Embed Size (px)
Citation preview

HAL Id: hal-00848692https://hal.archives-ouvertes.fr/hal-00848692
Submitted on 27 Jul 2013
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
LES RÉSEAUX TROPHIQUES MICROBIENS DESMILIEUX AQUATIQUES ET TERRESTRES
Behzad Mostajir, Christian Amblard, Evelyne Buffan-Dubau, Rutger De Wit,Robert Lensi, Télesphore Sime-Ngando
To cite this version:Behzad Mostajir, Christian Amblard, Evelyne Buffan-Dubau, Rutger De Wit, Robert Lensi, et al..LES RÉSEAUX TROPHIQUES MICROBIENS DES MILIEUX AQUATIQUES ET TERRESTRES.J.C. Bertrand et al. Presses Universitaires de Pau et des Pays de l’Adour, pp.28, 2012. <hal-00848692>

13.1. INTRODUCTION
13.1.1. LES ÉCOSYSTÈMES PÉLAGIQUESMARINS ET LACUSTRES
L’importance quantitative et fonctionnelle des micro-organismes dans les réseaux trophiques aquatiques n’asérieusement été considérée que depuis une trentained’années (Pomeroy, 1974). Ceci malgré le fait queLohmann, au début des années 1900 (cité dans Laval-Peuto et al., 1986), reconnaissait déjà l’importance dumicrozooplancton. De la même façon, Vernadskii (1926,cité dans Pomeroy, 1991) se référait fréquemment auxmicro-organismes comme une composante importantedu fonctionnement général des océans, faisantréférence, spécifiquement, aux protozoaires et à leursrôles dans les flux de matière et d’énergie et dans lescycles biogéochimiques. En fait, les interactions entreles organismes étaient appréhendées de façonsimpliste, avant la découverte du rôle fonctionnel desmicro-organismes, notamment hétérotrophes. Ainsi,pour les milieux aquatiques, on considérait la structuredu réseau trophique comme étant constituée de troisniveaux : la production de matière organique par lephytoplancton, sa consommation (broutage) par lezooplancton (métazoaire) et la prédation de celui-ci parles poissons. Dans cette chaîne trophique simple etlinéaire, l’accent était mis, principalement, sur lebroutage. Il avait ainsi été identifié, en milieu aquatique,
la chaîne trophique des brouteurs (“Grazer food chain”)et la chaîne trophique basée sur les détritus (“Detritusfood chain”) (Wetzel et al., 1972).
Cette vision simplifiée du réseau trophiqueaquatique résultait de méthodes d’échantillonnage, deconservation et surtout d’observation, qui étaient bienadaptées aux organismes planctoniques de grandetaille mais qui n’étaient pas appropriées pour lesmicro-organismes les plus petits. Pour des raisonspratiques, les organismes planctoniques ont étéséparés en plusieurs classes de taille (Sieburth et al.,1978) : femto (0,02-0,2 µm), pico (0,2-2 µm), nano (2-20 µm), micro (20-200 µm), méso (0,2-20 mm), macro(2-20 cm) et mega (20-200 cm) (Tableau 13.1.).
Le modèle conceptuel simple du réseau trophiquea donc été progressivement modifié. Tout d’abord, lesobservations recueillies dans de nombreux sitesocéaniques et lacustres ont montré que lesorganismes pico- et nanoplanctoniques (<20 µm)étaient responsables de la plus grande partie de larespiration aérobie de la communauté pélagique(Pomeroy, 1974). Par la suite, l’observation et laquantification du picoplancton auto- et hétérotrophedans les milieux aquatiques sont devenues possiblesgrâce à l’utilisation de la microscopie àépifluorescence (Hobbie et al., 1977) et,accessoirement, de la microscopie électronique (cf.Chapitre 17.2.1). Le picoplancton phototrophe s’est
1
LES RÉSEAUX TROPHIQUES MICROBIENSDES MILIEUX AQUATIQUES ET TERRESTRES
BEHZAD MOSTAJIR*, CHRISTIAN AMBLARD*, EVELYNE BUFFAN-DUBAU,RUTGER DEWIT, ROBERT LENSI & TÉLESPHORE SIME-NGANDO
Chapitre 13

avéré d’une importance majeure, notamment dans lesmilieux oligotrophes où plus de 50 % de la productionprimaire totale est due à des organismes de taille < 3µm (Li et al., 1983). Ces découvertes concernant lerôle majeur du pico- et du nanoplancton dans lefonctionnement des écosystèmes aquatiques ontabouti au développement d’une nouvelle conceptionde l’écologie microbienne aquatique, dont un deschamps d’étude est le réseau trophique microbien.
Les travaux conduits en milieu aquatique lacustre,au cours des trois dernières décennies, ont, de lamême façon, permis de montrer que les flux dematière et d’énergie ne s’organisent pas seulementselon la voie trophique linéaire classique basée surl’assimilation photosynthétique (Phytoplancton −>Zooplancton −> Poissons) mais empruntent aussi lavoie du réseau microbien formé des micro-organismesde très petite taille (pico- et nanoplancton) (cf.Carpenter, 1988) pour former un véritable réseautrophique.
La structure du réseau microbien dépend, d’unepart, du type de milieux, et, d’autre part, des forçagesenvironnementaux qui s’exercent sur ces milieux (cf.section 13.4). En milieu aquatique, les domainespélagique et benthique sont largement contrastés etceci aussi bien dans les océans, que dans les lagunesou dans les lacs. Les écosystèmes pélagiques sontsitués entre l’atmosphère et la zone benthique. Lesécosystèmes marins côtiers sont également àl’interface entre le continent et la zone hauturièreocéanique. De ce fait, les écosystèmes pélagiquescôtiers sont des zones tampon, sur le plan verticalentre l’air et la zone benthique et/ou sur le planhorizontal, entre le continent et le large. L’écosystèmepélagique est, par conséquent, très souvent sousl’influence de processus se déroulant dans lesécosystèmes attenants (la lumière, la pluie, le vent, lesapports du bassin versant, la remise en suspensiondes sédiments ou la remontée d’eau profonde, etc.).
13.1.2. LES ÉCOSYSTÈMES BENTHIQUES
Le benthos est défini comme étant l’ensemble descommunautés d’organismes vivant à la surface ou ausein des sédiments des écosystèmes aquatiques. Lesédiment est un support physique constitué de deuxphases, solide sous forme de particules, et liquidesous forme d’eau qui occupe les interstices entre lesparticules minérales et organiques (l’eau interstitielle).Il est à noter que les sols sont constitués de troisphases (solide, liquide et gaz). Les communautésmicrobiennes qui se développent à la surface des
macrophytes (périphyton) et des substrats durs desfonds des écosystèmes aquatiques appartiennentaussi au benthos.
Une grande partie des micro-organismesbenthiques procaryotes et eucaryotes adhère auxsurfaces et est vraiment sessile, tandis que l’autrepartie est composée de micro-organismes mobiles quise déplacent par glissement sur les surfaces ou parnatation dans l’eau interstitielle. Les densités desmicro-organismes procaryotes vivant dans lespremiers mm du sédiment sont souvent 103 à 104 foisplus élevées que celle des bactéries de la colonned’eau. Ainsi, le premier cm du sédiment seral’équivalent d’une colonne d’eau d’une dizaine demètres de hauteur. Le sédiment représente donc unvrai réacteur biogéochimique impliqué dans la réminé-ralisation de la matière organique (MO - cf. Chapitre14.2.4). Dans les systèmes profonds (océan hauturieret grands lacs profonds), cette MO provientessentiellement de la production dans la colonned’eau par sédimentation. Dans les écosystèmesaquatiques moins profonds, elle peut aussi êtreproduite au niveau du benthos par des producteursprimaires comme les macrophytes (phanérogames etmacroalgues), les microalgues et les cyanobactériesou provenir du bassin versant pour les lacs et pour lessystèmes côtiers. Ces systèmes benthiques dominéspar des macrophytes ressemblent beaucoup auxsystèmes terrestres.
Plusieurs espèces de procaryotes et demicroalgues benthiques excrètent de grandesquantités de polymères extracellulaires dénommésEPS selon l’abréviation du terme anglais« Extracellular polymeric substances ». Ces polymèresextracellulaires (EPS) jouent un rôle dans l’adhésiondes micro-organismes aux surfaces, dans la mobilitédes cellules par glissement et représentent égalementun stock important de matière organique.
Les EPS sont généralement très riches en carbone(C) et pauvres en azote (N) et phosphore (P). Ainsi,l’excrétion des EPS peut, aussi, être le résultat d’unecroissance non équilibrée comme, par exemple, dansle cas des carences en N et P.
Ce stock de matière organique, sous forme depolymères, constitue une source de nourriture pour demultiples micro-organismes benthiques et les EPSreprésentent donc un compartiment important dans leschaînes trophiques benthiques. Les EPS sont particu-lièrement importantes dans les biofilms où cespolymères forment une véritable matrice organique quienglobe les micro-organismes. Un cas particulier estconstitué par le biofilm photosynthétique où les
2
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S

microalgues et les cyanobactéries sont lesproducteurs primaires.
13.1.3. LES ÉCOSYSTÈMES TERRESTRES
Comme les milieux aquatiques, les sols jouent unrôle important dans la fourniture de services et biensaux êtres humains. Situés à l’interface entrel’atmosphère, la lithosphère, l’hydrosphère et labiosphère, ces écosystèmes participent aux grandscycles nécessaires à la vie sur Terre : cycle de l’eau etdes nutriments majeurs (carbone, azote, phosphore,etc.). Ils supportent la plupart des systèmes deproduction agricoles, sylvicoles et pastoraux etparticipent à la régulation du climat, au contrôle del’érosion, à la détoxification (Lavelle et al., 2006). Onpeut distinguer quatre niveaux d’organisation desconstituants du sol (Ruellan et Dosso, 1998) :
(i) l’agrégat (de quelques mm à quelques dm) :ensemble cohérent de particules élémentaires ;
(ii) l’assemblage (association d’agrégats) ;
(iii) l’horizon (de quelques cm à quelques mètres) :couches de sol reconnues homogènes sur le planmorphologique ;
(iv) la couverture pédologique :ensemble tridimensionnel d’horizons.
Le poids des processus d’agrégation des niveauxd’organisation 1 et 2 entraîne une distribution spatialetridimensionnelle des facteurs de l’environnement(pression partielle en oxygène, teneurs en MO, étatrédox, etc.) dans les premières couches du sol alorsque, dans les sédiments, la stratification verticale deces facteurs reste prépondérante, à l’exception dessédiments soumis à des processus de bioturbationimportants.
La présence d’une phase gazeuse dans le solgénère évidemment de très importantes différencesd’oxygénation entre la zone inter-agrégats et l’intérieurdes agrégats ainsi que des micro-gradients aérobiose– anaérobiose au sein même des agrégats (Sextoneet al., 1985, Renault et Stengel, 1994). Les modesd’agencement des différents constituants au sein et àla surface d’un agrégat de sol sont complexes.Cependant, l’importance de la spatialisation tridimen-sionnelle des conditions environnementales à micro-et méso-échelles, ainsi que la présence demicrogradients, ne sont pas équivalentes dans tousles sols. Elles jouent un rôle décroissant entre lesstructures fragmentaires (présence d’agrégatsvisibles à l’œil nu), les structures particulaires (danslesquelles les particules élémentaires du sol restent
dissociées) et les structures continues (absenced’agrégats visibles).
Le fonctionnement biologique des sols regroupel’ensemble des fonctions assurées par les organismesvivants qui, en interaction avec les composantesphysiques et chimiques, permettent la dynamique de lamatière organique, le recyclage des nutriments et lacirculation de l’eau. Ces fonctions sont assurées pardes organismes de taille variable comprenant les micro-organismes (archées, bactéries, eucaryotesunicellulaires hétérotrophes) et les invertébrés. De plus,des interactions fortes existent entre ces organismes,interactions dont dépend la qualité des servicesévoqués plus haut. En dépit de cette évidence, la notionde réseau trophique a beaucoup moins imprégné lesétudes sur les sols que les études sur les milieuxaquatiques. L’une des causes les plus importantesréside dans le fait que la production primaire desécosystèmes terrestres n’est pas (ou pratiquement pas)assurée par des micro-organismes mais par lesvégétaux supérieurs. Le fonctionnement de lacomposante biotique souterraine est sous ladépendance de la végétation non seulement parce queles plantes fournissent la quasi-totalité de l’énergie dusystème mais aussi parce que, via leur systèmeracinaire, elles exercent un contrôle important (1) sur lesinteractions bio-physico-chimiques dans les sols et (2)sur la structuration des sols, et peuvent, à ce titre, êtreconsidérées comme d’importants ingénieurs du sol*.
Une autre cause réside dans le fait que le sol estun milieu triphasique (eau, solide, gaz). Contrairementau milieu aquatique, l’équilibre de ces trois phases estsusceptible de changer rapidement en fonction desconditions climatiques, ce qui suppose une forteadaptabilité des acteurs biologiques du sol. A titred’exemple, comme nous le développeronsultérieurement, la présence d’un film d’eau estindispensable à l’activité (notamment prédatrice) desprotozoaires. Ces derniers s’enkystent et deviennentinactifs dès que la teneur en eau s’abaisse en dessousd’un seuil critique (Bardgett, 2005). Enfin, la majoritédes micro-organismes du sol, comme dans d’autresécosystèmes, n’est pas cultivable en utilisant lestechniques classiques de culture et donc difficile àétudier. Certes, le développement récent detechniques moléculaires (indépendantes de la mise enculture) permet d’explorer la diversité réelle desorganismes de l’environnement (voir Bardgett et al.,2005 – cf. Chapitre 17.6), mais l’extraction desorganismes ou de l’ADN de la matrice sol reste unverrou technique difficile à lever.
3
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES

13.2. LES ACTEURS DES RÉSEAUXTROPHIQUES MICROBIENS
13.2.1. LES MICRO-ORGANISMES DES MILIEUX
PÉLAGIQUES MARINS ET LACUSTRES
Au cours de ces dernières années, les progrèsréalisés au niveau des techniques d’identification, dedénombrement et de mesure d’activité métabolique,notamment en microscopie à épifluorescence et enbiologie moléculaire, ont permis d’entrevoir l’extraor-dinaire diversité des micro-organismes, l’étendue deleurs conditions de vie et leurs abondancesjusqu’alors largement sous-estimées, notammentdans les milieux aquatiques. De plus, l’améliorationsensible des différentes méthodes dechromatographie a permis de décrire la compositionbiochimique des communautés et d’aborder lestransferts de matière au sein des réseaux trophiquessous un angle qualitatif.
Dans les écosystèmes pélagiques marins etlacustres, le mode de vie planctonique (flottant) est lemieux adapté. Les micro-organismes planctoniquesprocaryotes ou eucaryotes peuvent êtrehétérotrophes, mixotrophes* ou autotrophes. Cesmicro-organismes assurent la quasi-totalité de laproduction de biomasse et une grande partie desfonctions de l’écosystème pélagique. Plusgénéralement, les systèmes pélagiques marins etd’eau douce sont les seuls écosystèmes dont labiomasse est essentiellement microbienne. Ladifférenciation en classes de taille des micro-organismes, comme mentionnée en introduction, estun moyen pratique pour les étudier et est égalementliée au rôle fonctionnel joué par ces micro-organismes dans les milieux pélagiques. Les acteursmajeurs des réseaux trophiques microbienspélagiques (Tableau 13.1.) sont présentés selon laclassification de Sieburth et al., (1978) :femtoplancton (0,02-0,2 µm), picoplancton (0,2-2 µm), nanoplancton (2-20 µm), microplancton(20-200 µm), et mesoplancton (0,2-20 mm).
Les principales méthodes de dénombrement etd’estimation de biomasse des micro-organismespélagiques sont la cytomètrie en flux et la microscopieà épifluorescence pour le pico- et le nanoplancton, lamicroscopie optique inversée pour le microplancton.La chromatographie liquide (HPLC : High performanceLiquid Chromatography) est également utilisée pourestimer la biomasse du phytoplancton à partir de laquantification des pigments.
Le femtoplancton
En plus des archées et des bactéries de trèspetite taille, le femtoplancton est essentiellementcomposé du virioplancton ou des particules de typeviral (VPL, Virus-Like Particles), particulesomniprésentes en milieu aquatique avec desconcentrations de l’ordre de 104 à 108 ml-1, ce qui enfait les entités biologiques les plus abondantes desmilieux aquatiques (Sime-Ngando, 1997, Sime-Ngando et al., 2003, cf. section 13.4.1).
Le picoplancton
Le picoplancton est composé d’organismesprocaryotes et eucaryotes : des archées, des bactérieshétérotrophes, du picophytoplancton (cyanobactérieset cellules eucaryotes autotrophes) et des picoflagelléshétérotrophes. Plus précisément, le bactérioplanctondes milieux aquatiques est représenté par deuxgrands domaines : les Archaea (ex archeabacteria) etles Bacteria (ex eubacteria). Les Archaea ne sont pasconfinées aux milieux dits « extrêmes » et ellespeuvent représenter jusqu’à 30% des procaryotesdénombrés dans les zones euphotiques oxygénéesdes écosystèmes lacustres (Jardillier et al., 2005) etmarins (Delong, 1992).
4
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
CATÉGORIECLASSESDE TAILLE
VIRUS ET MICRO-ORGANISMES
Femtoplancton 0,02-0,2 µmVirus (la plupart), archaea etbactéries de petite taille
Picoplancton 0,2-2 µm
Virus de grande taille, archées etbactéries (la plupart), cyanobac-téries, eucaryotes de petite tailleauto- et hétérotrophes
Nanoplancton 2-20 µm
Flagellés hétérotrophes, autotro-phes et mixotrophes, ciliés depetite taille, amibes nues de pe-tite taille, zoospores et filamentsde fungi
Microplancton 20-200 µm
Ciliés (la plupart), amibes (la plu-part), autres Sarcodines(Foraminifères, héliozoaires), di-noflagellés, eucaryotes unicellu-laires pigmentés (Desmidiées,Diatomées, etc.)
Mesoplancton 0,2-20 mm Cyanobactéries filamenteuses
Tableau 13.1. Classification selon le critère de taille des viruset micro-organismes planctoniques.

La densité des bactéries hétérotrophes des milieuxaquatiques pélagiques marins et lacustres varie de 105
ml-1 dans les eaux très oligotrophes (pauvre enéléments nutritifs) à 107 ml-1 dans les milieux les pluseutrophes (riche en éléments nutritifs). Généralement,on observe une faible variabilité saisonnière de ladensité des bactéries hétérotrophes avec, cependant,une biomasse sensiblement plus élevée lors dudéveloppement printanier du phytoplancton.
L’abondance des cyanobactéries dans les milieuxmarins côtiers est très variable mais est, généralement,d’environ 104 ml-1 (7-13 x 104 ml-1 en MéditerranéeNord-Ouest) et peut atteindre 17 x 104 ml-1 dans leslagunes méditerranéennes). L’abondance dupicoplancton eucaryote, comme Ostreoccocus tauri,est de l’ordre de 103 ml-1, notamment dans les milieuxmarins côtiers (lagunaires).
Dans les milieux lacustres, l’abondance dupicoplancton autotrophe (picocyanobactéries etpicoplancton autotrophe eucaryote) fluctue entre103ml-1 dans les lacs oligotrophes à 106 ml-1 dans leseaux eutrophes. Cependant, en raison d’un biovolumecellulaire moyen plus élevé que celui des bactérieshétérotrophes, le picoplancton autotrophe constitueune part importante (20 à 50 %) de la biomassepicoplanctonique. Alors que les picocyanobactéries sedéveloppent préférentiellement en période estivale, enrelation avec l’élévation de température, la densité dupicoplancton autotrophe eucaryote est généralementmaximale en période printanière, en lien avec lesfaibles éclairements auxquels cette communauté estparticulièrement bien adaptée d’un point de vuepigmentaire.
Les picoflagellés hétérotrophes sont, avec lesflagellés nanoplanctoniques, les principaux prédateursdu picoplancton dans les milieux marins et lacustresoù leur abondance est d’environ 1 000 cellules ml-1.
Le nanoplancton
Le nanoplancton est constitué de cellulesphytoplanctoniques et de protozooplancton (Planche13-1). Le nanophytoplancton représente, avec lepicophytoplancton, un maillon important des réseauxtrophiques microbiens marins et lacustres.L’abondance du nanophytoplancton est,généralement, inférieure d’un ordre de grandeur àcelle du picophytoplancton.
La principale composante hétérotrophe dunanoplancton est le protozooplancton constituéessentiellement de flagellés et de ciliés et,accessoirement, d’amibes. Les nanoflagelléshétérotrophes forment une communauté abondante et
ubiquiste des milieux marins et lacustres. Ils sontconsidérés comme les principaux prédateurs descellules picoplanctoniques et constituent, ainsi, unélément clé du réseau trophique microbien. C’est ungroupe particulièrement hétérogène à la fois sur unplan morphologique et sur un plan physiologique. Ilssont caractérisés par la présence d’un ou de plusieursflagelles qui jouent un rôle dans le déplacement descellules et la capture de nourriture. Les protistesflagellés peuvent être autotrophes, mixotrophes ouhétérotrophes. Les nanoflagellés hétérotrophes senourrissent par phagotrophie* ou par osmotrophie*.Leur densité en milieu marin est inférieure à 1000cellules ml-1. Ils sont plus abondants dans les milieuxlacustres eutrophes (105 ml-1) que dans les eauxoligotrophes (102 ml-1) avec une taille qui varie,généralement, entre 2 et 30 µm, mais dans les lacsdes zones tempérées la plupart des espèces ont unetaille inférieure à 5 µm.
Le protozooplancton cilié a longtemps été ignorédans les milieux pélagiques en raison de son extrêmesensibilité aux méthodes de prélèvement et de fixationgénéralement employées. Grâce à l’amélioration deces méthodes, plus de 7000 espèces de ciliés, libresou parasites, ont été décrites dans les écosystèmesaquatiques. Il s’agit d’un groupe caractérisé par laprésence d’un cortex cilié qui peut se présenter sousforme de simple cils ou de structures ciliées beaucoupplus complexes. La plupart des ciliés présententégalement un dimorphisme nucléaire puisqu’ilspossèdent un macronoyau et, généralement, plusieursmicronoyaux.
Certains groupes de ciliés, tels que lesOligotriches, les Scuticociliés et les Haptoridés sontparticulièrement bien représentés dans les lacs. Leurtaille varie de 5-10 µm à plus de 200 µm. Leurs modesde nutrition sont très variés ; les ciliés pouvant êtremixotrophes, bactérivores, algivores, détritivores oubien encore omnivores.
Leur abondance dans les milieux marins etlacustres est, en moyenne, de 1 cellule ml-1. Latempérature, l’énergie lumineuse disponible et lesressources nutritives induisent une répartition verticalehétérogène des ciliés dans la colonne d’eau. Leurabondance varie également selon le niveau trophiquedes milieux aquatiques.
Le micro- et le mésoplancton
Le micro-phytoplancton contribue de façonsignificative à la production primaire dans les milieuxpélagiques. Généralement, on estime sa biomasse pardes dénombrements cellulaires en microscopie
5
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES
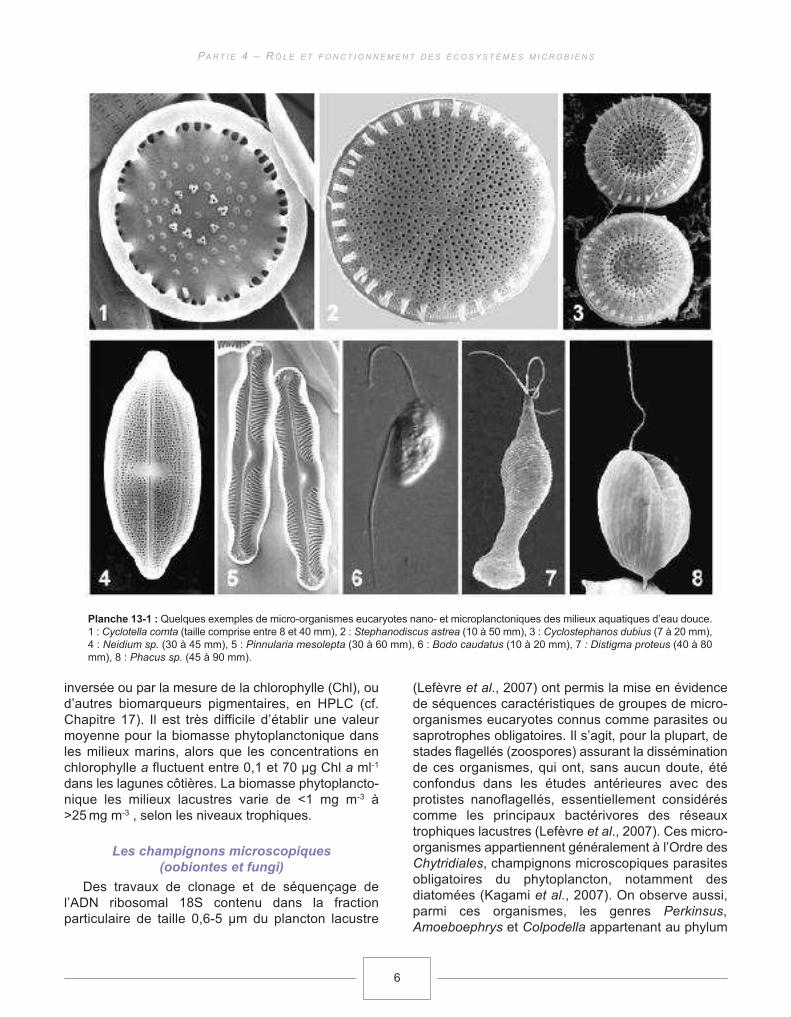
inversée ou par la mesure de la chlorophylle (Chl), oud’autres biomarqueurs pigmentaires, en HPLC (cf.Chapitre 17). Il est très difficile d’établir une valeurmoyenne pour la biomasse phytoplanctonique dansles milieux marins, alors que les concentrations enchlorophylle a fluctuent entre 0,1 et 70 µg Chl a ml-1
dans les lagunes côtières. La biomasse phytoplancto-nique les milieux lacustres varie de <1 mg m-3 à>25mg m-3 , selon les niveaux trophiques.
Les champignons microscopiques
(oobiontes et fungi)
Des travaux de clonage et de séquençage del’ADN ribosomal 18S contenu dans la fractionparticulaire de taille 0,6-5 µm du plancton lacustre
(Lefèvre et al., 2007) ont permis la mise en évidencede séquences caractéristiques de groupes de micro-organismes eucaryotes connus comme parasites ousaprotrophes obligatoires. Il s’agit, pour la plupart, destades flagellés (zoospores) assurant la disséminationde ces organismes, qui ont, sans aucun doute, étéconfondus dans les études antérieures avec desprotistes nanoflagellés, essentiellement considéréscomme les principaux bactérivores des réseauxtrophiques lacustres (Lefèvre et al., 2007). Ces micro-organismes appartiennent généralement à l’Ordre desChytridiales, champignons microscopiques parasitesobligatoires du phytoplancton, notamment desdiatomées (Kagami et al., 2007). On observe aussi,parmi ces organismes, les genres Perkinsus,Amoeboephrys et Colpodella appartenant au phylum
6
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
Planche 13-1 : Quelques exemples de micro-organismes eucaryotes nano- et microplanctoniques des milieux aquatiques d’eau douce.1 : Cyclotella comta (taille comprise entre 8 et 40 mm), 2 : Stephanodiscus astrea (10 à 50 mm), 3 : Cyclostephanos dubius (7 à 20 mm),4 : Neidium sp. (30 à 45 mm), 5 : Pinnularia mesolepta (30 à 60 mm), 6 : Bodo caudatus (10 à 20 mm), 7 : Distigma proteus (40 à 80mm), 8 : Phacus sp. (45 à 90 mm).

des Alvéolobiontes, qui sont parasites de protistes etd’autres organismes aquatiques. Dans le lac Pavin(Auvergne - France), ces micro-organismes parasitesou saprotrophes peuvent représenter plus de 40 % dela diversité génétique totale des organismes flagelléshétérotrophes de petite taille, considérés auparavantcomme bactérivores dans leur totalité. L’élaborationd’outils, notamment moléculaires, favorisant l’obtentionde données quantitatives et fonctionnelles, devraitpermettre de préciser le rôle de ces organismesparasites ou saprotrophes obligatoires dans lefonctionnement des réseaux trophiques aquatiques.D’ores et déjà, ces travaux permettent de proposerune nouvelle vision du fonctionnement des réseauxtrophiques microbiens, notamment lacustres, quiprend en compte des voies de cheminement de lamatière et de l’énergie généralement négligées(parasitisme, saprotrophie).
13.2.2. LES MICRO-ORGANISMES DES MILIEUXBENTHIQUES MARINS ET LACUSTRES
Les microphages benthiques regroupent desorganismes unicellulaires et des métazoaires. Leseucaryotes unicellulaires hétérotrophes constituent desconsommateurs potentiels importants de micro-alguesbenthiques et de bactéries. Les ciliés sont très souventobservés dans les sédiments marins et lacustres et ausein des biofilms où leur densité peut varier entre 87et 1 600 ind. cm². D’autres groupes comme les amibeset les foraminifères sont également présents.
Parmi les métazoaires, la méiofaune comprendune part importante des invertébrés benthiquesmicrophages. Les organismes méiobenthiques(passant à travers une maille de 1 mm et retenus parune maille de 63 µm) sont diversifiés et abondantsdans les sédiments superficiels (106 ind.m² enmoyenne). La microdistribution spatiale agrégative dela méiofaune est associée à celle du microphytoben-thos dans les sédiments superficiels côtiers. Cesinvertébrés sont particulièrement abondants au seindes agrégats microbiens phototrophes qui leurprocurent, à la fois, refuge et nourriture (Mathieu etal., 2007). La méiofaune est généralement dominée,en termes d’abondance et de biomasse, par lesnématodes libres, les copépodes Harpacticoïdes etles rotifères.
Les macro-invertébrés benthiques (de taillesupérieure à celle de la méiofaune) microphages sontprincipalement représentés par des gastéropodes etpar des larves d’insectes (e.g. Chironomidae,Ephemeroptera, Trichoptères).
Dans les sédiments, les techniques d’extractionsont très ciblées pour récolter certains groupes demicro-organismes. Cependant, aucune de cestechniques ne permet d’obtenir l’ensemble des micro-organismes benthiques remis en suspension et de lesséparer entièrement des particules sédimentaires etdétritiques. Les techniques d’extraction ont donc unrecouvrement variable selon les groupes étudiés et,par conséquent, les échantillons de micro-organismesbenthiques obtenus après remise en suspension nesont pas totalement représentatifs de la compositionde la communauté benthique.
Par ailleurs, et contrairement au milieu pélagique,le classement selon la taille est rarement utilisé pourles micro-organismes benthiques. Ces derniers sontclassés à l’aide de critères morphologiques,fonctionnels ou phylogénétiques évalués,respectivement, par des techniques de microscopie,de biogéochimie, ou d’empreintes génétiques.
Les microalgues et les cyanobactéries
Parmi les micro-organismes, les producteursprimaires sont représentés par les microalgues(eucaryotes) et les cyanobactéries (procaryotes,Bacteria). Ce sont des photoautotrophes qui réalisentune photosynthèse oxygénique. Ce compartiment peutêtre étudié à l’aide de l’observation microscopique, carles formes morphologiques sont très caractéristiques,mais aussi à l’aide de l’analyse des pigments photosyn-thétiques par HPLC. La production d’oxygène peut êtrequantifiée à l’aide de micro-electrodes d’oxygène et lafixation de CO2 en utilisant les traceurs isotopiques
14Cou 13C (cf. Chapitre 17). Cependant, ce ne sont pas lesseuls autotrophes au sein des sédiments. Les formesfilamenteuses multicellulaires des cyanobactéries sonttrès communes dans les systèmes benthiques.
Parmi les microalgues, les diatomées et, plusparticulièrement, les formes pennées avec un raphé,sont communes à la surface des sédiments et au seindes biofilms. Les diatomées pennées et la plupart descyanobactéries filamenteuses sont capables de migrerau sein de la communauté benthique par glissement.Ces organismes produisent également de grandesquantités de polymères extracellulaires (EPS). CesEPS peuvent apparaître sous différentes formes :
(i) des structures avec des morphologies caractéris-tiques comme les gaines ;
(ii) des masses muqueuses structurées entourant lescellules ;
(iii) des masses muqueuses dispersées dansl’environnement.
7
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES

Hormis les diatomées, d’autres groupes d’algues sontrégulièrement observés dans les biofilms et à lasurface des sédiments, comme les Chlorophyceae.
Les bactéries phototrophes anoxygéniques
Un autre groupe de micro-organismesphototrophes des milieux benthiques est représentépar les bactéries phototrophes anoxygéniques. Enraison de pigments très caractéristiques (bactério-chlorophylles et caroténoïdes spécifiques), leurprésence peut être révélée à l’aide d’une analysepigmentaire par HPLC, si elles y sont suffisammentabondantes. À la place de l’eau, elles utilisent un autredonneur d’électrons, comme H2S, S2O3
2- ou descomposés organiques et leur source de carbonecellulaire est le CO2 ou des composés organiquesdissous. Bien qu’elles convertissent l’énergielumineuse en biomasse, à la différence des « vrais »producteurs primaires, elles dépendent d’un substratriche en énergie comme donneur d’électrons etpeuvent être considérées comme des producteurs« para-primaires ». Ainsi, l’énergie pour la fixation duCO2 est partiellement fournie, d’une part, par ledonneur d’électrons et, d’autre part, par l’énergielumineuse.
Plusieurs espèces ont une morphologiecaractéristique qui est directement visible à l’aide de lamicroscopie optique comme, par exemple, les espècesde Chromatiaceae (présence de soufre intracellulaire)et les bactéries vertes filamenteuses (qui peuventcependant être facilement confondues avec desespèces de cyanobactéries organisées en filamentsétroits). En revanche, la morphologie d’autres bactériesphototrophes anoxygéniques (Chlorobiaceae,bactéries pourpres non sulfureuses, Ectothiorhodospi-raceae) se confond facilement avec celle des bactérieshétérotrophes chimiolithoautotrophes.
Les bactéries chimiolithotrophes
Les bactéries chimiolithotrophes comprennentdifférents groupes fonctionnels qui fixent le CO2 etdiffèrent selon leur donneur d’électrons (H2, H2S,autres composés soufrés réduits, NH4
+, NO2). Seul unnombre très restreint de bactéries sulfo-oxydantespossède une morphologie suffisamment caractéristiquepour être reconnue à l’aide de la microscopie optique(par exemple les genres Beggiatoa et Thioploca). Ausein de ce groupe fonctionnel on trouve, dans certainssédiments, des représentants de Bacteria « géantes »,visibles à l’œil nu, comme des espèces de Beggiatoa,Thioploca et Thiomargarita (Schulz etJørgensen, 2001, cf. Chapitre 5).
Les bactéries hétérotrophes et les archées
Les bactéries chimio-organohétérotrophes (par lasuite elles seront simplement qualifiéesd’hétérotrophes) utilisent des composés organiques àla fois comme donneur d’électrons et comme sourcede carbone. Elles comprennent un grand nombre degroupes fonctionnels qui peuvent être définis,notamment, selon leur accepteur d’électrons utilisépour la génération d’énergie (O2, NO3
-, composésorganiques, métaux oxydés (Mn4+, Fe3+), SO4
2- et CO2- cf. Chapitre 3). En général, la seule observationmicroscopique de ces procaryotes ne permet pas dedéterminer leur statut fonctionnel.
Au sein des sédiments, le dénombrement desbactéries peut être problématique à cause de laprésence de grandes quantités de matière détritique.Cependant, leur quantification a beaucoup progressédepuis l’introduction des fluorochromes (DAPI) etl’utilisation de la microscopie à épifluorescence (cf.Chapitre 17). Le nombre total de bactéries au seind’un sédiment varie, à la surface, entre 108 et 1010
cellules cm-3. Des bactéries persistent à 500 m deprofondeur dans les sédiments océaniques, desvaleurs allant jusqu’à 105 – 106 cellules cm-3 ont,ainsi, été rapportées. Ces techniques d’épifluores-cence, avec utilisation de fluorochromes, montrentégalement qu’un grand nombre de particules, avecdes densités de l’ordre de 1011 cm-3, ressemblent àdes virus dans les sédiments. Cependant, dans lemême temps, le nombre de bactéries infectées pardes virus paraît très faible et le rôle de ces derniersau sein des sédiments reste encore énigmatique(Filippini et al., 2006).
Comme dans la plupart des écosystèmes, leconcept de groupe fonctionnel est particulièrementpertinent pour l’analyse des activitésbiogéochimiques des communautés benthiques,mais son application peut devenir problématique caril n’y a pas toujours une correspondance entre lesgrands groupes phylogénétiques, issus de l’analysemicroscopique et de l’analyse des empreintesgénétiques selon le gène codant pour le 16S rRNA,et leurs activités biogéochimiques. Cependant,l’analyse des gènes fonctionnels est une méthode dechoix pour établir le lien entre phylogénie et activitébiogéochimique.
L’application du concept de groupe fonctionnel,basé sur les critères biogéochimiques, peut même êtreproblématique au niveau trophique, car les prédateurssélectionnent leurs proies plutôt selon leur taille ouleurs microhabitats que sur leurs différentesfonctionnalités. Le concept fonctionnel a également
8
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
ses limites au niveau individuel des micro-organismes,car plusieurs espèces sont des généralistes versatileset se trouvent, ainsi, au sein de plusieurs groupesfonctionnels. C’est le cas des mixotrophes quicombinent simultanément les voies de l’hétérotrophieet de l’autotrophie.
Pendant longtemps, les archées méthanogènesont été considérées comme les seules Archaeavivant dans la plupart des sédiments anoxiques,tandis que d’autres Archaea étaient considéréescomme typiques des environnements hyperthermo-philes et hypersalés. Bien que les méthanogènesdominent les Archaea de la plupart des sédimentsanoxiques, l’analyse des empreintes génétiquesmontre que d’autres Archaea peuvent exister dansles sédiments.
13.2.3. LES MICRO-ORGANISMESDES MILIEUX TERRESTRES
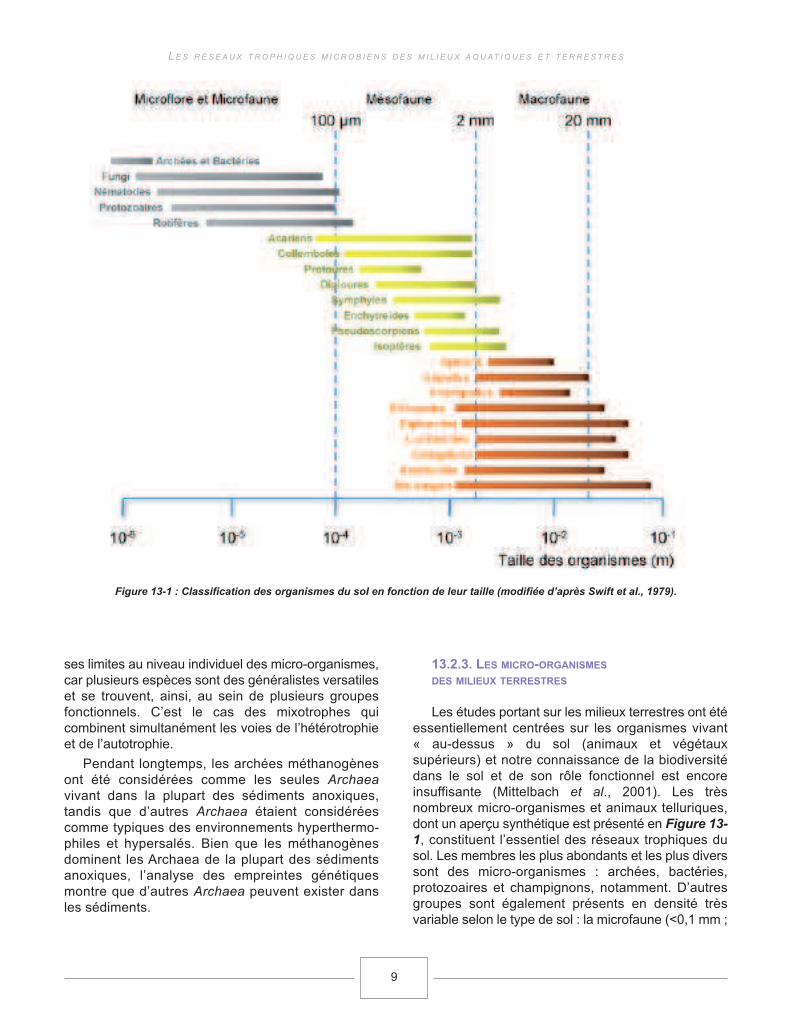
Les études portant sur les milieux terrestres ont étéessentiellement centrées sur les organismes vivant« au-dessus » du sol (animaux et végétauxsupérieurs) et notre connaissance de la biodiversitédans le sol et de son rôle fonctionnel est encoreinsuffisante (Mittelbach et al., 2001). Les trèsnombreux micro-organismes et animaux telluriques,dont un aperçu synthétique est présenté en Figure 13-1, constituent l’essentiel des réseaux trophiques dusol. Les membres les plus abondants et les plus diverssont des micro-organismes : archées, bactéries,protozoaires et champignons, notamment. D’autresgroupes sont également présents en densité trèsvariable selon le type de sol : la microfaune (<0,1 mm ;
9
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES
Figure 13-1 : Classification des organismes du sol en fonction de leur taille (modifiée d’après Swift et al., 1979).

ex : nématodes), la mésofaune (>0,1 mm et <2 mm ;ex : micro-arthropodes et enchitreïdes) et lamacrofaune (>2 mm ; ex : vers de terre, termites etdiplopodes). Les micro-organismes constituent lerégime alimentaire privilégié de nombreux organismesde la microfaune (de Ruiter et al., 1995).
Toutefois, certains auteurs considèrent que nombred’organismes du sol ont un régime omnivore et/ousont capables de se nourrir à partir de différentsniveaux trophiques.
Les modèles concernant les flux de carbone,d’énergie ou de nutriments, à travers le réseautrophique du sol, étant dominés par les lienstrophiques entre les bactéries, les champignons, lesprotozoaires et les nématodes (de Ruiter et al., 1997),notre analyse se limitera à ces organismes dans lasuite de ce paragraphe. Ils peuvent être classés enconsommateurs primaires et secondaires.
Les consommateurs primaires
Les consommateurs primaires jouent un rôleessentiel dans la dégradation des substancesorganiques complexes. Le rôle premier deschampignons (Oobiontes et fungi) est ladécomposition de la matière organique, via leurcapacité à produire une large gamme d’enzymesextracellulaires. Dans un écosystème donné, labiomasse fongique peut largement dépasser celle desautres composantes biotiques (procaryotes, végétaux,animaux). A titre d’exemple, dans un système prairial,les hyphes de champignons peuvent atteindre unebiomasse de 250 kg ha-1 de matière sèche dans lacouche superficielle du sol (5 cm). Cette biomassefongique peut être 10 fois plus importante dans dessols forestiers acides. Outre leur rôle dans ladécomposition de la MO, les champignons peuventêtre pathogènes des plantes, favoriser la structurationdu sol et constituer une ressource abondante pour lafaune consommatrice de micro-organismes. Il estégalement important de citer ici les champignonsmycorhiziens capables d’association mutualiste avecles racines des végétaux : le champignon facilitel’accès par la plante à des éléments souvent limitants(N et P) et, en retour, la plante apporte au champignondes composés carbonés exsudés par les racines, ainsiqu’un espace de moindre compétition (cf. Chapitre 11).
La très grande abondance des bactéries dans lesol est connue depuis longtemps. Plus récemment,les méthodes moléculaires nous ont permisd’appréhender l’immense, et inattendue, diversité dumonde bactérien tellurique (Tableau 13.2). Afin demieux quantifier cette considérable diversité, elle peut
être exprimée sur la base de l’analyse de l’ADNprésent dans le sol. C’est ainsi que Torsvik et al.,(2002) ont estimé que 5 cc3 de sol peuvent contenirjusqu’à 10 000 types de procaryotes génétiquementdistincts. Selon Curtis et al., (2002), il y aurait del’ordre de 4 x 106 taxons procaryotes dans le sol. Àl’instar des champignons, les bactéries jouent un rôleimportant dans la dégradation de la matièreorganique. À l’image de leur diversité taxonomique,leur diversité métabolique est considérable(cf. Chapitre 6).
Les consommateurs secondaires
Parmi les consommateurs secondaires, lesgroupes les plus abondants dans le sol sont lesprotozoaires et les nématodes.
Organismes unicellulaires, les protozoairescomprennent trois groupes principaux : les amibes, lesflagellés et les ciliés. Dans les sols, la présence d’unfilm d’eau est nécessaire pour assurer leur locomotionet, par conséquent, conditionne leur aptitude à senourrir. Leur activité est donc limitée à la porositéoccupée par l’eau (water-filled pore space). Enabsence d’une teneur en eau suffisante, ils arrêtentleur activité, mais sont aptes à former des structuresde résistance, les cystes. Leur nombre varie fortementen fonction du type du sol et des contextesclimatiques. On trouve néanmoins communément desabondances de 104 à 106 g-1 de sol dans la couchesuperficielle du sol, mais leur densité ne dépasse pas100 cellules par gramme de sol dans les horizons plusprofonds où la teneur en matière organique et lenombre de proies deviennent fortement limitants. Ilsprésentent également une grande diversité : on estimeque plus de 1000 espèces de ciliés existent sur laplanète au niveau des sols (Foissner, 1997a) etenviron 150 ont été recensées dans un seul et mêmesite (Foissner, 1997b). La plupart des protozoairessont bactérivores mais il existe également desconsommateurs de champignons et des saprophytes.Lorsque les conditions deviennent favorables aux
10
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
HabitatAbondance(cellules cm-3)
Diversité(équivalent génome)
Sols forestiersSols prairiauxSols cultivé
4,8 x 109
1,8 x 107
2,1 x 1010
60003500-8800140-350
Tableau 13.2. Abondance et diversité des procaryotes dans diffé-rents sols (d’après Torsvik et al., 2002).
protozoaires, la prédation qu’ils exercent sur lesprocaryotes se traduit par une diminution de leurnombre mais, généralement, par une augmentationconcomitante de la minéralisation.
Les nématodes sont les organismes pluricellulairesles plus abondants et les plus divers dans le sol. Ilspeuvent atteindre des densités de 107 m-2 (Yeates etal., 1997). Une étude portant sur un sol de forêt auCameroun a révélé la présence de plus de 400espèces. Les nématodes sont également capables decoloniser des milieux à priori hostiles. À titred’exemple, 27 espèces ont été trouvées dans un sitede dunes sableuses dépourvu de végétation enEcosse. Comme pour les protozoaires, la présence defilms d’eau est indispensable à l’activité (de nutrition etde reproduction, notamment) des nématodes quipossèdent également l’aptitude à résister à desconditions défavorables en réduisant fortement leurmétabolisme. On peut les classer en fonction de leurmode de nutrition sur la base de la morphologie despièces buccales. Les espèces les plus fréquemmentretrouvées consomment des plantes, des bactéries,des champignons ou d’autres nématodes et certainessont omnivores.
13.3. LE FONCTIONNEMENTDES RÉSEAUX TROPHIQUESMICROBIENS
13.3.1. LE FONCTIONNEMENT DES RÉSEAUX TROPHIQUESMICROBIENS EN MILIEUX PÉLAGIQUES MARINS
Dans les écosystèmes pélagiques marins, deuxtypes de réseau trophique peuvent être distingués :le réseau trophique herbivore ou classique(Cushing, 1989) et le réseau trophique microbien(Azam et al., 1983).
Le réseau trophique herbivore
Le réseau trophique herbivore (Tableau 13.3.) estdominé par les communautés micro-phytoplancto-niques (notamment les diatomées) qui assurentl’essentiel de la productivité dans certaines régions età certaines périodes (résurgences, floraisonsprintanières, etc.). Ces communautés phytoplancto-niques assimilent préférentiellement les nitrates(provenant des rivières, de la pluie ou de remontéesd’eaux profondes riches en nitrates) et sont doncimpliquées dans la production dite « nouvelle ». Leréseau trophique classique, par sa productivitéimportante, alimente le métazooplancton de grandetaille et, par la suite, les poissons. Le taux desédimentation du micro-phytoplancton et des produitsassociés (pelotes fécales produites par les herbivores)étant élevé, ce type de réseau est responsable d’uneforte exportation de la matière organique en dehors dela couche euphotique.
Le réseau trophique microbien
Le réseau trophique microbien, qui inclut desmicro-organismes auto- et hétérotrophes, est dominépar les communautés pico- et nano-planctoniques(cyanobactéries, bactéries hétérotrophes et flagellés)et est caractéristique des zones oligotrophes(Tableau 13.3.). La production primaire est plutôtsoutenue par des formes d’azote régénéré (NH4
+) et,de ce fait, considérée comme production« régénérée ». Le concept classique, qui a étédéveloppé en océanographie hauturière, selon lequella production primaire « nouvelle » est basée surl’utilisation de NO3
- et la production primaire« régénérée » est basée sur l’utilisation de NH4
+, nes’applique pas forcément dans les environnementspeu profonds comme certaines zones côtières etlagunaires ou la plupart des lacs. Ceci est lié au faitqu’une grande partie de NH4
+ n’est pas directementassociée à la reminéralisation de l’azote organique
11
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES
Tableau 13.3. Caractéristiques générales des deux types de réseaux trophiques en milieu marin.
Réseauxtrophiques
Tailledes organismesdominants
Statut trophiquedu milieu
Type deproductionprimaire
Sels nutritifsdominants
Productivité dusystème
Exportationou sédimentation
Herbivore < 200 µm Eutrophe Nouvelle NO3- Forte Forte
Microbien < 20 µm Oligotrophe Régénérée NH4+ Faible Faible
mais est produite dans le sédiment, à partir de NO3-
par la réduction dissimilatrice de nitrate en ammonium(RDNA), pour être ensuite relarguée dans la colonned’eau peu profonde.
Le taux de sédimentation de la matière organiquedans les zones oligotrophes étant très faible, lescellules ne contribuent donc pas significativement à lamatière exportée en dehors de la zone euphotique etla matière organique est plutôt recyclée sur place parle protozooplancton (e.g. flagellés et ciliés) qui assurele lien essentiel entre les micro-organismes et lesmacro-organismes.
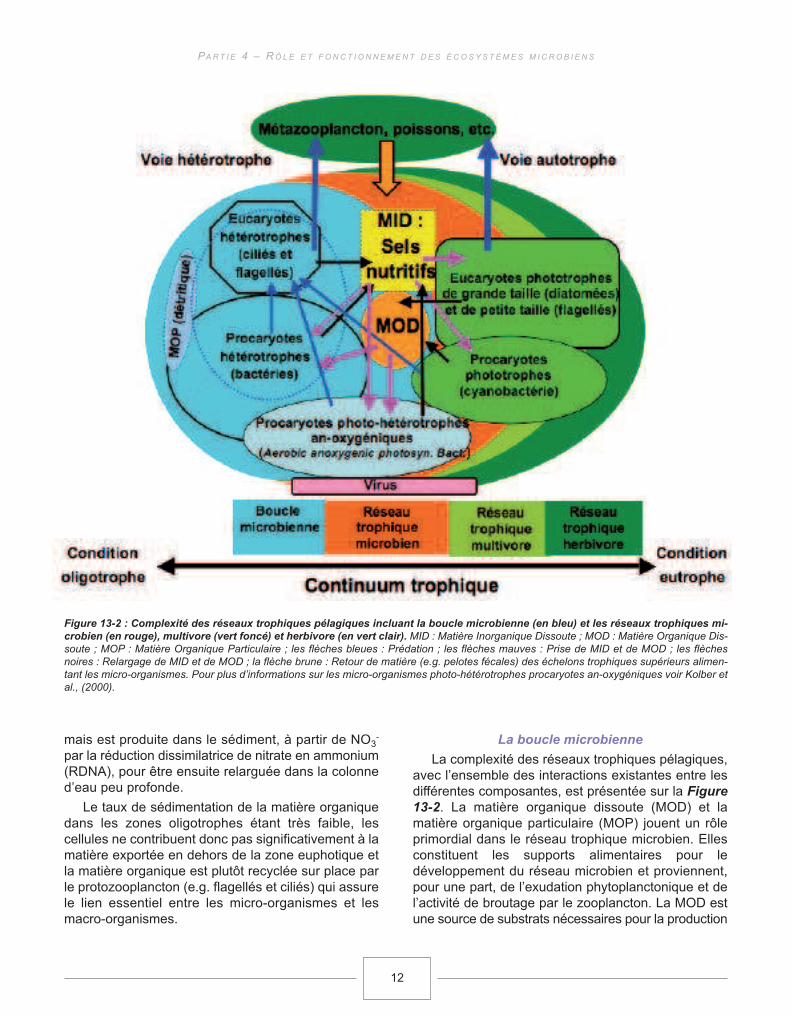
La boucle microbienne
La complexité des réseaux trophiques pélagiques,avec l’ensemble des interactions existantes entre lesdifférentes composantes, est présentée sur la Figure13-2. La matière organique dissoute (MOD) et lamatière organique particulaire (MOP) jouent un rôleprimordial dans le réseau trophique microbien. Ellesconstituent les supports alimentaires pour ledéveloppement du réseau microbien et proviennent,pour une part, de l’exudation phytoplanctonique et del’activité de broutage par le zooplancton. La MOD estune source de substrats nécessaires pour la production
12
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
Figure 13-2 : Complexité des réseaux trophiques pélagiques incluant la boucle microbienne (en bleu) et les réseaux trophiques mi-
crobien (en rouge), multivore (vert foncé) et herbivore (en vert clair). MID : Matière Inorganique Dissoute ; MOD : Matière Organique Dis-soute ; MOP : Matière Organique Particulaire ; les flèches bleues : Prédation ; les flèches mauves : Prise de MID et de MOD ; les flèchesnoires : Relargage de MID et de MOD ; la flèche brune : Retour de matière (e.g. pelotes fécales) des échelons trophiques supérieurs alimen-tant les micro-organismes. Pour plus d’informations sur les micro-organismes photo-hétérotrophes procaryotes an-oxygéniques voir Kolber etal., (2000).

bactérienne. Dans le réseau trophique microbien, lesbactéries hétérotrophes peuvent être en compétitionavec le phytoplancton selon la concentration de lamatière inorganique dissoute (MID), notamment del’azote inorganique dissous (AID) (Fouilland et al.,2007). Lorsque la concentration en AID est faible, lesbactéries hétérotrophes sont en compétition favorableavec le phytoplancton pour l’ammonium. Ceci conduitle système vers la prédominance de la bouclemicrobienne. La boucle microbienne est définie àl’intérieur du réseau trophique microbien comme étantla communauté des organismes hétérotrophes :archées, bactéries, flagellés et ciliés (Azam et al.,1983). La prédation des procaryotes par les flagelléshétérotrophes et par les ciliés, ainsi que celle desflagellés par les ciliés, produit de la matière organiqueet permet le recyclage de différents éléments nutritifs(ammonium, phosphore, etc.) qui sont mobilisés pourla croissance bactérienne.
Le réseau trophique multivore
Il existe toutefois un réseau trophique intermédiaireentre les réseaux trophiques herbivore et microbien, àsavoir le réseau trophique “multivore” (Legendre etRassoulzadegan, 1995). Dans le réseau trophique mul-tivore, le broutage par le métazooplancton (e.g. copé-podes) et le broutage par le protozooplancton (e.g.flagellés et ciliés) ont, tous les deux, un rôle significatif.
Les voies trophiques et le transfert de matière du
système microbien vers les macro-organismes
Les flux de matière résultant des activités de micro-organismes autotrophes et hétérotrophes de différentsréseaux trophiques sont acheminés vers les niveauxtrophiques supérieurs par deux voies trophiquesdistinctes (Figure 13-2).
La voie autotrophe est basée sur l’activité desmicro-organismes phototrophes des réseauxtrophiques herbivore et multivore, alors que la voiehétérotrophe repose sur l’activité des micro-organismes hétérotrophes de la boucle microbienne etdu réseau trophique microbien, notamment des ciliéset flagellés. Le protozooplancton constitue donc unmaillon clé du fonctionnement des réseaux trophiquesen transférant la matière et l’énergie des niveauxtrophiques inférieurs (picoplancton : bactérieshétérotrophes, cyanobactéries et des picoalgueseucaryotes) vers le métazooplancton, les coraux, leshuîtres et les larves de poissons. Dans lesécosystèmes pélagiques marins, l’importance relativedu flux d’éléments (e.g. carbone) acheminés par lavoie hétérotrophe est, en général, inférieure à celle
des éléments transférés par la voie autotrophe. Il estévident qu’une partie de la matière et de l’énergie,nécessaire aux activités métaboliques des micro-organismes, est assurée par le flux de retour dematières issues des activités des organismes desniveaux trophiques supérieurs, telles que les pelotesfécales.
13.3.2. LE FONCTIONNEMENT DES RÉSEAUX TROPHIQUESMICROBIENS EN MILIEUX PÉLAGIQUES LACUSTRES
Les travaux conduits dans des écosystèmeslacustres, au cours des trois dernières décennies, ontpermis, également, de montrer que les flux de matièreet d’énergie ne s’organisent pas seulement selon lavoie trophique linéaire classique basée surl’assimilation photosynthétique (Phytoplancton −>Zooplancton −> Poissons) mais empruntent aussi lavoie de la boucle microbienne, formée des organismeshétérotrophes de petite taille (pico- et nanoplancton),pour constituer un véritable réseau trophique (Figure13-3, Amblard et al., 1998).
Le transfert de matière et d’énergie
vers les niveaux trophiques supérieurs
Il est communément admis qu’une part trèssignificative de la production primaire phytoplanctonique(10 à 50 %) n’est pas consommée directement par lezooplancton métazoaire mais est excrétée dans lemilieu puis transite par la voie de la boucle microbienne.
Le carbone organique dissous (COD) excrété parle phytoplancton, généralement de faible poidsmoléculaire, est utilisé rapidement par les bactérieshétérotrophes qui participent aux transferts de matièreet d’énergie dans les écosystèmes aquatiques parleurs activités de minéralisation et de production debiomasse. Les protistes flagellés hétérotrophes sontconsidérés comme les principaux prédateurs descellules picoplanctoniques en général, et bactériennesen particulier.
Les flagellés mixotrophes*, les ciliés, lescladocères et, à un degré moindre, les rotifèresparticipent également à la prédation sur lescommunautés picoplanctoniques. Les travauxexpérimentaux conduits en conditions contrôlées ontpermis d’observer que le macrozooplancton, àl’exception notoire des cladocères, n’est pas capablede consommer avec efficacité les cellules picoplanc-toniques en raison de leur petite taille et de leurdensité relativement faible. Cette activité de prédation,qui peut être sélective en fonction de la taille, de lamobilité ou de l’activité métabolique des cellules,
13
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES
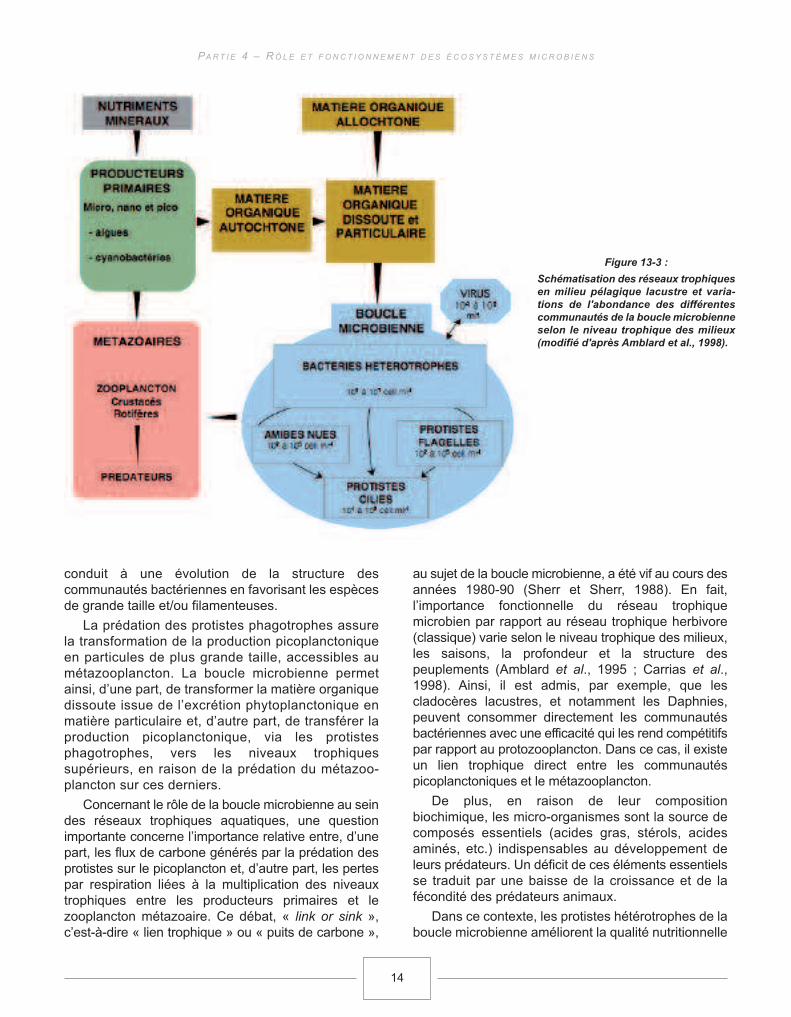
conduit à une évolution de la structure descommunautés bactériennes en favorisant les espècesde grande taille et/ou filamenteuses.
La prédation des protistes phagotrophes assurela transformation de la production picoplanctoniqueen particules de plus grande taille, accessibles aumétazooplancton. La boucle microbienne permetainsi, d’une part, de transformer la matière organiquedissoute issue de l’excrétion phytoplanctonique enmatière particulaire et, d’autre part, de transférer laproduction picoplanctonique, via les protistesphagotrophes, vers les niveaux trophiquessupérieurs, en raison de la prédation du métazoo-plancton sur ces derniers.
Concernant le rôle de la boucle microbienne au seindes réseaux trophiques aquatiques, une questionimportante concerne l’importance relative entre, d’unepart, les flux de carbone générés par la prédation desprotistes sur le picoplancton et, d’autre part, les pertespar respiration liées à la multiplication des niveauxtrophiques entre les producteurs primaires et lezooplancton métazoaire. Ce débat, « link or sink »,c’est-à-dire « lien trophique » ou « puits de carbone »,
au sujet de la boucle microbienne, a été vif au cours desannées 1980-90 (Sherr et Sherr, 1988). En fait,l’importance fonctionnelle du réseau trophiquemicrobien par rapport au réseau trophique herbivore(classique) varie selon le niveau trophique des milieux,les saisons, la profondeur et la structure despeuplements (Amblard et al., 1995 ; Carrias et al.,1998). Ainsi, il est admis, par exemple, que lescladocères lacustres, et notamment les Daphnies,peuvent consommer directement les communautésbactériennes avec une efficacité qui les rend compétitifspar rapport au protozooplancton. Dans ce cas, il existeun lien trophique direct entre les communautéspicoplanctoniques et le métazooplancton.
De plus, en raison de leur compositionbiochimique, les micro-organismes sont la source decomposés essentiels (acides gras, stérols, acidesaminés, etc.) indispensables au développement deleurs prédateurs. Un déficit de ces éléments essentielsse traduit par une baisse de la croissance et de lafécondité des prédateurs animaux.
Dans ce contexte, les protistes hétérotrophes de laboucle microbienne améliorent la qualité nutritionnelle
14
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
Figure 13-3 :
Schématisation des réseaux trophiques
en milieu pélagique lacustre et varia-
tions de l'abondance des différentes
communautés de la boucle microbienne
selon le niveau trophique des milieux
(modifié d'après Amblard et al., 1998).

du picoplancton autotrophe qu’ils ingèrent par leurscapacités de bioconversion, au niveau, par exemple,de la composition en acides gras, selon le concept de« trophic up grading » qui correspond à uneamélioration de la qualité nutritive de la biomasse lelong de la chaîne trophique, (Bec et al., 2006).
Plus généralement, ce processus de « trophic upgrading » semble jouer un rôle majeur dans lefonctionnement des écosystèmes lacustres et leurproductivité.
Le recyclage des éléments nutritifs
Les nutriments inorganiques, principalement l’azoteet le phosphore, sont indispensables à la croissancephytoplanctonique. Bien que l’apport en ces nutrimentspuisse avoir lieu directement par voix aérienne(transport atmosphérique, précipitations), les nutrimentsinorganiques allochtones proviennent majoritairementdu milieu terrestre, par l’intermédiaire d’une rivière oupar le lessivage du bassin versant. La remise ensuspension des sédiments peut également constituerun apport important en nutriments inorganiques.
Les micro-organismes hétérotrophes sontégalement largement impliqués dans les cyclesbiogéochimiques de l’azote et du phosphore, certainscomme assimilateurs et d’autres commerégénérateurs de nutriments inorganiques. Longtempsvu comme seul reminéralisateur de la matièreorganique dans les écosystèmes aquatiquesstagnants, le compartiment bactérien est considérécomme étant d’une importance capitale pour lacroissance phytoplanctonique, notamment en périodede limitation en éléments minéraux. Cependant, lesbactéries peuvent incorporer plus de nutrimentsinorganiques qu’elles n’en produisent. Il a aussi étémis en évidence que les bactéries, du fait de leur fortrapport surface / volume, sont plus compétitives quele phytoplancton vis-à-vis de l’assimilation desnutriments inorganiques, notamment en conditions defaibles concentrations. De ce fait, les bactériesconstituent un pool de phosphore et d’azoteinexploitable par le phytoplancton strictementautotrophe.
Quelques études expérimentales (Dolan, 1997), ontmis en évidence que les flagellés hétérotrophespourraient également jouer un rôle très important dansla reminéralisation de la matière organique. En effet, enexcrétant une partie des nutriments assimilés,principalement sous forme d’ammonium pour l’azote etde phosphates solubles pour le phosphore, les flagelléshétérotrophes contribueraient à la croissancephytoplanctonique de deux manières :
(i) par leur activité bactérivore, ils limiteraient lespopulations bactériennes et donc la compétitionqu’elles représentent pour le phytoplancton vis-à-visdes nutriments ;
(ii) par la production de nutriments inorganiques, ilsstimuleraient la croissance phytoplanctonique.
13.3.3. LE FONCTIONNEMENT DES RÉSEAUXTROPHIQUES MICROBIENS EN MILIEUX BENTHIQUES
MARINS ET LACUSTRES
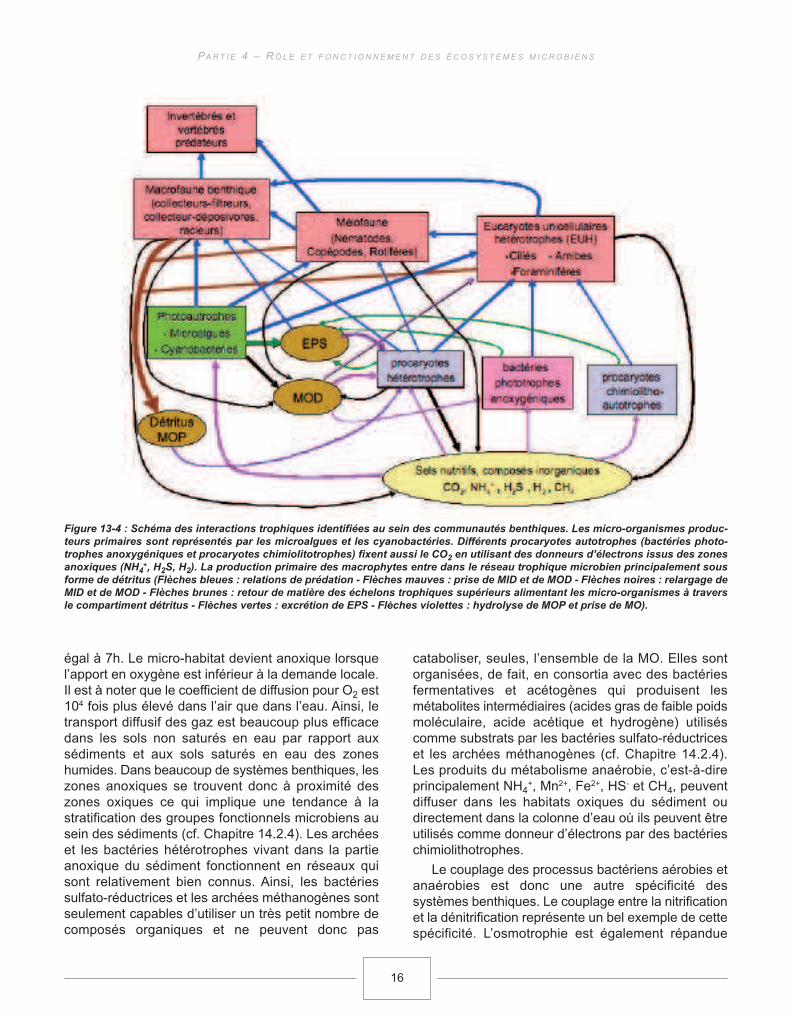
Différentes interactions trophiques ont été identifiéespermettant de conclure que dans les systèmesbenthiques, les transferts de matière et d’énergie nes’organisent pas seulement selon la voie trophiqueclassique (photoautotrophes benthiques→ invertébrésherbivores → invertébrés et vertébrés prédateurs). Eneffet, un grand nombre d’interactions trophiquespotentielles entre les micro-organismes ont étéidentifiées indiquant qu’un réseau trophique microbienexiste au sein des systèmes benthiques (Figure 13-4),même si la quantification de ces transferts trophiquesreste encore problématique (voir ci-après).
La densité élevée des micro-organismes et lesdistances relativement courtes les séparant favorisentun transfert trophique des composés dissous basé surla diffusion moléculaire et l’osmotrophie*. Cetransport diffusif est très efficace à micro-échelle, où ilfavorise l’osmotrophie, mais il devient extrêmementcontraignant à des échelles spatiales supérieures.Pour cette raison, la taille des organismesosmotrophes est généralement limitée. Desexceptions sont observées pour les bactéries chimio-lithotrophes de différents sédiments (Schulz etJørgensen, 2001). Ainsi, Thioploca est capable, parses mouvements, de créer un courant advectif quiaugmente le transport des solutés vers les cellules.D’autres bactéries « géantes » comme certainesespèces de Beggiatoa, et notamment Thiomargaritanamibiensis, sont capables de stocker de grandesquantités de soufre (donneur d’électrons) et de nitrate(accepteur d’électrons) dans leurs vacuoles etdeviennent, ainsi, indépendantes vis-à-vis de lavariabilité dans le temps ou dans l’espace de ladisponibilité de leurs donneurs et accepteursd’électrons (Schulz et Jørgensen, 2001).
Dans les sédiments non perméables comme lesvases, le déplacement de l’eau interstitielle est faibleet le transport des solutés dépend donc exclusivementde la diffusion moléculaire. Le temps pour parcourir 1µm linéaire par diffusion pour l’oxygène est égal à 0,3ms, alors que le temps pour traverser 1 cm linéaire est
15
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES
égal à 7h. Le micro-habitat devient anoxique lorsquel’apport en oxygène est inférieur à la demande locale.Il est à noter que le coefficient de diffusion pour O2 est104 fois plus élevé dans l’air que dans l’eau. Ainsi, letransport diffusif des gaz est beaucoup plus efficacedans les sols non saturés en eau par rapport auxsédiments et aux sols saturés en eau des zoneshumides. Dans beaucoup de systèmes benthiques, leszones anoxiques se trouvent donc à proximité deszones oxiques ce qui implique une tendance à lastratification des groupes fonctionnels microbiens ausein des sédiments (cf. Chapitre 14.2.4). Les archéeset les bactéries hétérotrophes vivant dans la partieanoxique du sédiment fonctionnent en réseaux quisont relativement bien connus. Ainsi, les bactériessulfato-réductrices et les archées méthanogènes sontseulement capables d’utiliser un très petit nombre decomposés organiques et ne peuvent donc pas
cataboliser, seules, l’ensemble de la MO. Elles sontorganisées, de fait, en consortia avec des bactériesfermentatives et acétogènes qui produisent lesmétabolites intermédiaires (acides gras de faible poidsmoléculaire, acide acétique et hydrogène) utiliséscomme substrats par les bactéries sulfato-réductriceset les archées méthanogènes (cf. Chapitre 14.2.4).Les produits du métabolisme anaérobie, c’est-à-direprincipalement NH4
+, Mn2+, Fe2+, HS- et CH4, peuventdiffuser dans les habitats oxiques du sédiment oudirectement dans la colonne d’eau où ils peuvent êtreutilisés comme donneur d’électrons par des bactérieschimiolithotrophes.
Le couplage des processus bactériens aérobies etanaérobies est donc une autre spécificité dessystèmes benthiques. Le couplage entre la nitrificationet la dénitrification représente un bel exemple de cettespécificité. L’osmotrophie est également répandue
16
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
Figure 13-4 : Schéma des interactions trophiques identifiées au sein des communautés benthiques. Les micro-organismes produc-
teurs primaires sont représentés par les microalgues et les cyanobactéries. Différents procaryotes autotrophes (bactéries photo-
trophes anoxygéniques et procaryotes chimiolitotrophes) fixent aussi le CO2 en utilisant des donneurs d’électrons issus des zones
anoxiques (NH4+, H2S, H2). La production primaire des macrophytes entre dans le réseau trophique microbien principalement sous
forme de détritus (Flèches bleues : relations de prédation - Flèches mauves : prise de MID et de MOD - Flèches noires : relargage de
MID et de MOD - Flèches brunes : retour de matière des échelons trophiques supérieurs alimentant les micro-organismes à travers
le compartiment détritus - Flèches vertes : excrétion de EPS - Flèches violettes : hydrolyse de MOP et prise de MO).

parmi les micro-organismes eucaryotes unicellulairesqui bénéficient ainsi des produits d’excrétion et de lysedes bactéries, ainsi que de leurs activités d’hydrolysedes polymères extracellulaires.
La macrofaune peut être classée (Figure 13-4)
selon le mode d’acquisition de la nourriture. Lesracleurs prélèvent les agrégats microbiens en surfaceen les décollant de leurs supports avant de les ingérer(e.g. les gastéropodes). Les déposivores (e.g.Chironomidae) ingèrent les particules superficiellesavec leur microbiota associé. Ces deux groupes senourrissent donc, potentiellement, à la fois desmicroalgues, des cyanobactéries et d’autres bactériesainsi que de leurs EPS associés. Les « collecteurs-filtreurs » (e.g . Simulidae) benthiques accèdent à leurnourriture par filtration de l’eau surnageante. Cegroupe se nourrit donc du plancton et constitue ainsiun lien trophique entre les communautés benthiqueset pélagiques. Cependant, par leur mise ensuspension, les micro-organismes benthiquespeuvent aussi constituer un apport nutritif pour les« collecteurs-filtreurs » benthiques. La méiofaune sedéplace de façon active à travers les interstices dansle sédiment ou au sein du biofilm (Mathieu et al.,2007) et consomme des bactéries et des microalguestelles que les diatomées (Buffan-Dubau et Carman,2000). Les eucaryotes unicellulaires se nourrissentpar phagocytose des microalgues, descyanobactéries et d’autres bactéries, ainsi que parosmotrophie de la MOD.
L’activité des animaux dans les sédiments ne serésume pas uniquement à leurs rôles trophiques ; laméiofaune et la macrofaune ont également un rôlefonctionnel et contribuent, en particulier, au mélange dusédiment, à la formation de terriers au sein dusédiment et à l’irrigation de ces derniers par le maintiend’un courant d’eau. L’ensemble de ces activités estdécrit par le terme de « bioturbation ». Ce processusfavorise les échanges d’oxygène et d’autres composésà l’interface eau-sédiment. Le creusement de terrierspar la macrofaune benthique donne lieu à unestructuration spatiale très complexe du sédiment avecdes zones relativement bien oxygénées comme lesparois des terriers et des zones anoxiques. Cette fauneexerce donc un rôle d’organismes ingénieurs* del’écosystème comme cela a aussi été observé dans lesécosystèmes terrestres.
Dans les sédiments perméables, qui sont composésde sables, de graviers et de galets, le transport del’oxygène dissous et des autres solutés peut êtrefavorisé par les courants d’advection qui percolent àtravers le sédiment sous l’impact de l’hydrodynamisme.
Les détritus jouent un rôle trophique très importantdans les écosystèmes benthiques. Les systèmesbenthiques où se développent des phanérogamespeuvent être comparés aux systèmes terrestres. Lesgrands brouteurs herbivores sont représentés par desvertébrés comme par exemple les anatidés.Cependant, une grande partie de la production demacrophytes entre dans le réseau trophique sous formede détritus. La macrofaune benthique comprend desinvertébrés broyeurs qui consomment ces détritus defaçon partielle et réduisent les particules grossières enparticules fines qui peuvent être colonisées etminéralisées par les micro-organismes hétérotrophes.Ces derniers sont consommés, par la suite, par lesmicro-organismes eucaryotes et les invertébrés.
La connaissance de l’importance quantitative de laprédation sur les communautés de bactéries au seindes sédiments et du transfert vers les échelonstrophiques supérieurs est encore réduite. Une desraisons pour lesquelles le rôle et l’importance desréseaux trophiques microbiens sont moins étudiésdans les sédiments que dans la colonne d’eau est quela communauté sédimentaire ne se prête pas auxmanipulations telles que les techniques de dilution etde filtration selon la taille des organismes, car cestechniques engendrent une désorganisation totale dela structure spatiale des sédiments. De nouvellestechniques basées sur l’utilisation des traceursisotopiques stables sont prometteuses. Des substratsspécifiques pour les microalgues et les bactérieshétérotrophes enrichis en 13C sont ainsi additionnésaux systèmes benthiques et leur devenir peut être suivipar traçage en étudiant les rapports isotopiques 13C/12Cdans les différents compartiments. Après un tempsd’incubation donné, des individus appartenant à lamacrofaune et à certaines espèces de la méiofaunesont récoltés et leurs signatures isotopiques δ13C sontanalysées à l’aide d’un spectromètre de masse.Cependant, il est très difficile de séparer physiquementles microalgues et les bactéries hétérotrophesbenthiques et des biomarqueurs spécifiques sont doncnécessaires pour mesurer le marquage de ces deuxgroupes. L’addition de NaH13CO3 à un biofilm dediatomées d’un sédiment intertidal a permis de montrerune incorporation croissante du traceur dans lesdiatomées pendant 4 heures et un transfert trophiquenotable vers les nématodes à partir d’une heured’incubation et de façon croissante jusqu’au 3e jourd’incubation. En même temps, les acides grasspécifiques des bactéries hétérotrophes s’enrichis-saient également en 13C, leur marquage en cet élémentétant maximal après un jour d’incubation (Middelburget al., 2000). Ceci montre un transfert trophique très
17
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES

rapide de la MO des diatomées vers les bactérieshétérotrophes probablement à travers le compartimentEPS (Figure 13-4). Dans une autre expérience utilisantle même type de biofilms photosynthétiques, lesbactéries hétérotrophes ont été marquées par l’additionde 13C glucose. De cette façon, il a été calculé que labiomasse bactérienne constitue 8 % et 11 % de lanourriture de la méiofaune et de la macrofaune,respectivement.
13.3.4. LE FONCTIONNEMENT DES RÉSEAUXTROPHIQUES MICROBIENS EN MILIEUX TERRESTRES
Le paragraphe 13.2.3 présente une partie desacteurs constituant les réseaux trophiques telluriques.Le présent paragraphe porte sur le rôle fonctionnel deces acteurs et de leurs interactions.
Il est maintenant largement admis que la structuredes réseaux trophiques du sol a des impacts directssur son fonctionnement en général (Beare et al.,1995).
La composante biotique contribue aux processusfonctionnels du sol via des mécanismes directs etindirects. Les mécanismes directs sont laconséquence de processus métaboliques dedégradation de molécules complexes et de libérationd’énergie, de carbone, d’azote et de minéraux àtravers la respiration, l’excrétion et la chimiotrophie.Les mécanismes indirects comprennent de nombreuxeffets de rétro-actions et les organismes responsablessont qualifiés d’« ingénieurs de l’écosystème* »(Anderson, 2000).
La décomposition de la MO dans les sols
La fonction de décomposition de la matièreorganique (MO) est évidemment l’un des rôlesmajeurs des réseaux trophiques du sol. Ladécomposition de la MO produit du CO2 mais aussides composés solubles qui sont des nutriments pourles végétaux. Dans les milieux terrestres « naturels »(non fertilisés), la quasi-totalité des besoins desplantes en azote et une fraction de leurs besoins enphosphore proviennent de la décomposition de la MO.Même dans les systèmes recevant des intrants*, lerôle de la décomposition de la MO demeure centraldans le maintien d’une production acceptable sur leplan environnemental.
En milieu non cultivé, 80-90 % de la productionprimaire retourne au sol sous forme de litière et deracines mortes. Les champignons et les procaryoteshétérotrophes sont les principaux acteurs de ladécomposition de ces apports végétaux. La faune
détritivore joue, au travers de la fragmentation et del’ingestion, essentiellement un rôle de facilitation del’activité microbienne (via notamment l’augmentationdes surfaces de colonisation). L’énorme palette decapacités métaboliques des micro-organismes permetà ces derniers de dégrader la quasi-totalité desmolécules présentes dans les végétaux. Dans denombreux écosystèmes, les champignons sont lesdécomposeurs « primaires » les plus abondants. Ilssont mieux adaptés que les bactéries pour débuter ladécomposition du matériel végétal non soluble dans lamesure où leurs hyphes peuvent pénétrer et proliférerà l’intérieur des cellules végétales. À l’instar desbactéries, ils possèdent une large gamme depossibilités métaboliques qui leur permettent,notamment, de dégrader des composants récalcitrantsdes cellules végétales, comme la lignine. De plus, lefait de pouvoir mobiliser des nutriments à travers toutle réseau d’hyphes leur donne la possibilité d’exploiterde nouveaux substrats même lorsque les teneurslocales en P et N sont faibles. Ces caractéristiquesd’exploitation active des substrats par leschampignons contrastent avec l’immobilité ou lamobilité passive (mouvements de l’eau du sol parexemple) des bactéries. En conséquence, cesdernières sont probablement plus fréquemmentinactives par manque de ressources dans leurenvironnement immédiat.
Le rôle de la prédation sélective
Comme cela a été évoqué précédemment, leschampignons et les procaryotes hétérotrophes sont lesacteurs prépondérants de la transformation de la MOdans les sols et, en particulier, de sadégradation/minéralisation. Cependant, l’activité deces micro-organismes est fortement affectée par desorganismes des niveaux trophiques supérieurs(Bardgett, 2005).
La consommation sélective des micro-organismespeut modifier l’abondance, la structure et l’activité descommunautés fongiques et bactériennes. Même si unnombre grandissant d’études fait état du caractèregénéraliste et/ou opportuniste des consommateurssecondaires, d’autres études mentionnent un véritable« choix » des prédateurs pour leurs proies. Certainsnématodes ont, par exemple, des pièces buccalesadaptées, soit à la consommation des bactéries(bactérivores), soit à la consommation deschampignons (fongivores) (Yeates et al., 1993). Il aégalement été montré que certains protozoairespeuvent détecter des signaux chimiques pourdiscriminer entre des proies bactériennes et algales
18
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S

potentielles (Verity, 1991). Cette consommationsélective des proies peut avoir un impact important surle fonctionnement de l’écosystème. Ainsi, en milieuforestier, la consommation sélective par le collemboleOnichiurus latus du champignon Marasmusandrosaceus se traduit par une diminution del’abondance de ce dernier au profit d’un autrechampignon Mycena galopus non consommé par lecollembole. Ce changement dans la structure de lacommunauté fongique a pour conséquence uneréduction du taux de décomposition des litières deconifère car M. galopus présente une efficiencespécifique de décomposition moindre que M.androsaceous.
La consommation d’une espèce de micro-organisme par un prédateur peut également stimulerl’activité de la proie sans que des mécanismes decompétition entrent en jeu. Hedlund et al. (1991) ontmontré, ainsi, que l’activité spécifique amylase duchampignon Mortiella Isabellina est très nettementsupérieure lorsque ce champignon est consommé parun collembole que lorsqu’il est protégé de la prédation.
Enfin, si l’on se réfère au rôle des animaux de tailleplus importante sur le devenir des acteurs de ladécomposition que sont les champignons et lesprocaryotes hétérotrophes, il convient de prendre encompte l’influence de l’ingestion des micro-organismeset de leur exposition au liquide intestinal. Les donnéesde la littérature sont à cet égard très contrastées. Eneffet, Brown (1995) a montré que la communautébactérienne était plus abondante et active aprèspassage dans le tractus intestinal des vers de terrealors qu’à l’inverse, Moody et al. (1996) ont observéune réduction de la germination de spores dechampignon après ingestion par un ver de terre.
Le rôle des autres interactions
biotiques et abiotiques
Certaines interactions reposent davantage surdes modifications physiques de l’habitat que sur desrelations trophiques. Elles ont une importanceconsidérable dans le fonctionnement du sol. C’est lecas des organismes qui ingèrent des débris végétauxet produisent des pelotes fécales dont la dimensionest très inférieure au matériel ingéré. La création deces agrégats d’origine biologique a pourconséquence de modifier fondamentalement lespropriétés physiques du sol (compaction élevée,capacité de rétention de l’eau accrue). Cette litièretransformée devient un milieu très favorable à lacomposante bactérienne et il en résulte uneaugmentation considérable du rapport
bactéries/champignons. Ainsi, il a été mis enévidence une multiplication par 100 des bactéries etpar seulement 3 des champignons présents dans despelotes fécales d’arthropodes par rapport au matérielinitial. Il est probable que la compaction du matérielconstitue le facteur directement impliqué dans ceteffet différentiel. Cette interaction est de typemutualiste dans la mesure où (1) la stimulation de lamicroflore dans les pelotes s’accompagne d’uneaugmentation de la minéralisation de la MO et (2) laré-ingestion par la faune des pelotes modifiéesparticipe à améliorer son alimentation. Ce systèmefonctionne comme un « rumen externe » et devienttrès important quand la faune consomme une litièrenon encore conditionnée par la microflore et pauvreen nutriments.
Outre la relation mutualiste décrite ci-dessus, lesingénieurs de l’écosystème* ou ingénieurs du solmodifient physiquement l’environnement et créent denouveaux habitats pour les micro-organismes et lamicrofaune du sol. C’est le cas des vers de terre, destermites et des fourmis. Par exemple, nombred’auteurs ont montré que la microflore était stimuléedans les structures créées par les vers de terre viaune amélioration de l’aération et des conditionshydriques du sol et/ou en favorisant l’accessibilitéaux ressources par la microflore (enfouissement dela litière, par exemple) (Brown, 1995). Il est à noterque, dans certains cas, cet effet stimulateur n’estque transitoire et que, sur le long terme, cesstructures peuvent, au contraire, freiner ladécomposition de la MO.
L’apport de matière organique, via la litièrevégétale, constitue la base vitale du fonctionnementde ces réseaux comportant une très grande diversitéd’organismes telluriques, essentiellementhétérotrophes. Ceci est à rapprocher de la notion dechaîne trophique basée sur les détritus (Detritus foodchain) dans les milieux aquatiques, évoquée enintroduction. Dans un contexte pédo-climatique donné,la quantité de litière retombant au sol, ainsi que saqualité, exercent un fort contrôle sur la structure desréseaux trophiques. Les études, visant à comprendrela manière dont cette structure est régulée dans le solpar l’environnement, qui restaient encore peunombreuses il y a quelques années (Wardle, 2002),s’intensifient à l’heure actuelle. Ceci est dû,notamment, à de nouveaux développementsméthodologiques. Parmi eux, on peut citer l’utilisationdes variations d’abondance naturelle des isotopesstables du carbone et de l’azote (13C/12C et 15N/14N)(Schmidt et al., 2004).
19
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES

13.4. LES FACTEURS DE CONTRÔLEDES RÉSEAUX MICROBIENS
13.4.1. LES FACTEURS DE CONTRÔLE DES RÉSEAUXMICROBIENS PÉLAGIQUES MARINS ET LACUSTRES
Dans les milieux pélagiques, différentesstructurations de réseaux trophiques peuvent co-exister. La prédominance d’un réseau trophique parrapport à un autre est sous le contrôle de forçagesenvironnementaux comme la nature et la concentrationdes nutriments, la température (responsable de lastratification ou du mélange vertical), la pollution, etc.Ainsi, la différence de structure du réseau trophiquepélagique entre les écosystèmes marins hauturiers etles environnements côtiers est liée aux différents typesde forçages environnementaux qui s’exercent. Lemilieu marin côtier est régulièrement enrichi enéléments nutritifs par les fleuves et la pluie. Celaconduit à la prédominance du réseau trophiqueherbivore avec une forte biomasse phytoplanctoniqueen milieu côtier par rapport au large, ce dernier milieuétant caractérisé, généralement, par un réseautrophique microbien.
Les forçages environnementaux font évoluer lastructure du réseau trophique au profit des micro-organismes qui s’adaptent le mieux au nouvelenvironnement. Cette évolution temporelle est àrapprocher du concept de succession écologique. Lasuccession des réseaux trophiques peut conduire, defaçon réversible, de la boucle microbienne auxréseaux trophiques microbien, multivore et herbivore.La notion de « bascule d’équilibre et de réversibilitéd’état » entre les différents réseaux trophiques signifieque sous l’effet des forçages environnementaux, unesituation peut basculer de l’une vers l’autre. Lesconséquences de la dominance d’un réseau trophiquepar rapport à un autre sont très importantes, car ellesconduisent l’écosystème vers des niveauxcomplètement différents en ce qui concerne laproduction, la régénération, les flux de matière, lebroutage, la séquestration ou l’exportation de matière(e.g. pêche). Le réseau trophique herbivore et laboucle microbienne seraient des systèmes transitoireset instables, alors que les réseaux trophiquesmultivore et microbien seraient beaucoup plus stableset durables (Legendre et Rassoulzadegan, 1995).
En dehors des forçages climatiques (vent,température, lumière), on considère généralementdeux groupes principaux de facteurs de contrôle desréseaux trophiques microbiens pélagiques. Lesfacteurs ascendants ou ‘bottom-up’ sont constitués par
les ressources et affectent plus le taux de croissancedes organismes que leur biomasse. Les facteursdescendants ou top-down sont représentés parl’ensemble des pressions de prédation qui conduisentà des prélèvement de biomasse. Par extension, tousles facteurs affectant la biomasse, comme la lysevirale où le parasitisme, sont considérés comme desfacteurs descendants, même si leurs effets sur lastructure et le fonctionnement des réseaux trophiquessont fondamentalement différents. Selon McQueen etal., (1989), les facteurs ascendants et descendants nesont pas exclusifs et l’intensité relative de chaque typede contrôle varie dans l’espace et dans le temps.
Les aspects de prédation sur les communautéspicoplanctoniques et le recyclage des élémentsminéraux au sein du réseau et de la bouclemicrobienne étant abordés dans les sous-chapitres13.3.1 et 13.3.2, seule la régulation via la lyse viraleest évoquée ci-dessous.
Les rôles des virus dans les réseaux trophiques
microbiens aquatiques
L’abondance et la diversité des virusL’écologie virale aquatique est une discipline
relativement récente. Mieux étudiés dans lesécosystèmes marins (Fuhrman, 1999 ; Suttle ; 2005 ;Suttle, 2007) que dans les eaux douces (Sime-Ngando, 1997, Sime-Ngando et al., 2003), les virussont omniprésents dans l’environnement aquatiquepélagique et benthique où, très souvent, ilsreprésentent l’entité biologique la plus abondante. Lesdensités moyennes fluctuent généralement entre 104
et 108 virus ml-1 dans les écosystèmes pélagiqueslacustres, et sont étroitement liées à l’abondance desprincipaux hôtes potentiels (les procaryotes) et austatut trophique du milieu. En milieu lacustre, les plusfortes abondances virales (6,5 x 108 à 1,83 x 1010 virusml-1) ont été rapportées dans les eaux interstitielles dusédiment superficiel d’un petit lac oligomésotrophe (lacGilbert, Canada) par Maranger et Bird (1996). Entermes de carbone, les virus représentent le deuxièmeréservoir de biomasse dans les écosystèmesaquatiques après les procaryotes (Mann, 2003 ;Weinbauer, 2004 ; Suttle, 2005).
La diversité des virus reste largement méconnueen milieu aquatique où il existe une forte variété deformes phénotypiques, généralement dominées pardes virus encapsidés de type phage (Planche 13-2).Les familles morphologiques les mieux représentéessont les Myovirus (phages à queue contractile,exemple phage T4) et les Siphovirus (phages à longue
20
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S
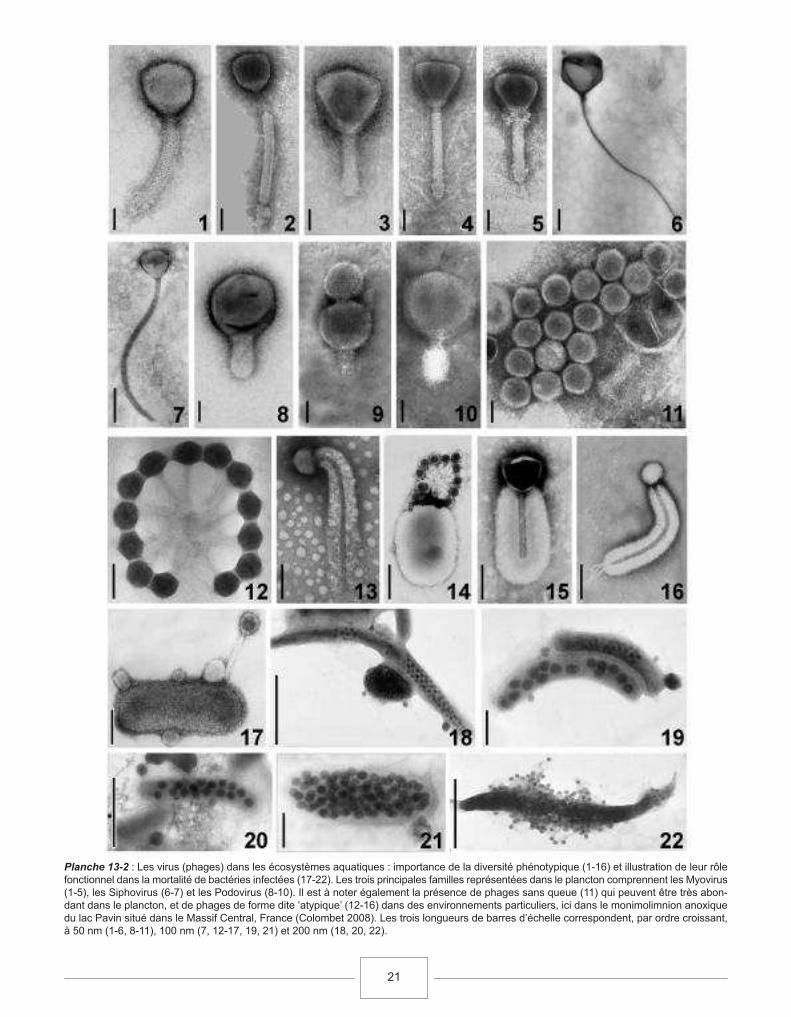
21
Planche 13-2 : Les virus (phages) dans les écosystèmes aquatiques : importance de la diversité phénotypique (1-16) et illustration de leur rôlefonctionnel dans la mortalité de bactéries infectées (17-22). Les trois principales familles représentées dans le plancton comprennent les Myovirus(1-5), les Siphovirus (6-7) et les Podovirus (8-10). Il est à noter également la présence de phages sans queue (11) qui peuvent être très abon-dant dans le plancton, et de phages de forme dite ‘atypique’ (12-16) dans des environnements particuliers, ici dans le monimolimnion anoxiquedu lac Pavin situé dans le Massif Central, France (Colombet 2008). Les trois longueurs de barres d’échelle correspondent, par ordre croissant,à 50 nm (1-6, 8-11), 100 nm (7, 12-17, 19, 21) et 200 nm (18, 20, 22).

queue flexible, exemple phage lambda), suivies desPodovirus (phages à courte queue non contractile,exemple phage T7). Les traits phénotypiques sontrévélateurs des pressions sélectives subies par lesvirus. Ainsi, les Myovirus sont essentiellement lytiquesavec un large spectre d’hôtes, présentant un avantagecompétitif assimilable à des stratèges r* avec defortes capacités de prolifération. Contrairement auxMyovirus, les Podovirus et, dans une moindre mesure,les Siphovirus, sont plus spécifiques de leurs hôtes.De plus, les Siphovirus peuvent facilement crypter leurpropre génome dans celui de leurs hôtes pourplusieurs générations (lysogénie). Ils présentent doncun faible taux de prolifération assimilable à unestratégie de développement de type K* (Suttle,2005). D’un point de vue génomique, on suppose queles virus aquatiques constituent un réservoir importantde diversité génétique et biologique, sans doute le plusvaste de la biosphère, étant donné leur mode dedéveloppement et la susceptibilité de toutes lescellules vivantes aux infections virales. Ainsi, uneétude métagénomique a montré que 200 litres d’eaude mer contiennent entre 400 et 7000 génotypesviraux différents et que 1 kg de sédiments contiendrait1 million de génotypes viraux. De manière générale,les données métagénomiques montrent que larichesse génétique des virus aquatiques, bien quesous échantillonnée, reste largement supérieure àcelle des procaryotes.
Les principaux rôles des virus
en milieu aquatique pélagique
La plupart des virus aquatiques libres seraientinfectieux, certains pouvant conserver ce pouvoirinfectieux très longtemps, plusieurs dizainesd’années pour ce qui concerne les virus dessédiments (Suttle, 2005).
Il est admis que l’intervention des virus estessentielle dans différents processus écologiques sedéroulant dans l’environnement aquatique, notammentdans les processus de perte affectant lescommunautés microbiennes (archées, bactérieshétérotrophes, cyanobactéries, microalgues, etc.),dans la dynamique de la biodiversité microbienne etdans le cycle des éléments chimiques (carbone, azote,phosphore, etc.) ; autant de processus qui sont à labase du fonctionnement des réseaux trophiques(Fuhrman, 1999).
Ainsi, on estime que la mortalité microbienne liée àla lyse virale (Planche 13-2) est équivalente à celle dueà l’activité des organismes bactérivores, même si la lysevirale semble être très faible au niveau du benthos où
les virus libres sont, paradoxalement, plus abondantsque dans la colonne d’eau (Filippini et al., 2006).
Les processus viraux qui sont susceptiblesd’intervenir dans la dynamique de la diversitéspécifique et génétique des micro-organismesprocaryotes (hôtes les plus abondants et les plusétudiés) sont résumés sur la Figure 13-5. Le principalfacteur limitant de la prolifération virale est ladisponibilité des hôtes. Outre l’action structurante desproduits de la lyse virale, les virus infectent préféren-tiellement les populations les plus abondantes,permettant ainsi le développement des espèces lesmoins compétitives vis-à-vis d’autres facteurs derégulation comme la disponibilité des ressources(hypothèse dite de « killing the winner », Thingstad,2000). La conversion lysogénique de bactériescompétentes permet la dissémination et le maintiendes peuplements viraux, tout en procurant aux hôtesde nouvelles propriétés phénotypiques, génotypiqueset métaboliques, comme la résistance à la surinfectionou la production de toxines.
Par leurs activités, les virus représentent donc devéritables catalyseurs dans les cyclesbiogéochimiques qui se déroulent dans lesécosystèmes aquatiques. Ils court-circuitent letransfert de la matière vers les niveaux trophiquessupérieurs, en réduisant les particules vivantes enmatière organique dissoute et en accélérant lesprocessus respiratoires. Les produits de lysecomprennent également des éléments autres que lecarbone, l’azote ou le phosphore. Ainsi, dans certainscas, le fer contenu dans les produits de la lyse viralepeut combler les besoins métaboliques desorganismes planctoniques. Les virus étant desparticules riches en azote et en phosphore, laproduction virale correspond à une séquestration deces éléments dans la colonne d’eau (Suttle 2005,2007). Cette séquestration change les rapports deséléments minéraux dans la colonne d’eau (C, N et P),ce qui peut affecter la production primaire essentielleau fonctionnement des réseaux trophiquesaquatiques.
Les virus interviendraient également, de manièredirecte (par transduction) ou indirecte (partransformation à partir des produits de la lyse) dans lestransferts horizontaux de gènes entre les micro-organismes (Weinbauer et Rassoulzadegan, 2004).Cet aspect de l’écologie virale reste, cependant, trèslargement inexploré. A titre d’exemple, il a été montréque certains génomes viraux, notamment ceux desmyovirus de picocyanobactéries, présentent desgènes typiquement viraux, qui codent pour des
22
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S

protéines photosynthétiques (Mann, 2003). Un virusgéant, comportant un phénotype mimant un micro-organisme et, de ce fait, nommé Mimivirus(étymologiquement ‘mimicking microbe’), a étérécemment découvert chez une amibe aquatique(Acanthamoeba polyphaga) isolée dans des tours derefroidissement en Angleterre (La Scola et al. 2003).Ce virus est géant par son génome qui comprend 1,2million de paires de bases, et code pour plus de 900protéines différentes.
De manière générale, les recherches récentes enécologie virale sont source de nouvellesconnaissances, non seulement dans le domaine de ladiversité du vivant et du fonctionnement desécosystèmes, mais également dans le domaine del’évolution (Forterre, 2007).
13.4.2. LES PRINCIPAUX FACTEURS DE CONTRÔLEDES RÉSEAUX MICROBIENS BENTHIQUES
MARINS ET LACUSTRES
Les effets du broutage (« top-down ») exercé par lamacrofaune benthique ont principalement été mis enévidence sur les communautés microbiennes debiofilms. Ces effets du broutage sont directs etprovoquent une réduction de la diversité et de labiomasse des microphytes de grande taille. De façonindirecte, le broutage engendre une augmentation de
la capacité photosynthétique des micro-organismesphototrophes du biofilm non consommés.
Il est à noter que le comportement des prédateursest également un facteur à considérer dans lecontrôle des réseaux trophiques benthiques. Ainsi, lebroutage par des gastropodes, en eau douce et enmilieu marin, dépend de leurs déplacements et deleurs visites aux habitats benthiques. Leur broutagesur les biofilms est souvent drastique et c’est pourcette raison que ces visites sont appelées « les raidstrophiques ». A la suite d’un raid trophique, unnouveau cycle de colonisation peut s’enclencher etun nouveau biofilm photosynthétique est formélorsque l’intensité du broutage par la macrofaunedevient très faible.
Ainsi, la pression de broutage par la macrofauneempêche souvent une accumulation de la MO commeon peut l’observer pour les tapis microbiens. Laprésence de tapis microbiens est actuellement limitéeaux zones où la pression de prédation est moindre,c’est-à-dire les environnements extrêmes (milieuxhypersalés, hautes et basses températures, forthydrodynamisme). Néanmoins, il est à noter qu’àl’époque du précambrien, avant l’apparition desmétazooaires eucaryotes, les tapis cyanobactériensétaient très largement répandus sur terre (cf.Chapitre 4.x.x).
23
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES
Figure 13-5 :
Rôles potentiels des virus dans la
dynamique de la diversité micro-
bienne et le fonctionnement des
écosystèmes aquatiques.
Ce schéma est centré sur la diversitédes procaryotes qui représentent leplus grand réservoir d’hôtes pour lesvirus aquatiques. Les interactionsentre ces deux communautés sont lesmieux étudiées en écologie viraleaquatique. Modifiée de Weinbauer &Rassoulzadegan (2004).

La production des micro-organismes photosynthé-tiques benthiques dans les sédiments superficielsintertidaux est principalement soumise aux processusde régulation de type « bottom-up » c’est-à-direessentiellement limitée par la disponibilité desressources telles que la lumière et les nutriments.Cependant, des études ont indiqué que la biomasse etla production des micro-organismes benthiques sontétroitement couplées à l’intensité du broutage exercépar les consommateurs méiobenthiques (Buffan-Dubau et Carman, 2000). Le broutage assure unerégulation qui, toutefois, ne représente pas leprocessus de contrôle primordial. En revanche, laproduction des microphytes benthiques estindirectement favorisée par le broutage de la méiofaunequi modifie le contrôle « bottom-up » potentiel enaméliorant les conditions de pénétration de la lumièreet la régénération des nutriments dans les sédimentssuperficiels côtiers. La bioturbation et le remaniementdes sédiments par l’activité de la méiofaune peuventaussi augmenter la porosité et les taux de transport dessolutés dans les sédiments et les agrégats microbiens,ce qui améliore la disponibilité des nutriments dissouspour les micro-organismes photo-autotrophes (Aller etAller, 1992 ; Mathieu et al., 2007).
Le broutage par la méiofaune induit des effetsindirects pouvant agir sur la structure descommunautés bactériennes (composition etabondance relative des taxons bactériens) etsusceptibles de provoquer une augmentation del’activité bactérienne des sédiments superficiels decours d’eau en raison de l’excrétion de métabolites parles invertébrés (Traunspurger, 1997).
13.4.3. LES FACTEURS DE CONTRÔLEDES RÉSEAUX MICROBIENS TERRESTRES
Compte tenu du fait que la quasi-totalité de laproduction primaire des écosystèmes terrestresprovient des végétaux supérieurs, il convientévidemment de considérer les plantes comme deséléments centraux de la régulation des organismes dusol et de leurs interactions, au moins aussi importantsque les facteurs environnementaux et lesperturbations.
La régulation par les facteurs environnementaux
et les perturbations
Comme dans les milieux aquatiques, le réseautrophique microbien du sol dépend à la fois de facteursde contrôle top-down et bottom-up. Il est généralementadmis que les bactéries sont prioritairement régulées
par la prédation alors que les champignons sontsurtout régulés par les ressources. Il est à notercependant qu’il existe de nombreux cas de contrôles« top down » des champignons. A titre d’exemple, laprésence de protozoaires et de nématodes dans larhizosphère des arbres réduit la colonisation desracines d’arbres par des champignons mycorhiziens(Wardle, 2002).
La diversité au sein d’un groupe donné dans le soldépend d’un grand nombre de facteurs parmi lesquelson peut mentionner la disponibilité des ressources, latempérature, le pH ou l’état hydrique, ainsi que deperturbations telles que la mise en culture ou desévènements climatiques extrêmes et rapides.Certaines études montrent qu’une augmentation desressources s’accompagne d’une stimulation de ladiversité fonctionnelle microbienne du sol (Degens etHarris, 2000). Cette observation va à l’encontre de lathéorie élaborée à partir des végétaux, à savoir quel’augmentation des ressources devrait avoir pourconséquence une réduction de la biodiversité. Dansl’étude de Degens et Harris (2000), la diversité estmesurée in vitro par l’aptitude de la microflore du sol àutiliser des substrats apportés et il est probable queles résultats observés traduisent davantage lesaptitudes métaboliques des bactéries que deschampignons. Les bactéries sont, en effet, aptes àcroître rapidement sur les substrats et seraient, de cefait, moins régulées par la compétition que desgroupes à croissance plus lente comme leschampignons.
Il est pourtant généralement reconnu que ladiversité de la faune du sol se maintient même lorsqueles conditions deviennent plus favorables. Yeates etBongers (1999) observent que la diversité desnématodes (groupe que l’on suppose peu régulé parla compétition) se réduit progressivement avec unaccroissement de la concentration en métaux lourdset n’est pas réduite par un apport calcique au sol. Plusgénéralement, la diversité de nombreux groupes dansle sol est plus importante sur un mull* à Fagussylvatica où règnent des conditions relativementfavorables pour l’activité des organismes (pH : 4,3-6,8,capacité d’échange des cations élevée) que dans unmull sous la même couverture végétale où lesconditions sont nettement moins favorables (pH : 3-4,capacité d’échange des cations faible). La plupart desétudes tendent à montrer que la diversité desorganismes décomposeurs ne diminue pas avecl’amélioration des conditions de milieu. Cesorganismes ne semblent donc pas, de ce point de vue,suivre les modèles les plus fréquemment observés
24
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S

chez les plantes. A l’inverse, et comme indiqué audébut de ce paragraphe, certains groupes comme leschampignons peuvent être régulés par la compétition.Ainsi, Egerton-Warburton et Allen (2000) montrent quel’addition d’azote réduit la diversité des champignons.Ce résultat suggère que l’accroissement desressources favorise l’exclusion compétitive*.
La régulation par les facteurs biotiques
Les études portant sur la régulation biotique desorganismes du sol et de leurs activités sont peunombreuses, à l’exception notable des relationsplantes-micro-organismes. La présence d’unemicrofaune consommatrice de champignons influencegénéralement, mais non systématiquement, ladiversité fongique. Ceci peut s’expliquer par l’effetcombiné de l’importance du caractère sélectif desconsommateurs (voir plus haut) et du rôle de lacompétition dans la structuration des communautésfongiques (McLean et Parkinson, 2000).
Par ailleurs, les organismes ingénieurs tels que lesvers de terre peuvent avoir un impact sur la diversité –notamment des champignons – par leur rôle sur lamodification de la structure du sol, qui conduit à lacréation d’habitats nouveaux. McLean et Parkinson(2000) ont observé, ainsi, à partir d’expériences enmicrocosmes, que le ver de terre Dendrobaenaoctaedra pouvait modifier la structure de lacommunauté fongique dans le sol et, dans certainesconditions, augmenter la diversité des champignons endiminuant l’intensité de la compétition. Dans d’autresexpériences, ces mêmes auteurs montrent que laprésence de D. octaedra peut, au contraire, réduire ladiversité en favorisant une espèce à croissance rapide,cette dernière devenant très dominante.
De très nombreux travaux prouvent, par ailleurs,que les espèces végétales peuvent orienter ladiversité, l’abondance et l’activité des micro-organismes du sol. Ainsi, Badejo et Tian (1999)montrent, dans des parcelles en monoculture auNigeria, que la diversité de certains groupes de lamicrofaune du sol était plus élevée sous l’espèceLeucaena leococephala que sous trois autres espècesvégétales étudiées. De même, il a été montré que larichesse spécifique des champignons mycorhizienssur les racines des végétaux est très différente entrecinq espèces de plantes sur le même site.
Enfin, le rôle de la composition de la végétation surla diversité des organismes du sol a également étémontré dans les différents stades de la successionvégétale qui sont dominés par des types fonctionnelsde plantes différents (Wardle, 2002).
13.5. CONCLUSION
Les organismes des réseaux trophiquesmicrobiens (RTM), incluant les virus, les archées, lesbactéries, les champignons, le phytoplancton, lephytobenthos et les protozoaires, jouent un rôleessentiel dans le fonctionnement des écosystèmesaquatiques, benthiques et terrestres. Il existecependant des différences fondamentales entre cesdifférents écosystèmes au niveau du rôle fonctionneldes micro-organismes.
Le fonctionnement des écosystèmes aquatiquespélagiques est largement dépendant de phénomènesd’advection, de mélange vertical ou de stratification dela masse d’eau et de leurs effets induits. Le mode devie planctonique est le mieux adapté à cetenvironnement dynamique. En conséquence, lesécosystèmes aquatiques pélagiques sont particulière-ment originaux puisque les communautésmicrobiennes assurent la quasi-totalité de laproduction primaire, constituent l’essentiel de labiomasse présente et forment, ainsi, la base de tousles réseaux trophiques aquatiques.
En lien avec la nature physique des sédiments etdes sols, le fonctionnement des RTM benthiques etterrestres est sensiblement différent.
Dans les écosystèmes terrestres, au contraire desmilieux aquatiques pélagiques et même s’il existe lesmêmes groupes de micro-organismes que dans l’eau,avec une diversité au moins aussi importante, la basefonctionnelle de ces écosystèmes est constituée parles plantes supérieures comme principaux producteursprimaires. De plus, ces dernières exercent, via leursystème racinaire, un contrôle important sur lastructuration des sols et sur les interactions bio-physico-chimiques. Dans ce contexte, lesmicro-organismes jouent, cependant, un rôleprimordial dans les cycles biogéochimiques terrestreset ils interviennent comme des « bioréacteurs », entransformant et en recyclant la matière organiqueprimaire provenant des plantes.
Sur ce dernier point, les sédiments possèdent,selon les cas, des similitudes soit avec les sols,lorsqu’une végétation de phanérogames aquatiquesassure l’essentiel de la production primaire, soit avecles systèmes pélagiques lorsque les micro-organismesautotrophes de la surface du sédiment ou ceuxprovenant par sédimentation de la colonne d’eaureprésentent l’essentiel des apports en MO.
En raison des caractéristiques biophysiques dessols, à macro-, méso- et micro-échelles, les réseaux
25
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES

trophiques microbiens terrestres sont généralementappréhendés de manière sensiblement différente et lanotion de réseau trophique a beaucoup moinsimprégné les études sur les sols que sur lesécosystèmes aquatiques. Les processus d’agrégation,en favorisant une structuration tridimensionnelle desfacteurs environnementaux, conduisent à unedistribution tridimensionnelle des caractéristiquestaxonomique et fonctionnelle des micro-organismes.Comme dans les sédiments, cette structuration setraduit par une dissociation partielle dans l’espace destypes de micro-organismes.
En outre, la présence d’oxygène sous formegazeuse dans le sol, associée aux mécanismesd’agrégation, a pour conséquence de favoriser la co-existence à micro-échelle de zones aérobies etanoxiques. Ces micro-gradients d’oxygénationorientent la « colonisation » des agrégats par lesmicro-organismes en fonction de leur caractèreaérobie strict, aérobie-anaérobie ou anaérobie strict.De tels gradients d’oxygénation existent également auvoisinage immédiat des racines et affectent ladistribution des micro-organismes en fonction de lagamme d’accepteurs d’électrons qu’ils sont capablesd’utiliser dans les processus de respiration.
Il est à noter, cependant, que les milieuxaquatiques pélagiques ne sont pas, eux non plus,strictement homogènes, notamment en raison demicro-agrégats qui peuvent conduire à la présence, àmicro-échelle, de zones anoxiques au sein d’unecolonne d’eau globalement aérobie.
Plus généralement, la structure des réseauxtrophiques microbiens dans les milieux aquatiquespélagiques, benthiques et terrestres reflètel’adaptation des micro-organismes face aux forçagesphysiques, chimiques et biologiques del’environnement. La compréhension des mécanismesresponsables de ces adaptations aux conditionsfluctuantes de l’environnement constitue un enjeumajeur pour les années à venir.
REMERCIEMENTS
Nous remercions L. Legendre, F. Rassoulzadegan, M.Troussellier, F. Vidussi et les autres membres duCNRS-GDR 2476 « Réseaux Trophiques Aquatiques »pour toutes les discussions enrichissantes que nousavons eues sur la thématique développée dans cechapitre.
LECTURES COMPLÉMENTAIRES
OUVRAGES GÉNÉRAUX
Bardgett, R.D., Usher, M.B., and Hopkins D.W. (2005)Biological Diversity and Functions in Soils. CambridgeUniversity Press, Cambridge, UK.
Carpenter, S.R. (1988) Complex interactions in lakecommunities. Springer-Verlag, N.Y.
Reid, P.C., Turley, C.M., and Burkill, P.H. (1991)Protozoa and their role in marine processes. Springer,New York.
ARTICLES
Aller, R.C., and Aller, J.Y. (1992) Meiofauna and solutetransport in marine muds. Limnol Oceanogr 37: 1018-1033.
Amblard, C., Boisson, J.C., Bourdier, G., Fontvieille,D., Gayte, X., and Sime-Ngando, T. (1998) Ecologiemicrobienne en milieu aquatique : des virus auxprotozoaires. Rev. Sc. Eau, N° Spécial : Les Sciencesde l’Eau : Bilan et perspectives, 145-162.
Amblard, C., Carrias, J.F., Bourdier, G., and Maurin, N.(1995) The microbial loop in a humic lake : seasonaland vertical variations in the structure of the differentcommunities. Hydrobiologia 300/301: 71-84.
Anderson, J.M. (2000) Food web functioning andecosystem processes. In Invertebrates as webmastersin ecosystems (Eds Colemab D.C., and Hendrix P.F.,C.A.B. International, Wallunford, UK. Pp. 3-24.
Azam, F., Fenchel, T., Field, J.G., Gray, J.S., Meyer-Reil, L.A., and Thingstad, F. (1983) The ecological roleof water-column microbes in the sea. Mar Ecol ProgSer 10: 257-263.
Badejo, M.A., and Tian, G. (1999) Abundance of soilmites under four agroforestry tree species withcontrasting litter quality. Biology and Fertility of Soils30: 107-112.
Bardgett, R.D. (2005) Organism interactions ans soilprocesses. In The Biology of Soil, A community andEcosystem Approach (Eds M.J. Crawley, C. Little,T.R.E. Southwood, and S. Ulfstrand), Oxford UniversityPress, Oxford, UK, pp. 57-85.
Bardgett, R.D. (2005) The diversity of life in soil. In TheBiology of Soil, A community and EcosystemApproach(eds M.J. Crawley, C. Little, T.R.E. Southwood, and S.Ulfstrand), Oxford University Press, Oxford, UK,pp. 24-56.
Beare, M.H., Coleman D.C., Crossley, D.A., Hendrix,P.F., and Odum, E.P. (1995) A hierarchical approach toevaluating the significance of soil biodiversity tobiogeochemical cycling. Plant and soil 170: 5-22.
26
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S

Bec, A., Martin-Creuzburg, D., and von Elert, E. (2006)Trophic upgrading of autotrophic picoplankton by theheterotrophic nanoflagellate Paraphysomonas sp.Limnol Oceanogr 51: 1699-1707.
Brown, G.G. (1995) How do earthworms affectmicrofloral and faunal community diversity. Plant andSoil 170: 209-231.
Buffan-Dubau, E., and Carman, K.R. (2000) Diel feedingbehavior of meiofauna and their relationships withmicroalgal resources. Limnol Oceanogr 45: 381-395.
Carrias, J.F., Amblard, C., and Bourdier, G. (1998)Seasonal dynamics and vertical distribution ofplanktonic ciliates and their relationship to microbialfood resources in the oligomesotrophic lake Pavin.Archiv für Hydrobiol 143: 227-255.
Colombet, J. (2008) Importance de la variabilitéverticale dans un lac méromictique profond : diversitéet activité lysogène des communautés virales. Thèsede Doctorat, Université Blaise Pascal, 204 p.
Curtis, T.P., Sloan, W.T., and Scannel, J.W. (2002)Estimating prokaryotic diversity and its limits.Proceedings of the National Academy of Science USA99: 10494-10499.
Cushing, D.H. (1989) A difference in structure betweenecosystems in strongly stratified waters and in thosethat are only weakly stratified. J. Plank. Res 11: 1-13.
De Ruiter, P.C., Neutel, A.N., and Moore, J.C. (1995)Energetics, patterns of interactions strengths andstability in real ecosystems. Science 269: 1257-1260.
De Ruiter, P.C., Neutel, A.N., and Moore, J.C. (1997)Soil foodweb interactions and modelling: In Fauna inSoil Ecosystems (ed. G. Benckiser) Dekker, New York,pp. 363-386.
Degens, B., and Harris, J.A. (2000) Decreases inorganic C reserves in soil can reduce the catabolicdiversity of soil microbial communities. Soil Biologyand Biochemistry 32: 189-196.
Delong, E.F. (1992) Archaea in coastal marineenvironments. Proceedings of the National Academyof Science USA 89 : 5685-5689.
Dolan, J.R. (1997) Phosphorus and ammoniaexcretion by planktonic protists. Mar Geol 139: 109-122.
Egerton-Warburton, L., and Allen, E.B. (2000) Shifts inarbuscular myccorhizal communities alond ananthropogenic nitrogen deposition gradient. EcologicalApplications 10: 484-496.
Filippini, M., Buesing, N., Bettarel, Y., Sime-Ngando,T., and Gessner, M.O. (2006) Infection paradox: highabundance but low impact of freshwater benthic
viruses. Appl Environ Microbiol 72: 4893-4898.
Foissner, W. (1997a) Global soil ciliate (Protozoa :Ciliophora) diversity : a probability-based approachusing large sample collections from Africa, Australiaand Antartica. Biodiversity and Conservation 5: 1627-1638.
Foissner, W. (1997b) Soil ciliates (Protozoa :Ciliophora) from evergreen rain forest of Australia,South America and Costa Rica: diversity anddescription of new species. Biology and Fertility ofSoils 25: 317-339.
Forterre, P. (2007) Microbes de l’enfer. Edition Belin,Paris 250pp.
Fouilland, É., Gosselin, M., Rivkin, R.B., Vasseur, C.,and Mostajir, B. (2007) Nitrogen uptake byheterotrophic bacteria and phytoplankton in Arcticsurface waters. J. of Plankton Res 29: 369-376.
Fuhrmann, J.A. (1999) Marine viruses and theirbiogeochemical and ecological effects. Nature 399:541-548.
Hedlund, K., Boddy, L., and Preston, C.M.(1991) Mycelial response of the soil fungus, MortiellaIsabellina, to grazing by Onichiurus armatus(collembolan). Soil Biology and Biochemistry 23:361-366.
Hobbie, J.E., Daley, R.J., and Jasper, S. (1977) Use ofNuclepore filters for counting bacteria by fluorescencemicroscopy. Appl. Environ. Microbiol 33: 1225-1228.
Jardillier, L., Boucher, D., Personnic, S., Jacquet, S.,Thenot, A., Sargos, D., Amblard, C., and Debroas, D.(2005) Relative importance of nutrients and mortalityfactors on prokaryotic community composition in twolakes of different trophic status: Microcosmexperiments. FEMS Microbiol Ecol 53: 429-443.
Kagami, M., De Bruin, A., Ibelings, B.W., and VanDonk, E. (2007) Parasitic chytrids: their effects onphytoplankton communities and food-web dynamics.Hydrobiologia 578: 113-129.
Kolber, Z.S., Van Dover, C.L., Niederman, R.A., andFalkowski, P.G. (2000) Bacterial photosynthesis insurface waters of the open ocean. Nature 407: 177-179.
La Scola B., Audic S., Robert C., Jungang L., deLamballerie X., Drancourt M., Birtles R., Claverie J.M.,Raoult D. (2003) A giant virus in amoebae. Science299: 2033.
Laval-Peuto, M., Heinbokel, J.F., Anderson, R.,Rassoulzadegan, F., and Sherr B.F. (1986) Role ofmicro- and nanozooplankton in marine food webs.Insect Sci. Applic 7: 387-395.
Lavelle, P., Decaëns, T., Aubert, M., Barot, S., Blouin,
27
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES

M., Bureau, F., Margerie P., Mora P., and Rossi J.P.(2006) Soil invertebrates and ecosystem services.European Journal of Siol Biology 42: 3-15.
Lefèvre, E., Bardot, C., Noël, C., Carrias, J.-F.,Viscogliosi, E., Amblard, C., and Sime-Ngando, T.(2007) Unveiling fungal zooflagellates as members offreshwater picoeukaryotes: evidence from a moleculardiversity study in a deep meromictic lake. EnvironMicrobiol 9: 61-71.
Legendre, L. and Rassoulzadegan, F. (1995) Planktonand nutrient dynamics in marine waters. Ophelia 41:153-172.
Li, K.W., Subba Rao, D.V., Harrison, G.W., Smith, C.J.,Cullen, J.J., Irwin, B., and Platt, T. (1983) Autotrophicpicoplankton in the tropical ocean. Science 219: 292-295.
Mann, N.H. (2003) Phages of the marinecyanobacterial phytoplankton. FEMS Microb Rev 27:17-34.
Mathieu, M., Leflaive, J., Ten-Hage, L., de Wit, R., andBuffan-Dubau, E. (2007) Free-living nematodes affectoxygen turn-over of artificial diatom biofilms. AquatMicrob Ecol 49: 281-291.
McLean, M.A., and Parkinson, D. (2000) Fieldevidence of the effects of the epigenic earthwormDendrobaena octaedra on the microfungal communityin pine forest floor. Soil Biology and Biochemistry 32:351-360.
McQueen, D.J., Johannes, M.R.S., Post, J.R.,Steward, T.J., and Lean, D.R.S. (1989). Bottum-up andtop-down impacts on freshwater pelagic communitystructure. Ecol Monogr 59: 289-310.
Middelburg, J.J., Barranguet, C., Boschker, H.T.S.,Herman, P.M.J., Moens, T., and Heip, C.H.R. (2000)The fate of intertidal microphytobenthos carbon: an insitu 13C labelling study. Limnol. Oceanogr 45: 1224-123.
Mittelbach, G.G., Steiner, C.F., Scheiner, S.M., Gross,K.L., Reynolds, H.L.; and Waide, R.B. (2001) What isthe observed relationship between species richnessand productivity? Ecology 82: 2381-2396.
Moody, S.A., Piearce, T.G., and Dighton, J. (1996)Fates of some fungal sporesassociated with wheatstraw decomposition on passage through the guts ofLumbricus terrestris and Aporrectoeda longa. SoilBiology and Biochemistry 28: 533-537.
Newbould, P. (1982) Biological nitrogen fixation inupland and marginal areas. Philosophical Transactionsof The Royal Society, London B 296: 405-417.
Pomeroy, L.R. (1974) The ocean’s food web, a
changing paradigm. BioScience 24: 499-504.
Pomeroy, L.R. (1991) Status and future needs inprotozoan ecology. In: Reid, P.C., Turley, C.M., andBurkill, P.H. (Eds), Protozoa and their role in marineprocesses. p. 475-492. Nato Asi series. Springer,New York.
Renault, P., and Stengel, P. (1994) Modelling oxygendiffusion in aggregated soils: anaerobiosis inside theaggregates. Soil Sci. Soc. Am. J 58: 1017-1023.
Ruellan, A., and Dosso, M. (1998) Regard sur le sol(Foucher et Aupelf éditeurs), 192 p.
Schmidt, O., Curry, J.P., Dyckmans, J., Rota, E., andScrimgeour, C.M. (2004) Dual stable isotope analysis(delta C-13 and delta N-15) of soil invertebrates andtheir food source. Pedobiologia 48: 171-180.
Schulz, H.N., and Jorgensen, B.B. (2001) Big bacteria.Annu Rev Microbiol 55: 105-137.
Sextone, A.J., Revbesh, N.P., Parkin, T.B., and Tiedje,J.M. (1985) Direct measurement of oxygen profilesand denitrification rates in soil aggregates. Soil Sci.Soc. Am. J. 49: 645-651.
Sherr, E.B., and Sherr, B.F. (1988) Role of microbes inpelagic food webs: A revised concept. LimnolOceanogr 33: 1225-1227.
Sieburth, J. Mc.N., Smetacek, V., and Lenz, J. (1978)Pelagic ecosystem structure: heterotrophiccompartments of the plankton and their relationship toplankton size fractions. Limnol. Oceanogr 23: 1256-1263.
Sime-Ngando, T. (1997). Importance des virus dans lastructure et le fonctionnement des réseaux trophiquesmicrobiens aquatiques. Ann. Biol 36: 181-210.
Sime-Ngando, T., Bettarel, Y., Chartogne, C., andSean, K. (2003) The imprint of wild viruses onfreshwater microbial ecology. Recent Res. Devel.Microbiology 7: 481-497.
Suttle, C.A. (2005) Viruses in the sea. Nature 437:356-361.
Suttle, C.A. (2007) Marine viruses – major players inthe global ecosystem. Nature Reviews 5: 801-812.
Swift, M.J., Heal, O.W., and Anderson, J.M. (1979)Decomposition in terrestrial ecosystems. University ofCalifornia Press, Berkeley.
Thingstad, T.F. (2000) Element of a theory for themechanisms controlling abundance, diversity, andbiogeochemical role of lytic bacteria viruses in aquaticsystems. Limnol Oceanogr 45: 1320-1328.
Torsvik, V., Ovreas, L., and Thingstad, T.F. (2002)Prokariotic diversity: magnitude, dynamics and
28
PART I E 4 – RÔ LE E T FONC T I ONNEMEN T DES ÉCOSYS T ÈMES M I CROB I E N S

controlling factors. Science 296: 1064-1066.
Traunspurger, W., Bergtold, M., and Goedkoop, W.(1997) The effects of nematodes on bacterial activityand abundance in a freshwater sediment. Oecologia112: 118-122.
Verity, P.G. (1991) Feeding in planktonic protozoans –evidence for non-random acquisition of prey. Journalof Protozoology 38: 69-76.
Wardle, D.A. (2002) Linking the aboveground andbelowground components. Princeton, PrincetonUniversity Press.
Weinbauer, M.G. (2004) Ecology of prokaryoticviruses. FEMS microb Rev 28: 127-181.
Weinbauer, M.G., and Rassoulzadegan, F. (2004) Areviruses driving microbial diversification and diversity?Environ Microbiol 6: 1-11.
Wetzel, R.G., Rich, P.H., Miller, M.C., and Allen, H.L.
(1972) Metabolism of dissolved and particulate detritalcarbon in a temperate hard-water lake. In: Melchiorri-Santolini, U., and Hopton, J.W. (Ed.) Detritus and itsrole in aquatic ecosystems. Memoire Inst. Ital. Idrobiol29: 185-243.
Yeates, G.W., Bardgett, R.D., Cook, R., Hobbs, P.J.,Bowling, P.J., and Potter, J.F. (1997) Faunal andmicrobial diversity in three Welsh grassland soils underconventional and organic managements regimes.Journal of Applied Ecology 34: 453-471.
Yeates, G.W., and Bongers, T. (1999) Nematodebiodiversity in agroecosystems. Agriculture,Ecosystems and Environment 74: 113-135.
Yeates, G.W., Bongers, T., De Groede, R.G.M.,Freckman, D.W., and Georgieva, S.S. (1993) Feedinghabits in soil nematode families and genera – an outlinefor ecologists. Journal on Nematology 25: 315-331.
29
L ES RÉSEAUX TROPH I QUES M I CROB I E N S DES M I L I E U X AQUAT I QUES E T T ERRES TRES





























