Embed Size (px)
Citation preview

Kinesin-2 family in vertebrate ciliogenesisChengtian Zhaoa, Yoshihiro Omorib, Katarzyna Brodowskaa, Peter Kovacha, and Jarema Malickia,1
aDivision of Craniofacial and Molecular Genetics and Program in Genetics, Sackler School of Graduate Biomedical Studies, Tufts University, Boston, MA 02111;and bDepartment of Developmental Biology, Osaka Bioscience Institute, Osaka 565-0874, Japan
Edited by Kathryn V. Anderson, Sloan–Kettering Institute, New York, NY, and approved December 28, 2011 (received for review October 12, 2011)
The differentiation of cilia is mediated by kinesin-driven transport.As the function of kinesins in vertebrate ciliogenesis is poorlycharacterized, we decided to determine the role of kinesin-2 familymotors—heterotrimeric kinesin-II and the homodimeric Kif17 kine-sin—in zebrafish cilia. We report that kif17 is largely dispensablefor ciliogenesis; kif17 homozygous mutant animals are viable anddisplay subtle morphological defects of olfactory cilia only. In con-trast to that, the kif3b gene, encoding a heterotrimeric kinesinsubunit, is necessary for cilia differentiation in most tissues, al-though exceptions exist, and include photoreceptors and a subsetof hair cells. Cilia of these cell types persist even in kif3b/kif17double mutants. Although we have not observed a functional re-dundancy of kif3b and kif17, kif17 is able to substitute for kif3b insome cilia. In contrast to kif3b/kif17 double mutants, simultaneousinterference with kif3b and kif3c leads to the complete loss ofphotoreceptor and hair cell cilia, revealing redundancy of function.This is in agreement with the idea that Kif3b and Kif3c motorsubunits form complexes with Kif3a, but not with each other. In-terestingly, kif3b mutant photoreceptor cilia differentiate witha delay, suggesting that kif3c, although redundant with kif3b atlater stages of differentiation, is not active early in photoreceptorciliogenesis. Consistent with that, the overexpression of kif3c inkif3bmutants rescues early photoreceptor cilia defects. These datareveal unexpected diversity of functional relationships betweenvertebrate ciliary kinesins, and show that the repertoire of kinesinmotors changes in some cilia during their differentiation.
intraflagellar transport | opsin | outer segment | ciliary axoneme
Cilia and flagella are thin elongated cell surface protrusions thatperform diverse biological functions. Motile cilia or flagella of
protozoans or sperm cells facilitate the movement of entire cells.In multicellular organisms, motile cilia drive fluid flow in the lu-men of ducts and chambers such as kidney tubules or brain ven-tricles (1, 2). Apart from its ability to generatemotion, the cilium isa separate subcellular compartment that features cell surface andcytoplasmic components. These characteristics provide basis forthe ability of cilia to mediate signal transduction in a variety ofcontexts, including developmental signaling by the hedgehog ornotch pathways, as well as sensory signal transduction.As the cilium itself is devoid of protein synthesis, polypeptides
that contribute to the structure and function of cilia are trans-ported from the cell body. This transport, referred to as theintraflagellar transport (IFT), relies on the function of two typesof plus-end directed motors that are members of the kinesin-2family (3), named heterotrimeric kinesin-II and homodimericOSM-3 or KIF17 (4–7). In the flagella of algae Chlamydomonas,mainly kinesin-II is thought to function in IFT (6). In Caeno-rhabditis elegans, subunits KLP-20, KLP-11, and KAP-1 form thesingle kinesin-II complex, which functions in parallel to homo-dimeric OSM-3 (8). Vertebrates, on the contrary, are thought toform two kinesin-II complexes (KIF3A/3B/KAP3 and KIF3A/3C/KAP3) in addition to homodimeric KIF17 (9, 10). Bothkinesin-II and kif17 have been proposed to function in vertebrateciliogenesis (11–13).Given the diversity of cilia-mediated processes in vertebrate
tissues, it appears that kinesins may function differently in cilia ofdifferent cells. The analysis of this issue is complicated, however,because kinesin-II mouse knock-outs (KOs) lead to midgestation
lethality (12, 13). Consequently, conditional KOs of Kif3a havebeen performed in several organs, including kidney tubules, pan-creas, and skin, and in all cases resulted in a loss of cilia, demon-strating that Kif3a is necessary for ciliogenesis (14–16). As thevertebrate photoreceptor outer segment is among the best char-acterized cilia-derived structures (17, 18), several studies focusedon the role of kinesins in photoreceptor ciliogenesis. A conditionalKO of Kif3a in photoreceptor cells revealed a role for the het-erotrimeric kinesin II in the transport of opsin and arrestin, butnot transducin or peripherin, suggesting the presence of multipleanterograde ciliary motor mechanisms (19). Rod and conephotoreceptors appear to have different requirements for motorproteins: following conditional KOs of Kif3a specifically in rodsor in cones, rods degenerate much faster compared with cones(20). Given these results, it appears likely that multiple kinesinscontribute to photoreceptor ciliogenesis.Based on biochemical studies, Kif3a functions as a hetero-
trimeric complex, consisting of another motor subunit, eitherKif3b or Kif3c, and an accessory protein, Kap3 (9, 10, 21).Similar to kif3a, the role of kif3b in different tissues is difficult toevaluate because homozygous mouse mutants die at midg-estation, and few, if any, conditional KOs are available (13). Therole of kif3c in ciliogenesis also remained obscure, as mouse KOsof this gene do not display any obvious phenotype (22, 23). Thecontribution of kif17 to vertebrate ciliogenesis is even less clearas its studies produced contradictory results (11, 24).By using zebrafish as a model system, we show that vertebrate
kif3b and kif17 kinesins are required for the formation of dif-ferent subsets of cilia. Although the loss of kif3b function affectsthe majority of cilia, cone photoreceptor cilia and a subset ofkinocilia in the otic vesicle do not require kif3b function. Incontrast to that, among tissues analyzed so far, kif17 function isconfined to the morphogenesis of olfactory cilia. Consistent withbiochemical studies, kif3b acts redundantly with kif3c in photo-receptor and some hair cell cilia. Interestingly, the repertoire ofkinesins that function in photoreceptor cilia changes during de-velopment; whereas kif3b alone appears to drive ciliogenesisearly on, kif3b and kif3c function redundantly at later stagesof differentiation.
Resultsjj203 Mutant Embryos Display Cilia Defects. We identified the jj203mutant strain following N-ethyl-N-nitrosourea (ENU) muta-genesis. The most obvious external phenotype of jj203 is curvedbody axis (Fig. 1B). As such a phenotype is characteristic of ciliamutants (25, 26), we investigated ciliogenesis in jj203. Thezebrafish embryo and larva feature a number of well character-ized ciliated organs (27). Similar to defects in ovl and elipsa loci,which affect IFT particle components, the jj203 mutation resultsin a shortening or absence of cilia in the olfactory pit (Fig. 1 C
Author contributions: C.Z. and J.M. designed research; C.Z., Y.O., K.B., and P.K. performedresearch; C.Z. and J.M. analyzed data; and C.Z. and J.M. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence should be addressed: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1116035109/-/DCSupplemental.
2388–2393 | PNAS | February 14, 2012 | vol. 109 | no. 7 www.pnas.org/cgi/doi/10.1073/pnas.1116035109
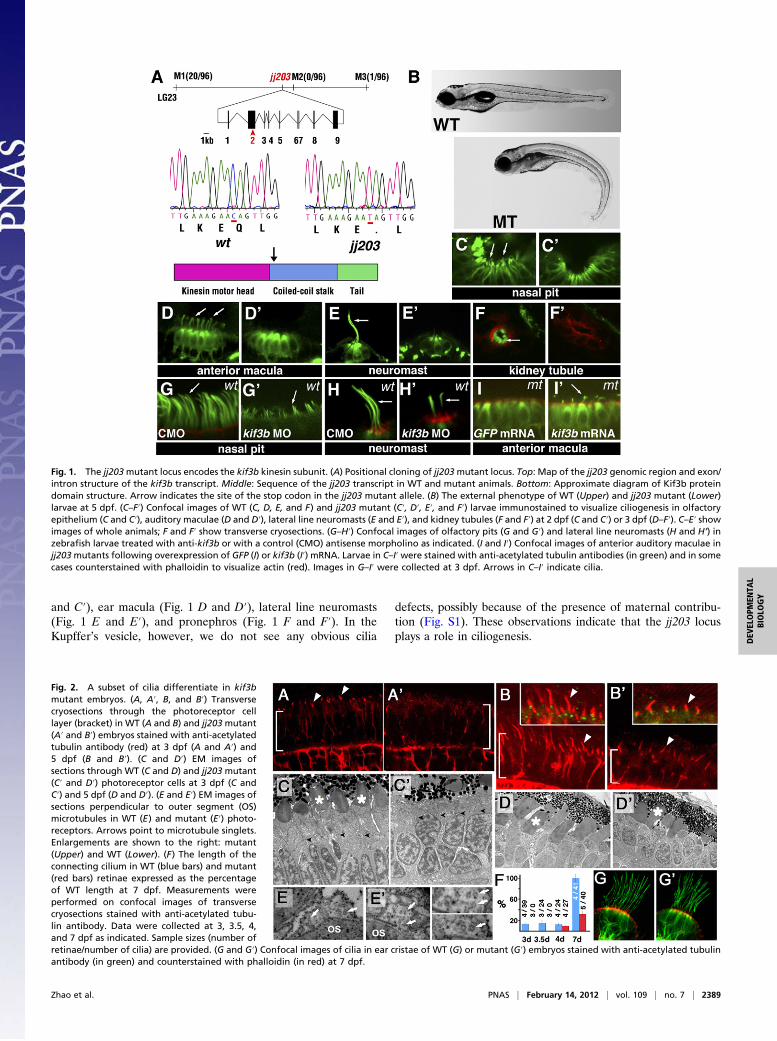
and C′), ear macula (Fig. 1 D and D′), lateral line neuromasts(Fig. 1 E and E′), and pronephros (Fig. 1 F and F′). In theKupffer’s vesicle, however, we do not see any obvious cilia
defects, possibly because of the presence of maternal contribu-tion (Fig. S1). These observations indicate that the jj203 locusplays a role in ciliogenesis.
Fig. 1. The jj203mutant locus encodes the kif3b kinesin subunit. (A) Positional cloning of jj203mutant locus. Top: Map of the jj203 genomic region and exon/intron structure of the kif3b transcript. Middle: Sequence of the jj203 transcript in WT and mutant animals. Bottom: Approximate diagram of Kif3b proteindomain structure. Arrow indicates the site of the stop codon in the jj203 mutant allele. (B) The external phenotype of WT (Upper) and jj203 mutant (Lower)larvae at 5 dpf. (C–F′) Confocal images of WT (C, D, E, and F) and jj203 mutant (C′, D′, E′, and F′) larvae immunostained to visualize ciliogenesis in olfactoryepithelium (C and C′), auditory maculae (D and D′), lateral line neuromasts (E and E′), and kidney tubules (F and F′) at 2 dpf (C and C′) or 3 dpf (D–F′). C–E′ showimages of whole animals; F and F′ show transverse cryosections. (G–H′) Confocal images of olfactory pits (G and G′) and lateral line neuromasts (H and H’) inzebrafish larvae treated with anti-kif3b or with a control (CMO) antisense morpholino as indicated. (I and I′) Confocal images of anterior auditory maculae injj203mutants following overexpression of GFP (I) or kif3b (I′) mRNA. Larvae in C–I′were stained with anti-acetylated tubulin antibodies (in green) and in somecases counterstained with phalloidin to visualize actin (red). Images in G–I′ were collected at 3 dpf. Arrows in C–I′ indicate cilia.
Fig. 2. A subset of cilia differentiate in kif3bmutant embryos. (A, A′, B, and B′) Transversecryosections through the photoreceptor celllayer (bracket) in WT (A and B) and jj203 mutant(A′ and B′) embryos stained with anti-acetylatedtubulin antibody (red) at 3 dpf (A and A′) and5 dpf (B and B′). (C and D′) EM images ofsections through WT (C and D) and jj203 mutant(C′ and D′) photoreceptor cells at 3 dpf (C andC′) and 5 dpf (D and D′). (E and E′) EM images ofsections perpendicular to outer segment (OS)microtubules in WT (E) and mutant (E′) photo-receptors. Arrows point to microtubule singlets.Enlargements are shown to the right: mutant(Upper) and WT (Lower). (F) The length of theconnecting cilium in WT (blue bars) and mutant(red bars) retinae expressed as the percentageof WT length at 7 dpf. Measurements wereperformed on confocal images of transversecryosections stained with anti-acetylated tubu-lin antibody. Data were collected at 3, 3.5, 4,and 7 dpf as indicated. Sample sizes (number ofretinae/number of cilia) are provided. (G and G′) Confocal images of cilia in ear cristae of WT (G) or mutant (G′) embryos stained with anti-acetylated tubulinantibody (in green) and counterstained with phalloidin (in red) at 7 dpf.
Zhao et al. PNAS | February 14, 2012 | vol. 109 | no. 7 | 2389
DEV
ELOPM
ENTA
LBIOLO
GY

jj203 Encodes Kif3b Subunit of Heterotrimeric Kinesin II. To de-termine the molecular nature of the jj203 locus, we performedpositional cloning (Fig. 1A). This effort revealed a single nu-cleotide substitution (nucleotide 1,105, C to T) in the secondexon of the kif3b gene, which introduces a nonsense codon in theplace of a glutamine at the position 369 of the polypeptide (Fig.1A). The truncated mutant protein lacks the coiled-coil stalkdomain and the C-terminal tail domain (Fig. 1A). To confirmthat defects in the kif3b gene are responsible for the jj203 phe-notype, we performed a knockdown with two morpholinos tar-geted against this gene. This treatment produces a curly bodyaxis phenotype by 3 d postfertilization (dpf), which closelyresembles that of jj203 mutant larvae [88% (n = 87) vs. 2% (n =85) in control morpholino-treated animals]. Cilia length in theolfactory placode and lateral line neuromasts is also obviouslyshorter in morphant animals (Fig. 1 G–H′). In a complementaryexperiment, we overexpressed the kif3b mRNA in embryos fromcrosses between jj203 heterozygotes. This treatment rescues cil-iogenesis in the ear of homozygous mutant embryos (n = 7 of 7vs. 0 of 3 in embryos treated with control GFP mRNA; Fig. 1 Iand I′). Taken together, these data demonstrate that a defect inthe kif3b gene is responsible for the jj203 phenotype.
kif3b Function Is Not Required in a Subset of Cilia. Vertebratephotoreceptor cells feature highly differentiated cilia, known asouter segments (17, 18). Loss of IFT genes, such as oval/ift88,leads to outer segment absence (25, 28). At 3 dpf, antibodystaining reveals ciliary axoneme at the base of the outer segmentin WT animals but not in kif3bjj203 larvae (Fig. 2A′ vs. 2A). By 5dpf, however, connecting cilia form in mutants, although they areshorter than the WT ones (Fig. 2B′ vs. 2B; graph in Fig. 2F).Ultrastructural analysis revealed that, at 3 dpf, no outer seg-ments are found in kif3bjj203 retinae (Fig. 2 C and C′). By 5 dpf,however, outer segments also form in kif3bjj203 mutants (Fig. 2 Dand D′). Mutant and WT outer segments feature similar mi-crotubule organization, including the presence of microtubulesinglets at 6 dpf (Fig. 2 E and E′). Nonetheless, outer segmentsof some mutant photoreceptors are abnormally shaped or bro-ken. These observations suggest that the kif3bjj203 locus playsa particularly prominent role in the initiation of outer segment
formation, whereas another motor contributes to outer segmentdifferentiation in parallel to kif3b at later stages.Although we determined that cilia are missing in ear maculae
of kif3bjj203 mutants by 3 dpf, the staining of ear of cristae ciliais normal even as late as at 7 dpf (Fig. 2 G and G′). Althoughwe cannot exclude minor defects in the structure of crista cilia,these results indicate that mechanisms of cilia formation aredifferent in maculae and cristae, and that kif3b is not requiredfor the differentiation of cristae cilia at least during the first7 d of development.The presence of outer segments in kif3bjj203 mutants suggests
that opsin is transported fairly normally in mutant photo-receptors. To evaluate opsin transport efficiency, we expressedGFP–Opsin–CT44 fusion gene under the control of a heat shockpromoter in kif3bjj203 mutants as described previously (29) (Fig.S1). Heat shock was applied at 5 dpf to eliminate the contribu-tion of rods, which degenerate almost completely by that time(Fig. S2). Larvae were collected at 4, 9, and 24 h after heat shock,and GFP intensity was measured in the photoreceptor cell body.Four hours after heat shock, GFP intensity is much higher in cellbodies of mutant photoreceptors compared with cells in the WT(Fig. S1 D and E). This result indicates that kif3b contributes toopsin transport. Importantly, we found that GFP–opsin fusion iscleared from the cell body by 9 h after heat shock even in theabsence of kif3b function (Fig. S1), indicating that another motormechanism transports opsin in parallel to the kif3b kinesincomplex. The persistence of some cilia in kif3b mutant homo-zygotes is surprising given that kif3a KOs in the mouse result inthe absence of cilia (14–16).
kif3b Functions Differently in Rods and Cones. To assess the ciliaryphenotype in photoreceptors of kif3bjj203 mutant animals further,we carried out whole-mount in situ hybridization using probes togenes specifically expressed in cones or rods. The expression ofcone-specific genes, such as cone transducin, cone arrestin, orcone opsins, is normal in kif3bjj203 animals. On the contrary, theexpression of rod-specific genes, such as rod transducin and rodopsin, dramatically decreases in kif3bjj203 mutants by 5 dpf (Fig.S2) as a result of cell death (Fig. S3). However, rod opsin ex-pression is normal at 3 dpf (Fig. S2A). These results were con-firmed by using antibody staining to opsins and the rod-specific
Fig. 3. kif17 mutant phenotype. (A) Upper: Sequences ofWT and kif17sa0119 mutant allele. Lower: Diagram of theKif17 protein. Arrow indicates the site of the stop codon.(B) Upper: Quantitative RT-PCR. Expression level relative toWT is provided. Data were normalized for actin expression(P < 0.05). Lower: RT-PCR amplification of the kif17 tran-script in WT and mutant embryos at 5 dpf. (C and C′) Ex-ternal phenotype of WT (C) and kif17sa0119 (C′) larvae at 7dpf. (D) Confocal and ultrastructural analysis of cilia in WTand kif17sa0119 mutants. Larvae were stained with anti-acetylated tubulin antibody to visualize cilia in the nasal pit(a and d) and ear cristae (b and e). c and f, Ultrathin sec-tions perpendicular to the distal tips of olfactory cilia in WTand mutant, respectively. (E) Graph showing the lengthdistribution of nasal cilia in WT and kif17sa0119 mutants at 7dpf. Each dot represents the average cilia length measuredin a single individual. “n” is the number of individualsanalyzed (P ≤ 0.001). (F) Confocal images of transversecryosections through the retina of adult WT (a and c) andkif17 homozygous mutant (b and d) individuals stainedwith Zpr-1 (a and b) or anti-rod opsin (c and d) antibodies(green) and counterstained with phalloidin (red). Arrow-heads indicate the outer limiting membrane, asterisks theouter plexiform layer. (G) Measurement of opsin transportefficiency from the inner to the outer segment in WT and kif17sa0119 mutant animals at 4 dpf. The intensity of GFP–opsin signal in the cell body is measured at4, 9, and 24 h after heat shock. For each data point, 15 to 20 retinae, and 30 to 36 photoreceptors were analyzed. This experiment was performed as published(29) and illustrated in Fig. S1 (*P < 0.05; **P < 0.001).
2390 | www.pnas.org/cgi/doi/10.1073/pnas.1116035109 Zhao et al.

Nr2e3 transcription factor (Fig. S2D). Similar to WT cells, coneopsins localize mainly to outer segments of mutant photo-receptors. These data suggest that rod photoreceptors degeneratein mutants between 3 and 5 dpf. In contrast to that, cone photo-receptors survive longer and frequently feature grossly normalouter segments.
kif17 Plays a Minor Role in Ciliogenesis. The formation of C. eleganscilia involves the function of the homodimeric osm-3 kinesin (30–32). To determine whether this kinesin functions in vertebrateciliogenesis, perhaps redundantly with the heterotrimeric kinesinII, we investigated mutant phenotypes of its vertebrate ortho-logue, kif17. We studied a chemically induced mutant allele,kif17sa0119, which contains a stop codon at position 551 (Fig. 3A).The resulting truncated Kif17 protein lacks 271 C-terminalamino acids. Several lines of evidence suggest that this mutationresults in a null or a near-null phenotype. First, it eliminatesseveral highly conserved sequences in the C terminus of theKif17 protein. Second, the C-terminal region of this polypeptideis involved in cargo interactions (33, 34). Third, this mutationresults in a nonsense-mediated decay that severely reduces thekif17 transcript expression (Fig. 3B). Finally, morpholinoknockdown of kif17 function in kif17sa0119 mutant homozygotesdoes not result in any obvious enhancement of cilia defects (Fig.S4B). kif17sa0119 mutant homozygotes do not display any obviousexternal phenotype (Fig. 3 C and C′) and survive to adulthood.
Immunostaining did not reveal any ciliogenesis defects in theear, the retina, the spinal cord, or the pronephric duct of mutanthomozygotes (Fig. 3D and Fig. S4). However, the nasal cilia weresomewhat shorter at 7 dpf (P < 0.001; Fig. 3 D and E), and 8 dpf(P < 0.01; n ≥ 10 for WT and mutant). In contrast to nematodeosm-3 mutant phenotype (30), singlet microtubules persist innasal cilia of kif17 mutants (Fig. 3D), which continue to bemotile (Movies S1 and S2). Antibody staining did not reveal rodopsin or green cone opsin mislocalization in mutant retinae at 5dpf (Fig. S4). Similarly, we did not observe obvious morpho-logical defects or opsin mislocalization in the retinae of adultmutant homozygotes (Fig. 3F). As kif17 may not be required foropsin localization and may only contribute to its transport, weapplied GFP–opsin transient overexpression assay at 4 dpf (Fig.S1). Opsin transport is not affected in this test (Fig. 3G). Theseresults indicate that the kif17 kinesin plays a minor role in ciliaformation, and does not contribute to the transport of opsin, themost abundant cargo in photoreceptor cilia.
kif17 Can Substitute for Loss of kif3 Function in Some Cilia. Giventhat kif3 and kif17 function redundantly in C. elegans, we testedwhether they are functionally interchangeable in a vertebrate.We injected kif17 mRNA into embryos from crosses betweentwo kif3bjj203 heterozygotes, and found a significant decrease inthe frequency of severe curly body phenotype. Four percent(n = 433) of embryos displayed this phenotype, compared with
Fig. 4. kif3c function in ciliogenesis. (A and A′) Confocalimages of transverse cryosections through the retinastained with Zpr-1 (to visualize double cones; green), andanti-green opsin antibodies (red). (B and B′) Confocalimages of lateral cristae in whole animals stained with anti-acetylated tubulin antibody to visualize kinocilia (green)and counterstained with phalloidin (red). WT andkif3bjj203−/−;kif17sa0119−/− double mutant animals were an-alyzed as indicated. (C) Cilia length in WT, mutant, anddouble mutant cristae as indicated. The average WT lengthis set at 100%. “n” is the number of individuals analyzed.(D) A schematic drawing of the exon/intron structure forthe kif3c and kif3c-like genes. Morpholino-targeted exonsare in color. Target sites are indicated as red horizontalbars. (E) Confocal images of lateral cristae in whole animalsstained as in B. The number of individuals tested that dif-ferentiate cilia is indicated in lower left corner of eachpanel. (F) Confocal images of transverse cryosectionsthrough the retina stained with an anti-acetylated tubulinantibody to visualize cilia (arrows). For E and F, genotypesare indicated below. Brackets indicate the photoreceptorcell layer, arrowheads the outer limiting membrane. (G)The frequency of photoreceptor cilia at 4 dpf in differentmutant/morphant backgrounds as indicated. (H) The fre-quency of photoreceptor cilia in kif3b−/− mutant homo-zygotes at 3 dpf, following rescue with kif3c or control GFPmRNA. In G and H, each dot represents the number of ciliaper an arbitrary segment of the photoreceptor cell layer ina single retina. (I and I′) Confocal images of photoreceptorcilia (arrows) at 3 dpf visualized as in F. (J) Relative sizes ofapical opsin-positive compartments (presumably outersegments) in kif3bjj203 homozygous mutant and pheno-typically WT animals treated with kif3c or control GFPmRNA. Each dot represents the total size of apical opsin-positive domains on a single section through the retina,adjusted for the length of the photoreceptor cell layer. Theaverage WT size equals 100%. (K) Efficiency of GFP–opsintransport in kif3c morphants measured as in Fig. S1. “n” isthe number of photoreceptors analyzed. A minimum of 30sections from six retinae were used to calculate each datapoint. In G, H, and J, “n” is the number of retinae analyzed.In all images, apical direction is up (*P < 0.05; **P < 0.001).
Zhao et al. PNAS | February 14, 2012 | vol. 109 | no. 7 | 2391
DEV
ELOPM
ENTA
LBIOLO
GY

18% (n = 348) in GFP mRNA-treated controls. Immunostain-ing revealed that spinal canal cilia but not nasal cilia were res-cued at 3 dpf in mutant embryos treated with kif17 mRNA (Fig.S5). We did not observe rescue in control kif3bjj203 embryostreated with GFP mRNA. These results indicate that kif17kinesin can substitute for kif3b function to drive ciliogenesis insome tissues.
kif3b;kif17 Double Mutant.The persistence of cilia in kif3bmutantssuggested that kif3 and kif17 may function redundantly. To testthis idea, we investigated the phenotype of kif17sa0119/kif17sa0119;kif3bjj203/kif3bjj203 double homozygotes. Staining with anti-greenopsin antibodies in double mutants did not reveal obvious mis-localization of this visual pigment (Fig. 4 A and A′). Similarly, inthe absence of both kinesins, cilia of auditory cristae differentiatenormally and display normal length (Fig. 4 B, B′, and C). Theseresults indicate that neither kif3b nor kif17 is required for cilio-genesis in cone photoreceptors and in a subset of hair cells.Moreover, unless the third parallel motor mechanism is involved,these two kinesins do not function redundantly in the cilia ofthese cells.
kif3b and kif3c Display Redundant Functions. The Kif3c kinesinsubunit is thought to form complexes with Kif3a (9, 10). Al-though the mouse Kif3c protein appears to be absent from thephotoreceptor cell layer (10, 22, 23), we considered the possi-bility that the zebrafish kif3c functions redundantly with kif3b.The zebrafish has two kif3c-related genes, which we will refer toas kif3c and kif3c-like (Fig. 4D and Fig. S6). To block their ex-pression, we designed anti-splice site morpholinos and verifiedtheir efficiency by real-time-PCR (Fig. S7). We tested the ideathat in the absence of kif3b, these kinesins may be necessary forthe formation of cilia in cristae and in photoreceptor cells. Thisturned out to be the case: although neither kif3c nor kif3c-likemorphants display cilia defects, simultaneous morpholinoknockdown of kif3c and kif3c-like in kif3bjj203 mutants results ina loss of cristae and photoreceptor cilia (Fig. 4 E and F; graph inFig. 4G). Subsequent single knockdowns of kif3c or kif3c-likerevealed that kif3c, but not kif3c-like, functions redundantly withkif3b (P < 0.0001; Fig. 4G).
Developmental Changes in Kinesin Repertoire. The redundancy ofkif3c and kif3b function becomes obvious only at 4 dpf afterphotoreceptor cilia are formed in kif3b mutants, suggesting thatkif3c does not function during early stages of ciliogenesis. GFP–opsin transient overexpression assay in kif3c morphants at 3 dpfdid not reveal significant differences in opsin transport efficiencybetween kif3c and control morphants at 3 dpf (Fig. 4K), in-dicating that, consistent with the kif3b phenotype, kif3c does notcontribute to this process at this stage. By 5 dpf, however, GFP-opsin transport was somewhat slower in kif3c morphants com-pared with control animals (P < 0.05; Fig. 4K). To further testthe idea that the absence of cilia at the early stages of photo-receptor differentiation in kif3b mutants is caused by the absenceof kif3c function, we overexpressed kif3c mRNA in kif3bjj203
mutant homozygotes. This treatment resulted in a significantrescue of body curvature (13% vs. 23% following kif3c and GFPRNA injections, respectively; n = 520 and 261, respectively) andcilia differentiation in kif3bjj203 mutants at 3 dpf (P < 0.001; Fig.4 H, I, and I′) and 4 dpf (P < 0.001; Fig. S8B). Finally, thistreatment increased opsin accumulation at the apical terminus ofthe photoreceptor cell, a phenotype that most likely reflectsouter segment formation (P < 0.001; Fig. 4J and Fig. S8A). Anincomplete rescue is most likely a result of limited stability ofinjected mRNA. Based on these results, we propose that thefunctional repertoire of ciliary kinesins undergoes a develop-mental change during photoreceptor differentiation: whereasonly kif3b drives cilia formation early on, kif3b and kif3c perform
this function at later stages. The significance of this develop-mental change is not clear, but it may facilitate the transport ofthe massive amount of cargo that needs to be moved from thecell body into the outer segment.
DiscussionExperiments presented in this work reveal that kif3b and kif17,the two kinesins known to play major roles in nematode cilio-genesis, display very different contributions to the formation ofvertebrate cilia. In C. elegans, the homodimeric kif17/osm-3kinesin is required for the differentiation of distal microtubulesinglets of amphidial channel cilia (30). In addition, in this set ofcilia, and in the cilia of AWC cells, the homodimeric and theheterotrimeric kinesin function redundantly. In contrast tochannel cilia, kif17/osm-3 is not required for the differentiation ofdistal singlets in neighboring AWB amphidial cilia, although thetwo kinesins still function redundantly (35). Functional inter-actions between kinesins are even more complex in the cephalicmale cilia (32). Our data reveal that diverse functional rela-tionships also exist between vertebrate ciliary kinesins (Fig. S9).A limited role for kif17 in zebrafish ciliogenesis, especially in
double mutants with kif3b, is surprising, given the prominentfunction of this kinesin in C. elegans cilia (31, 36). In vertebrates,the function of kif17 has been studied in tissue culture as well asin mouse and zebrafish models. Tissue culture studies providedevidence that kif17 is necessary for the transport of a cyclic nu-cleotide-gated channel subunit but not for the elongation of cilia(37), whereas the mouse KO analysis revealed a role in learningand memory (24). Our studies demonstrate a function for kif17in the morphogenesis of a subset of vertebrate cilia. However,the role of kif17 is surprisingly limited. Combined with previousstudies, our data indicate that kif17 functions in the transport ofstructural and signaling components in a limited subset of cilia.The results of our analysis are in agreement with reports that
kif3c KO mice are viable and do not display photoreceptordefects (22, 23). However, a redundancy of kif3c/kif3b function inphotoreceptor cells is surprising in light of previous reports thatkif3c is primarily expressed in ganglion cell axons and in ama-crine cells (9, 10). The same studies reported that Kif3c isstructurally related to Kif3b and, similar to Kif3b, associates withKif3a, which suggests that Kif3b and Kif3c proteins may functionredundantly as alternative binding partners of Kif3a. Althoughredundancy of kif3b and kif3c function has been hypothesized (9,22), to our knowledge, this is the first study to demonstrate that itactually exists. In the ear, this redundancy is limited to cilia ina subset of auditory hair cells. Hair cells that display the re-dundancy of function are very similar to those that do not. Thismay be a result of somewhat different cargo requirements. Ciliaof hair cells in cristae are particularly long, so they may requirea more robust transport compared with neighboring cells in earmaculae. Similarly, somewhat different cargo molecules aretransported in rod and cone cilia, which may account for dif-ferences of kif3bjj203 phenotype in these two cell types (18, 38).Further biochemical analysis as well as in vivo imaging of IFTmovement will be necessary to fully understand the diversity ofkinesin functions in cilia.Interestingly, cilia formation in photoreceptor cells is initially
driven by kif3b only. At later stages, kif3c also contributes tociliogenesis in this cell type and is largely sufficient to drive thedifferentiation of outer segments. This, to our knowledge, is thefirst example of a developmental change in the repertoire ofciliary kinesins during differentiation of a single cell type. Theouter segments of kif3b mutant homozygotes, although relativelyrobust, do not appear to be entirely normal. It remains to beinvestigated whether this results from quantitative differences inkinesin expression, or reflects differences in cargo specificitybetween Kif3c and Kif3b kinesins.
2392 | www.pnas.org/cgi/doi/10.1073/pnas.1116035109 Zhao et al.

Materials and MethodsAnimals. Zebrafish strains were maintained following the standard protocolsapproved by the Tufts University Animal Care Committee.
Genetic Screen. ENU mutagenesis was performed as described (39). To en-hance our ability to identify subtle photoreceptor defects, we crosseda transgene that expresses GFP in rod photoreceptors into ENU-mutagen-ized animals. Mutants were identified in the F2 generation by using theearly pressure screening approach (40).
Knockdown and Rescue Experiments. Morpholino knockdowns and rescueexperiments were performed as described previously (25, 26). During rescueexperiments, embryos were genotyped by sequencing. The following morpho-linos were used: kif3c-like, GACGTACTTGAATTTCATCTCTCTT; kif3c, TCAGT-CCTCA GACACATACC TTAAA; kif3b (ATG), AGCTCTTGCT TTTAGACATT TTGAC;kif3b (SP), AGCTTGAAGT TTCTAACCTT AACT; and kif17 (SP) the same as in thestudy of Insinna et al. (11): TTGTAAACTG GTTACCTGGA TTGTC. Knockdownefficiency was confirmed by using the following RT-PCR primers: for kif3c,ATCCGCGACCTGCTCACCAAAG and CAGTGATGATGAAGATGGCGTGAG; forkif3c-like, AACGAGGGATGCTGGCGAAAGA and TGGTCTCTTCATCTTGTTCA.
Immunohistochemistry. Sectioning and immunohistochemistry were per-formed by using standard protocols (27). The following antibodies wereused: anti-acetylated α-tubulin (1:500; Sigma), zpr3 (1:1,000; Zebrafish In-ternational Resource Center), zpr1 (1:250; Zebrafish International ResourceCenter), anti–γ-tubulin (1:500; Sigma), anti-green opsin (gift from ThomasVihtelic), and anti-Nr2e3 (gift from Jeremy Nathans).
Opsin Transport Analysis. Opsin transport was analyzed as described pre-viously (29). GFP–opsin fusion construct was injected into kif17 or kif3bmutant homozygotes or, alternatively, injected together with anti-kif3cmorpholinos into WT embryos.
Quantitative PCR. kif17 transcripts were amplified from 5-d-old WT or kif17homozygous mutant larvae. β-actin was used as an internal reference con-trol. Analysis was performed by using Qiagen SYBR Green PCR Kit ona Stratagene MX3000 cycler with the following settings: 95 °C for 30 s, 57 °Cfor 1 min, and 72 °C for 1 min for 40 cycles. The data were collected fromthree independent experiments (each in triplicate). The relative expressionlevel of kif17 transcripts in WT and mutant larvae were analyzed with Qia-gen REST 2009 software. The following primers were used for amplification:kif17 forward, GCTTCACAAGAACAGGCTAAG; kif17 reverse, CATCTCAAAC-TCTGCCTGTAG; β-actin forward, ATGGATGATGAAATTGCCGCAC; and β-actinreverse, ACCATCACCAGAGTCCATCACG.
Videomicroscopy and Image Analysis. To image nasal cilia movement, larvaewere embedded head down in 1% agarose on a glass bottom dish and filmedusing a QuantEM 512SC camera and a 63× water immersion lens on a ZeissAxio Observer Z1 inverted microscope. To evaluate ciliogenesis in the pho-toreceptor cell layer, cilia were counted relative to the length of the outerlimiting membrane on each confocal image. For the analysis of rod opsindistribution, the areas of apical rod opsin accumulation were selected withthe wand tool (tolerance of 20), and their sizes were calculated by usingImageJ software (National Institutes of Health).
ACKNOWLEDGMENTS. We thank Dr. Thomas Vihtelic for anti-green opsinand Dr. Jeremy Nathans for anti-Nr2e3 antibodies; Dr. James Fadool forproviding the rod-GFP transgenic line; the Louisville mapping facility forassistance in cloning the jj203 mutant allele; Dr. Xinjun He for help withphylogenetic analysis and Dr. Viktoria Andreeva with quantitative PCR;Dr. Tomer Avidor-Reiss for commenting on an earlier version of this manu-script; and the Sanger Institute Zebrafish Mutation Resource, sponsored byWellcome Trust Grant WT 077047/Z/05/Z, for providing the zebrafishkif17sa0119 allele. This work was supported by National Institutes of HealthGrants R01 EY018176 and R01 EY016859 (to J.M.).
1. Rosenbaum JL, Witman GB (2002) Intraflagellar transport. Nat Rev Mol Cell Biol 3:813–825.
2. Satir P, Christensen ST (2007) Overview of structure and function of mammalian cilia.Annu Rev Physiol 69:377–400.
3. Lawrence CJ, et al. (2004) A standardized kinesin nomenclature. J Cell Biol 167:19–22.4. Cole DG, et al. (1993) Novel heterotrimeric kinesin-related protein purified from sea
urchin eggs. Nature 366:268–270.5. Shakir MA, Fukushige T, Yasuda H, Miwa J, Siddiqui SS (1993) C. elegans osm-3 gene
mediating osmotic avoidance behaviour encodes a kinesin-like protein. Neuroreport4:891–894.
6. Walther Z, Vashishtha M, Hall JL (1994) The Chlamydomonas FLA10 gene encodesa novel kinesin-homologous protein. J Cell Biol 126:175–188.
7. Jana SC, Girotra M, Ray K (2011) Heterotrimeric kinesin-II is necessary and sufficient topromote different stepwise assembly of morphologically distinct bipartite cilia inDrosophila antenna. Mol Biol Cell 22:769–781.
8. Signor D, Wedaman KP, Rose LS, Scholey JM (1999) Two heteromeric kinesin com-plexes in chemosensory neurons and sensory cilia of Caenorhabditis elegans. Mol BiolCell 10:345–360.
9. Yang Z, Goldstein LS (1998) Characterization of the KIF3C neural kinesin-like motorfrom mouse. Mol Biol Cell 9:249–261.
10. Muresan V, et al. (1998) KIF3C and KIF3A form a novel neuronal heteromeric kinesinthat associates with membrane vesicles. Mol Biol Cell 9:637–652.
11. Insinna C, Pathak N, Perkins B, Drummond I, Besharse JC (2008) The homodimerickinesin, Kif17, is essential for vertebrate photoreceptor sensory outer segment de-velopment. Dev Biol 316:160–170.
12. Marszalek JR, Ruiz-Lozano P, Roberts E, Chien KR, Goldstein LS (1999) Situs inversusand embryonic ciliary morphogenesis defects in mouse mutants lacking the KIF3Asubunit of kinesin-II. Proc Natl Acad Sci USA 96:5043–5048.
13. Nonaka S, et al. (1998) Randomization of left-right asymmetry due to loss of nodalcilia generating leftward flow of extraembryonic fluid in mice lacking KIF3B motorprotein. Cell 95:829–837.
14. Cano DA, Sekine S, Hebrok M (2006) Primary cilia deletion in pancreatic epithelial cellsresults in cyst formation and pancreatitis. Gastroenterology 131:1856–1869.
15. Croyle MJ, et al. (2011) Role of epidermal primary cilia in the homeostasis of skin andhair follicles. Development 138:1675–1685.
16. Lin F, et al. (2003) Kidney-specific inactivation of the KIF3A subunit of kinesin-II in-hibits renal ciliogenesis and produces polycystic kidney disease. Proc Natl Acad SciUSA 100:5286–5291.
17. Rodieck RW (1973) The Vertebrate Retina. Principles of Structure and Function (W. H.Freeman, San Francisco).
18. Kennedy B, Malicki J (2009) What drives cell morphogenesis: A look inside the ver-tebrate photoreceptor. Dev Dyn 238:2115–2138.
19. Marszalek JR, et al. (2000) Genetic evidence for selective transport of opsin and ar-restin by kinesin-II in mammalian photoreceptors. Cell 102:175–187.
20. Avasthi P, et al. (2009) Trafficking of membrane proteins to cone but not rod outersegments is dependent on heterotrimeric kinesin-II. J Neurosci 29:14287–14298.
21. Yamazaki H, Nakata T, Okada Y, Hirokawa N (1996) Cloning and characterization of
KAP3: A novel kinesin superfamily-associated protein of KIF3A/3B. Proc Natl Acad Sci
USA 93:8443–8448.22. Yang Z, Roberts EA, Goldstein LS (2001) Functional analysis of mouse kinesin motor
Kif3C. Mol Cell Biol 21:5306–5311.23. Jimeno D, Lillo C, Roberts EA, Goldstein LS, Williams DS (2006) Kinesin-2 and pho-
toreceptor cell death: requirement of motor subunits. Exp Eye Res 82:351–353.24. Yin X, Takei Y, Kido MA, Hirokawa N (2011) Molecular motor KIF17 is fundamental
for memory and learning via differential support of synaptic NR2A/2B levels. Neuron
70:310–325.25. Tsujikawa M, Malicki J (2004) Intraflagellar transport genes are essential for differ-
entiation and survival of vertebrate sensory neurons. Neuron 42:703–716.26. Omori Y, et al. (2008) Elipsa is an early determinant of ciliogenesis that links the IFT
particle to membrane-associated small GTPase Rab8. Nat Cell Biol 10:437–444.27. Malicki J, Avanesov A, Li J, Yuan S, Sun Z (2011) Analysis of cilia structure and function
in zebrafish. Methods Cell Biol 101:39–74.28. Doerre G, Malicki J (2002) Genetic analysis of photoreceptor cell development in the
zebrafish retina. Mech Dev 110:125–138.29. Zhao C, Malicki J (2011) Nephrocystins and MKS proteins interact with IFT particle and
facilitate transport of selected ciliary cargos. EMBO J 30:2532–2544.30. Snow JJ, et al. (2004) Two anterograde intraflagellar transport motors cooperate to
build sensory cilia on C. elegans neurons. Nat Cell Biol 6:1109–1113.31. Evans JE, et al. (2006) Functional modulation of IFT kinesins extends the sensory
repertoire of ciliated neurons in Caenorhabditis elegans. J Cell Biol 172:663–669.32. Morsci NS, Barr MM (2011) Kinesin-3 KLP-6 regulates intraflagellar transport in male-
specific cilia of Caenorhabditis elegans. Curr Biol 21:1239–1244.33. Setou M, Nakagawa T, Seog DH, Hirokawa N (2000) Kinesin superfamily motor pro-
tein KIF17 and mLin-10 in NMDA receptor-containing vesicle transport. Science 288:
1796–1802.34. Guillaud L, Wong R, Hirokawa N (2008) Disruption of KIF17-Mint1 interaction by
CaMKII-dependent phosphorylation: A molecular model of kinesin-cargo release. Nat
Cell Biol 10:19–29.35. Mukhopadhyay S, et al. (2007) Distinct IFT mechanisms contribute to the generation
of ciliary structural diversity in C. elegans. EMBO J 26:2966–2980.36. Ou G, Blacque OE, Snow JJ, Leroux MR, Scholey JM (2005) Functional coordination of
intraflagellar transport motors. Nature 436:583–587.37. Jenkins PM, et al. (2006) Ciliary targeting of olfactory CNG channels requires the
CNGB1b subunit and the kinesin-2 motor protein, KIF17. Curr Biol 16:1211–1216.38. Pugh E, Lamb T (2000) Phototransduction in vertebrate rods and cones. Handbook of
Biological Physics (Elsevier, Amsterdam), Vol 3, pp 183–255.39. Malicki J, et al. (1996) Mutations affecting development of the zebrafish retina.
Development 123:263–273.40. Malicki J (2000) Harnessing the power of forward genetics—analysis of neuronal di-
versity and patterning in the zebrafish retina. Trends Neurosci 23:531–541.
Zhao et al. PNAS | February 14, 2012 | vol. 109 | no. 7 | 2393
DEV
ELOPM
ENTA
LBIOLO
GY