Embed Size (px)
Citation preview

ARTICLE IN PRESS
European Journal of
PROTISTOLOGY
0932-4739/$ - se
doi:10.1016/j.ej
�Current addDuve Institute
B1200 Brussels
E-mail addr
European Journal of Protistology 42 (2006) 115–123
www.elsevier.de/ejop
Isolation and molecular identification of free-living amoebae of
the genus Naegleria from Arctic and sub-Antarctic regions
Johan F. De Jonckheere�
Scientific Institute of Public Health, 1050 Brussels, Belgium
Received 5 December 2005; received in revised form 25 January 2006; accepted 1 February 2006
Abstract
Twenty-three freshwater samples with sediment taken from two regions in the Arctic, Spitzbergen and Greenland,and one region in sub-Antarctica, Ile de la Possession, were cultured for amoebae at 37 1C and room temperature (RT).Only two samples yielded amoebae at 37 1C and the two isolates were identified from their morphological features tobelong to the genus Acanthamoeba. Vahlkampfiid amoebae were isolated from 11 samples at RT. Morphologicalanalysis of the cysts identified all 11 isolates as belonging to the genus Naegleria, although only about half of them(45%) transformed into flagellates. Ribosomal DNA sequence analysis demonstrated that these isolates representnovel species and that N. antarctica, N. dobsoni and N. chilensis are their closest relatives. Not surprisingly, these threespecies also grow at lower temperatures (o37 1C) than the majority of described Naegleria spp. Two of the eight newspecies were found in both Arctic and sub-Antarctic regions, and other new species from the Arctic are closely relatedto new species from the sub-Antarctic. Therefore, it seems the Naegleria gene pool present in the polar regions isdifferent from that found in temperate and tropical regions.r 2006 Elsevier GmbH. All rights reserved.
Keywords: 5.8S rDNA; Arctic species; ITS; Naegleria; New species; Sub-Antarctic species
Introduction
Free-living amoebae of the genus Naegleria areroutinely isolated while searching for the pathogenicN. fowleri in various environments. High incubationtemperatures, up to 45 1C, are used to screen for thepresence of this human pathogen. Therefore, mostNaegleria spp. identified until now have a high maximumgrowth temperature, usually between 37 and 45 1C(De Jonckheere 2002). A few isolates belonging to other
e front matter r 2006 Elsevier GmbH. All rights reserved.
op.2006.02.001
ress: Research Unit for Tropical Diseases, Christian de
of Cellular Pathology, Avenue Hippocrate, 74-75,
, Belgium. Fax: +322 7626853.
ess: [email protected].
genera of the Vahlkampfiidae have also been isolated (e. g.Tetramitus entericus, Singhamoeba horticola, Learamoeba
waccamawensis, see De Jonckheere and Brown 2005a).There are some scattered reports on vahlkampfiids
isolated from Antarctic sites, in which the isolatesturned out to be new species (Murtagh et al. 2002;Garstecki et al. 2005), specially adapted to coldtemperatures. Page (1988) reports on a strain ofV. ustiana (now called Paravahlkampfia ustiana) isolatedfrom Antarctica. This is surprising since this species wasoriginally isolated from an indoor swimming pool in theCzech Republic (Page 1974). Caution should beexercised, however, since no details on the source ofthe Antarctic strain were given and the identificationwas based solely on morphological features.

ARTICLE IN PRESSJ.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123116
There is one more comprehensive publication on theoccurrence of free-living amoebae in Antarctic soil andwater (Brown et al. 1982), but in this case most of theisolates were only identified to genus on the basis ofmorphological characters. In this study, isolations wereperformed at 30 1C, and many of the isolates (50%) wereable to grow at 37 1C upon subculture. It was mostly theisolated vahlkampfiids, and especially the Naegleria
isolates, which were unable to grow at 37 1C.To my knowledge, there are no reports on isolation of
vahlkampfiids from Arctic regions. I report here theisolation of vahlkampfiids from water samples withsediment collected in Arctic and sub-Antarctic regions.The isolates were identified with molecular methods toensure accurate species designations.
Material and methods
Arctic water with sediment samples were collected inSpitzbergen (Norway), in July 2004, and on DiskoIsland (Greenland), in September 2004. Sub-Antarcticsamples were collected on Ile de la Possession (CrozetArchipelago), in January 2005. After being kept for afew weeks at room temperature (RT), the samples wereinoculated onto plates containing non-nutrient (NN)agar coated with Escherichia coli (Page 1988). The sealedplates were incubated at RT and 37 1C. Because thecultures grown at 37 1C were negative for vahlkampfiids,the samples were pre-incubated for 7 days at 37 1Cbefore attempting a re-isolation at this temperature.
Only isolates that showed the typical vahlkampfiidmorphology (Page 1988) were further investigated.Isolates from RT (20–24 1C) plates were tested forgrowth at 37 and 30 1C. Flagellate formation wasinvestigated by suspending the trophozoites in distilledwater at RT.
DNA was isolated from pelleted trophozoites usingthe UNSET method (Hugo et al. 1992). The ITS1, 5.8Sand ITS2 rDNA were PCR-amplified using the ITSforward primer and the ITS reverse primer, correspond-ing to the 30 end of the SSU rDNA and the 50 end of thelarge subunit (LSU) rDNA, respectively (De Jonckheere2004). Two pairs of ITS primers were employed, onepair designed for amplifying DNA from Naegleria spp.specifically (De Jonckheere 1998) and a second lessspecific pair for amplifying DNA from vahlkampfiidspecies (De Jonckheere and Brown 2005a). The PCRproduct was sequenced (both strands) with the ampli-fication primers without cloning with a BeckmanCEQ2000 sequencer using the CEQ Dye TerminatorCycle Sequencing kit (Beckman Coulter Inc., Fullerton,CA, USA).
The sequences were aligned with rDNA sequences ofother vahlkampfiids (De Jonckheere 2004; De Jonc-
kheere and Brown 2005a) using the programmeClustalX (Thompson et al. 1997). The alignment wasmanually adjusted. Phylogenetic trees were constructedusing the DNAPARS (parsimony), DNADIST (dis-tance matrix), NEIGHBOR (neighbor-joining andUPGMA), FITCH, KITSCH, DNAML, DNAMLKand SEQBOOT (bootstrapping) programs of the PHY-LIP (version 3.572c) package (Felsenstein 1989). Thecomplete ITS1-5.8S rDNA-ITS2 sequences of the newlyidentified Naegleria strains have been deposited inEMBL with accession numbers AM157657–AM157664.
To investigate the presence of group I introns at site516 of the small subunit (SSU) rDNA, the DNA of allstrains was amplified with a SSU forward primer andprimer 1000RV, an internal reverse primer of the SSUrDNA sequence (De Jonckheere 1994). DNA of the typestrain of N. italica, containing the group I intron, wasincluded as a positive control. The length of the PCRamplified products was compared on a 0.7% agarose gelto that of the positive control.
A strain of each new species was deposited at theCulture Collection of Algae and Protozoa, Dunbeg,Argyll, UK.
Results
Morphological identification
Only one out of 16 samples from the Arctic regionyielded amoebae at 37 1C, and the amoeba was identifiedfrom morphological characters to belong to the genusAcanthamoeba. Out of the seven samples from the sub-Antarctic region also only one yielded amoebae at 37 1C,and again this was identified as an Acanthamoeba. Pre-incubation of the samples at 37 1C prior to isolation didnot increase the number of isolates at 37 1C.
A high percentage of the samples (87%) from theArctic region yielded amoebae at RT. Six isolates (46%)were identified as belonging to the genus Naegleria
(Table 1) and identified further by molecular methods.None of them grew at 37 1C upon subculture, and onlytwo isolates (SP8 and GR11) grew at 30 1C. A similarhigh percentage of the samples (86%) from the sub-Antarctic region yielded amoebae at RT. Five isolates(83%) were identified as belonging to the genusNaegleria (Table 1) and investigated further by mole-cular study. None of them could be grown at 37 1C uponsubculture and only one isolate (IP7) grew at 30 1C.
Although all vahlkampfiid isolates showed a cystmorphology typical for the genus Naegleria (i.e. mostcysts were spherical to angular with pores) only five outof these 11 isolates (45%) could be induced to formflagellates (Table 1). However, molecular methodsconfirmed that they all belonged to the genus Naegleria.

ARTICLE IN PRESS
Table 1. Amoeba isolation at room temperature (all negative at 37 1C) and vahlkampfiid identification in the samples from Arctic
and sub-Antarctic regions
Origin Sample no. Amoeba Vahlkampfiid identificationa Enflagellation
Spitzbergen Sp1 + � NA
Sp2 � NA NA
Sp3 + Naegleria �
Sp4 + Naegleria +
Sp5 + Naegleria �
Sp6 + � NA
Sp7 � NA NA
Sp8 + Naegleria +
Sp9 � NA NA
Greenland Gr10 + Naegleria +
Gr11 + Naegleria +
Gr12 + � NA
Gr13 +b� NA
Gr15 + � NA
Gr16 + � NA
Gr17 + � NA
Ile de la possession IP1 � NA NA
IP2 + Naegleria +
IP3 + � NA
IP4 + Naegleria �
IP5 + Naegleria �
IP6 +b Naegleria �
IP7 + Naegleria �
NA: not applicable.aNone of the isolates grows at 37 1C upon subculture, and only SP8, Gr11 and IP7 grow at 30 1C.bAlso an isolate at 37 1C, identified as Acanthamoeba sp.
J.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123 117
Molecular identification
The DNA of all the vahlkampfiid isolates wasamplified using both the general ITS primers and themore specific Naegleria ITS primers, indicating that allisolates belonged to the genus Naegleria.
Isolates SP3, GR10 and IP6 have identical ITS1, 5.8Sand ITS2 sequences. The ITS1 and 5.8S rDNA sequencesare identical to those of N. antarctica. However, the ITS2sequences of strains SP3, GR10 and IP6 are much shorter,at 297bp (Table 2), compared to 439bp for N. antarctica.The ITS1 of N. antarctica and these three new isolates isspecial because it is 36 bp long, which is different from themajority of the Naegleria spp., where it is only 33bp long.Because strains of this new species were isolated from bothpole regions I propose the new name N. polaris.
Isolate SP4 has the same ITS1 and 5.8S sequences asstrains SP3, GR10 and IP6, and although the ITS2length is the same, the ITS2 sequence of SP4 differsby 8 bp (substitutions) from that of the new speciesN. polaris (Table 2). Therefore, a new species name,N. neopolaris, is proposed, to indicate the close relation-ship with the new species 1.
The ITS1 and 5.8S sequences of isolate SP5 areidentical to those of N. antarctica while the ITS2 differs
by 39 bp (24 substitutions and 15 indels) from it(Table 2). Because of the differences from its closestrelative this new isolate is considered to represent a newspecies. I propose the name N. arctica for strain SP5because it was isolated only from the Arctic region.
The 5.8S sequence of isolate SP8 differs by 4 bp(substitutions) from that of N. niuginensis and 6 bp(substitutions) from that of N. dobsoni. But both theITS1 and the ITS2 sequences have more resemblance tothose of N. dobsoni (Table 2). For strain SP8 I proposethe name N. spitzbergenensis to denote the place oforigin.
Isolates GR11 and IP4 have identical ITS1, 5.8S andITS2 sequences. Their closest relative is N. dobsoni, withonly 3 bp difference (substitutions) in the ITS1, and 4 bpdifference (2 deletions and two substitutions) in the ITS2(Table 2). The 5.8S rDNA sequence of these new strainsis identical to that of N. dobsoni. The latter is reportedalso to grow at less than 35 1C, although it was isolatedin South Australia. I propose the name N. neodobsoni
for strains GR11 and IP4 to denote the close relation-ship with N. dobsoni.
The ITS2 of isolate IP2 differs in length by 1 bpfrom that of N. antarctica. There is a 4 bp difference(3 substitutions and one deletion) between the ITS2 of
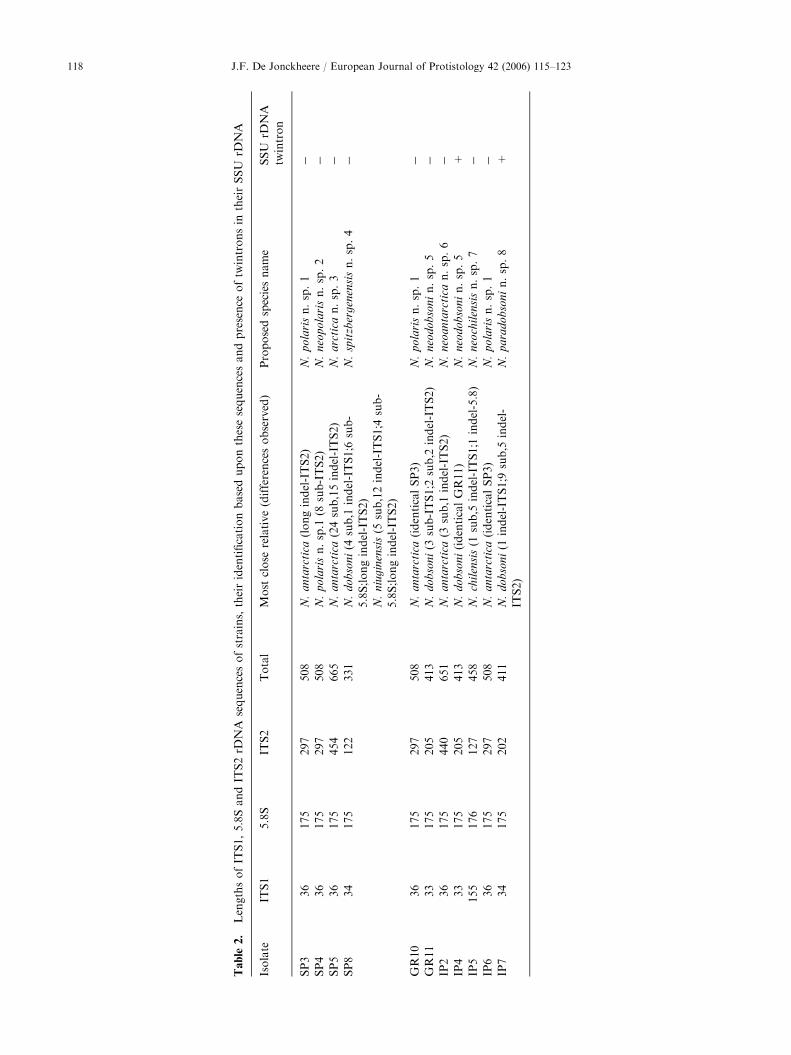
ARTICLE IN PRESS
Table
2.
LengthsofIT
S1,5.8SandIT
S2rD
NA
sequencesofstrains,theiridentificationbaseduponthesesequencesandpresence
oftw
intronsin
theirSSU
rDNA
Isolate
ITS1
5.8S
ITS2
Total
Most
close
relative(differencesobserved)
Proposedspeciesname
SSU
rDNA
twintron
SP3
36
175
297
508
N.
an
tarc
tica
(longindel-ITS2)
N.
po
lari
sn.sp.1
�
SP4
36
175
297
508
N.
po
lari
sn.sp.1
(8sub-ITS2)
N.
neo
po
lari
sn.sp.2
�
SP5
36
175
454
665
N.
an
tarc
tica
(24sub,15indel-ITS2)
N.
arc
tica
n.sp.3
�
SP8
34
175
122
331
N.
do
bso
ni(4
sub,1
indel-ITS1;6
sub-
5.8S;longindel-ITS2)
N.
spit
zber
gen
ensi
sn.sp.4
�
N.
niu
gin
ensi
s(5
sub,12indel-ITS1;4
sub-
5.8S;longindel-ITS2)
GR10
36
175
297
508
N.
an
tarc
tica
(identicalSP3)
N.
po
lari
sn.sp.1
�
GR11
33
175
205
413
N.
do
bso
ni(3
sub-ITS1;2
sub,2
indel-ITS2)
N.
neo
do
bso
nin.sp.5
�
IP2
36
175
440
651
N.
an
tarc
tica
(3sub,1
indel-ITS2)
N.
neo
an
tarc
tica
n.sp.6
�
IP4
33
175
205
413
N.
do
bso
ni(identicalGR11)
N.
neo
do
bso
nin.sp.5
+
IP5
155
176
127
458
N.
chil
ensi
s(1
sub,5
indel-ITS1;1
indel-5.8)
N.
neo
chil
ensi
sn.sp.7
�
IP6
36
175
297
508
N.
an
tarc
tica
(identicalSP3)
N.
po
lari
sn.sp.1
�
IP7
34
175
202
411
N.
do
bso
ni(1
indel-ITS1;9
sub,5
indel-
ITS2)
N.
pa
rad
ob
son
in.sp.8
+
J.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123118

ARTICLE IN PRESSJ.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123 119
this new isolate and that of N. antarctica (Table 2). TheITS1 and 5.8S sequences are identical. The name N.
neoantarctica is proposed for this new isolate.The ITS2 of isolate IP5 is identical to that of
N. chilensis, but it differs by 1 bp (deletion) in the5.8S, and 6 bp (one substitution and 5 indels) in the ITS1from N. chilensis (Table 2). The maximum growthtemperature of N. chilensis has also been recorded asbeing 30 1C (De Jonckheere et al. 2001). This species isdifferent from other Naegleria spp. because of its verylong ITS1 sequence. Because of the close relationship tothis unique ITS1 sequence of N. chilensis the nameN. neochilensis is proposed for strain IP5.
The sequence of isolate IP7 is closely related to that ofN. dobsoni, but also differs from those of isolates GR11and IP4 (N. neodobsoni n. sp. 5). Isolate IP7 differs by1 bp (indel) in the ITS1 and 14 bp (9 substitutions and 5indels) in the ITS2 from those of N. dobsoni (Table 2).Therefore the new species name N. paradobsoni isproposed.
Phylogenetic analysis
In all trees produced by different methods using the5.8S rDNA sequences strains SP3 (N. polaris n. sp. 1),SP4 (N. neopolaris n. sp. 2), SP5 (N. arctica n. sp. 3) andIP2 (N. neoantarctica n. sp. 6) cluster with N. antarctica
(Fig. 1), forming a new cluster 7. Interestingly, the 5.8Ssequence of all the strains in this cluster starts withTAC, instead of AAC as in most other Naegleria spp. Incluster 2 only N. morganensis and N. sturti have 5.8Ssequences which start with TAC, while in the thirdspecies in cluster 2, N. niuginensis, it starts with AGC. Inaddition the ITS1 of all the strains in the new cluster 7 is36 bp long and starts with CC, which is different from allother Naegleria spp.
Strain IP5 (N. neochilensis n. sp.7) clusters withN. chilensis (Fig. 1) in cluster 6. Both have a very longITS2, while in both strains the 5.8S sequence startswith AAT. Both these features are unique amongstNaegleria spp.
Strains GR11 (N. neodobsoni n. sp. 5) and IP7(N. paradobsoni n. sp. 8) cluster with N. dobsoni
(Fig. 1) in all phylogenetic methods used, forming anew cluster 8.
Strain SP8 (N. spitzbergenensis n. sp. 4) is branchingnear the cluster containing N. dobsoni and the onecontaining N. niuginensis (Fig. 1). In the latter the 5.8Sstarts with AGC, which is the only Naegleria sp. withthis 5.8S start. But the 5.8S sequence of strain SP8 startswith AAC, as in most Naegleria spp., also includingN. dobsoni. The ITS1 and ITS2 sequences of strain SP8are more related to those of N. dobsoni, although theITS2 is much shorter in strain SP8 than in N. dobsoni.
In trees based on the alignment of the ITS1, 5.8S andITS2 sequences the differences between the old and newspecies within the clusters concerned are more easilyseen (Fig. 2).
Group I introns
Only strain IP4 of N. neodobsoni n. sp. 5 and strainIP7 of N. paradobsoni n. sp. 8 have a group I intron inthe SSU rDNA (Table 2).
Discussion
This is the first time that vahlkampfiids have beenreported from Arctic region samples. All the vahlk-ampfiids that I isolated from the Arctic and sub-Antarctic region samples turned out to belong to thegenus Naegleria although about half of them (55%)could not be induced to form flagellates. Some isolatesdid not transform by suspending them in distilled water,but formed flagellates in the liquid that accumulates atthe border between the agar surface and the agar blockon which they are transferred for subculturing, asreported earlier (De Jonckheere 2002). Flooding theagar plates with an E. coli suspension induced enfla-gellation in N. fultoni, while the standard procedure didnot (De Jonckheere et al. 2001). At the same time bothof these procedures failed to induce enflagellation inN. chilensis and N. indonesiensis. In N. angularis
flagellates were only obtained by suspending thetrophozoites in Prescott’s and James’s solution (DeJonckheere and Brown 2005b). It is obvious thatconditions for enflagellation differ according to thespecies and even in different isolates of the same species(De Jonckheere et al. 2001; Behets et al. 2003).
It has been shown that in almost all species of thegenus Naegleria different strains within the same specieshave identical ITS2 sequences (De Jonckheere 2004).Only in two species, N. byersi and N. andersoni, was adifference of 1 bp detected in the ITS2 of one straincompared with all strains of that species. In allspecies, except in N. fowleri, the ITS1 and 5.8Ssequences are identical in different strains withinthe same species. Based on ITS1, 5.8S and ITS2sequences none of the strains isolated during the presentinvestigation belong to already described Naegleria
spp. However, the new isolates are related to threespecies with low maximum temperature tolerance, i.e.N. antarctica (28 1C), N. chilensis (30 1C) and N. dobsoni
(o35 1C). None of the new isolates grows at 37 1C, andat 30 1C only strains SP8 (N. spitzbergenensis), GR11(N. neodobsoni) and IP7 (N. paradobsoni) grow. Theclosest relative of the three latter strains is N. dobsoni,which is reported to grow ato35 1C, so this species does

ARTICLE IN PRESS
N. pageiN. gruberiN. endoiN. laresiN. tihangensisN. australiensisN. mexicanaN. schusteriN. gallicaN. pringsheimiN. italicaN. clarkiN. philippinensisN. byersiN. americanaN. galeacystisN. fultoniN. indonesiensisN. robinsoniN. dobsoniN. neodobsoni GR11N. paradobsoni IP7N. carteriN. antarcticaN. arctica SP5N. polaris SP3N. neopolaris SP4N. neoantarctica IP2N. sturtiN. morganensisN. niuginensisN. spitzbergenensis SP 8N. jamiesoniN. and ersoni
N. martineziN. lovaniensisN. johanseniN. fowleriN. pussardiN. angularisN. chilensisN. neochilensis IP5
5
1
2
4
6
8 (new)
7 (new)
N. minorN. jadini
Fig. 1. Unrooted phylogenetic tree inferred from comparisons of the 5.8S rDNA sequences using the Kimura two-parameter
correction and the UPGMA method. Numbers at the right indicate previous clusters, based on both SSU and 5.8S rDNA analyses,
and the two new clusters. Arrows indicate the two strains that form cluster 3 based on SSU rDNA, but not 5.8S analysis.
J.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123120
grow at 30 1C as well. One of the related species,N. chilensis, is one of the few species that is reported notto transform into flagellates (De Jonckheere et al. 2001).
All phylogenetic analysis methods give similar resultsand show 2 new clusters within the genus Naegleria.Four of the new species form a new cluster withN. antarctica. The latter species was outside any clusterin trees based on rDNA phylogeny (De Jonckheere2004). Therefore, this branching is considered a newcluster 7 (Fig. 1). Also N. dobsoni is not in any cluster intrees based on rDNA phylogeny. As two new species
branch with N. dobsoni these three species are con-sidered to constitute a new cluster 8. One new species isbranching within cluster 6, containing N. pussardi,N. chilensis, and the recently described N. angularis
(De Jonckheere and Brown 2005b). This is the mostdistantly related cluster within the genus Naegleria.
Only strain IP4 of N. neodobsoni n. sp. 5 and strainIP7 of N. paradobsoni n. sp. 8 have a group I intronin the SSU rDNA. In this new cluster 8 the closelyrelated N. dobsoni does not have a group I intron, whilealso strain GR11 of N. neodobsoni, which has identical

ARTICLE IN PRESS
N. antarctica
N. neoantarctica IP2
N. arctica SP5
N. polaris SP3
N. neopolaris SP4
N. niuginensis
N. spitzbergenensis SP8
N. dobsoni
N. neodobsoni GR11
N. paradobsoni IP7
N. chilensis
N. neochilensis IP5
100
100
100
100
100
100
96
61
93
94
8 (new)
7 (new)
6
Fig. 2. Unrooted phylogenetic tree inferred from comparisons
of the ITS1-5.8S-ITS2 rDNA sequences using the Kimura two-
parameter correction and the UPGMA method. The whole
ITS1 sequence is included except for N. chilensis and N.
neochilensis, where the majority of the sequence is omitted
because it is about five times longer in these two species than in
the others. Bootstrap values (100 replicates) are shown above
nodes. Clusters are indicated as obtained with the 5.8S rDNA
phylogeny (see Fig. 1).
J.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123 121
ITS1-5.8S-ITS2 sequences to strain IP4, does not have agroup I intron. Except in cluster 5, intron+ species arenot usually present together with intron� species in thesame cluster.
Two of the new species (N. polaris n. sp. 1 andN. neodobsoni n. sp. 5) were found in both Arctic andsub-Antarctic samples. The new species neoantarctica
and neopolaris are closely related to the antarctica andpolaris species. All this indicates that there is only alimited gene pool of the genus Naegleria present in thesecold environments.
In a previous study (Brown et al. 1982), fourNaegleria strains and six Vahlkampfia strains wereisolated but the species identity is not known asmolecular typing methods for vahlkampfiids based onrDNA sequencing (De Jonckheere 1998; De Jonckheereand Brown 2005a) were not available at the time. Thesemolecular identification methods later demonstratedthat the genus Vahlkampfia as previously recognisedconsists of different genera, and that several speciesbelong to the genus Tetramitus. The only two previouslydescribed vahlkampiids from Antarctica turned out bymolecular typing to be a new species of Tetramitus
(Murtagh et al. 2002) and a new species of Vahlkampfia
(Garstecki et al. 2005). It comes as no surprise that allthe Naegleria isolates recovered from both the Arctic
and sub-Antarctic regions represent new species as well,because all these isolates only grow at low temperature,while the majority of already described Naegleria spp.tolerate rather high maximum temperatures.
Diagnostic summaries
Naegleria polaris n. sp. 1
The trophozoites have the typical appearance ofNaegleria spp. but only one out of three isolates couldbe induced to transform into flagellates. The cysts have around to angular shape with pores in the walls. Thestrains grow at RT, not at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS2sequence, which differs by a long deletion from that ofN. antarctica. The ITS1 and 5.8S rDNA sequences areidentical to those of N. antarctica. The sequence of thetype strain SP3 is available from EMBL under accessionno. AM157657.
Strains of this new species were isolated both from theArctic (Spitzbergen and Greenland) and sub-Antarctic(Ile de la Possession). The name N. polaris is chosenbecause strains of this new species are found in bothpolar regions.
Naegleria neopolaris n. sp. 2
The trophozoites have the typical appearance ofNaegleria spp. and they could be induced to transforminto flagellates. The cysts have a mostly round shapewith pores in the walls. The only known strain grows atRT, not at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS2sequence, which differs by 8 bp (substitutions) fromthat of N. polaris n. sp. 1. The ITS1 and 5.8S rDNAsequences are identical to those of N. polaris n. sp. 1 andN. antarctica. The sequence of the type strain SP4 isavailable from EMBL under accession no. AM157658.
The type strain SP4 was isolated from the Arctic(Spitzbergen). The name N. neopolaris is chosen becauseof the close ITS2 sequence resemblance with that ofN. polaris.
Naegleria arctica n. sp. 3
The trophozoites have the typical appearance ofNaegleria spp. but they could not be induced totransform into flagellates. The cysts have a mostlyround shape with pores in the walls. The only knownstrain grows at RT, not at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS2

ARTICLE IN PRESSJ.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123122
sequence, which differs by 39 bp (24 substitutions and 15indels) from that of N. antarctica. The ITS1 and 5.8SrDNA sequences are identical to those of N. antarctica.The sequence of the type strain SP5 is available fromEMBL under accession no. AM157659.
The type strain SP5 was isolated from the Arctic(Spitzbergen). The name N. arctica is chosen as it wasonly isolated from the Arctic.
Naegleria spitzbergenensis n. sp. 4
The trophozoites have the typical appearance ofNaegleria spp. and could be induced to transform intoflagellates. The cysts have a mostly round shape withpores in the walls. The only known strain grows at RTand at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS1,5.8S and ITS2 sequence. Its 5.8S sequence is mostclosely related to N. niuginensis and N. dobsoni, differingby 4 (2.3%) and 6 bp substitutions (3.5%), respectively.The sequence of the type strain SP8 is available fromEMBL under accession no. AM157660.
The type strain SP8 was isolated from the Arctic(Spitzbergen). The name N. spitzbergenensis is chosenbecause of its geographic origin.
Naegleria neodobsoni n. sp. 5The trophozoites have the typical appearance of
Naegleria spp. but only one of the two isolates (thetype strain GR11) could be induced to transform intoflagellates. The cysts have a round to angular shape withpores in the walls. The strains grow at RT, and the typestrain GR11 also grows at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS1and ITS2 sequences, which differ by 3 bp (substitutions)and 4 bp (2 substitutions and 2 indels), respectively,from that of N. dobsoni. The 5.8S rDNA sequence isidentical to that of N. dobsoni. The sequence of the typestrain GR11 is available from EMBL under accessionno. AM157661.
While strain IP4 has a group I intron in the SSUrDNA, strain GR11 does not.
Strains of this new species were isolated both fromthe Arctic (Greenland) and sub-Antarctic (Ile de laPossession). The name is chosen because of the closesequence resemblance of the ITS1 and ITS2 with thoseof N. dobsoni.
Naegleria neoantarctica n. sp. 6
The trophozoites have the typical appearance ofNaegleria spp. and could be induced to transform intoflagellates. The cysts have a round to angular shape with
pores in the walls. The only known strain grows at RT,not at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS2sequence, which differs by 4 bp (3 substitutions and 1indel) from N. antarctica. The ITS1 and 5.8S rDNAsequences are identical to those of N. antarctica. Thesequence of the type strain IP2 is available from EMBLunder accession no. AM157662.
The type strain IP2 was isolated from Ile dela Possession in the sub-Antarctic. The name N.
neoantarctica is chosen because of the close sequenceITS2 resemblance with that of N. antarctica.
Naegleria neochilensis n. sp. 7
The trophozoites have the typical appearance ofNaegleria spp. but could not be induced to transforminto flagellates. The cysts have a round to angular shapewith pores in the walls. The only known strain grows atRT, not at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS1, 5.8Sand ITS2 sequences. Although its closest relative is N.
chilensis their 5.8S sequence differs by 2bp (1 substitutionand 1 indel) (1%). The sequence of the type strain IP5 isavailable from EMBL under accession no. AM157663.
Only one strain is known. The type strain IP5 wasisolated from Ile de la Possession in the sub-Antarctic.The name N. neochilensis is chosen because the ITS1,5.8S and ITS2 sequence closely resembles that ofN. chilensis. Both of these species have a long ITS1sequence, which is unique amongst Naegleria spp.
Naegleria paradobsoni n. sp. 8The trophozoites have the typical appearance of
Naegleria spp. but could not be induced to transforminto flagellates. The cysts have a round to angular shapewith pores in the walls. The only known strain grows atRT and at 30 1C.
Because of the morphological similarity of the cysts tothose of other Naegleria spp. molecular identification isrequired. The species can be identified from the ITS1and ITS2 sequences, which differ by 1 bp (indel) and14 bp (9 substitutions and 5 indels), respectively, fromthose of N. dobsoni. The 5.8S rDNA sequence isidentical to that of N. dobsoni. The sequence of thetype strain IP7 is available from EMBL under accessionno. AM157664.
The only known strain, IP7, has a group I intronin the SSU rDNA. The type strain IP7 was isolatedfrom Ile de la Possession in the sub-Antarctic. Thename N. paradobsoni is chosen because of the closesequence resemblance of the ITS1 and ITS2 with thoseof N. dobsoni.

ARTICLE IN PRESSJ.F. De Jonckheere / European Journal of Protistology 42 (2006) 115–123 123
Acknowledgements
I thank L. Beyens and his collaborators (PolarEcology, Limnology and Paleobiology, University ofAntwerp) for providing the samples from the Arctic andsub-Antarctic regions.
References
Behets, J., Seghi, F., Declerck, P., Verhelst, L., Duvivier, L.,
Van Damme, A., Ollevier, F., 2003. Detection of Naegleria
spp. and N. fowleri: a comparison of flagellation tests,
ELISA and PCR. Water Sci. Technol. 147, 117–122.
Brown, T.J., Cursons, R.T.M., Keys, E.A., 1982. Amoebae
from Antarctic soil and water. Appl. Environ. Microbiol.
44, 491–493.
De Jonckheere, J.F., 1994. Evidence for the ancestral origin of
group I introns in the SSUrDNA of Naegleria spp. J.
Eukaryotic Microbiol. 41, 457–463.
De Jonckheere, J.F., 1998. Sequence variation in the ribosomal
internal transcribed spacer, including 5.8S, of Naegleria
spp. Protist 149, 221–228.
De Jonckheere, J.F., 2002. A century of research on the
amoeboflagellate genus Naegleria. Acta Protozool. 41,
309–342.
De Jonckheere, J.F., 2004. Molecular definition and the
ubiquity of species in the genus Naegleria. Protist 155,
89–103.
De Jonckheere, J.F., Brown, S., 2005a. The identification of
vahlkampfiid amoebae by ITS sequencing. Protist 156,
89–96.
De Jonckheere, J.F., Brown, S., 2005b. Description of a new
species with a remarkable cyst structure in the genus
Naegleria: Naegleria angularis n. sp. Acta Protozool. 44,
61–65.
De Jonckheere, J.F., Brown, S., Dobson, P.J., Robinson, B.S.,
Pernin, P., 2001. The amoeba-to-flagellate transformation
test is not reliable for the diagnosis of Naegleria spp.
Description of three new Naegleria spp. Protist 152,
115–121.
Felsenstein, J., 1989. PHYLIP: phylogenetic inference package
(version 3.2). Cladistics 5, 164–166.
Garstecki, T., Brown, S., De Jonckheere, J.F., 2005. Descrip-
tion of Vahlkampfia signyensis n. sp. (Heterolobosea), based
on morphological, ultrastructural and molecular character-
istics. Eur. J. Protistol. 41, 119–127.
Hugo, E.R., Stewart, V.J., Gast, R.J., Byers, T.J., 1992.
Purification of amoeba mtDNA using the UNSET proce-
dure. In: Soldo, A.T., Lee, J.J. (Eds.), Protocols in
Protozoology. Allen Press, Lawrence, KS, p. D-7.1-2.
Murtagh, G.J., Dyer, P.S., Rogerson, A., Nash, G.V.,
Laybourn-Parry, J., 2002. A new species of Tetramitus in
the benthos of a saline Antarctic lake. Eur. J. Protistol. 37,
437–443.
Page, F.C., 1974. A further study of taxonomic criteria for
limax amoebae, with descriptions of new species and a key
to genera. Arch. Protistenkd. 116, 149–184.
Page, F.C., 1988. A New Key to Freshwater and
Soil Gymnamoebae. Freshwater Biological Association,
Ambleside.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F.,
Higgins, D.G., 1997. The ClustalX windows interface:
flexible strategies for multiple sequence alignment aided by
quality analysis tools. Nucleic Acids Res. 24, 4876–4882.





























