptide (b-alanyl-L-His) found inskeletal muscles and brain in vertebrates at millimolarlevels. It can covalently trap carbonyls at the aminoend of its b-alanyl moiety (Aldini et al., 2002; Burchamet al., 2002). Hydralazine (1-hydrazinylphthalazine)can trap carbonyls at its hydrazine moiety (Burchamet al., 2002).
Our findings confirmed that carnosine and hydralazinesuppressed the carbonyl-induced PCD. When carnosine
(1–5 mM) or hydralazine (0.2 mM) was added withacrolein, HNE, n-hexanal, n-heptanal, (Z)-3-hexenal, orpropionaldehyde in the culture medium, all of the PCD-associated events caused by these aldehydes weresuppressed: the DNA laddering (Fig. 4A; SupplementalFigs. S3A, S4A, and S5), the increase in TUNEL-positivenuclei (Fig. 4B; Supplemental Figs. S3B and S4B), andthe cytosol retraction (Fig. 4C; Supplemental Figs. S3Cand S4C).
In the H2O2-treated cells, the addition of carnosineand hydralazine greatly suppressed the carbonyl levels(Fig. 3B). Carnosine effectively prevented the H2O2-induced growth inhibition and the death of BY-2 cells(Fig. 1A-i). Hydralazine also suppressed cell death, al-though the compound itself showed weak cytotoxicity(Fig. 1A-ii). As expected, all of the PCD-associatedevents caused by H2O2 were strongly suppressed bycarnosine and hydralazine: the DNA laddering (Fig. 2A),
Figure 2. PCD-associated events in BY-2 cellsinduced by H2O2 were suppressed by carbonylscavengers. Four-day-cultured cells were treatedwith 1 mM H2O2 with or without a carbonylscavenger (1 mM carnosine or 0.2 mM hydral-azine). After a 20-h incubation, the cells wereused for genomic DNA extraction, a TUNELassay, and cytoplasm retraction observation asdescribed in “Materials and Methods.” A, Aga-rose gel electrophoresis of genomic DNA. Cellswere treated as indicated at the top of eachlane. The left-most lane is for molecular weightmarkers. White arrows indicate DNA fragmentsof 0.18, 0.36, and 0.54 kb. B, Fraction of thecells with TUNEL-positive nuclei. Cells forminga single layer under microscopy were chosenfor evaluation. The total cell number wascounted under phase contrast observation, andthe TUNEL-positive cells were counted underfluorescence observation. All values are means 6 SE,and the data represent three independent ex-periments. Differences among treatments wereanalyzed by Tukey’s test: P , 0.05. C, Typicalfluorescence microscopy images of the TUNELassay results: untreated cells as a blank control(i), 1 mM H2O2 (ii), 1 mM H2O2 + 1 mM carnosine(iii), 1 mM H2O2 + 0.2 mM hydralazine (iv), 1 mM
carnosine (v), 0.2 mM hydralazine (vi), positivecontrol (vii), and negative control (viii). Bar = 50mm.D, Typical phase-contrast microscopy images ofcell morphology for cytoplasm retraction: un-treated control cells (i), 1 mM H2O2 (ii), 1 mM
H2O2 + 1 mM carnosine (iii), and 1 mM H2O2 +0.2 mM hydralazine (iv). The white arrow in-dicates cytosolic retraction. Bar = 50 mm.
888 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

the increase in TUNEL-positive nuclei (Fig. 2B), and thecytosol retraction (Fig. 2D). It appears that hydralazinehad a weaker effect on PCD markers, but this wasprobably due to the low dose of hydralazine used here.Because it showed a toxicity on the growth of the cells(Fig. 1A-ii), we did not add it at a higher concentration.At the concentration we used, hydralazine might havebeen depleted below optimal scavenger levels by thecarbonyls that were continuously produced followingthe addition of H2O2.These pharmacological results suggested that the
H2O2-induced PCD was prevented by the scavengingof oxylipin carbonyls. There is, however, a possibilitythat these scavengers also suppressed ROS, becausecarnosine and hydralazine have been reported to have
antioxidant capacities (Aruoma et al., 1989; Daiberet al., 2005). We investigated the possibility that thesecompounds suppressed ROS and LOOH in the fol-lowing four ways. First, to examine the direct scav-enging of H2O2 by these compounds, we incubatedH2O2 at 5 mM with either carnosine at 5 mM or hy-dralazine at 1 mM in 50 mM phosphate buffer (pH 7.4)and 0.5 mM diethylenetriaminepentaacetic acid at25°C. After a 30-min incubation, we found no signifi-cant decrease in the H2O2 amount as determined by thecatalase-mediated oxygen evolution (data not shown).Thus, these carbonyl scavengers do not significantlyscavenge H2O2 at the concentrations we used.
Second, we examined the effects of these com-pounds on the intracellular H2O2 level by using
Figure 3. Effects of H2O2 and carbonylscavengers on the carbonyl contents in BY-2cells. Four-day-cultured cells were treatedwith water as a control, 1 mM H2O2, and1 mM H2O2 plus 2 mM carnosine or 0.2 mM
hydralazine for 2 h. Carbonyls were ex-tracted from them, derivatized with 2,4-dinitrophenylhydrazine, and separated byHPLC as described in “Materials andMethods.”A, Typical chromatograms showing the car-bonyls in the control (top) and H2O2-treated(bottom) BY-2 cells. The identified aldehydesare labeled at the top of each peak. B, Intra-cellular contents of malondialdehyde, acet-aldehyde, HHE, acrolein, propionaldehyde,HNE, n-hexanal, and n-heptanal. Means6 SE
of three independent experiments are shown.Differences among treatments were ana-lyzed by Tukey’s test: P , 0.05. FW, Freshweight.
Plant Physiol. Vol. 168, 2015 889
Oxylipin Carbonyls Cause Programmed Cell Death
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
BES-H2O2-Ac, a highly specific H2O2 indicator (Maeda,2008; Supplemental Fig. S6). At 2 h after the addition ofH2O2 to BY-2 cells, the BES-H2O2 fluorescence levelwas lower. The decrease in H2O2 was probably due tothe induction of H2O2-scavenging enzymes such asperoxidases (Tsukagoshi et al., 2010; Xu et al., 2011). Inboth H2O2-stimulated and untreated cells, carnosineand hydralazine, at the concentrations we used, didnot affect the intracellular H2O2 level.
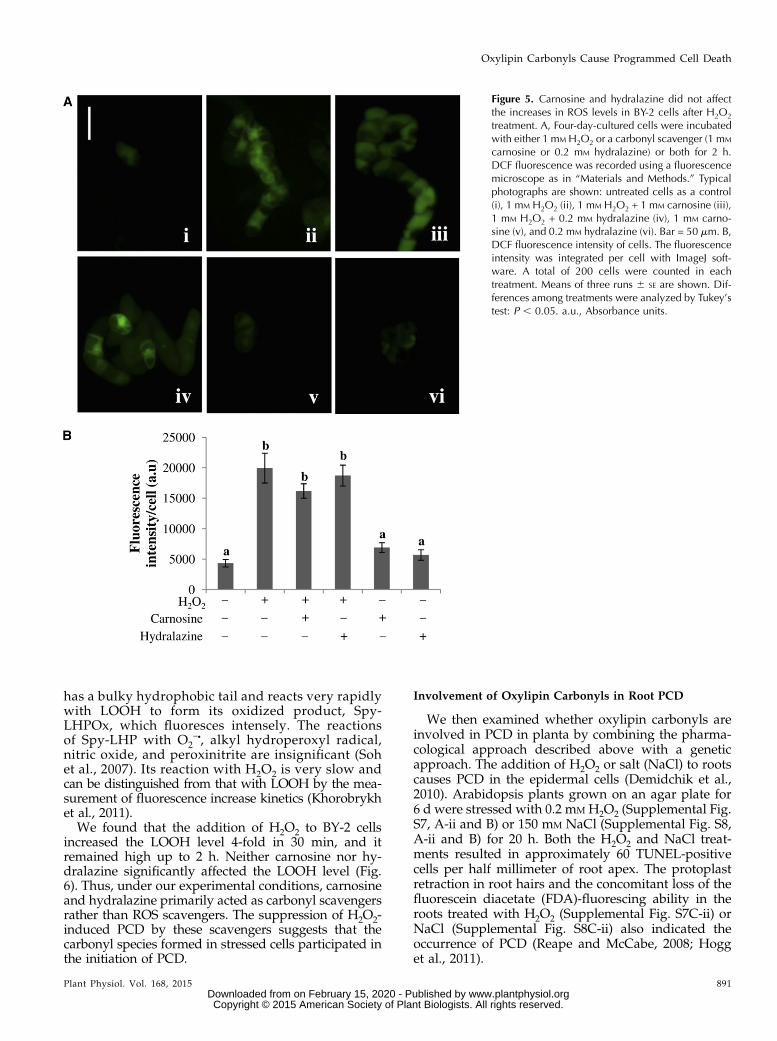
In the third experiment, we determined the intra-cellular level of a broader range of ROS by monitoringthe oxidation of 29,79-dihydrodichlorofluoresein (H2DCF;Fig. 5). H2DCF, formed from the exogenously addedH2DCF diacetate via enzymatic hydrolysis in the cell, isoxidized by hydroxyl radical, organic peroxyl radicals,and the reactive nitrogen species NOc and ONOO– toform the fluorescent dye dichlorofluorescein (DCF). Thisdye is used for determinations of the formation of gen-eral reactive species rather than specific ROS (Halliwelland Gutteridge, 2007).
Untreated control cells showed weak fluorescence,representing the basal level of constitutively formedROS. When H2O2 was added to the cells, the fluores-cence level was increased slightly at 30 min, and at 2 h,it became 4-fold stronger than that in the untreated
control cells (Fig. 5B). The fluorescence had becomeweaker at 5 h (data not shown). This indicated that theintracellular ROS level was transiently increased bythe H2O2 stimulus. It should be noted that the changesin DCF fluorescence and BES-H2O2 fluorescence af-ter H2O2 treatment were apparently different. Whilethe H2O2 level was suppressed in stressed cells, thegeneral ROS level was increased. Thus, in the H2O2-treated BY-2 cells, DCF fluorescence represented notonly the level of H2O2 but also the levels of a broaderrange of ROS. Carnosine and hydralazine did notaffect the intracellular ROS levels before or after theH2O2 treatment (Fig. 5B), providing evidence thatthese reagents did not scavenge ROS efficiently in theBY-2 cells.
In the fourth experiment, we examined the effectsof carnosine and hydralazine on the LOOH level.LOOHs are products of ROS and the immediateprecursors of oxylipin carbonyls. If the LOOH level isnot affected by carnosine or hydralazine, then theobserved suppression of carbonyls by these com-pounds can be explained primarily as a direct scav-enging of carbonyls. We determined the LOOH levelwith the fluorescent probe Spy-LHP (Soh et al., 2007).Spy-LHP, a derivative of diphenyl-1-pyrenylphosphine,
Figure 4. Induction of PCD in BY-2 cells by acrolein.Four-day-cultured cells were treated with 0.2 mM
acrolein or a carbonyl scavenger (1 mM carnosine or0.2 mM hydralazine) or both as indicated. After 20 hof incubation, the cells were used for genomic DNAextraction, the TUNEL assay, and cytoplasm retrac-tion observation as described in “Materials andMethods.” A, Agarose gel electrophoresis of genomicDNA. White arrows indicate the DNA fragments of0.18, 0.36, and 0.54 kb. B, Fraction of the cells withTUNEL-positive nuclei. The total cell number and theTUNEL-positive cells were counted as in Figure 2B.Means 6 SE of three independent experiments areshown. Differences among treatments were analyzedby Tukey’s test: P , 0.05. C, Typical fluorescencemicroscopy images of the TUNEL assay results: un-treated cells as a blank control (i), 0.2 mM acrolein(ii), 0.2 mM acrolein + 1 mM carnosine (iii), and 0.2 mM
acrolein + 0.2 mM hydralazine (iv). Bar = 50 mm. D,Typical phase-contrast microscopy images of cellmorphology for cytoplasm retraction: untreatedcontrol cells (i), 0.2 mM acrolein (ii), 0.2 mM acrolein +1 mM carnosine (iii), and 0.2 mM acrolein + 0.2 mM
hydralazine (iv). The white arrow indicates cytosolicretraction. Bar = 50 mm.
890 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
has a bulky hydrophobic tail and reacts very rapidlywith LOOH to form its oxidized product, Spy-LHPOx, which fluoresces intensely. The reactionsof Spy-LHP with O2
–c, alkyl hydroperoxyl radical,nitric oxide, and peroxinitrite are insignificant (Sohet al., 2007). Its reaction with H2O2 is very slow andcan be distinguished from that with LOOH by the mea-surement of fluorescence increase kinetics (Khorobrykhet al., 2011).We found that the addition of H2O2 to BY-2 cells
increased the LOOH level 4-fold in 30 min, and itremained high up to 2 h. Neither carnosine nor hy-dralazine significantly affected the LOOH level (Fig.6). Thus, under our experimental conditions, carnosineand hydralazine primarily acted as carbonyl scavengersrather than ROS scavengers. The suppression of H2O2-induced PCD by these scavengers suggests that thecarbonyl species formed in stressed cells participated inthe initiation of PCD.
Involvement of Oxylipin Carbonyls in Root PCD
We then examined whether oxylipin carbonyls areinvolved in PCD in planta by combining the pharma-cological approach described above with a geneticapproach. The addition of H2O2 or salt (NaCl) to rootscauses PCD in the epidermal cells (Demidchik et al.,2010). Arabidopsis plants grown on an agar plate for6 d were stressed with 0.2 mM H2O2 (Supplemental Fig.S7, A-ii and B) or 150 mM NaCl (Supplemental Fig. S8,A-ii and B) for 20 h. Both the H2O2 and NaCl treat-ments resulted in approximately 60 TUNEL-positivecells per half millimeter of root apex. The protoplastretraction in root hairs and the concomitant loss of thefluorescein diacetate (FDA)-fluorescing ability in theroots treated with H2O2 (Supplemental Fig. S7C-ii) orNaCl (Supplemental Fig. S8C-ii) also indicated theoccurrence of PCD (Reape and McCabe, 2008; Hogget al., 2011).
Figure 5. Carnosine and hydralazine did not affectthe increases in ROS levels in BY-2 cells after H2O2
treatment. A, Four-day-cultured cells were incubatedwith either 1 mM H2O2 or a carbonyl scavenger (1 mM
carnosine or 0.2 mM hydralazine) or both for 2 h.DCF fluorescence was recorded using a fluorescencemicroscope as in “Materials and Methods.” Typicalphotographs are shown: untreated cells as a control(i), 1 mM H2O2 (ii), 1 mM H2O2 + 1 mM carnosine (iii),1 mM H2O2 + 0.2 mM hydralazine (iv), 1 mM carno-sine (v), and 0.2 mM hydralazine (vi). Bar = 50 mm. B,DCF fluorescence intensity of cells. The fluorescenceintensity was integrated per cell with ImageJ soft-ware. A total of 200 cells were counted in eachtreatment. Means of three runs 6 SE are shown. Dif-ferences among treatments were analyzed by Tukey’stest: P , 0.05. a.u., Absorbance units.
Plant Physiol. Vol. 168, 2015 891
Oxylipin Carbonyls Cause Programmed Cell Death
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
When carnosine (3 mM) or hydralazine (0.2 mM) wassupplemented in the medium, all of the PCD-associatedevents caused by H2O2 and NaCl were largely sup-pressed. Specifically, the number of TUNEL-positivenuclei was decreased (Supplemental Figs. S7 and S8A,iii and iv) and protoplast retraction in root hairs wasprevented (Supplemental Figs. S7 and S8C, iii and iv).
Carbonyl scavengers suppressed oxylipin carbonylsbut did not affect the intracellular levels of H2O2 in theroot tissues. In Arabidopsis plants exposed to 150 mM
NaCl for 20 h, an accumulation of carbonyls along thevascular cylinder was observed as a pink color de-velopment of Schiff’s reagent (Supplemental Fig. S9-ii).When carnosine or hydralazine was added to theplants concomitantly with NaCl, the color developedonly weakly (Supplemental Fig. S9, iii and iv). Thesecarbonyl scavengers also did not affect the BES-H2O2fluorescence level in H2O2-treated root apex (SupplementalFig. S10). Thus, oxylipin carbonyls appeared to be in-volved in in planta PCD as well.
To obtain genetic evidence for the involvement ofoxylipin carbonyls in PCD, we used transgenic tobaccoplants overexpressing Arabidopsis AER (Mano et al.,2005). AER catalyzes the NADPH-dependent reduc-tion of the a,b-unsaturated bond in 2-alkenals such asHNE (Mano et al., 2002). The AER-overexpressing to-bacco plants accumulate smaller amounts of carbonylsupon oxidative stress than the wild-type SR1 (Manoet al., 2010; Yin et al., 2010).
In this study, SR1 and the AER-overexpressing lineP1#18 plants grown on agar plates were exposed toH2O2 or NaCl. Treatment with 2 mM H2O2 and 175 mM
NaCl for 20 h respectively resulted in approximately 50and 40 TUNEL-positive nuclei per 1 mm of root apex inSR1 plants, whereas in the P1#18 plants, both treatmentsproduced only approximately 10 TUNEL-positive nuclei
(Fig. 7B). Most of the root hair cells in the SR1 plantsexposed to H2O2 and NaCl showed protoplast retractionand lost the ability to exhibit FDA fluorescence, whereasthe root hair cells in the P1#18 plants showed muchfewer PCD symptoms (Fig. 8).
The overexpression of AER did not affect the intra-cellular ROS level (Fig. 9). The DCF fluorescence levelwas low under nonstressed conditions, and it was in-creased by treatment with H2O2 and NaCl, especiallyat the elongation zone in the root apex. The SR1 andP1#18 plants showed the same fluorescence level beforethe stress treatments and after the treatments. Specifi-cally, the lowered PCD rate in the AER-overexpressingplants was ascribed to the scavenging of carbonyls byAER. These pharmacological and genetic results indi-cate the direct contribution of oxylipin carbonyls toPCD in the H2O2- and NaCl-stressed roots.
Strength of Each Carbonyl to Cause PCD
The a,b-unsaturated aldehydes such as HNE andacrolein are highly electrophilic and generally highlyreactive, whereas saturated aldehydes such as n-hexanaland propionaldehyde are less reactive. To obtain insights
Figure 7. PCD in roots of the wild-type tobacco line SR1 and the AER-overexpressing line P1#18. Six-day-old plants were transferred to freshMurashige and Skoog (MS) medium supplemented with either 2 mM
H2O2 or 175 mM NaCl. After a 20-h incubation, root tips of about5-mm length were excised from the plant and used for TUNEL assay asdescribed in “Materials and Methods.” A, Typical fluorescence imagesfor TUNEL assay. Bar = 250 mm. B, Fraction of the cells with TUNEL-positive nuclei. TUNEL-positive cells were counted in 1 mm from thetip of the primary root. A total of nine roots were counted in eachtreatment. Differences between the lines were examined by Student’st test. n.s., Not significant.
Figure 6. The carbonyl scavengers carnosine and hydralazine did notsuppress LOOH levels in BY-2 cells. Four-day-cultured cells were in-cubated with either 1 mM H2O2 or a carbonyl scavenger (1 mM car-nosine or 0.2 mM hydralazine) or both for the indicated times. TheLOOH levels were detected as in “Materials and Methods.” Means 6 SE
of the data represent three independent experiments. Differencesamong treatments at the same time point were analyzed by Tukey’stest: P , 0.05. FW, Fresh weight.
892 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.
into the roles of individual carbonyls that are formed instressed plant cells, we evaluated the PCD-inducingstrength of these carbonyls. We defined the strength of acarbonyl species as the lowest concentration that wouldcause DNA laddering in tobacco BY-2 cells (Table I;Supplemental Figs. S4 and S5). As expected, the strongestcarbonyls were acrolein and HNE. These caused theDNA laddering, TUNEL-positive nuclei, and cytoplasmretraction (Fig. 4; Supplemental Fig. S3) at concentrationsas low as 0.2 mM. The next strongest carbonyls weren-hexanal, n-heptanal, and (Z)-3-hexenal (3 mM requiredfor PCD), and the weakest was propionaldehyde (50 mM).
We also tested the PCD-inducing ability of carbonylsin planta and found that acrolein and HNE at 1 mM
and propionaldehyde at 10 mM effectively developedPCD markers in root hairs of Arabidopsis, such asprotoplast retraction and a concomitant loss of theFDA-fluorescing ability (Supplemental Fig. S11). Thus,the PCD-inducing strength of carbonyls differs byspecies, but most species of oxylipin carbonyls thatwere generated in H2O2-stressed cells (Fig. 3) can causePCD.
DISCUSSION
Oxylipin Carbonyls Can Mediate H2O2- andNaCl-Induced PCD
We investigated the roles of oxylipin carbonyls inplant injury under oxidative stress. As experimentalmodels, we used H2O2-induced damage to tobacco BY-2cells and H2O2- and NaCl-induced damage to roots oftobacco and Arabidopsis. In these experiments, wefound that the stress treatment commonly inducedPCD and that the scavenging of endogenous oxylipincarbonyls with chemical scavengers or a specific en-zyme blocked the PCD. We identified the oxylipincarbonyl species that were increased in H2O2-treatedBY-2 cells. These species when added exogenouslycould induce PCD in BY-2 cells and roots. From theseresults, we concluded that oxylipin carbonyls, down-stream products of ROS, significantly contributed tothe initiation of PCD in H2O2- or NaCl-stressed cells.
The involvement of oxylipin carbonyls in plant PCDshown in this study may not be surprising. For animalcells, it is established that reactive oxylipin carbonylssuch as HNE have the potential to cause apoptosis,and their involvement in oxidative signal-inducedPCD has been studied extensively (for review, seeDalleau et al., 2013). For plant cells, the toxicity ofoxylipin carbonyls has been recognized recently, buttheir ability to produce cellular damage has not been
Figure 8. Root hair PCD in the wild-type tobacco line SR1 and theAER-overexpressing line P1#18. Six-day-old plants were treated withH2O2 or NaCl as in Figure 7. After a 20-h incubation, root tips of 5-mmlength were excised and used for FDA staining as described in “Ma-terials and Methods.” Typical phase-contrast images for cytoplasmretraction (top row for each line) and fluorescence images of the samefield (bottom row) are shown. White arrows indicate cytoplasm re-traction. Bars = 100 mm.
Figure 9. Distribution of ROS in roots of the wild-type tobacco lineSR1 and the AER-overexpressing line P1#18. Six-day-old plants weretreated with H2O2 or NaCl as in Figure 7. After a 20-h incubation, roottips of 5-mm length were excised from the primary root. DCF fluo-rescence was recorded as in “Materials and Methods.” Typical imagesof DCF fluorescence are shown. Bar = 250 mm.
Plant Physiol. Vol. 168, 2015 893
Oxylipin Carbonyls Cause Programmed Cell Death
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

investigated. An achievement in this study was thatwe identified the carbonyl species that are generatedand can cause PCD in stressed cells. In addition, ourfindings demonstrated that not only ROS but alsooxylipin carbonyls were necessary for the develop-ment of PCD under H2O2 and NaCl stress. Specifically,these oxylipin carbonyls were the chemical entities thatinitiated plant PCD. Although such critical involve-ment of oxylipin carbonyls may be valid only for cer-tain types of stress, our results provide a new clue tounderstanding the toxicity of ROS in plant cells.
Multiple Species of Oxylipin Carbonyls Appear toContribute to PCD
The oxylipin carbonyls identified in the H2O2-stressedcells included the a,b-unsaturated carbonyls acroleinand HNE, the saturated carbonyls n-hexenal, n-heptanal,and propionaldehyde, and (Z)-3-hexenal. All carbonylscan modify proteins via Schiff-base formation with anamino group. The a,b-unsaturated carbonyls are knownto have higher cytotoxicity than other types of carbonylsbecause they are strong electrophiles and can formMichael adducts with the Cys, His, and Lys residues inaddition to the Schiff base. Indeed, HNE and acroleininduced PCD in BY-2 cells at 0.3 mM. What we did notexpect was that saturated carbonyls such as n-hexanaland n-heptanal also have the ability to initiate PCD. Thestrength order of these carbonyls to induce PCD corre-sponded with the order of chemical reactivity (Table I).
Considering that n-hexenal and n-heptanal weremore greatly increased than acrolein and HNE in theH2O2-stimulated cells, we suggest the possibility thatthese saturated carbonyls also substantially contrib-uted to the induction of PCD in the stressed cells. Theinhibition of PCD by the overexpression of AER doesnot exclude the involvement of saturated carbonyls,because in the AER-overproducing plants, the scav-enging of the a,b-unsaturated carbonyls resulted in thesuppression of the stress-induced increase in saturatedcarbonyls, probably as a secondary effect (Yin et al.,2010). These two types of carbonyls might mediatePCD by different mechanisms (i.e. via different
receptors/sensors [discussed below]). It is necessaryto investigate the effect of distinct carbonyls ondifferent pathways.
In addition, the finding that various carbonyls havethe capacity to induce PCD may explain the diversityand number of carbonyl-scavenging enzymes. Thereare four classes of carbonyl-scavenging enzymes inplants: aldehyde dehydrogenases (Kirch et al., 2001), aldo-keto reductases (Oberschall et al., 2000), 2-alkenal reduc-tases (Mano et al., 2005), and glutathione S-transferases(Gronwald and Plaisance, 1998). In the Arabidopsisgenome, a dozen isozymes or more are encoded foreach class. These enzymes in different classes havedifferent substrate specificities, and even in the sameclass, isozymes show different specificities. There musthave been selective pressure on plants to obtain a largeset of scavenging enzymes to harness oxylipin car-bonyls so that they would not trigger PCD unless it isto be commanded.
Oxylipin Carbonyls as Mediators of Oxidative Signals inEnvironmental Stress Responses
ROS-induced cell death in plants has been demon-strated for various environmental stressors (Mittleret al., 2011; Petrov et al., 2015). In this study, we in-vestigated the cell death induced by H2O2 and NaCl.NaCl treatment activates the plasma membraneNADPH oxidases to facilitate superoxide production,from which H2O2 is formed via catalysis with super-oxide dismutase. NaCl treatment also activates poly-amine oxidase, which catalyzes the production of H2O2in the oxidation of spermine and spermidine. Thus,NaCl treatment also imposes H2O2 stress on plant cells(Petrov et al., 2015). When they were stressed, bothcultured cells and root cells showed TUNEL-positivenuclei, cytosolic retraction, and DNA laddering, whichare typical of apoptosis-like PCD (Reape and McCabe,2008).
Two oxidative signal pathways for PCD are reported:one is the mitochondria-dependent activation of caspase-like proteases, as in animal cell apoptosis (Reape andMcCabe, 2008), and the other is the mitogen-activated
Table I. Comparison of the strength of carbonyls to induce PCD in BY-2 tobacco cells as determined byDNA fragmentation
Cells were incubated with a carbonyl at the indicated concentrations for 20 h. Fragmentation of thechromosomal DNA was judged based on the generation of 180 bp and its multiples by agarose gelelectrophoresis. + and –, Detectable and undetectable levels of the DNA fragments, respectively. nt, Nottested.
CarbonylConcentration
0.1 mM 0.2 mM 0.5 mM 3 mM 10 mM 50 mM
Acrolein – + + + nt ntHNE – + + nt nt nt(Z)-3-Hexenal – – – + + ntn-Hexanal – – – + + ntn-Heptanal – – – + + ntPropionaldehyde – – – – – +
894 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

protein kinase (MAPK) cascade (Nakagami et al., 2006).In either case, the receptors or sensors for ROS have beenlargely elusive (Gadjev et al., 2008). Our finding that theoxylipin carbonyls caused PCD suggests that thesecompounds participate in the signal pathway, at least inpart, as mediators just downstream of ROS.The PCD-initiating role of oxylipin carbonyls in
whole plants may not be restricted to NaCl stress. Theoverexpression of carbonyl-scavenging enzymes im-proves a plant’s tolerance not only to salinity but alsoto heavy metals, drought (Sunkar et al., 2003), UV-Blight (Hideg et al., 2003), strong light (Mano et al.,2010), and aluminum (Yin et al., 2010). Among thesestressors, UV-B light and aluminum are known tocause PCD. Analysis of the mode of cell death andoxylipin carbonyls will clarify the roles of these com-pounds in PCD induced by the different conditions.From a broader perspective, ROS-triggered PCD has
been observed in a hypersensitive response afterpathogen infection (Torres et al., 2005) and certaindevelopmental events such as lateral root cap shed-ding (Pennell and Lamb, 1997), aleurone layer deathduring the germination of cereal grains (Bethke andJones, 2001), the development of tracheary elements inthe xylem of vascular plants (Kuriyama and Fukuda,2002), leaf senescence (Zapata et al., 2005), and thenucellar cell elimination during endosperm formationin Sechium edule (Lombardi et al., 2010). Because theproduction of oxylipin carbonyls is closely associatedwith the occurrence of ROS, we can expect their in-volvement in these physiological responses. The use ofcarbonyl scavengers such as carnosine or enzymes incombination with the chemical analysis of the involvedcarbonyls will be a good test to judge the broaderphysiological aspects of the operation of the carbonylsignal in plant PCD.
How Do Oxylipin Carbonyls Initiate PCD?
Because of their high reactivity, the a,b-unsaturatedcarbonyls such as HNE and acrolein may exert indis-criminate damage to proteins, but they target onlylimited types of proteins in vivo. Winger et al. (2007)analyzed the HNE modification of mitochondrialproteins in Arabidopsis cells. They identified 16 dis-tinct proteins that were sensitively modified underoxidative stress. In salt-stressed Arabidopsis leaves,even when the tissue started to die, only 17 proteinspecies showed more than a 2-fold increase in HNEmodification (Mano et al., 2014b). Thus, oxylipin car-bonyls have chemical specificity required for ROSsignaling (Møller and Sweetlove, 2010). In other words,specific receptors/sensors of oxylipin carbonyls mayexist. Indeed, the redox-regulated Nonexpresser of PRgene proteins and the TGA transcription factors arecandidates for the carbonyl sensors (Farmer and Mueller,2013) but not for PCD induction.As judged by the morphological change and DNA
fragmentation, the H2O2- and NaCl-induced PCD in
this study was of the apoptosis-like mode, in which amitochondria-dependent activation of proteases (caspase-like proteases) is involved (Reape and McCabe, 2008). Inthe mitochondria-dependent apoptosis in animal cells,several proteins are suggested as HNE targets, such asthe Fas protein and the tumor suppressor/cell cycleregulator protein p53 (Dalleau et al., 2013), but plant cellsdo not have proteins apparently homologous to theseHNE targets. The primary effect of HNE on plant mito-chondria might be the inactivation of sensitive targetenzymes such as pyruvate dehydrogenase complex andGly decarboxylase complex (Taylor et al., 2002). A rapidconsumption of glutathione via Michael adduct forma-tion with the a,b-unsaturated carbonyls (Esterbauer et al.,1991) would also affect the redox status of mitochondria.These changes would facilitate the deterioration of themitochondrial membranes and the formation of pores onthe outer membrane to allow the release of mitochondrialPCD-associated proteins.
On the other hand, in the H2O2-inducible PCD inArabidopsis, a signal transduction mechanism via theregulation of proteasomal degradation is known to beinvolved in the MAPK cascade. Specifically, MEKK1,the MAPK kinase kinase, which is the most upstreamfactor of the H2O2-induced Arabidopsis MAPK3(AtMPK3)- and AtMPK6-dependent PCD pathway, isregulated in a proteasome-dependent manner (Nakagamiet al., 2006). It was reported in a study of mammaliancells that oxylipin carbonyls are involved in proteasome-dependent signal regulation via the carbonyl sensorKelch-like ECH (for erythroid cell-derived protein withCap‘n’Collar [CNC] homology)-associated protein1(Keap1; Higdon et al., 2012). In the relaxed state of thecells, Keap1 binds the transcription factor Nuclear factorE-2-related factor2 (Nrf2), and this binding facilitates theubiquitination and the subsequent proteasome-dependentdegradation of Nrf2, so as to repress the Nrf2-dependentgene expression. HNE formed under oxidative stressmodifies Keap1 on the Cys-151 residue. The modifiedKeap1 no longer binds Nrf2, which then escapes fromubiquitination. Thus, Keap1 receives and transduces thecarbonyl signal via the attenuation of the proteasome-dependent degradation of Nrf2 to induce defense re-sponses (Higdon et al., 2012). It is unclear how H2O2suppresses the proteasome-dependent MEKK1 degrada-tion, but it is likely that MEKK1 degradation is regulatedby a protein that is functionally similar to Keap1. Oxylipincarbonyls may act on such a regulator.
We recently identified 17 species of proteins thatare sensitively modified with HNE in the leaves ofArabidopsis under salt stress (Mano et al., 2014b). Amongthem, several proteins are candidates to determine cellfate, as follows. One is the chloroplastic peptidyl-prolylcis-trans-isomerase (also named cyclophilin20-3), whichplays a key role in Cys biosynthesis. The modification ofits redox-active Cys residues with 12-oxo-phytodienoicacid, a long-chain oxylipin carbonyl, triggers the forma-tion of Cys synthase complex, thereby activating sulfurassimilation and building up cellular redox potential(Park et al., 2013). Modification of these carbonyl-prone
Plant Physiol. Vol. 168, 2015 895
Oxylipin Carbonyls Cause Programmed Cell Death
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Cys residues with short-chain oxylipin carbonyls mighthave different effects on the protein’s function.
Another candidate is Nitrilase1 (Nit1), a highly con-served protein that functions as a tumor suppressor(Sun et al., 2009). In Arabidopsis, changes in the poly-mer structure and the intracellular localization of Nit1are closely associated with the transition from prolifer-ation to differentiation (Dosko�cilová et al., 2013). It isunclear, however, whether the HNE modification ofNit1 causes such changes. We are now investigating theeffects of the carbonyl modification of each target pro-tein on cell fate.
MATERIALS AND METHODS
Culture of Cells and Plants
The suspension of tobacco (Nicotiana tabacum) BY-2 cells was cultured in MSmedium supplemented with Suc (30 g L21), myoinositol (100 mg L21), KH2PO4(200 mg L21), thiamine HCl (0.5 mg L21), and 2,4-dichlorophenoxyacetic acid(0.2 mg L21), pH 5.6. The cells were cultured in darkness at 25°C with con-tinuous rotation at 120 rpm. Every 7 d, 50 mg of cells was transferred to a flaskof 50 mL of fresh medium. Four-day culture cells were used for PCD treat-ment. In this exponential growth phase, cells remain in the maximum mitoticstage (Nagata et al., 2004). Cells were collected by filtration and washed oncewith distilled water for analysis.
Wild-type tobacco ‘Petit Havana’ SR1, the transgenic line P1#18 thatoverexpresses Arabidopsis (Arabidopsis thaliana) AER (Mano et al., 2005), andArabidopsis ecotype Columbia-0 were grown vertically on 1% (w/v) agarplates (one-half-strength MS medium), in sterile conditions, at 23°C in a 14-h-light/10-h-dark cycle (light intensity, 100 mmol m–2 s–1 with white fluorescencelamps).
Trypan Blue Staining for Cell Death
A 0.2-mL aliquot of cell suspension was added to a mixture of 0.3 mL of3% (w/v) Suc and 0.5 mL of 4% (w/v) Trypan blue solution. After 10 min, cellswere washed once with phosphate-buffered saline (PBS), and then 10 mL of thecell suspension was transferred to a microscopic slide, and the dead (Trypanblue-stained) and live (unstained) cells were counted. Cell death is expressedas the percentage of dead cells.
Isolation of Genomic DNA and Gel Electrophoresis
Approximately 200 mg of BY-2 cells was ground with a mortar and pestlethat were chilled with liquid nitrogen. The resulting powder was transferredto a 1.5-mL microtube, and 0.7 mL of extraction buffer containing 2% (w/v)cetyltrimethylammonium bromide, 1.4 M NaCl, 20 mM EDTA, 100 mM Tris-HCl, pH 8, and 0.2% (v/v) b-mercaptoethanol was added immediately. Themixture was incubated at 65°C for 45 min. After centrifugation at 12,000g for15 min at room temperature, the supernatant was treated with RNase A(10 mg mL21) at 37°C for 1 h and then mixed well with an equal volume of thechloroform:isoamyl alcohol mixture (24:1). After centrifugation at 10,000g for12 min, the upper aqueous phase was collected. The DNA was precipitatedwith a two-thirds volume of isopropanol, washed in 70% (v/v) ethanol, dried,and resuspended in sterile distilled water. The DNA was then electrophoresedon a 1% (w/v) agarose gel followed by visualization with ethidium bromide.
Detection of Nuclear DNA Fragmentation Using aTUNEL Assay
We used the In Situ Cell Death Detection Kit: Fluorescence (Roche Diag-nostics) to determine cell death, according to the manufacturer’s instructionswith minor modification. Four-day culture cells were treated for 20 h withH2O2 or acrolein plus scavengers and then harvested for staining. The cellswere fixed with 4% (w/v) fresh paraformaldehyde in PBS and labeled by theTUNEL reaction mixture: the terminal deoxynuleotidyl transferase solution
and the label solution described in the kit’s manual. The transferase wasomitted for the negative control. For the positive control, we incubated fixedand permeabilized cells with DNase I to induce DNA strand breaks prior tothe TUNEL reaction. The technique was adjusted for the study of the wholeroot and observation of the epidermal cells. Roots were permeabilized for20 min in a solution containing 0.1% (w/v) Triton-X, 0.1% (w/v) sodiumcitrate, 0.1 mM KCl, and 0.1 mM CaCl2. The stained cells and roots were an-alyzed with a fluorescence microscope (Leica AF6000). The excitation andemission wavelengths were 488 and 515 nm, respectively.
Detection and Quantification of Carbonyls Using HPLC
Freshly harvested cells (0.45 g) were added to 3 mL of acetonitrile con-taining 60 nmol of 2-ethylhexanal (as an internal standard) and 0.005% (w/v)butylhydroxytoluene and then incubated in a screw-capped glass tubeat 60°C for 30 min. Carbonyls in the extract were derivatized with 2,4-dinitrophenylhydrazine, and the dinitrophenylhydrazone derivatives wereextracted and analyzed by HPLC as described (Yin et al., 2010). We iden-tified dinitrophenylhydrazone derivatives of carbonyls by their retentiontimes and determined their contents by a comparison with authentic com-pounds (Matsui et al., 2009).
ROS Detection with H2DCF Diacetate and H2O2 Detectionwith the BES-H2O2-Ac Probe
BY-2 cells, collected with a brief centrifugation followed by a washing withdistilled water, were incubated in 20 mM H2DCF diacetate (Molecular Probes)in PBS at 37°C for 30 min or 50 mM BES-H2O2-Ac (Wako Pure Chemical) for 30min in darkness. The cells were washed twice with PBS. For tobacco andArabidopsis roots, 5 mm of root apex was excised and incubated in 50 mM
H2DCF diacetate in 20 mM potassium phosphate, pH 6, or in 50 mM BES-H2O2-Ac in darkness for 30 min and washed two times with PBS. The fluorescencewas monitored using a microscope with excitation at 488 nm and emission at530 nm for both DCF and BES-H2O2.
Detection of LOOHs with Spy-LHP
BY-2 cells were collected via filtration and washed once with distilledwater. Then, 0.4 g of cells was taken in a tube, and 100 mL of suspensionmedium (50 mM MES-NaOH, pH 6.5, and 35 mM NaCl) and 2.7 mM Spy-LHP(Dojindo Laboratories) in ethanol (900 mL) were added. After a 30-min in-cubation at 37°C, the samples were centrifuged at 12,000g for 2 min. Thesupernatant was collected, and its fluorescence was measured with a spec-trofluorometer (FP 8300; JASCO) with excitation at 524 nm and emission at538 nm. To determine the amount of peroxides, a standard curve was pre-pared with the model LOOH m-chloroperbenzoic acid (Khorobrykh et al.,2011).
Visualization of the Accumulation of Carbonyls in Roots
Root tips (approximately 2 cm from the tip) were excised and stained withSchiff’s reagent (Wako) for 20 min at room temperature, rinsed with a freshlyprepare sulfite solution (0.5% [w/v] K2S2O2 in 0.05 M HCl), and kept in thesulfite solution and observed using a light microscope (Leica LED3000).
Root Hair PCD Assay
Roots were stained with FDA (Wako) for the detection of PCD. Only viableroot hairs are able to cleave FDA to form fluorescein and fluoresce green usinga microscope with excitation at 485 nm. Dead cells do not fluoresce, and theirprotoplasts retract from the cell wall. Roots were stained in a 10 mg mL–1
solution of FDA on microscope slides, and root hairs were immediately ob-served using a fluorescence microscope (Leica AF6000).
Supplemental Data
The following supplemental materials are available.
Supplemental Figure S1. Chromatograms of carbonyls in BY-2 cells (5 hexperiment).
896 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

Supplemental Figure S2. Scavengers’ effects on carbonyl levels in BY-2 cells(5 h experiment).
Supplemental Figure S3. Induction of PCD in BY-2 cells by HNE.
Supplemental Figure S4. Induction of PCD in BY-2 cells by n-hexanal.
Supplemental Figure S5. DNA fragmentation by carbonyl treatments onBY-2 cells.
Supplemental Figure S6. Scavengers’ effects on H2O2 levels in BY-2 cells.
Supplemental Figure S7. H2O2-induced PCD in Arabidopsis roots.
Supplemental Figure S8. NaCl-induced PCD in Arabidopsis roots.
Supplemental Figure S9. Accumulation of carbonyls in Arabidopsis roots.
Supplemental Figure S10. Scavengers’ effects on H2O2 in Arabidopsisroots.
Supplemental Figure S11. Root hair PCD made by carbonyls in Arabidopsis.
ACKNOWLEDGMENTS
We thank Atsushi Sakamoto (Hiroshima University) for providing thetobacco BY-2 cells and Kenji Matsui (Yamaguchi University) for critical read-ing of the article.
Received January 6, 2015; accepted May 24, 2015; published May 29, 2015.
LITERATURE CITED
Aldini G, Carini M, Beretta G, Bradamante S, Facino RM (2002) Carnosineis a quencher of 4-hydroxy-nonenal: through what mechanism of reac-tion? Biochem Biophys Res Commun 298: 699–706
Alméras E, Stolz S, Vollenweider S, Reymond P, Mène-Saffrané L,Farmer EE (2003) Reactive electrophile species activate defense geneexpression in Arabidopsis. Plant J 34: 205–216
Aruoma OI, Laughton MJ, Halliwell B (1989) Carnosine, homocarnosine andanserine: could they act as antioxidants in vivo? Biochem J 264: 863–869
Asada K (2006) Production and scavenging of reactive oxygen species inchloroplasts and their functions. Plant Physiol 141: 391–396
Bethke PC, Jones RL (2001) Cell death of barley aleurone protoplasts ismediated by reactive oxygen species. Plant J 25: 19–29
Burcham PC, Kaminskas LM, Fontaine FR, Petersen DR, Pyke SM (2002)Aldehyde-sequestering drugs: tools for studying protein damage bylipid peroxidation products. Toxicology 181-182: 229–236
Daiber A, Oelze M, Coldewey M, Kaiser K, Huth C, Schildknecht S,Bachschmid M, Nazirisadeh Y, Ullrich V, Mülsch A, et al (2005) Hy-dralazine is a powerful inhibitor of peroxynitrite formation as a possibleexplanation for its beneficial effects on prognosis in patients with con-gestive heart failure. Biochem Biophys Res Commun 338: 1865–1874
Dalleau S, Baradat M, Guéraud F, Huc L (2013) Cell death and diseasesrelated to oxidative stress: 4-hydroxynonenal (HNE) in the balance. CellDeath Differ 20: 1615–1630
Demidchik V, Cuin TA, Svistunenko D, Smith SJ, Miller AJ, Shabala S,Sokolik A, Yurin V (2010) Arabidopsis root K+-efflux conductance ac-tivated by hydroxyl radicals: single-channel properties, genetic basisand involvement in stress-induced cell death. J Cell Sci 123: 1468–1479
Dosko�cilová A, Kohoutová L, Volc J, Kourová H, Benada O, Chumová J,Plíhal O, Petrovská B, Halada P, Bögre L, et al (2013) NITRILASE1regulates the exit from proliferation, genome stability and plant devel-opment. New Phytol 198: 685–698
Esterbauer H, Schaur RJ, Zollner H (1991) Chemistry and biochemistry of4-hydroxynonenal, malonaldehyde and related aldehydes. Free RadicBiol Med 11: 81–128
Farmer EE, Mueller MJ (2013) ROS-mediated lipid peroxidation and RES-activated signaling. Annu Rev Plant Biol 64: 429–450
Foyer CH, Noctor G (2003) Redox sensing and signalling associated withreactive oxygen in chloroplasts, peroxisomes and mitochondria. PlantPhysiol 119: 355–364
Gadjev I, Stone JM, Gechev TS (2008) Programmed cell death in plants:new insights into redox regulation and the role of hydrogen peroxide.Int Rev Cell Mol Biol 270: 87–144
Gronwald JW, Plaisance KL (1998) Isolation and characterization of gluta-thione S-transferase isozymes from sorghum. Plant Physiol 117: 877–892
Grosch W (1987) Reactions of hydroperoxides: products of low molecularweight. In HWS Chan, ed, Autoxidation of Unsaturated Lipids. Aca-demic Press, London, pp 95–139
Halliwell B, Gutteridge JM (2007) Free Radicals in Biology and Medicine,Ed 4. Oxford University Press, Oxford
Hideg É, Nagy T, Oberschall A, Dudits D, Vass I (2003) Detoxificationfunction of aldose/aldehyde reductase during drought and ultraviolet-B(280-320 nm) stresses. Plant Cell Environ 26: 513–522
Higdon A, Diers AR, Oh JY, Landar A, Darley-Usmar VM (2012) Cellsignalling by reactive lipid species: new concepts and molecular mech-anisms. Biochem J 442: 453–464
Hogg BV, Kacprzyk J, Molony EM, O’Reilly C, Gallagher TF, Gallois P,McCabe PF (2011) An in vivo root hair assay for determining rates ofapoptotic-like programmed cell death in plants. Plant Methods 7: 45
Houot V, Etienne P, Petitot AS, Barbier S, Blein JP, Suty L (2001) Hy-drogen peroxide induces programmed cell death features in culturedtobacco BY-2 cells, in a dose-dependent manner. J Exp Bot 52: 1721–1730
Kai H, Hirashima K, Matsuda O, Ikegami H, Winkelmann T, Nakahara T,Iba K (2012) Thermotolerant cyclamen with reduced acrolein andmethyl vinyl ketone. J Exp Bot 63: 4143–4150
Khorobrykh SA, Khorobrykh AA, Yanykin DV, Ivanov BN, Klimov VV,Mano J (2011) Photoproduction of catalase-insensitive peroxides on thedonor side of manganese-depleted photosystem II: evidence with aspecific fluorescent probe. Biochemistry 50: 10658–10665
Kirch HH, Nair A, Bartels D (2001) Novel ABA- and dehydration-induciblealdehyde dehydrogenase genes isolated from the resurrection plantCraterostigma plantagineum and Arabidopsis thaliana. Plant J 28: 555–567
Kotchoni SO, Kuhns C, Ditzer A, Kirch HH, Bartels D (2006) Over-expression of different aldehyde dehydrogenase genes in Arabidopsisthaliana confers tolerance to abiotic stress and protects plants againstlipid peroxidation and oxidative stress. Plant Cell Environ 29: 1033–1048
Kruman I, Bruce-Keller AJ, Bredesen D, Waeg G, Mattson MP (1997)Evidence that 4-hydroxynonenal mediates oxidative stress-inducedneuronal apoptosis. J Neurosci 17: 5089–5100
Kuriyama H, Fukuda H (2002) Developmental programmed cell death inplants. Curr Opin Plant Biol 5: 568–573
Liu-Snyder P, Borgens RB, Shi R (2006) Hydralazine rescues PC12 cellsfrom acrolein-mediated death. J Neurosci Res 84: 219–227
Lombardi L, Ceccarelli N, Picciarelli P, Sorce C, Lorenzi R (2010) Nitricoxide and hydrogen peroxide involvement during programmed celldeath of Sechium edule nucellus. Physiol Plant 140: 89–102
Maeda H (2008) Which are you watching, an individual reactive oxygenspecies or total oxidative stress? Ann N Y Acad Sci 1130: 149–156
Mano J (2002) Early events in environmental stresses in plants: inductionmechanisms of oxidative stress. In D Inzé, M Van Montagu, eds, Oxi-dative Stress in Plants. Taylor & Francis, London, pp 217–245
Mano J (2012) Reactive carbonyl species: their production from lipid per-oxides, action in environmental stress, and the detoxification mecha-nism. Plant Physiol Biochem 59: 90–97
Mano J, Belles-Boix E, Babiychuk E, Inzé D, Torii Y, Hiraoka E, TakimotoK, Slooten L, Asada K, Kushnir S (2005) Protection against photooxi-dative injury of tobacco leaves by 2-alkenal reductase: detoxication oflipid peroxide-derived reactive carbonyls. Plant Physiol 139: 1773–1783
Mano J, Khorobrykh S, Matsui K, Iijima Y, Sakurai N, Suzuki H, ShibataD (2014a) Acrolein is formed from trienoic fatty acids in chloroplasts: atargeted metabolomics approach. Plant Biotechnol 31: 535–543
Mano J, Miyatake F, Hiraoka E, Tamoi M (2009) Evaluation of the toxicityof stress-related aldehydes to photosynthesis in chloroplasts. Planta 230:639–648
Mano J, Nagata M, Okamura S, Shiraya T, Mitsui T (2014b) Identifica-tion of oxidatively modified proteins in salt-stressed Arabidopsis: acarbonyl-targeted proteomics approach. Plant Cell Physiol 55: 1233–1244
Mano J, Tokushige K, Mizoguchi H, Fujii H, Khorobrykh S (2010) Ac-cumulation of lipid peroxide-derived, toxic a,b-unsaturated aldehyde(E )-2-pentenal, acrolein and (E )-2-hexenal in leaves under photo-inhibitory illumination. Plant Biotechnol 27: 193–197
Mano J, Torii Y, Hayashi S, Takimoto K, Matsui K, Nakamura K, Inzé D,Babiychuk E, Kushnir S, Asada K (2002) The NADPH:quinone oxido-reductase P1-z-crystallin in Arabidopsis catalyzes the a,b-hydrogenation of
Plant Physiol. Vol. 168, 2015 897
Oxylipin Carbonyls Cause Programmed Cell Death
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.

2-alkenals: detoxication of the lipid peroxide-derived reactive aldehydes.Plant Cell Physiol 43: 1445–1455
Matsui K, Sugimoto K, Kakumyan P, Khorobrykh SA, Mano J (2009)Volatile oxylipins and related compounds formed under stress in plants.Methods Mol Biol 580: 17–28
Matsui K, Sugimoto K, Mano J, Ozawa R, Takabayashi J (2012) Differentialmetabolisms of green leaf volatiles in injured and intact parts of a woundedleaf meet distinct ecophysiological requirements. PLoS ONE 7: e36433
Mène-Saffrané L, Davoine C, Stolz S, Majcherczyk P, Farmer EE (2007)Genetic removal of tri-unsaturated fatty acids suppresses developmen-tal and molecular phenotypes of an Arabidopsis tocopherol-deficientmutant: whole-body mapping of malondialdehyde pools in a complexeukaryote. J Biol Chem 282: 35749–35756
Mittler R, Vanderauwera S, Suzuki N, Miller G, Tognetti VB, Vandepoele K,Gollery M, Shulaev V, Van Breusegem F (2011) ROS signaling: the newwave? Trends Plant Sci 16: 300–309
Møller IM, Sweetlove LJ (2010) ROS signalling: specificity is required.Trends Plant Sci 15: 370–374
Montillet JL, Cacas JL, Garnier L, Montané MH, Douki T, Bessoule JJ,Polkowska-Kowalczyk L, Maciejewska U, Agnel JP, Vial A, et al(2004) The upstream oxylipin profile of Arabidopsis thaliana: a tool toscan for oxidative stresses. Plant J 40: 439–451
Nagata T, Sakamoto K, Shimizu T (2004) Tobacco BY-2 cells: the presentand beyond. In Vitro Cell Dev Biol Plant 40: 163–166
Nakagami H, Soukupová H, Schikora A, Zárský V, Hirt H (2006) Amitogen-activated protein kinase kinase kinase mediates reactive oxy-gen species homeostasis in Arabidopsis. J Biol Chem 281: 38697–38704
Oberschall A, Deák M, Török K, Sass L, Vass I, Kovács I, Fehér A, DuditsD, Horváth GV (2000) A novel aldose/aldehyde reductase protectstransgenic plants against lipid peroxidation under chemical and droughtstresses. Plant J 24: 437–446
Park SW, Li W, Viehhauser A, He B, Kim S, Nilsson AK, Andersson MX,Kittle JD, Ambavaram MM, Luan S, et al (2013) Cyclophilin 20-3 relaysa 12-oxo-phytodienoic acid signal during stress responsive regulation ofcellular redox homeostasis. Proc Natl Acad Sci USA 110: 9559–9564
Pennell RI, Lamb C (1997) Programmed cell death in plants. Plant Cell 9:1157–1168
Petrov V, Hille J, Mueller-Roeber B, Gechev TS (2015) ROS-mediatedabiotic stress-induced programmed cell death in plants. Front PlantSci 6: 69
Reape TJ, McCabe PF (2008) Apoptotic-like programmed cell death inplants. New Phytol 180: 13–26
Shin JH, Kim SR, An G (2009) Rice aldehyde dehydrogenase7 is needed forseed maturation and viability. Plant Physiol 149: 905–915
Soh N, Ariyoshi T, Fukaminato T, Nakajima H, Nakano K, Imato T (2007)Swallow-tailed perylene derivative: a new tool for fluorescent imagingof lipid hydroperoxides. Org Biomol Chem 5: 3762–3768
Sun J, Okumura H, Yearsley M, Frankel W, Fong LY, Druck T, Huebner K(2009) Nit1 and Fhit tumor suppressor activities are additive. J Cell Bio-chem 107: 1097–1106
Sunkar R, Bartels D, Kirch HH (2003) Overexpression of a stress-induciblealdehyde dehydrogenase gene from Arabidopsis thaliana in transgenicplants improves stress tolerance. Plant J 35: 452–464
Suzuki N, Rivero RM, Shulaev V, Blumwald E, Mittler R (2014) Abioticand biotic stress combinations. New Phytol 203: 32–43
Tang W, Sun J, Liu J, Liu F, Yan J, Gou X, Lu BR, Liu Y (2014) RNAi-directed downregulation of betaine aldehyde dehydrogenase 1 (OsBADH1)results in decreased stress tolerance and increased oxidative markerswithout affecting glycine betaine biosynthesis in rice (Oryza sativa). PlantMol Biol 86: 443–454
Taylor NL, Day DA, Millar AH (2002) Environmental stress causes oxi-dative damage to plant mitochondria leading to inhibition of glycinedecarboxylase. J Biol Chem 277: 42663–42668
Torres MA, Jones JD, Dangl JL (2005) Pathogen-induced, NADPHoxidase-derived reactive oxygen intermediates suppress spread of celldeath in Arabidopsis thaliana. Nat Genet 37: 1130–1134
Tsukagoshi H, Busch W, Benfey PN (2010) Transcriptional regulation ofROS controls transition from proliferation to differentiation in the root.Cell 143: 606–616
Winger AM, Taylor NL, Heazlewood JL, Day DA, Millar AH (2007) Thecytotoxic lipid peroxidation product 4-hydroxy-2-nonenal covalentlymodifies a selective range of proteins linked to respiratory function inplant mitochondria. J Biol Chem 282: 37436–37447
Xu FJ, Jin CW, Liu WJ, Zhang YS, Lin XY (2011) Pretreatment with H2O2
alleviates aluminum-induced oxidative stress in wheat seedlings. J In-tegr Plant Biol 53: 44–53
Yamauchi Y, Hasegawa A, Mizutani M, Sugimoto Y (2012) ChloroplasticNADPH-dependent alkenal/one oxidoreductase contributes to the de-toxification of reactive carbonyls produced under oxidative stress. FEBSLett 586: 1208–1213
Yin L, Mano J, Wang S, Tsuji W, Tanaka K (2010) The involvement of lipidperoxide-derived aldehydes in aluminum toxicity of tobacco roots. PlantPhysiol 152: 1406–1417
Zapata JM, Guéra A, Esteban-Carrasco A, Martín M, Sabater B (2005)Chloroplasts regulate leaf senescence: delayed senescence in transgenicndhF-defective tobacco. Cell Death Differ 12: 1277–1284
898 Plant Physiol. Vol. 168, 2015
Biswas and Mano
www.plantphysiol.orgon February 15, 2020 - Published by Downloaded from Copyright © 2015 American Society of Plant Biologists. All rights reserved.





























