Embed Size (px)
Citation preview

Isolation and Characterization of Naphthalene-Degrading Bacteria from Sediments ofCadiz Area (SW Spain)
D. Nair,1 F. J. Fernandez-Acero,2 E. Garcıa-Luque,1,3 I. Riba,1,3 T. A. Del Valls1,3
1Catedra UNESCO/UNITWIN/WiCop, Facultad de Ciencias del Mar y Ambientales,Universidad de Cadiz, Polıgono Rıo San Pedro s/n 11510 Puerto Real (Cadiz), Espana
2Laboratorio de Microbiologıa, Facultad de Ciencias del Mar y Ambientales, Universidad deCadiz, Polıgono Rıo San Pedro s/n 11510 Puerto Real (Cadiz), Espana
3Departamento de Quımica Fısica, Facultad de Ciencias del Mar y Ambientales, Universidadde Cadiz, Polıgono Rıo San Pedro s/n 11510 Puerto Real (Cadiz), Espana
Received 4 October 2007; revised 4 March 2008; accepted 16 April 2008
ABSTRACT: Petroleum hydrocarbon contamination of harbor sediments from shipping activity, fuel oilspills, and runoffs are becoming a great concern because of the toxicity and recalcitrance of many of thefuel components. Polycyclic aromatic hydrocarbons (PAHs) are of most concern due to their toxicity, lowvolatility, resistance to degradation, and high affinity for sediments. Microorganisms, especially bacteria,play an important role in the biodegradation of these hydrocarbons. The objective of the present studywas to characterize and isolate PAH-(naphthalene) degrading bacteria in the coastal sediments of Cadiz(SW Spain), since this area is mostly polluted by PAH occurrence. A total of 16 naphthalene-utilizing bac-teria were isolated from these sites. Introduction of bacteria isolated from contaminated sediments intomineral medium contributed to the increased rate of hydrocarbon utilization. The bacterial isolatesobtained from these sites are very potent in utilizing naphthalene and crude oil. It would be interesting toassess if the selected naphthalene-degrading isolates may degrade other compounds of similar structure.Hence these isolates could be very helpful in bioremediating the PAH-contaminated sites. Further pursueon this work might represent eco-friendly solution for oil contamination on sea surface and coastal area.# 2008 Wiley Periodicals, Inc. Environ Toxicol 23: 576–582, 2008.
Keywords: polycyclic aromatic hydrocarbons; naphthalene; biodegradation; bacteria; Cadiz
INTRODUCTION
The degradation of polycyclic aromatic hydrocarbons
(PAHs) by bacteria has been widely studied as many pure
cultures have been isolated and characterized for their
ability to grow on PAHs over many previous investigators
(Chadhain et al., 2006). The extensive investigation on
PAHs degradation still provide limited information about
the diversity of microbes involved in PAH degradation in
the environment. Bioremediation of PAH-polluted soil is
severely hampered by the low rate of degradation of the
higher PAHs, particularly the four- and five-ring PAH, due
to their very low water solubility and are often tightly
bound to soil particles. This results in very low bioavaila-
bility for bacterial degradation (Kotterman et al., 1998).
PAH concentrations in the environment vary widely,
depending on the proximity of the contaminated site to the
Correspondence to: E. G. Luque; e-mail: [email protected]
Contract grant sponsor: Spanish Ministry of Science and Education.
Contract grant numbers: CTM2005-07282-C03-C01/TECNO, VEM2003-
20563/INTER.
Contract grant sponsor: ERASMUS MUNDUS.
Published online 4 June 2008 in Wiley InterScience (www.interscience.
wiley.com). DOI 10.1002/tox.20408
�C 2008 Wiley Periodicals, Inc.
576

production source, the level of industrial development, and
the modes of PAH transport. Soil and sediment PAH con-
centrations at contaminated and uncontaminated sites rang-
ing from 1 lg kg21 to over 300 g kg21 have been reported
(Kanaly and Harayama, 2000). In the open oceans, PAHs
carried in the air are likely to be the main inputs. However,
other more localized inputs of PAHs include waterborne
industrial effluents (particularly from the metal and oil
industries), discharges of oil from shipping, and emissions
and flaring operations by the offshore oil and gas industry.
PAHs are always present as complex mixtures in PAH-
contaminated soils and sediments, and many organisms
isolated from contaminated systems have been found to
grow on a number of different PAHs as sole carbon sources
(Singleton et al., 2005). Several novel marine PAH
degraders, e.g., Cycloclasticus spp. and Neptunomonasnaphthovorans, have also been isolated from contaminated
sediments (Dyksterhouse et al., 1995; Geiselbrecht et al.,
1996; Hedlund et al., 1999). Marine contamination of water
and sediments has become a prime concern due to continu-
ous increase of maritime traffic and accidents in European
Atlantic waters. The English Channel and waters around
Galicia in Spain were the areas with most accidents. Recent
black tides from the Erika and Prestige vessels provided
new evidence for the high risk of accidents with serious ec-
ological impact, which is historically the most important oil
spill hotspot worldwide (Vieites et al., 2004). The interest in
the biodegradation mechanisms of PAHs is initiated because
of its wide ubiquitous distribution in marine ecosystem,
their low availability in soil, high hydrophobicity, where the
aqueous solubility of PAHs decreases almost logarithmi-
cally with increasing molecular mass, and solid–water ratio
consequences (Anders et al., 2005). The biodegradation of
PAHs has been reported to occur under anaerobic, sulfate
reducing and denitrifying conditions (Galushko et al., 1999;
Rockne et al., 2000; Karthikeyan and Bhandari, 2001). The
most important hydrocarbon-degrading bacteria in both ma-
rine and soil environments are Achromobacter spp., Acine-tobacter spp., Alcaligenes spp., Arthrobacter spp., Bacillusspp., Flavobacteriu spp., Nocardia spp. and Pseudomonasspp., and the coryneforms (Leahy and Colwell, 1990; Chung
and King, 2001). Daane et al. (2001) reported that wetlands
show a higher biological activity than many other ecosys-
tems and support enhanced biotransformation of toxic
chemicals. Considering the potential role of bacteria in
PAHs biodegradation, the present location was worth inves-
tigating as source of PAH-degrading bacteria.
Biological methods have an edge over the physicochem-
ical treatment regimes in removing spills since they offer
in situ biodegradation of PAH compounds and oil fractions
by the microorganisms. Thus, this research work explores
the isolation and characterization of isolates that could be
useful to degrade naphthalene by indigenous bacteria from
PAH-contaminated sediments of Cadiz and Algeciras areas
(South West of Spain).
MATERIALS AND METHODS
Isolation of Naphthalene Degrading Bacteria
Sampling for sediment and water was conducted during
September 2006. All the samples were aseptically collected
from different sites of Algeciras and Cadiz areas (SW
Spain) (Fig. 1).
Cadiz Area
Sampling stations: (i) ‘‘Cadiz-1’’ (Ca-1), a clean beach area,
considered as control site. (ii) ‘‘Cadiz-2’’ (Ca-2), a harbor
area, with Ro–Ro vessels, and fishing as main activities.
Algeciras Area
The port of Algeciras is one of the busiest ports in the world
with regard to maritime traffic (oil tankers, passengers’ fer-
ries, Ro/Ro vessels). Two rivers flow into the bay of Alge-
ciras, Palmones River and Guadarranque River. Sampling
stations: (i) ‘‘Palmones Estuary’’ (P-1) station is located in
the middle section of the Palmones River, (ii) ‘‘Guada-
rranque Estuary I’’ (GR30), and (iii) ‘‘Guadarranque Estuary
II’’ (GR4): both stations are located in the mouth of
Guadarranque estuary (which is polluted by industrial
effluents), close to Bay of Algeciras. The samples were
collected from the banks of opposite side of the river.
In all cases, water samples were transferred into polycar-
bonate bottles and the sediment samples were aseptically
transferred into sterile polythene bags and brought to the
laboratory for the study. The samples were stored in the
dark at 48C for further analysis. The pH and temperature of
the sediment samples were checked from the sampling site.
Salinity was measured with an induction salinometer
(Beckman, RS-10) in the laboratory.
Sediment samples were suspended in normal sterile sa-
line (0.85%) at the ratio of 1:100 (w/v) and incubated on the
shaker for 1 h at 150 rpm. The suspension was allowed to
settle down and 0.1 mL of clear supernatant was used to
carry out serial dilutions up to 105 for nutrient agar (NA) and
up to 102 for mineral salt medium (MSM agar: FeSO4, 0.12
Fig. 1. General areas sampled and locations of the five fieldsampling stations.
577NAPHTALENE-DEGRADING BACTERIA FROM CADIZ AREA
Environmental Toxicology DOI 10.1002/tox

3 g L21; K2HPO4, 12.60 g L21; KH2PO4, 3.64 g L21;
NH4NO3, 2.0 g L21; MgSO4�7H2O, 0.2 g L21; MnSO4,
0.0012 g L21; Na2MoO4�2H2O, 0.0012 g L21; CaCl2, 0.15 g
L21). Viable count was determined by spread-plating super-
natant directly as well as its dilutions on NA prepared with
artificial sea water for heterotrophic total counts and MSM
agar containing 0.01% naphthalene (Scharlau Chemie, Bar-
celona, Spain) as well as in MSM agar with 0.2 g of naphtha-
lene added to the lid of Petri plates, and then the plates were
inverted and sealed with parafilm so that the organisms could
utilize naphthalene in vapor phase for naphthalene-degra-
ding organisms (Guerin and Boyd, 1992). NA plates were
incubated at room temperature for 24 h and MSM plates
were incubated for 7 days at 228C. The distinct morphologi-
cal colonies from MSM media were picked, and the isolates
were further purified and maintained on MSM agar with
0.2% naphthalene for further studies.
Characterization and Identification of Isolates
After plating the samples, the isolates obtained on NA and
MSM agar plates were picked and transferred to new plates,
which were incubated at room temperature for 1 week. The
growth rate and morphological characteristics of the iso-
lates was observed. These selected isolates were further
characterized using different percentages of naphthalene.
The obtained isolates were plated on MSM agar with diffe-
rent concentrations of naphthalene, based on their growth
with high percentage of naphthalene (up to 1%) on agar
and their growth on broth with naphthalene was the criteria
used for the selection of the potent isolates.
Growth curve study was carried out by selecting all the
selected isolates and were inoculated in MSM broth with
1% naphthalene and incubated at 288C on the shaker at 150
rpm. The growth was measured after every 24 h at 600 nm
for 3 days. As negative control, the cultures from Spanish
Type Culture Collection (CECT, in Spanish initials): Baci-llus spp. CECT 40, Pseudomonas fluorescens CECT 378,
and E. coli CECT 101 were also inoculated.
Considering the importance of these isolates showing
interesting growth and utilization of higher percentage of
naphthalene, and to confirm their identity based on the mo-
lecular level, isolates were sent out for further identification
to Spanish Culture Collection Center (CECT). The cultures
were identified on the basis of 16S rRNA homology analysis.
RESULTS AND DISCUSSION
To check the total heterotrophic bacterial count of the sedi-
ment, the diluted sediment samples were plated on NA. It
was expected that different kinds of microorganisms would
grow, which gives a total viable count (TVC) of bacteria in
the sediment sample with its enormous diversity. The TVC
was ranging from 1.5 3 104 to 1.93 3 106, expressed in cfu
g21 of sediment. ‘‘GR3’’ site was found to be the most
microbiologically diverse location among the sampling
sites, which may be due to the very high fluctuations in
salinity, wherein all kinds of bacteria which are tolerant to
wide salinity range could grow and also due to the number
of industries situated around this area. As has been previ-
ously reported (Shiaris, 1989a) for ecosystems with low
salinity estuarine sediments subject to salt water intrusion,
it was observed that phenanthrene degraders are tolerant to
a wide range of salinities. As the first objective of the study
was to screen the PAH degrading bacteria, the same sedi-
ment samples were plated on MSM to check the TVC with
PAH (0.01% of naphthalene) and it was expected that only
those bacteria which can utilize PAH as carbon source
would grow. A significant number of bacteria were
observed growing solely on the selected PAH-containing
MSM agar. The ‘‘GR3’’ site was found to be most diversely
populated with PAH-tolerant bacteria, which in turn sub-
stantiate the fact that the estuarine soil is a rich source of
diverse microorganisms (Shiaris, 1989a,b). This work point
out clearly that the ability to utilize hydrocarbons is widely
distributed among diverse microbial populations, as direct
isolation method is used to isolate dominant members in a
microbial community (Zhuang et al., 2002). Hydrocarbons
are naturally occurring organic compounds and, therefore,
it is not surprising that microorganisms have evolved with
the ability to utilize these compounds. When natural eco-
systems are contaminated with petroleum hydrocarbons,
the indigenous microbial communities are likely to contain
microbial populations of differing taxonomic relationships,
which are capable of degrading the contaminating hydro-
carbons (Leahy and Colwell, 1990). The isolation of naph-
thalene-degrading bacteria was done by two methods, viz.,
incorporating naphthalene into medium and introducing
naphthalene vapors as carbon source. Although the method
of isolation was different, the results obtained (TVC)
appeared with minor differences. Thus, this result reinfor-
ces a paradigm of environmental microbiology that diffe-
rent isolation strategies can result in the isolation of different
bacteria with the same physiological characteristic of inter-
est. A total of 442 colonies were obtained from NA plates
and 189 colonies from MSM agar from all the sites. The
colonies with varied morphology were also obtained with
size ranging from pinpoint to over 3-mm diameter. Out of
this, only 16 colonies were selected for further studies since
many were identical. These cultures were subcultured later
and maintained on the MSM agar and NA.
The Gram and colony characters were noted. Although
the isolation methods could select for both Gram-positive
and Gram-negative bacteria, all candidate strains were Gram-
positive except for one, isolate 7. The dominance of Gram-
positive bacteria should not be surprising, since
Gram-positive bacteria have a stronger cell envelope than
Gram-negative bacteria that allows them to thrive in the
578 NAIR ET AL.
Environmental Toxicology DOI 10.1002/tox
highly variable intertidal sediment environment, where
sediment temperatures are high in the day and osmotic
pressures and nutrient supply may change periodically over
a daily cycle (Zhuang et al., 2003a). Many different species
of bacteria with the ability to degrade naphthalene and
other PAHs have been isolated, mostly from soil environ-
ments (Wilson and Jones, 1993). The majority of the PAH-
degrading bacteria were previously found to belong to the
genus Pseudomonas (Gram-negative) (Garcıa-Valdes et al.,
1988) but from the present study, most of the PAH-degrad-
ing isolates were found to be Gram-positive bacteria.
Many studies have shown that the number of hydrocar-
bon-utilizing microorganisms and their proportion in the het-
erotrophic community increases with exposure to petroleum
or other hydrocarbon pollutants and the levels of hydrocarbon
utilizing microorganisms generally reflect the degree of
contamination of the ecosystem (Leahy and Colwell, 1990;
Wilson and Jones, 1993). The addition of hydrocarbons to an
ecosystem may result in a selective increase in microorga-
nisms capable of utilizing the hydrocarbons and those that are
capable of utilizing metabolites produced by the hydrocar-
bon-degrading bacteria (Venkateswaran et al., 1995; Ferrari
et al., 1996). The enhancement or reduction will depend on
the chemical composition of the contaminating hydrocarbons
and on the species of microorganisms present within the mi-
crobial community of the particular ecosystem (Atlas, 1995).
Several studies have shown a rise in populations of hydrocar-
bon-utilizing microorganisms after oil spills (Atlas, 1981;
Regina et al., 2006). The bacterial cultures that were consis-
tent in growth on the MSM agar with 0.01% naphthalene
medium were selected for further study.
Characterization of the Bacteria with Respectto Utilization of Various NaphthaleneConcentrations
The selected 16 isolates were grown in different concentra-
tion of naphthalene with and without yeast extract
(0.005%) as nutritional supplement. The concentrations of
naphthalene used ranged between 0.01% and 1% in MSM
agar. Most of the isolates showed almost identical pattern
of growth in the assayed conditions. A previous study has
shown that yeast extract supplement initiates the growth of
the bacterial isolates in MSM (Law and Teo, 1997). How-
ever, this study indicated that the presence or absence of
yeast extract did not produce significant effect on the
growth of isolates. For this reason, further studies were
done without the addition of yeast extract.
Growth behavior of all the selected isolates was observed
in 0.01% naphthalene in MSM medium. After 7 days of
incubation, the optical density (OD) was observed and di-
fference in OD was measured as rate of growth in 7 days.
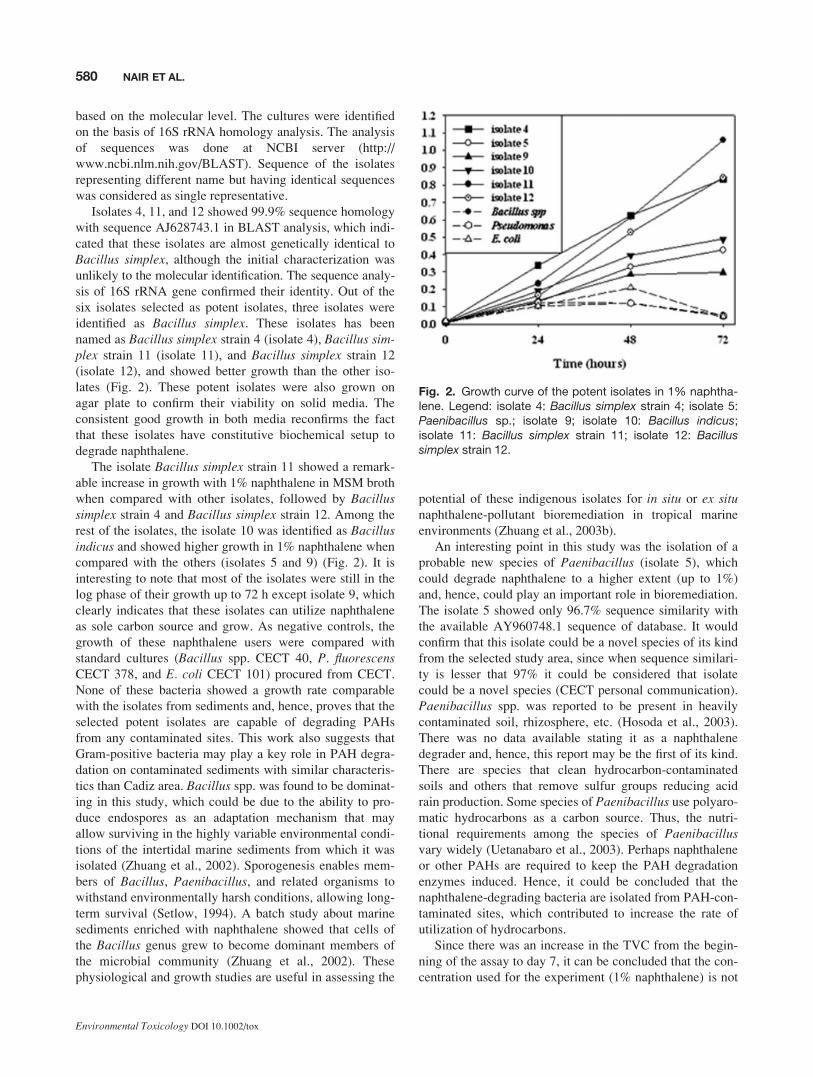
The isolates 4, 5, 9, 10, 11, and 12 grow in much higher rate
than the rest of the isolates (Table I), which could be related
to a potent degradation of naphthalene. Growth characteris-
tics of these isolates also emphasize the fact that these iso-
lates can grow in minimal medium utilizing naphthalene as
sole carbon source. Further increase in concentration of
naphthalene would eliminate most of the loosely resistant
bacteria and screen for naphthalene-degrading bacteria.
Since the isolates showed growth in 1% naphthalene show-
ing high OD they were selected for further study. The rest of
the isolates showed negligible growth.
The isolates which showed a very good growth with
highest percentage of naphthalene on solid media (1%
naphthalene) and the isolates which showed faster growth
in liquid media (0.01% naphthalene) were inoculated in
MSM broth with 1% naphthalene. The OD was assessed
after 7 days of incubation at 288C on the shaker at 150 rpm.
The viability of these isolates was observed with the due
course of incubation with increasing OD. The increasing
number of viable bacterial counts and increasing OD of the
broth clearly indicated that the culture can grow, tolerate,
and utilize up to 1% of naphthalene as carbon source.
The selected seven isolates were sent to the Spanish Cul-
ture Collection Center (CECT) to confirm their identity
TABLE I. The growth of different isolates in 0.5% and 1.0% of naphthalene
Isolates
Concentration of
Naphthalene (%)
OD at 600 nm
(0 h)
TVC at 0 h
(cfu mL21)
OD at 600 nm
(after 7 days)
TVC after 7 days
(cfu mL21)
4 0.5 0.006 2.1 3 102 1.018 8.7 3 105
1.0 0.010 2.0 3 102 0.677 4.3 3 104
5 0.5 0.008 1.8 3 102 0.964 4.9 3 105
1.0 0.010 1.6 3 102 0.248 3.8 3 104
9 0.5 0.004 1.3 3 102 0.045 2.3 3 103
1.0 0.018 1.0 3 102 0.028 1.2 3 102
10 0.5 0.002 1.5 3 102 0.642 3.8 3 104
1.0 0.014 1.2 3 102 0.453 2.7 3 104
11 0.5 0.004 3.1 3 103 1.235 8.9 3 106
1.0 0.013 2.2 3 103 1.062 6.8 3 104
12 0.5 0.003 3.0 3 103 1.156 7.8 3 106
1.0 0.011 2.7 3 103 0.836 5.9 3 104
579NAPHTALENE-DEGRADING BACTERIA FROM CADIZ AREA
Environmental Toxicology DOI 10.1002/tox
based on the molecular level. The cultures were identified
on the basis of 16S rRNA homology analysis. The analysis
of sequences was done at NCBI server (http://
www.ncbi.nlm.nih.gov/BLAST). Sequence of the isolates
representing different name but having identical sequences
was considered as single representative.
Isolates 4, 11, and 12 showed 99.9% sequence homology
with sequence AJ628743.1 in BLAST analysis, which indi-
cated that these isolates are almost genetically identical to
Bacillus simplex, although the initial characterization was
unlikely to the molecular identification. The sequence analy-
sis of 16S rRNA gene confirmed their identity. Out of the
six isolates selected as potent isolates, three isolates were
identified as Bacillus simplex. These isolates has been
named as Bacillus simplex strain 4 (isolate 4), Bacillus sim-plex strain 11 (isolate 11), and Bacillus simplex strain 12
(isolate 12), and showed better growth than the other iso-
lates (Fig. 2). These potent isolates were also grown on
agar plate to confirm their viability on solid media. The
consistent good growth in both media reconfirms the fact
that these isolates have constitutive biochemical setup to
degrade naphthalene.
The isolate Bacillus simplex strain 11 showed a remark-
able increase in growth with 1% naphthalene in MSM broth
when compared with other isolates, followed by Bacillussimplex strain 4 and Bacillus simplex strain 12. Among the
rest of the isolates, the isolate 10 was identified as Bacillusindicus and showed higher growth in 1% naphthalene when
compared with the others (isolates 5 and 9) (Fig. 2). It is
interesting to note that most of the isolates were still in the
log phase of their growth up to 72 h except isolate 9, which
clearly indicates that these isolates can utilize naphthalene
as sole carbon source and grow. As negative controls, the
growth of these naphthalene users were compared with
standard cultures (Bacillus spp. CECT 40, P. fluorescensCECT 378, and E. coli CECT 101) procured from CECT.
None of these bacteria showed a growth rate comparable
with the isolates from sediments and, hence, proves that the
selected potent isolates are capable of degrading PAHs
from any contaminated sites. This work also suggests that
Gram-positive bacteria may play a key role in PAH degra-
dation on contaminated sediments with similar characteris-
tics than Cadiz area. Bacillus spp. was found to be dominat-
ing in this study, which could be due to the ability to pro-
duce endospores as an adaptation mechanism that may
allow surviving in the highly variable environmental condi-
tions of the intertidal marine sediments from which it was
isolated (Zhuang et al., 2002). Sporogenesis enables mem-
bers of Bacillus, Paenibacillus, and related organisms to
withstand environmentally harsh conditions, allowing long-
term survival (Setlow, 1994). A batch study about marine
sediments enriched with naphthalene showed that cells of
the Bacillus genus grew to become dominant members of
the microbial community (Zhuang et al., 2002). These
physiological and growth studies are useful in assessing the
potential of these indigenous isolates for in situ or ex situnaphthalene-pollutant bioremediation in tropical marine
environments (Zhuang et al., 2003b).
An interesting point in this study was the isolation of a
probable new species of Paenibacillus (isolate 5), which
could degrade naphthalene to a higher extent (up to 1%)
and, hence, could play an important role in bioremediation.
The isolate 5 showed only 96.7% sequence similarity with
the available AY960748.1 sequence of database. It would
confirm that this isolate could be a novel species of its kind
from the selected study area, since when sequence similari-
ty is lesser that 97% it could be considered that isolate
could be a novel species (CECT personal communication).
Paenibacillus spp. was reported to be present in heavily
contaminated soil, rhizosphere, etc. (Hosoda et al., 2003).
There was no data available stating it as a naphthalene
degrader and, hence, this report may be the first of its kind.
There are species that clean hydrocarbon-contaminated
soils and others that remove sulfur groups reducing acid
rain production. Some species of Paenibacillus use polyaro-
matic hydrocarbons as a carbon source. Thus, the nutri-
tional requirements among the species of Paenibacillusvary widely (Uetanabaro et al., 2003). Perhaps naphthalene
or other PAHs are required to keep the PAH degradation
enzymes induced. Hence, it could be concluded that the
naphthalene-degrading bacteria are isolated from PAH-con-
taminated sites, which contributed to increase the rate of
utilization of hydrocarbons.
Since there was an increase in the TVC from the begin-
ning of the assay to day 7, it can be concluded that the con-
centration used for the experiment (1% naphthalene) is not
Fig. 2. Growth curve of the potent isolates in 1% naphtha-lene. Legend: isolate 4: Bacillus simplex strain 4; isolate 5:Paenibacillus sp.; isolate 9; isolate 10: Bacillus indicus;isolate 11: Bacillus simplex strain 11; isolate 12: Bacillussimplex strain 12.
580 NAIR ET AL.
Environmental Toxicology DOI 10.1002/tox

toxic for the bacteria isolated from PAH-contaminated sedi-
ments. A similar result was also reported by Maleszak et al.
(2004) with crude oil. Bacterial degradation of naphthalene
was not directly quantified because of its volatility and uti-
lization of very high concentrations. Naphthalene solubility
(32 mg L21) (Liu et al., 1995) was high enough to allow
unrestricted growth. However, at high cell density, the
availability of poorly soluble substrates becomes limiting,
because PAHs are utilized only in the dissolved state
(Deziel et al., 1996). The three isolates reported in this
study extend our knowledge of the range of naphthalene-
degrading bacteria found in marine and estuarine
environments.
The isolate 7, identified as Pseudomonas pohangensis,showed 97% similarity with the available sequence. Weong
et al. (2006) have reported P. pohangensis as a hydrocar-
bon-degrading bacteria from Korean soils, but identifying
this species as a naphthalene degrader from Cadiz harbor
has been done for the first time. The isolate 8, identified as
Paenibacillus agaridevorans, showed 97.5% similarity
with the available sequence. P. agaridevorans which was
first reported by Uetanabaro et al. (2003) and Hosoda
(2003) in sediments and rhizosphere was identified in this
study. The isolate 10 was identified as Bacillus indicus with
99% homology with database sequence. It is the first time
that this bacterium is reported as a naphthalene degrader. It
was first isolated and reported by Suresh et al. (2004) from
sand of an arsenic-contaminated aquifer in West Bengal,
India.
This study gives a comparative analysis of genetic and
enzymological evidences based identification of natural iso-
lates obtained from the marine sediments. It also confirmed
the fact that the analysis of conserved sequences is a tool
for bacterial identification although the biochemical method
can also be considered as a basic tool to characterize the
isolates.
CONCLUSIONS
The isolation and characterization of several strains of
naphthalene-degrading bacteria obtained from PAH-conta-
minated marine and estuarine sediments from Cadiz area
has been carried out, which is the first report from this area.
The introduction of bacteria isolated from contaminated
sediments into mineral medium contributed to increased
rate of hydrocarbon utilization and thus any other easily
available carbon source is not required for the growth of
these isolates.
Gram-positive bacteria were found to be predominant in
this study, out of which Bacillus was found to be the pre-
dominant genus as a naphthalene utilizer.
The cultures can grow, tolerate, and utilize up to 1% of
naphthalene as carbon source. This high concentration of
naphthalene is not toxic for the bacteria isolated.
Hence, these isolates can be very helpful in bioremedia-
ting the PAH-contaminated sites and oil spills thus preser-
ving the ecosystem in a natural way. Further pursue on this
work might represent eco-friendly solution for oil contami-
nation on sea surface and coastal area.
REFERENCES
Anders JR, Lukas WY, Hauke H. 2005. Principles of microbial
PAH-degradation in soil-organic compounds in the soil environ-
ment: Formation, potential for re-mobilisation and environmen-
tal significance of bound residues. Environ Pollut 133:71–84.
Atlas RM. 1981. Microbial degradation of petroleum hydrocarbons:
An environmental perspective. Microbiol Rev 45:180–209.
Atlas RM. 1995. Bioremediation of petroleum pollutants. Int Bio-
deterior Biodegrad 21:317–327.
Chadhain SMN, Norman RS, Pesce KV, Kukor JJ, Zylstra GJ.
2006. Microbial dioxygenase gene population shifts during
polycyclic aromatic hydrocarbon biodegradation. Appl Environ
Microbiol 72:4078–4087.
Chung WK, King GM. 2001. Isolation, characterization, and polyar-
omatic hydrocarbon degradation potential of aerobic bacteria
from marine macrofaunal burrow sediments and description of
Lutibacterium anuloederans gen. nov., sp. nov., and Cycloclasti-cus spirillensus sp. Nov. Appl Environ Microbiol 67:5585–5592.
Daane LL, Harjono I, Zylstra GJ, Haggblom MM. 2001. Isolation
and characterization of polycyclic aromatic hydrocarbon-
degrading bacteria associated with the rhizosphere of salt marsh
plants. Appl Environ Microbiol 67:2683–2691.
Dyksterhouse SE, Gray JP, Herwig RP, Lara JC, Staley JT. 1995.
Cycloclasticus pugetii gen. nov., sp. nov., an aromatic hydro-
carbon-degrading bacterium from marine sediments. Int J Syst
Bacteriol 45:116–123.
Deziel E, Paquette G, Villemur R, Lepine OF, Bisaillon JG. 1996.
Biosurfactant production by a soil Pseudomonas strain growing
on polycyclic aromatic hydrocarbons. Appl Environ Microbiol
62:1908–1912.
Ferrari MD, Neirotti E, Albornoz C, Mostazo MR, Cozzo M.
1996. Biotreatment of hydrocarbons from petroleum tank bot-
tom sludge in soil slurries. Biotechnol Lett 18:1241–1246.
Galushko A, Minz D, Schink B, Widdel F. 1999. Anaerobic degrada-
tion of naphthalene by a pure culture of a novel type of marine sul-
phate-reducing bacterium. Environ Microbiol 1:415–420.
Garcia-Valdes E, Cozar E, Rotger R, Lalucat J, Ursing J. 1988.
New naphthalene-degrading marine Pseudomonas strains. Appl
Environ Microbiol 54:2478–2485.
Geiselbrecht AD, Herwig RP, Deming JW, Staley JT. 1996. Enu-
meration and phylogenetic analysis of polycyclic aromatic
hydrocarbon-degrading marine bacteria from puget sound sedi-
ments. Appl Environ Microbiol 62:3344–3349.
Guerin WF, Boyd SA. 1992. Differential bioavailability of soil-
sorbed naphthalene to two bacterial species. Appl Environ
Microbiol 58:1142–1152.
Hedlund BP, Geiselbrecht AD, Bair TJ, Staley JT. 1999. Polycy-
clic aromatic hydrocarbon degradation by a new marine
581NAPHTALENE-DEGRADING BACTERIA FROM CADIZ AREA
Environmental Toxicology DOI 10.1002/tox

bacterium. Neptunomonas naphthovorans gen. nov., sp. nov.
Appl Environ Microbiol 65:251–259.
Hosoda A, Sakai M, Kanazawa S. 2003. Isolation and characteri-
zation of agar-degrading Paenibacillus spp. associated with the
rhizosphere of spinach. Biosci Biotechnol Biochem 67:1048–
1055.
Kanaly RA, Harayama S. 2000. Biodegradation of high-molecular-
weight polycyclic aromatic hydrocarbons by bacteria. J Bacteriol
182:2059–2067.
Karthikeyan R, Bhandari A. 2001. Anaerobic biotransformation of
aromatic and polycyclic aromatic hydrocarbons in soil micro-
cosms: A review. J Hazard Subst Res 3:3–19.
Kotterman MJJ, Vis EH, Field JA. 1998. Successive mineraliza-
tion and detoxification of benzo[a]pyrene by the white rot fun-
gus Bjerkandera sp. strain BOS55 and indigenous microflora.
Appl Environ Microbiol 64:2853–2858.
Law AT, Teo KS. 1997. Oil biodegradation in the Straits of
Malacca: Phenanthrene degradation by AR-3. J Mar Biotechnol
5:162–167,
Leahy JG, Colwell RR. 1990. Microbial degradation of hydrocar-
bons in the environment. Microbiol Rev 54:305–315.
Liu Z, Jacobson AM, Luthy RG. 1995. Biodegradation of naphtha-
lene in aqueous nonionic surfactant systems. Appl Environ
Microbiol 61:145–151.
Maleszak HB, Przestrzelska A, Bieszkiewicz E, Horoch M. 2004.
Biodegradation of petroleum products by microorganisms
adapted to high crude oil concentration in presence of easy
assimilated carbon source. Pol J Microbiol 53:183–187.
Regina OE, Emuobonuvie IF, Roseline UE. 2006. Growth
responses of bacterial isolates on various concentrations of
crude oil. J Am Sci 2:13–16.
Rockne KJ, Chee-Sanford JC, Sanford RA, Helund BP, Staley JT,
Strand SE. 2000. Anaerobic naphthalene degradation by micro-
bial pure cultures under nitrate-reducing conditions. Appl Envi-
ron Microbiol 66:1595–1601.
Setlow P. 1994. Mechanisms which contribute to long term sur-
vival of spores of Bacillus. J Appl Bacteriol 76:49–60.
Shiaris MP. 1989a. Phenanthrene mineralization along a natural
salinity gradient in an urban estuary, Boston Harbor, Massachu-
setts. Microbial Ecol 18:135–146.
Shiaris MP. 1989b. Seasonal biodegradation of naphthalene,
phenanthrene and benzo(a) pyrene in estuarine sediments. Appl
Environ Microbiol 55:1391–1399.
Singleton DR, Powell SN, Sangaiah R, Gold A, Ball LM, Aitken
MD. 2005. Stable-isotope probing of bacteria capable of
degrading salicylate, naphthalene, or phenanthrene in a bioreac-
tor treating contaminated soil. Appl Environ Microbiol 71:
1202–1209.
Suresh K, Prabagaran SR, Sengupta S, Shivaji S. 2004. Bacillusindicus sp. nov., an arsenic-resistant bacterium isolated from an
aquifer in West Bengal, India. Int J Syst Evol Microbiol
54:1369–1375.
Uetanabaro AP, Wahrenburg C, Hunger W, Pukall R, Sproer C,
Stackebrandt E, de Canhos VP, Claus D, Fritze D. 2003. Paeni-bacillus agarexedens sp. nov., nom. rev., and Paenibacillusagaridevorans sp. nov. Int J Syst Evol Microbiol 53:1051–
1057.
Venkateswaran K, Hoaki T, Kato M, Maruyama T. 1995. Micro-
bial degradation of resins fractionated from Arabian light crude
oil. Can J Microbiol 41:418–424.
Vieites DR, Roman SN, Palanca A, Ferrer X, Vences M. 2004.
European Atlantic: The hottest oil spill hotspot worldwide.
Naturwissenschaften 91:535–538.
Weong H, Yoo SH, Baek YK, Lee SY, Kwon SW, Go SJ, Stacke-
brandt E. 2006. Pseudomonas pohangensis sp. nov. isolated
from seashore sand in Korea. Int J Syst Evol Microbiol
56:2153–2156.
Wilson SC, Jones KC. 1993. Bioremediation of soil contaminated
with polynuclear aromatic hydrocarbons (PAHs): A review.
Environ Pollut 81:229–249.
Zhuang WQ, Tay ST, Maszenan AM, Tay STL. Bacillus naphtho-vorans sp. nov. from oil-contaminated tropical marine sedi-
ments and its role in naphthalene biodegradation. Appl Micro-
biol Biotechnol 58:547–553, 2002.
Zhuang WQ, Tay JH, Maszenan AM, Krumholz LR, Tay STL.
2003a. Importance of Gram-positive naphthalene-degrading
bacteria in oil-contaminated tropical marine sediments. Lett
Appl Microbiol 36:251–257.
Zhuang WQ, Tay JH, Maszenan AM, Tay ST. 2003b. Isolation of
naphthalene-degrading bacteria from tropical marine sediments.
Water Sci Technol 47:303–308.
582 NAIR ET AL.
Environmental Toxicology DOI 10.1002/tox





![Monitoring Naphthalene Catabolism Bioluminescence with ... · Mineralization. Naphthalene metabolism was monitored byusing a mineralization procedure to measure conversion of["4C]naphthalene](https://img.dokumen.tips/doc/110x75/5e86f458fffce403b43df98f/monitoring-naphthalene-catabolism-bioluminescence-with-mineralization-naphthalene.jpg)























