Embed Size (px)
Citation preview

La relation Structure Dynamique Fonctions
Ex 1 : Mouvement de chaînes latérales pour la fonction de la Myoglobine
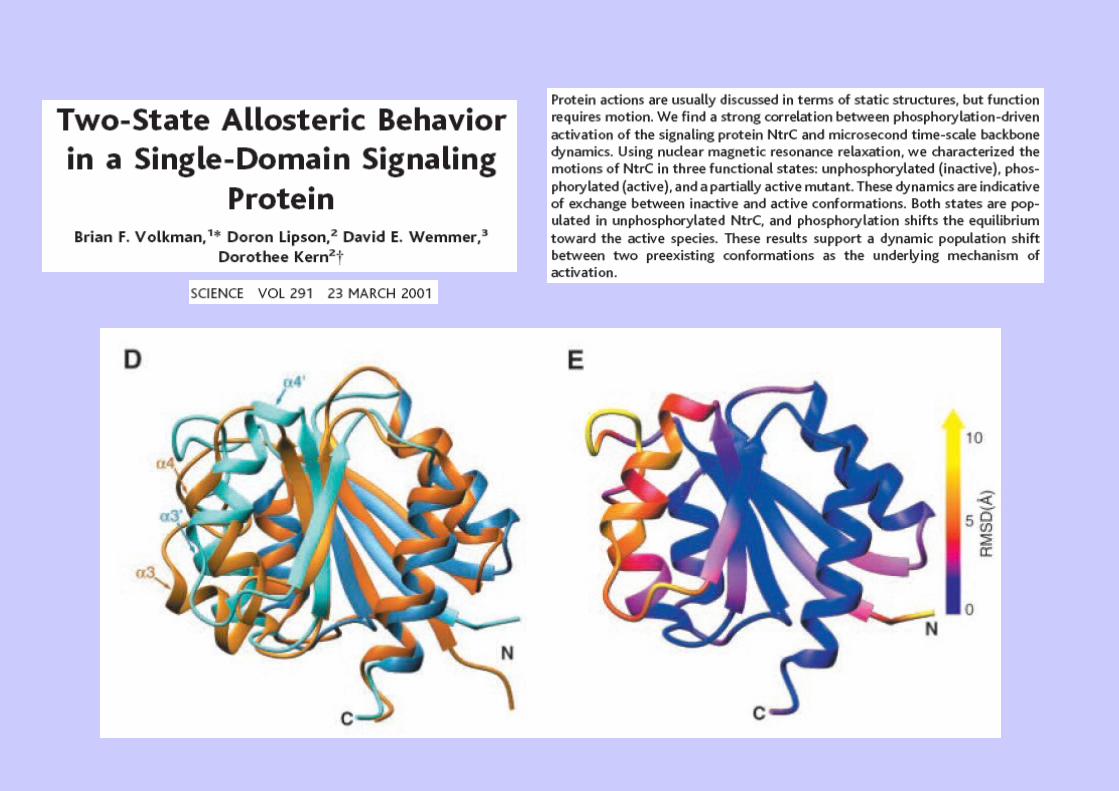
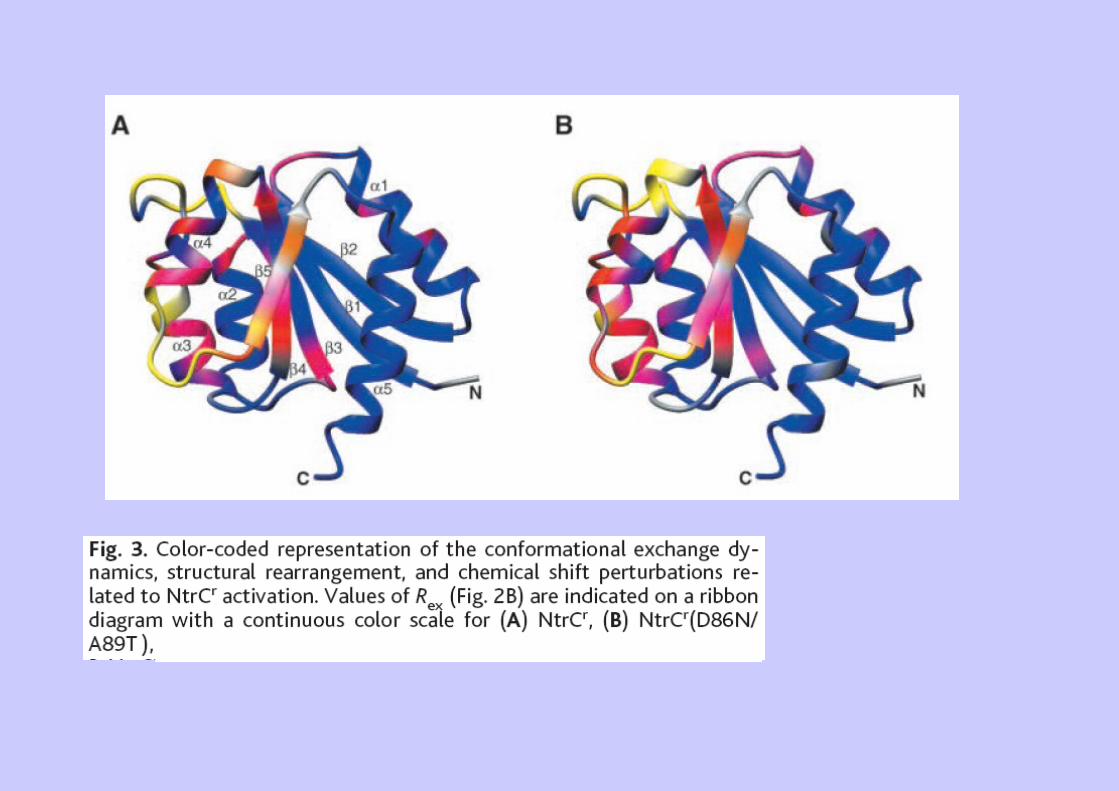
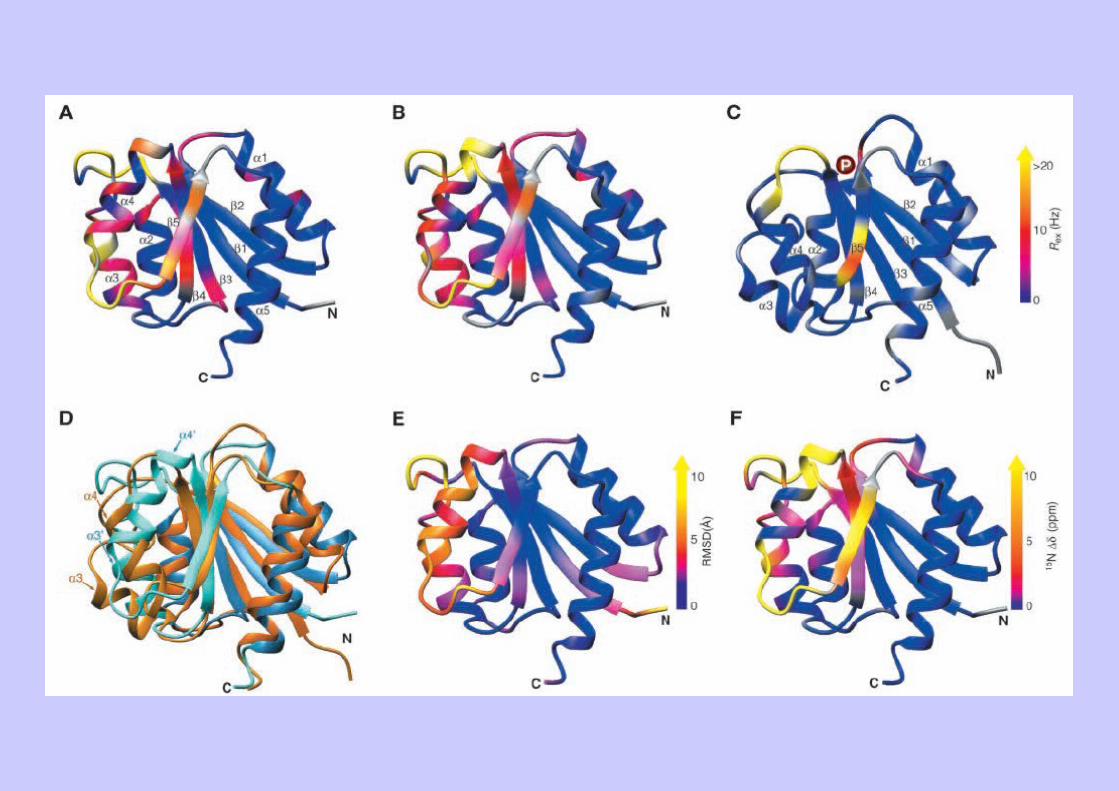
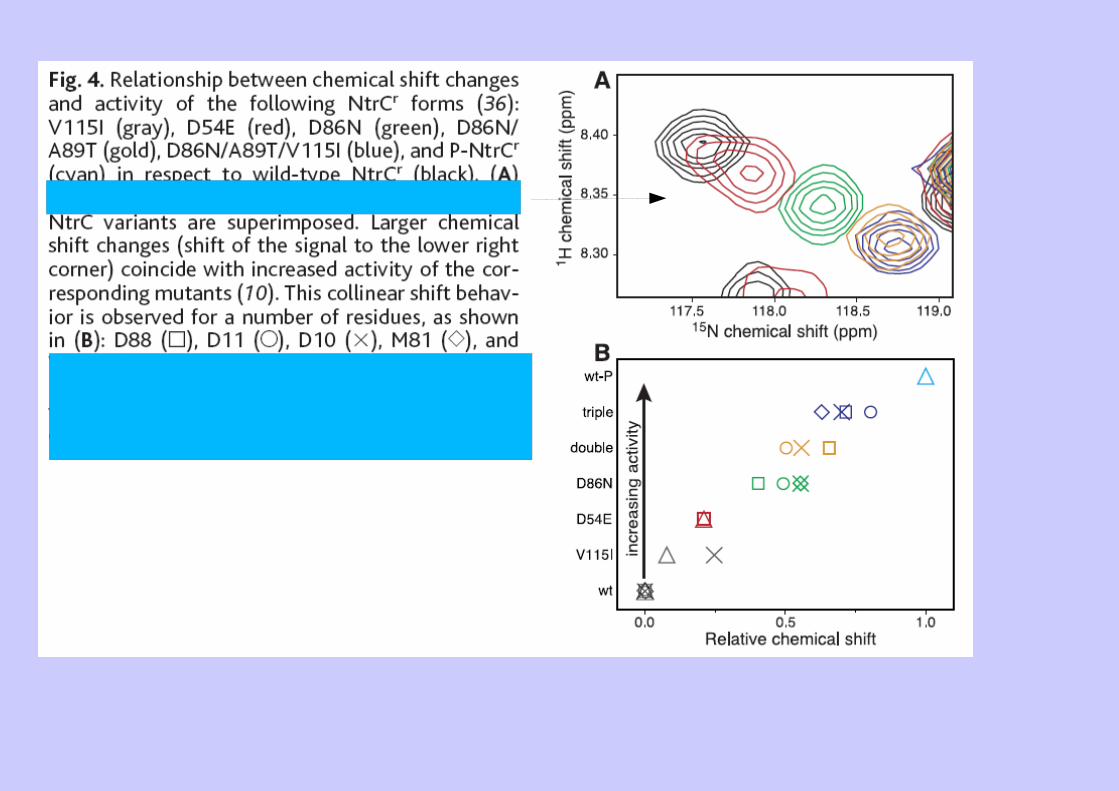
Ex 2 : Dynamique et équilibres allostériques (NtrC)
Ex 3 : La fonction enzymatique de Purine nucléotide phosphorylase et de la Thymidilate Synthase
Ex 4 : une protéine plusieurs structures : la base de données MolMovdb
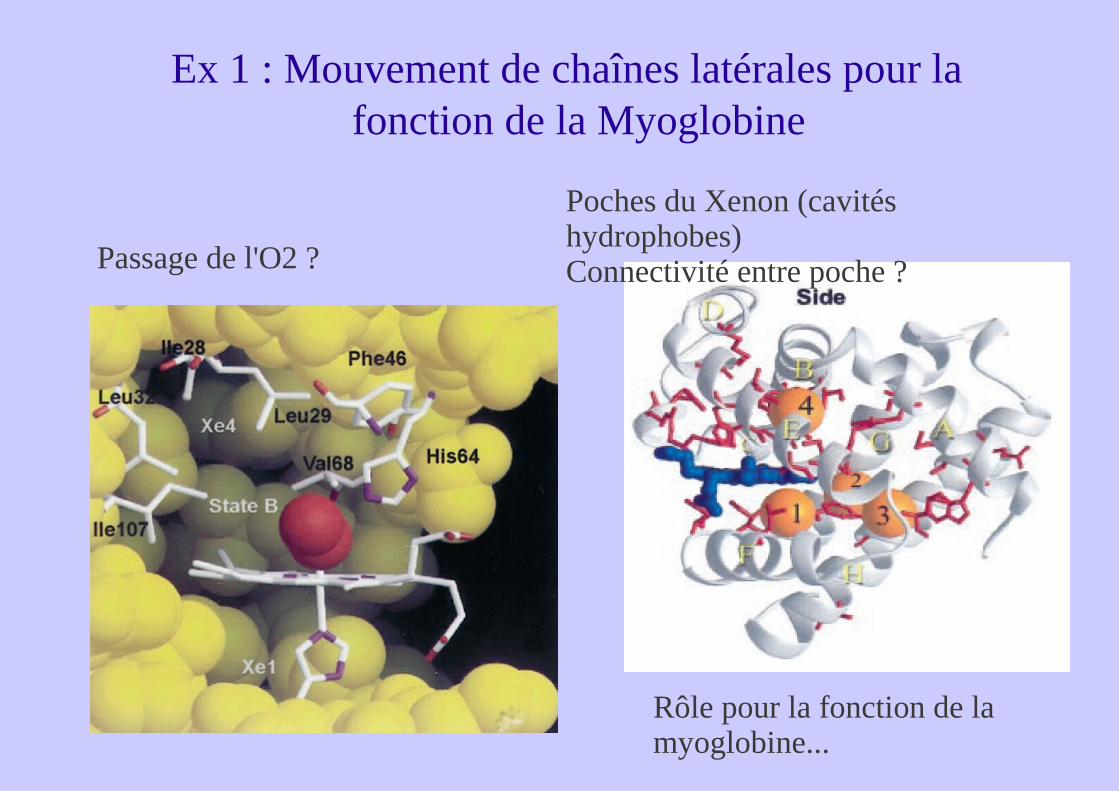
Ex 1 : Mouvement de chaînes latérales pour la fonction de la Myoglobine
Passage de l'O2 ?
Rôle pour la fonction de la myoglobine...
Poches du Xenon (cavités hydrophobes)Connectivité entre poche ?
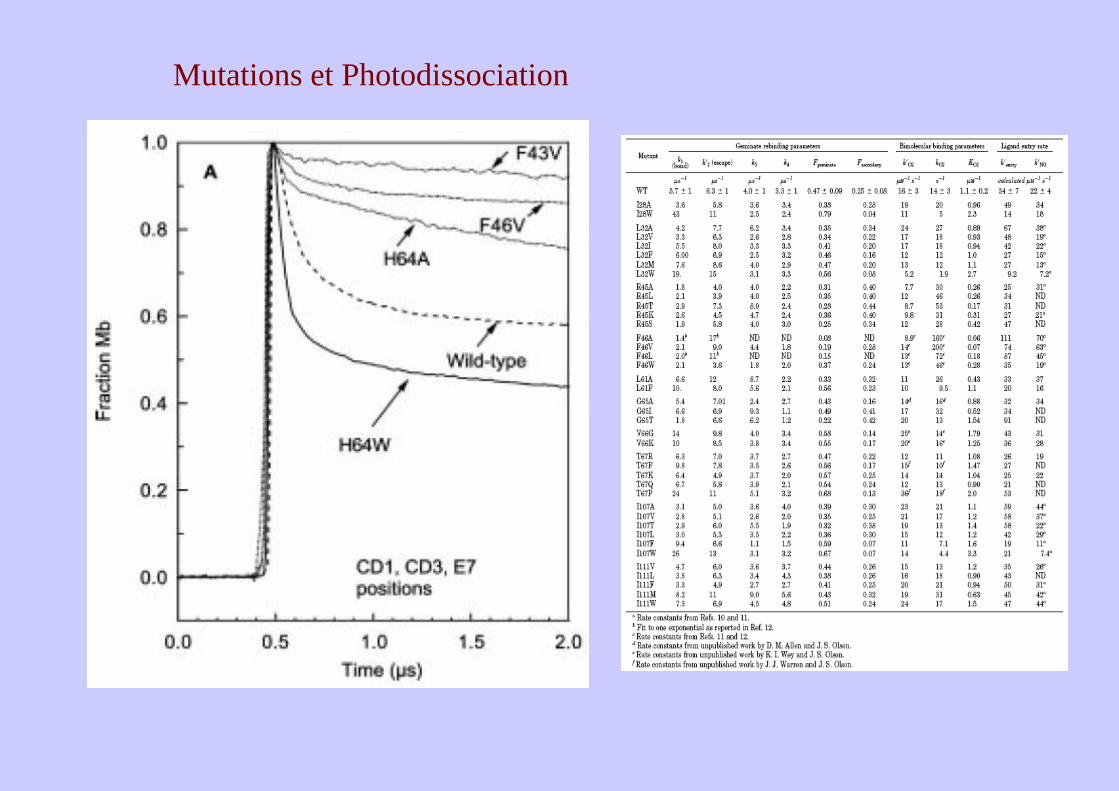
Mutations et Photodissociation
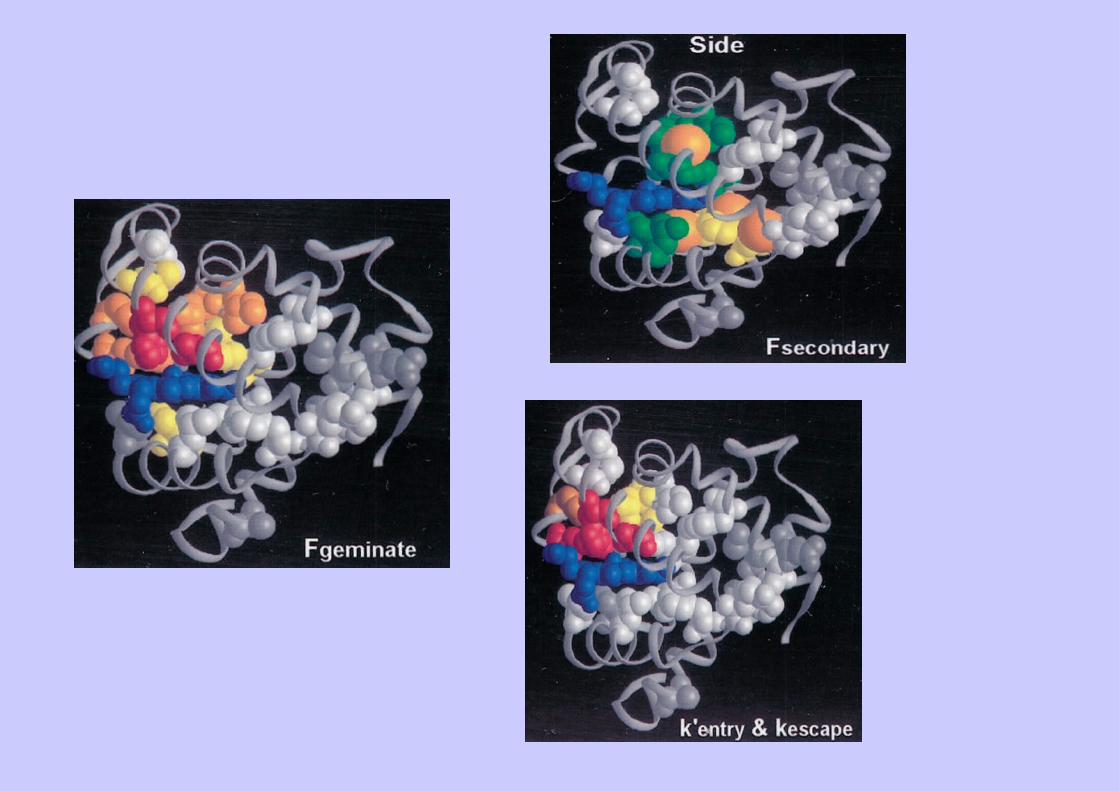
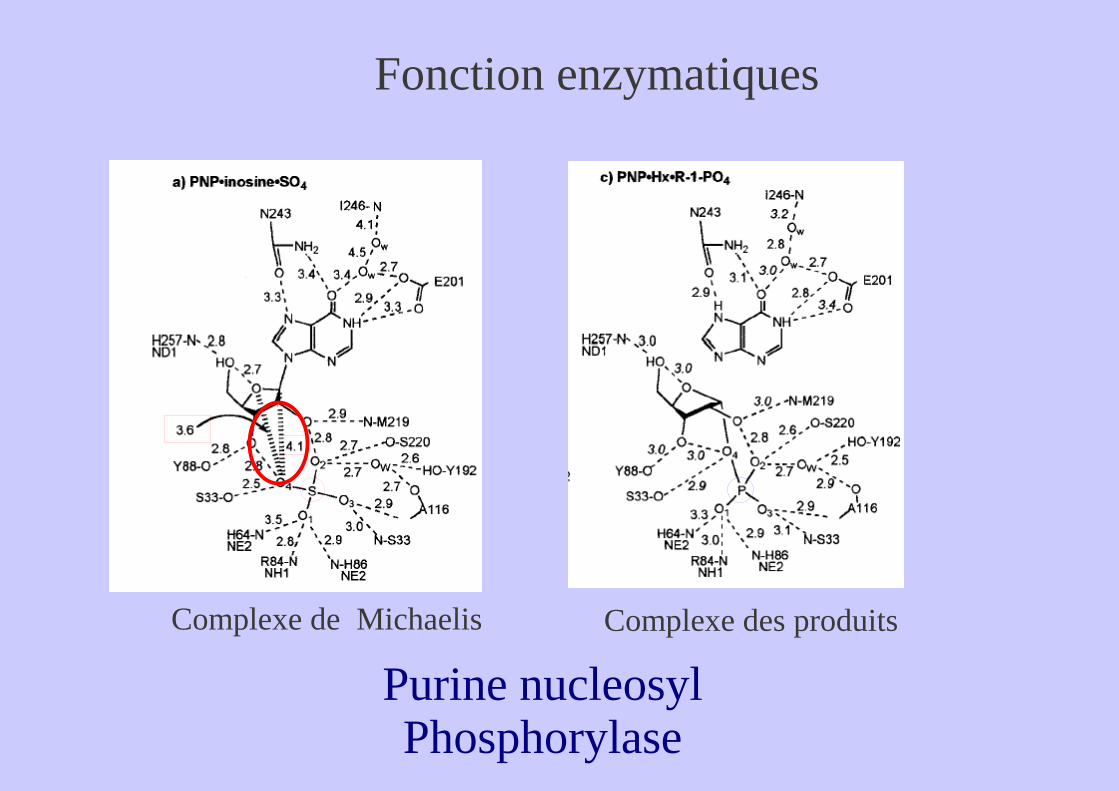
Complexe de Michaelis Complexe des produits
Purine nucleosyl Phosphorylase
Fonction enzymatiques
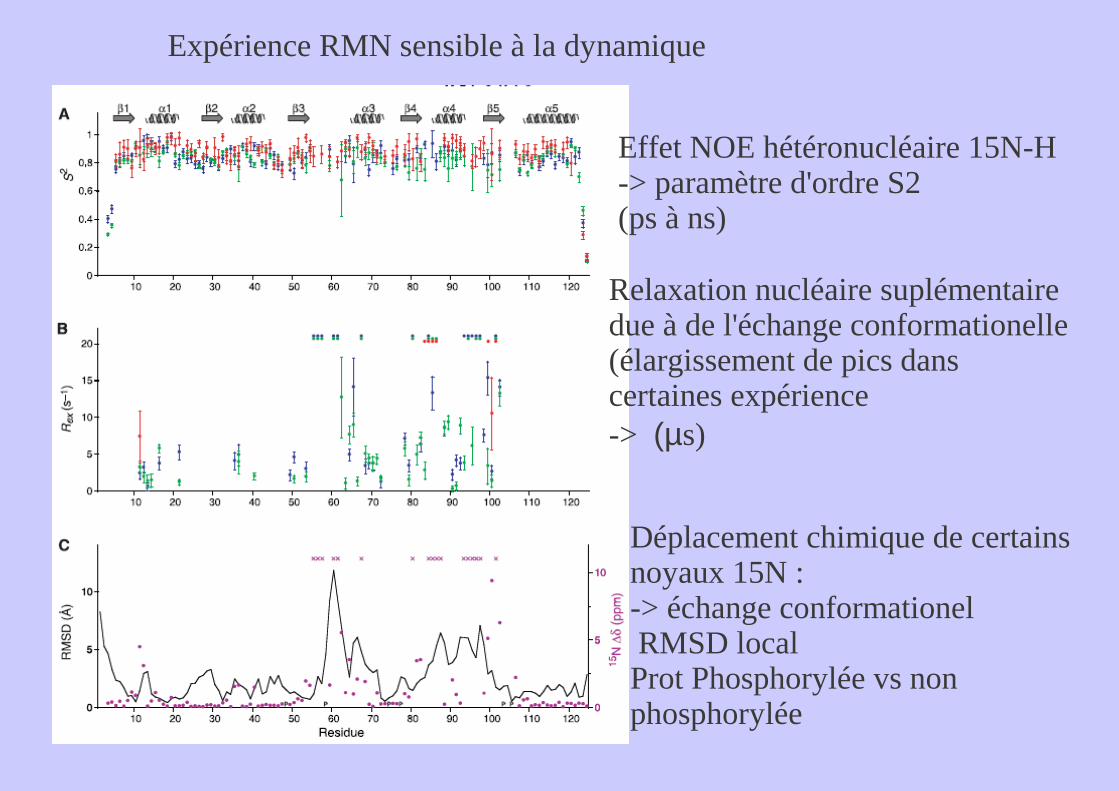
Expérience RMN sensible à la dynamique
Effet NOE hétéronucléaire 15N-H-> paramètre d'ordre S2(ps à ns)
Relaxation nucléaire suplémentaire due à de l'échange conformationelle (élargissement de pics dans certaines expérience-> (µs)
Déplacement chimique de certains noyaux 15N : -> échange conformationel RMSD local Prot Phosphorylée vs non phosphorylée
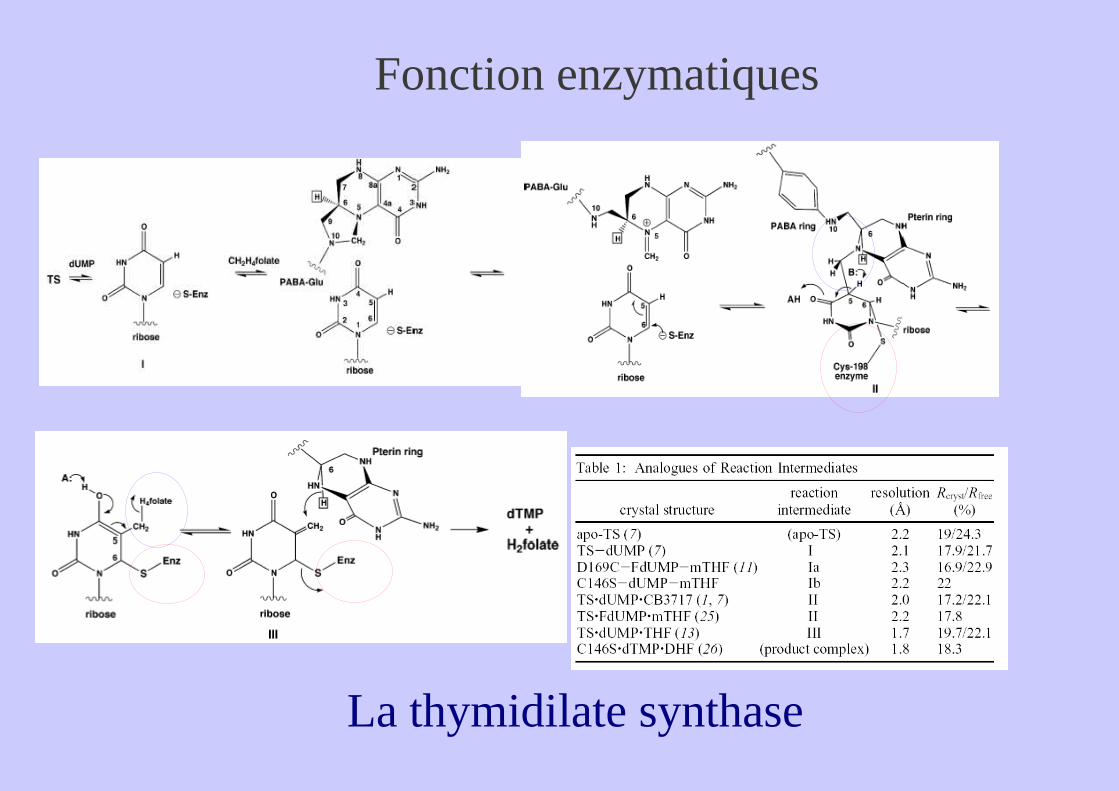
La thymidilate synthase
Fonction enzymatiques

Le Film
La thymidilate synthase
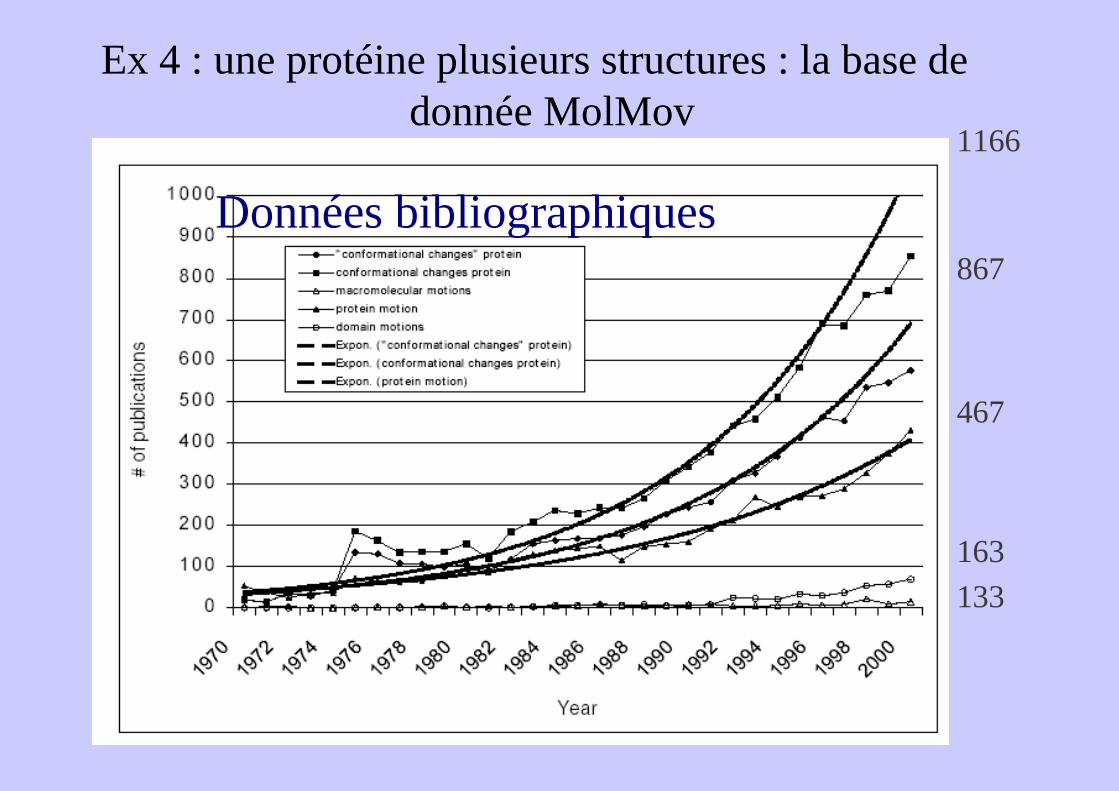
Ex 4 : une protéine plusieurs structures : la base de donnée MolMov
Données bibliographiques
467
867
163
133
1166

MolMovDB ...
http://molmovdb.mbb.yale.edu/cgi-bin/browse.cgi
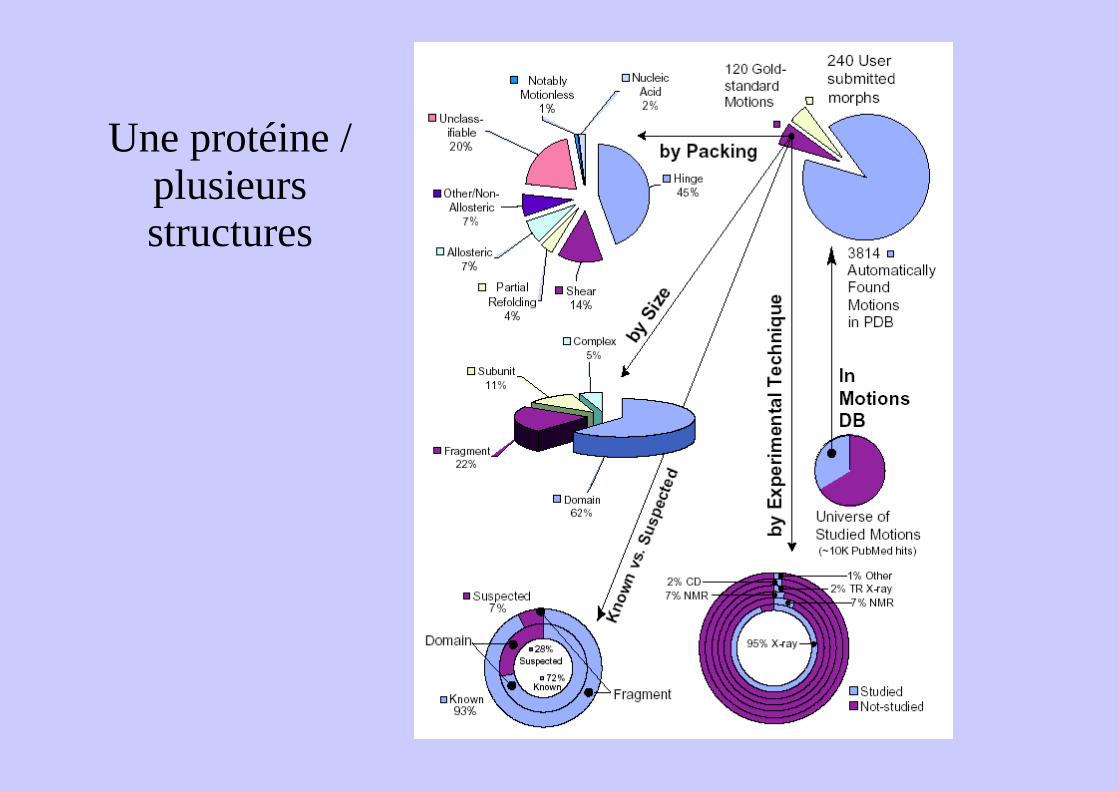
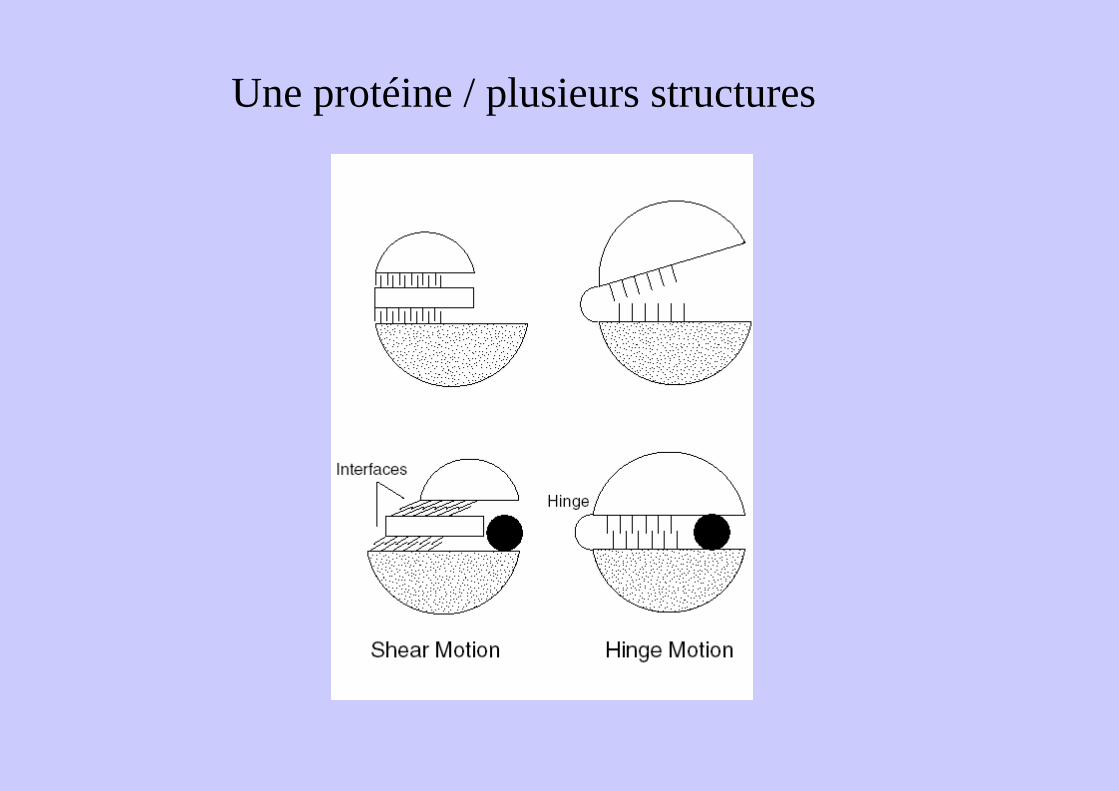
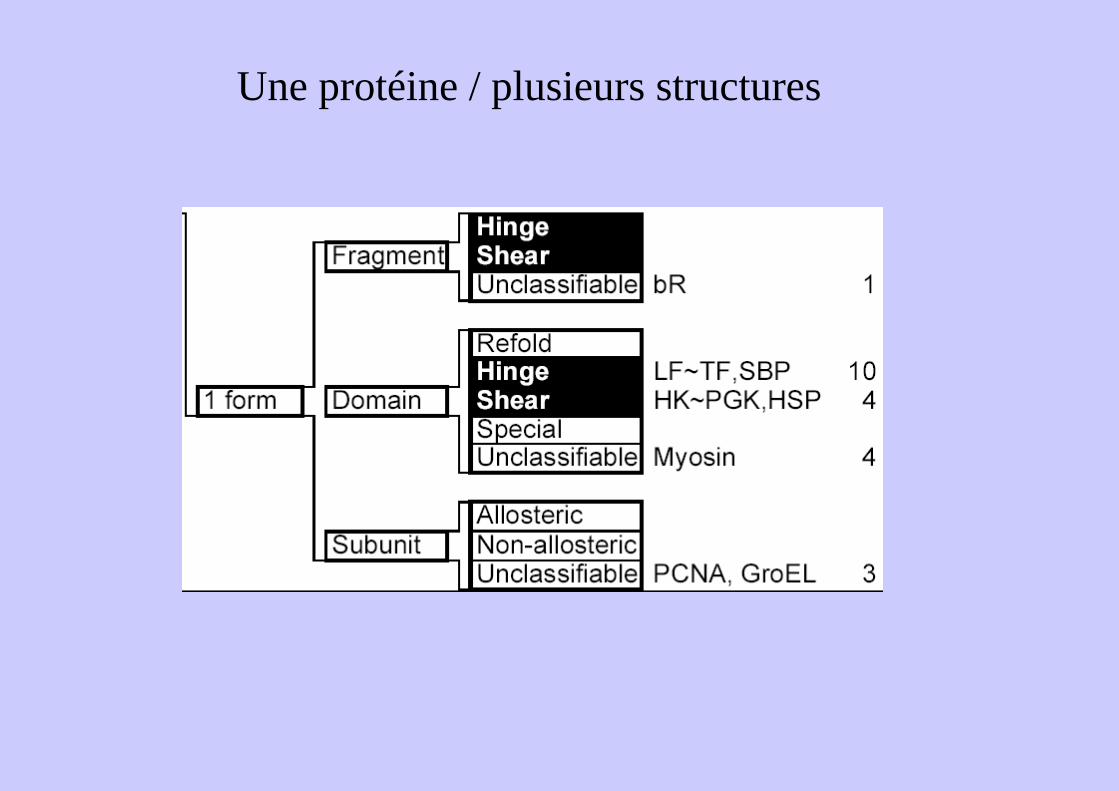
Une protéine / plusieurs structures

Une protéine / plusieurs structuresGlutamine Binding Protein
Mécanisme de charnière (hinge)Quelques liens interdomaine30° de rotationGrands mouvements sur très peu
d'angles de tortion

Une protéine / plusieurs structuresGroEL
Mécanisme de charnière

Aspartate Amino TransferaseShear motion at 2 interfaces combined with hinge in a kinked helix. The domain motion is mainly the result of just two shear motions, which occur in perpendicular directions. AAT has an active site situated between a large and a small domain, and on substrate binding the small domain closes over the active site. The major shear motion involves a 13 degree rotation of the the small domain relative to the large one. A secondary shear motion moves a helix on one side of the small domain in a direction perpendicular to both the interdomain interface and the direction of the of the other shear motion. With a 1.2 A shift and a 10 degree rotation, it drops down to cover the active site. The shear motions in AAT are facilitated by a hinge motion in a long interdomain helix. This helix is kinked by 17 degrees in the open form and changes its kink angle by 12 degrees on closure.
Mécanisme de cisaillement (shear)

Une protéine / plusieurs structuresDNA polymerase β

Hémoglobine
Hemoglobin is a tetramer containing 2 copies of 2 (slightly) different subunits (α and β).
● When oxygen binds to the deoxy form (the T-state), the heme ring slightly flattens. ● This causes one helix (EF turn, F helix, FG turn) to move, ● This, in turn, destabilizes salt bridges between this helix and a helix on another subunit. ● A new inter-subunit salt bridge configuration is formed (with in total fewer bridges). ● This switch of configurations at the subunit interface causes one pair of subunits (e.g. α-1
and β-1) to rotate 15° and translate 0.8 A relative to the other pair (producing the high-affinity R-state).
● This, in turn, affects the interactions about the heme in other subunits, giving rise to cooperative oxygen binding.
α subunit

Une protéine ... plusieurs structures ?Normal Mode Analysis
Méthode théorique :La protéine est considérée comme un ressort ...Quelles sont les zones qui « vibrent » le plus ?
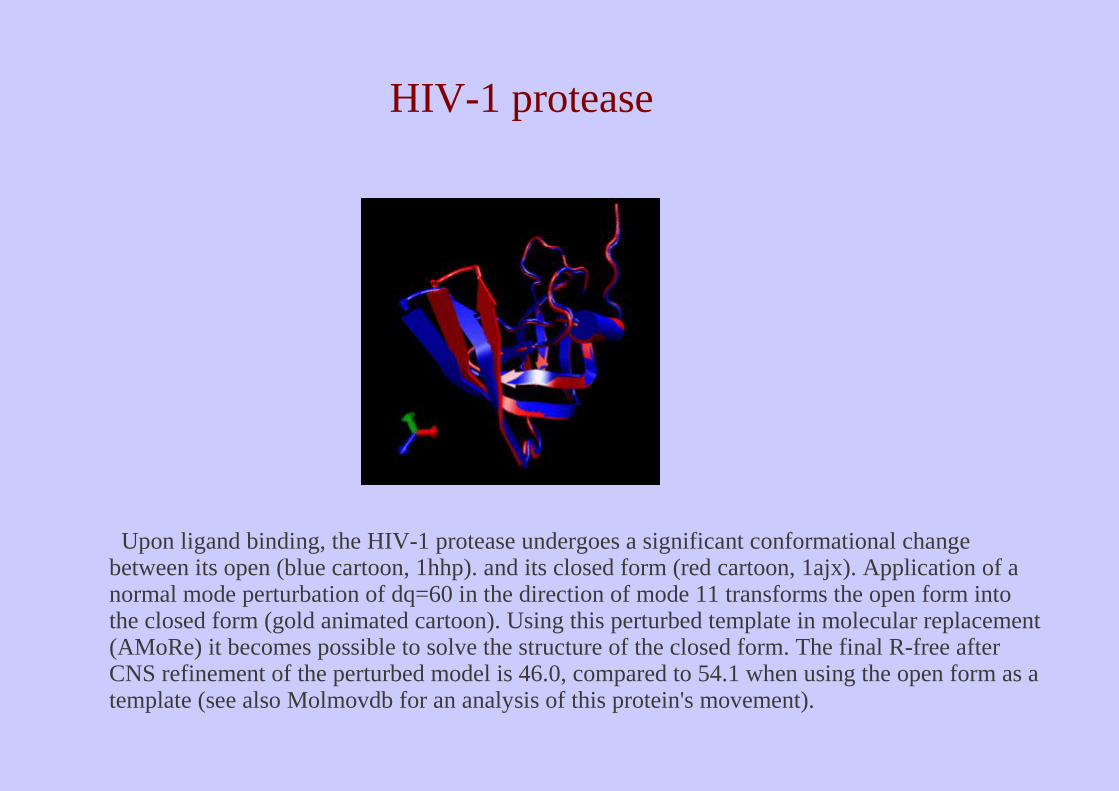
Upon ligand binding, the HIV-1 protease undergoes a significant conformational change between its open (blue cartoon, 1hhp). and its closed form (red cartoon, 1ajx). Application of a normal mode perturbation of dq=60 in the direction of mode 11 transforms the open form into the closed form (gold animated cartoon). Using this perturbed template in molecular replacement (AMoRe) it becomes possible to solve the structure of the closed form. The final R-free after CNS refinement of the perturbed model is 46.0, compared to 54.1 when using the open form as a template (see also Molmovdb for an analysis of this protein's movement).
HIV-1 protease
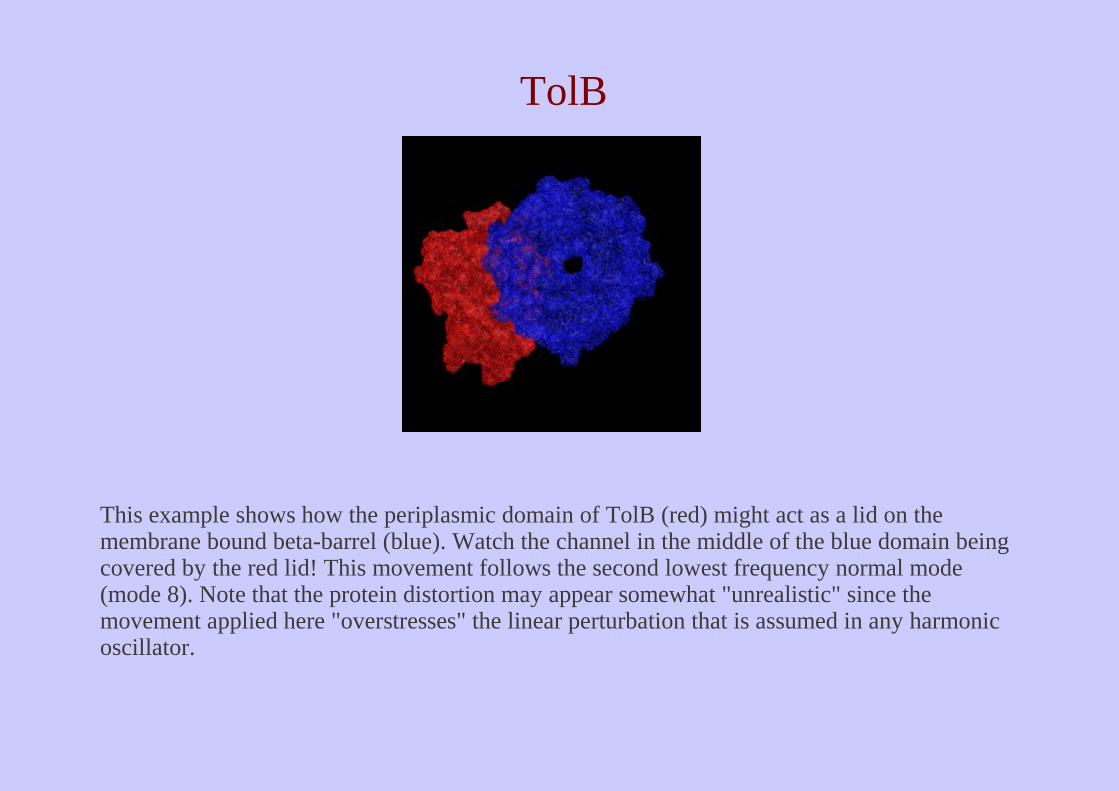
This example shows how the periplasmic domain of TolB (red) might act as a lid on the membrane bound beta-barrel (blue). Watch the channel in the middle of the blue domain being covered by the red lid! This movement follows the second lowest frequency normal mode (mode 8). Note that the protein distortion may appear somewhat "unrealistic" since the movement applied here "overstresses" the linear perturbation that is assumed in any harmonic oscillator.
TolB
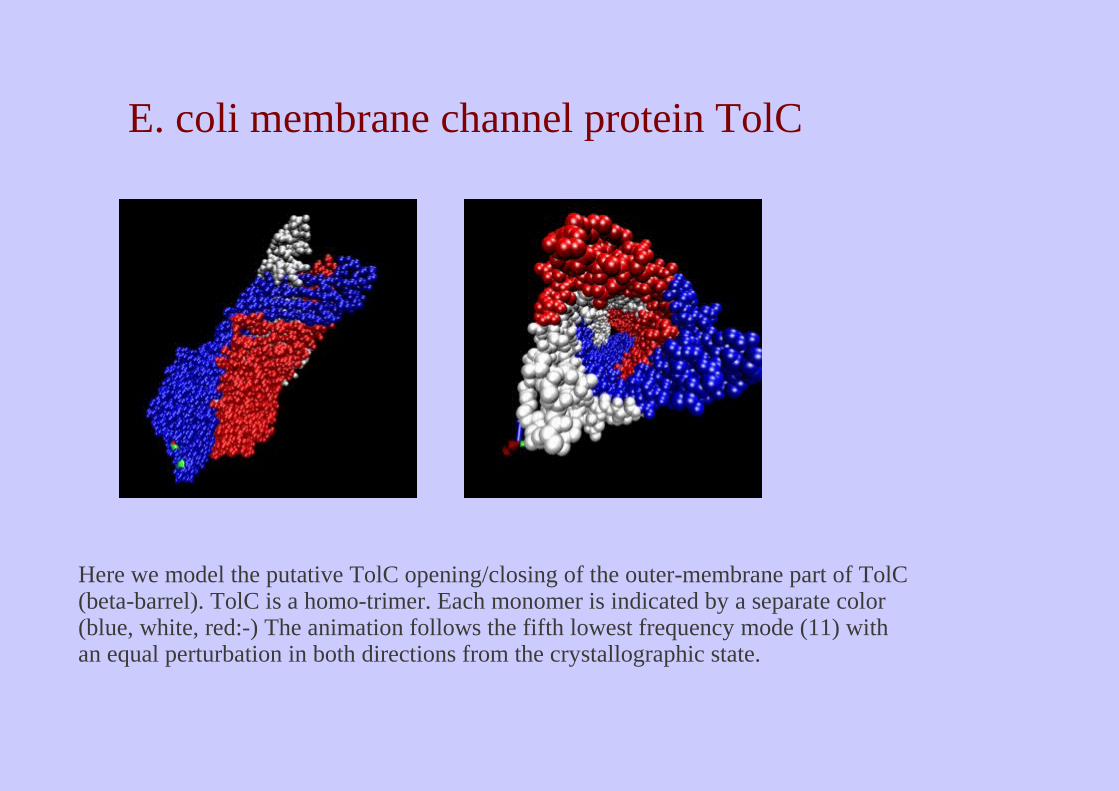
Here we model the putative TolC opening/closing of the outer-membrane part of TolC (beta-barrel). TolC is a homo-trimer. Each monomer is indicated by a separate color (blue, white, red:-) The animation follows the fifth lowest frequency mode (11) with an equal perturbation in both directions from the crystallographic state.
E. coli membrane channel protein TolC
Une protéine / plusieurs structures

Une protéine / plusieurs structures
Une protéine / plusieurs structures

Comment bougent les macromolécules biologique ?
Ex 1 : Simulation de la dynamique moléculaire de l'ADN
Ex2 : Insertion d'un peptides hydrophobe dans un environnement mimétique membranaire
Ex 3 : Passage d'un hexose au travers d'une porine membranaire
Ex 4 :Agrégation spontanée d'un détergent en micelle




![LES FONCTIONS DE RELATION ET L’HOMÉOSTASIE [1] COMMENT … · COMMENT SE RÉGULE L ... la régulation de ces fonctions biologiques dépend d’au moins 4 ... mais la spécificité](https://img.dokumen.tips/doc/110x75/5b9a82f309d3f2c3468d682b/les-fonctions-de-relation-et-lhomeostasie-1-comment-comment-se-regule.jpg)
























