Embed Size (px)
Citation preview

Interactive effects of elevated CO2, phosphorus deficiency,and soil drought on nodulation and nitrogenase activityin Alnus hirsuta and Alnus maximowiczii
Hiroyuki Tobita & Akira Uemura & Mitsutoshi Kitao &
Satoshi Kitaoka & Hajime Utsugi
Received: 6 June 2009 /Accepted: 16 November 2009 /Published online: 10 December 2009# Springer Science+Business Media B.V. 2009
Abstract We examined the interactive effects of elevatedCO2, soil phosphorus (P) availability, and soil drought onnodulation, nitrogenase activity, and biomass allocation inAlnus hirsuta and Alnus maximowiczii. Potted seedlingswere grown in either ambient or elevated CO2 (36 Pa and72 Pa CO2), with different levels of P (7.7 and 0.77 mgPpot−1 week−1 for high-P and low-P, respectively) and watersupply in a natural daylight phytotron. Measurements ofnitrogenase activity by an acetylene reduction assay failedto reveal significant effects of the treatments in any species.In high-P, nodule biomass increased under elevated CO2
and decreased under drought. In low-P, nodule biomassdecreased substantially compared to high-P, but the effectof elevated CO2 on nodule biomass was unclear. Soildrought increased the partitioning of biomass into nodules,especially in A. hirsuta. These results suggest that with highP availability, elevated CO2 could promote N2 fixation by
increasing nodule biomass even under drought. On theother hand, if soil P is limiting, elevated CO2 may notenhance N2 fixation because of the suppression of growth.
Keywords Alnus .Frankia . Nodulation . CO2.
Phosphorus .Water
1 Introduction
The increasing atmospheric CO2 concentration is an impor-tant factor in global climate change (Houghton et al. 2001).Many plant species grown under elevated CO2 conditionsexhibit enhanced photosynthetic rates and growth when otherenvironmental resources do not limit productivity (Norby et al.1999; Körner 2006). However, because forest ecosystems areusually nitrogen (N)-limited (Vitousek and Howarth 1991),soil N availability might become the limiting factor for growthunder elevated CO2 (Luo et al. 2004). This hypothesis iscalled Progressive Nitrogen Limitation (PNL). It has beentested (Hungate et al. 2006; Norby and Iversen 2006), leading tothe suggestion that an exogenous N input from outside of theforest ecosystems will be required to maintain enhancedgrowth under increased CO2 (Johnson 2006; Reich et al. 2006).
For this reason, symbiotic N2 fixation will become animportant factor in future environments under elevated CO2
(Houghton et al. 2001). As N2-fixers are largely independentof soil N, they may respond to increased CO2 more directlythan non-N2-fixers (Temperton et al. 2003). In fact, increasesof growth, N2 fixation, nitrogenase activity in nodules, andnodule biomass under elevated CO2 have been documented(Norby 1987; Arnon and Gordon 1990; Vogel et al. 1997;Tobita et al. 2005). In temperate to boreal zones, actinorhizalplants including Alnus species can contribute significantamounts of fixed N to forest ecosystems (Dawson 1983).
Presented at the 15th International Frankia and Actinorhizal PlantsMeeting, Bariloche, Argentina, October 19–23, 2008.
H. Tobita (*) :A. Uemura : S. Kitaoka :H. UtsugiHokkaido Research Center,Forestry and Forest Products Research Institute,Sapporo 062-8516, Japane-mail: [email protected]
A. Uemurae-mail: [email protected]
S. Kitaokae-mail: [email protected]
H. Utsugie-mail: [email protected]
M. KitaoForestry and Forest Products Research Institute,Tsukuba 305-8687, Japane-mail: [email protected]
Symbiosis (2010) 50:59–69DOI 10.1007/s13199-009-0037-7

Two subgenera of Alnus, Gymonothyrsus and Alnuster, arenative to Japan (Somego 1985).
N2 fixation in Alnus is influenced by various environ-mental factors (Hungate et al. 2003; Dawson 2008). In thefuture, soil phosphorus (P) availability and drought mightbecome primary problems. In the context of increasedatmospheric N deposition (Vitousek et al. 1997; Hyvönen etal. 2007; Wallace et al. 2007), soil P limitation might occurdue to a disturbed soil nutrient balance (Gress et al. 2007).Actinorhizal plants as well as legumes require phosphate forgrowth and root nodulation (Huss-Danell 1997; Valverde etal. 2002). P deficiency limits nodule formation and N2
fixation of N2-fixing plants even under ambient CO2
conditions (Gentili and Huss-Danell 2003; Edwards et al.2006). It has been argued that the P demand of trees willincrease under increased CO2 (Kogawara et al. 2006).
Regarding drought, it has been predicted that increasingtemperatures will decrease precipitation during the summerseason (Naumburg et al. 2004; Geßler et al. 2007). Droughtmay decrease photosynthetic rates and limit growth throughstomatal closure (Flexas and Medrano 2002), which mayalso affect N2 fixation. When grown under moderatedrought for prolonged periods, some plant species adaptby increasing photosynthetic capacity, the maximum rate ofcarboxylation (Vcmax; Kitao et al. 2003, 2007; Qaderi et al.2006). On the other hand, most plants react to increasedCO2 by photosynthetic down-regulation, a decrease inphotosynthetic capacity (Long et al. 2004; Nowak et al.2004; Ainsworth and Rogers 2007). Therefore, the photo-synthetic response to long term moderate drought is theopposite of the reaction to increased CO2, and it has beensuggested that the effect of increased CO2 on photosynthe-sis could be alleviated by moderate drought (Qaderi et al.2006; Kitao et al. 2007). Photosynthesis indirectly drivesN2 fixation through the provision of energy in the form ofphotosynthates. Therefore, photosynthetic acclimation un-der drought would affect nodule formation and N2 fixation.
We evaluated the effects of elevated CO2, soil Pdeficiency, and drought on N2 fixation in one Alnus speciesof each subgenus to examine, (i) whether nodule activityand nodule biomass can increase under elevated CO2; (ii)whether enhanced plant growth, the change in biomassallocation, or both cause the responses to elevated CO2; and(iii) whether photosynthetic acclimation to drought affectsnodule formation.
2 Material and methods
2.1 Plant material
One Alnus species of each subgenus was investigated. Alnushirsuta Turcz. belongs to the Subgenus Gymonothyrsus and
is a tall tree living at relatively low elevation. The speciesshows successive leaf expansion patterns (Kikuzawa 1983).Alnus maximowiczii belongs to the Subgenus Alnuster whichis a shrub endemic to Japan (Somego 1985); it occurs athigher elevations than A. hirsuta. Alnus maximowiczii showsa heterophyllous leaf expansion pattern and forms shortshoots (Kikuzawa 1983).
One-year-old seedlings of A. hirsuta and A. maximowic-zii obtained from a commercial nursery (Oji Forestry &Landscaping, Sapporo, Japan) were transplanted individu-ally into free-draining 5-liter plastic pots (diameter: 21 cm)filled with 1:1 (v/v) Kanuma pumice and clay loam. Eachpot was placed in a tray to prevent nutrient drainage. Theinitial height of the seedlings was 14–18 cm in bothspecies. Nodule formation in the root systems was observedin both species at the time of transplanting.
2.2 Elevated CO2, soil phosphorus supply, and soil droughttreatments
Seedlings were placed in a natural daylight phytotron (KoitoIndustries, Yokohama, Japan) equipped with CO2 concentra-tion controllers (DAIWA Air Co. Ltd., Sapporo, Japan) atHokkaido Research Center, FFPRI, Sapporo, Japan (43˚N,141˚E; 180 m a.s.l.). Seedlings were grown at 36 Pa(ambient CO2 treatment) or 72 Pa (elevated CO2 treatment)CO2 from mid-May. Each CO2treatment was replicated inthree chambers; details of the CO2 treatments have beendescribed previously (e.g. Tobita et al. 2005; Kitao et al.2007; Watanabe et al. 2008). Twenty-four seedlings perspecies were grown in each chamber. Twelve seedlings weresupplied with phosphorus (P) at 7.74 mg P pot−1 week−1
(high-P treatment), while the other twelve seedlings received0.774 mg P pot−1 week−1 (low-P treatment) in modified 0.5 ×Hoagland solution (Asher and Edwards 1983; 0.3 mMKNO3, 0.2 mM Ca(NO3)2/4 H2O, 0.05 mM NH4H2PO4,2.25 mM KCl, 1.8 mM CaCl2, 0.45 mM KH2PO4, 1 mMMgSO4/7 H2O, 25 µM EDTA-Fe, 4.5 µM MnCl2/4 H2O,23 µM H3BO3, 0.4 µM ZnSO4/7 H2O, 0.15 µM CuSO4/5H2O, 0.007 µM (NH4)6Mo7O24/4 H2O). In order to avoid aninhibition of nodule formation by nitrogen, only one tenth ofthe N of the standard nutrient solution was applied. In thelow-P treatment, KCl, CaCl2, and KH2PO4 were added toprovide the same concentrations of K+ and Ca2+ as in thehigh-P tests. Six seedlings in each P treatment received250 ml water three times per week (well-watered treatment:wet); the other seedlings received no water in addition to thenutrient solution (drought treatment: dry). In order to avoidsalt accumulation, water instead of nutrient solution wasapplied once per month. Air temperature was maintained at26/16˚C (day/night). Predawn leaf water potential wasmeasured with a pressure chamber (Model 3000, Soilmoisture Equipment Corp., Santa Barbara, CA) before and
60 H. Tobita et al.

after the scheduled watering. Volumetric water contents ofthe soil was also measured with a Hydrosense (CampbellScientific, Inc. United States). The seedlings were grown fortwo growing seasons. In accordance with the increased plantsize, the amount of P and water supplied was increased 1.5times in the second season. After defoliation by decreasingroom temperature gradually (20/16, 20/10, and 16/10˚C) inthe first autumn of the experimental period, the seedlingswere maintained in the open during winter. In the secondspring, the seedlings were moved into the chambers againbefore the buds began to open. The initial temperature was16/10˚C and gradually increased to the standard 26/16˚C.
2.3 Nitrogenase activity of nodules
In August of the second season, nitrogen-fixing activity wasmeasured by the acetylene reduction methods (Hardy et al.1973) in six seedlings of each treatment. Before measure-ment, seedlings were kept in their chambers after the rootsystems had been cleaned. Root nodules were excised withsmall segments of intact roots (> 2 cm long) and used tomeasure acetylene reduction activity (ARA). Since manipu-lations have to be minimized to avoid a reduction of noduleactivity (Huss-Danell 1978; Wheeler et al. 1978), theacetylene reduction assays were conducted immediately afterexcision by incubation for 30 minutes with 10% acetylene inair within glass containers of 200 ml capacity. The containerswere maintained at 25˚C in an incubator. The measurementswere taken between 10:00 and 14:00. After the incubationperiod, gas samples (1 mL) were taken from the containerand analyzed. The production of ethylene was determinedusing a gas chromatograph equipped with a flame ionizationdetector (GC-14B, SHIMADZU, Kyoto, Japan). Hydrocar-bon separation was achieved using a 1.0 m glass columnpacked with 100/120-mesh Porapak N. The temperature ofdetection was maintained at 75˚C, and column temperatureat 60˚C. Nitrogen gas was used as carrier. Acetylene gassamples were also checked for potential ethylene contami-nation and the acetylene reduction rates were corrected forthe contamination level. Preliminary studies under constantconditions showed that ethylene production was linear withincubation time for at least 3 hours. One major difficulty ofthe acetylene reduction method is that upon addition ofacetylene to root nodules, nitrogenase activity declineswithin minutes. In Alnus spp., the error from using closedcuvettes for the acetylene reduction assay probably is smallbecause the acetylene-induced decline is small (Schwintzerand Tjepkema 1994).
2.4 Growth and biomass allocation
After the measurement of nitrogenase activity, dry massesof seedling leaves, shoots, stems, coarse roots (> 2 mm),
fine roots (<2 mm), and nodules were determined afterdrying at 80°C, and the total leaf area (LA) was measured.The leaf area ratio (LAR; LA per whole-plant biomass) andnodule weight ratio (NWR; nodule biomass per whole-plantbiomass) of each seedling were calculated.
2.5 Leaf N and P concentration
The leaves of cultivated seedlings for each treatment weremeasured mass based N content by a combustion methodusing an NC-analyzer (Vario Max CN; Elementar Analy-sensysteme GmbH, Hanau, Germany) after dry at 80˚C for48 hr. Ground leaf samples of about 0.1 g were digested by60% HNO3 and 60% HClO4. The amount of P wasdetermined using the Murphy-Riley method (Murphy andRiley 1962; Schoenau and O’Halloran 2008) on a spectro-photometer at 712 nm.
2.6 Photosynthetic properties
Before the measurement of nitrogenase activity, leaf gasexchange measurements were taken on six mature leaves ofeach treatment by using an open gas exchange system (LI-6400; Li-Cor Inc., Lincoln, NE, USA). The age of theleaves was 38 – 56 days (A. hirsuta) and 34 – 46 days (A.maximowiczii). In A. hirsuta, the sixth to eighth leaves,counted from the tip of leader shoots, were used, whereas inA. maximowiczii, the second to third leaves from the base ofleader shoots were examined. The full light-saturated netphotosynthetic rates per leaf area (Asat) versus internal CO2
(Ci) curves were determined. Saturating photon flux density(PFD) at the upper leaf surface was 1,200 µmol m−2 s−1, asdetermined based on light response curves of photosyn-thetic rates (data not shown). Each curve consisted of 10steps, starting from ambient CO2 concentration (360) anddecreasing to 200, 100, and 50 µmol CO2 mol−1, beforeincreasing to 360, 500, 720, 1,000, 1,500, and 1,800 µmolCO2 mol−1. Leaf temperature was maintained at 25˚C andleaf to air vapor pressure saturation deficit (VPD) was keptat less than 1.2 kPa. The maximum rate of carboxylation(Vcmax) was estimated using the equation of the Furquhartype model (Farqhar et al. 1980):
Aarea ¼ Vcmax � Cc � Γ*� �.
Cc þ Kc � 1þ Oi=Koð Þf g � Rd
where Cc is the CO2 concentration in the chloroplasts, Γ* isthe CO2 compensation point, Kc and Ko are the Michaelis-Menten constants for carboxylation and oxygenation, Oi isthe intercellular O2 concentration, and Rd is the rate of day-time respiration. In this study, we ignored possible CO2
diffusion limitation within the leaf, and Ci was used as Cc.For the coefficients Kc, Ko, and Γ*, we used 404.9 µmol mol−1,
Interactive effects of abiotic factors on N2 fixation of two alder species 61
278.4 mmol mol−1, and 42.75 µmol mol−1, respectively, at25°C (Bernacchi et al. 2001). For leaves in which full A/Ci
curves were measured, Vcmax was calculated by fitting theequation to the initial slope of the A/Ci curve (Ci<300).
2.7 Statistical analysis
Three-way analysis of variance (ANOVA) for split-plotdesigns was used to evaluate the effects of CO2-treatment,P-treatment, and water-treatment on nitrogenase activity,growth properties (biomass of each tissue, whole-plantbiomass, LA, and LAR), physiological properties (Asat andVcmax), and leaf characters (leaf N, P, N/P ratio) at theprobability level P<0.05 with the JMP software (SASInstitute 2003). The probability level P<0.15 was consideredto indicate a trend. Differences among treatments in rootweight, nodule weight, and LA were analyzed with anANCOVA, including treatments as a factor. These parametersare size dependent, and whole-plant biomass was thereforeincluded as a covariable. These parameters did not alwaysshowed significant correlation with whole-plant biomass,except for root biomass in A. hirsuta. Therefore, the two soilP treatments data were combined to conduct analysis. It wasfirst tested whether there was a significant treatments (CO2
and water treatments) × whole-plant biomass interaction. Ifthis interaction was not significant, then the analysis wasrepeated excluding the interactive effect. If this interactionwas significant, further analysis was conducted for eachwater treatment separately whether there was a significantCO2 treatments × whole-plant biomass interaction.
3 Results
3.1 Nitrogenase activity of nodules
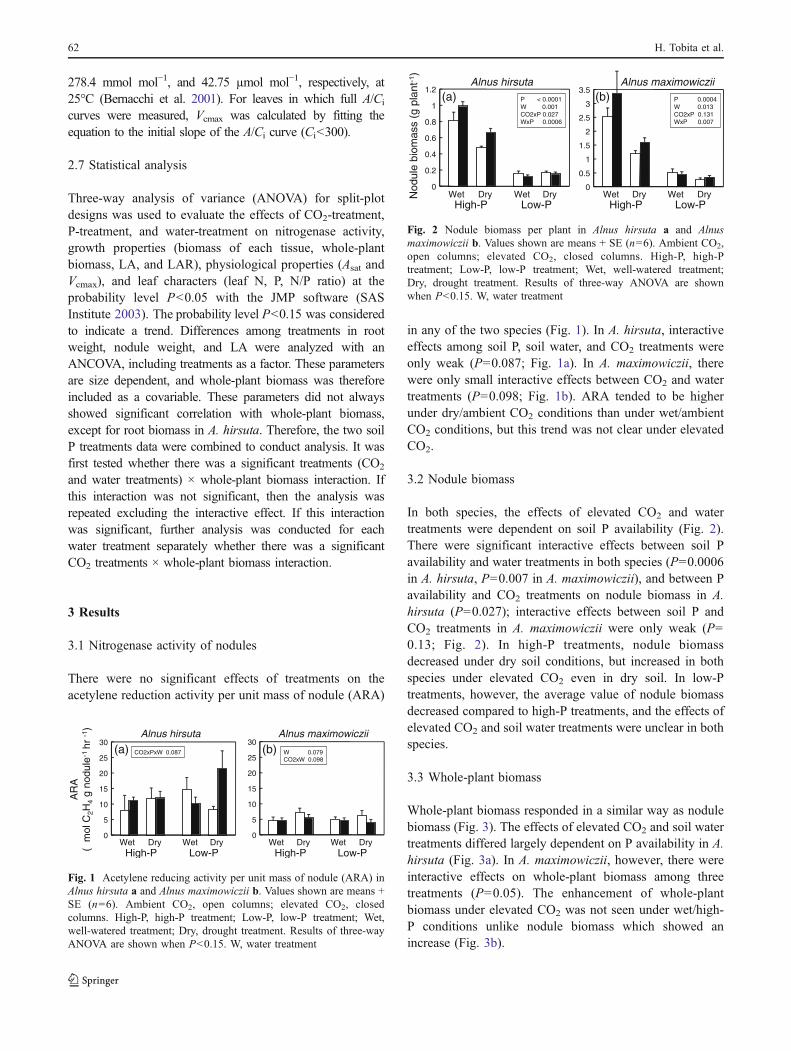
There were no significant effects of treatments on theacetylene reduction activity per unit mass of nodule (ARA)
in any of the two species (Fig. 1). In A. hirsuta, interactiveeffects among soil P, soil water, and CO2 treatments wereonly weak (P=0.087; Fig. 1a). In A. maximowiczii, therewere only small interactive effects between CO2 and watertreatments (P=0.098; Fig. 1b). ARA tended to be higherunder dry/ambient CO2 conditions than under wet/ambientCO2 conditions, but this trend was not clear under elevatedCO2.
3.2 Nodule biomass
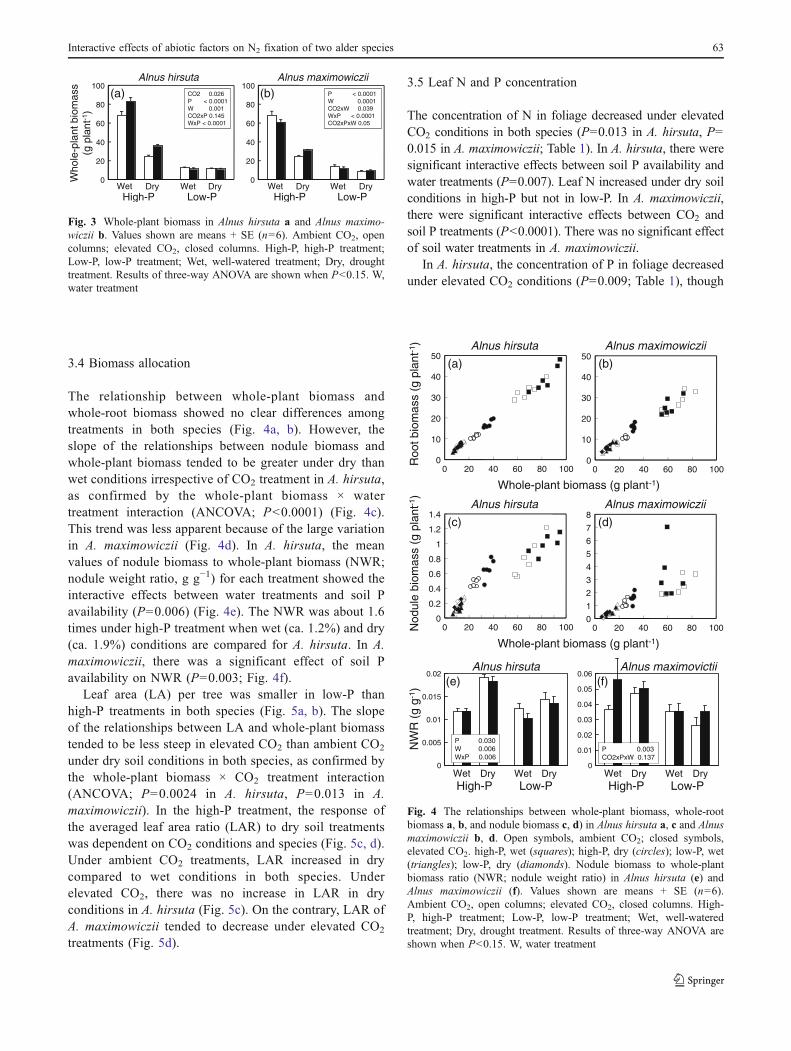
In both species, the effects of elevated CO2 and watertreatments were dependent on soil P availability (Fig. 2).There were significant interactive effects between soil Pavailability and water treatments in both species (P=0.0006in A. hirsuta, P=0.007 in A. maximowiczii), and between Pavailability and CO2 treatments on nodule biomass in A.hirsuta (P=0.027); interactive effects between soil P andCO2 treatments in A. maximowiczii were only weak (P=0.13; Fig. 2). In high-P treatments, nodule biomassdecreased under dry soil conditions, but increased in bothspecies under elevated CO2 even in dry soil. In low-Ptreatments, however, the average value of nodule biomassdecreased compared to high-P treatments, and the effects ofelevated CO2 and soil water treatments were unclear in bothspecies.
3.3 Whole-plant biomass
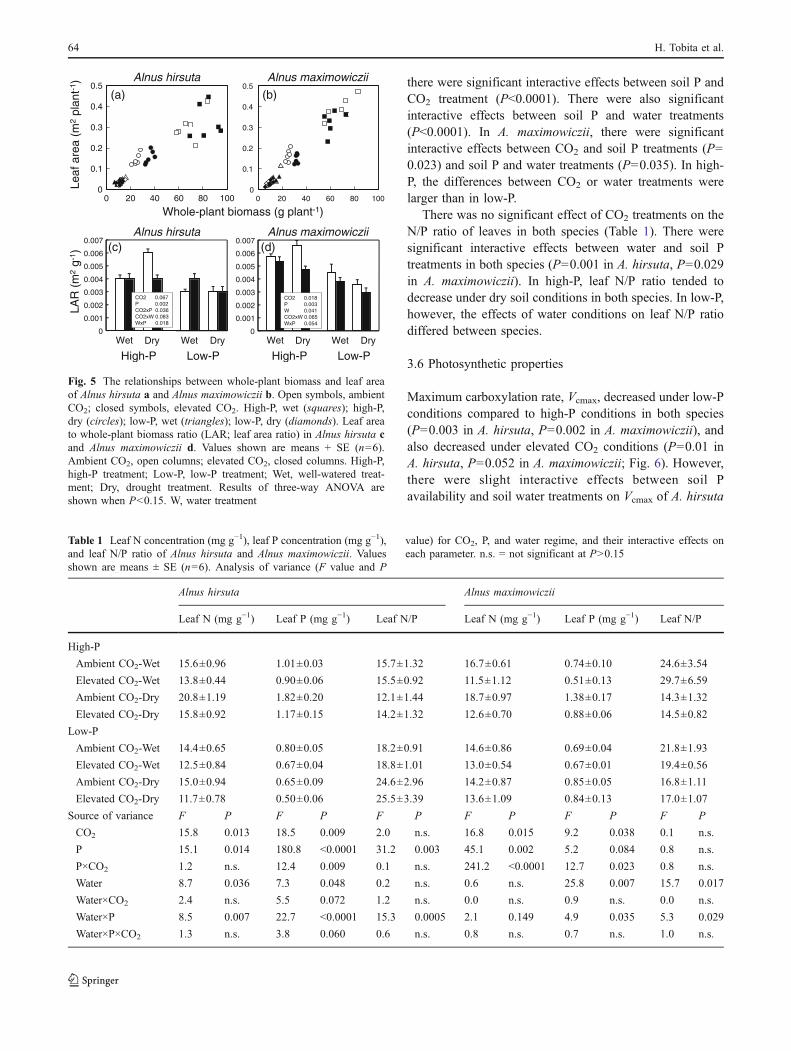
Whole-plant biomass responded in a similar way as nodulebiomass (Fig. 3). The effects of elevated CO2 and soil watertreatments differed largely dependent on P availability in A.hirsuta (Fig. 3a). In A. maximowiczii, however, there wereinteractive effects on whole-plant biomass among threetreatments (P=0.05). The enhancement of whole-plantbiomass under elevated CO2 was not seen under wet/high-P conditions unlike nodule biomass which showed anincrease (Fig. 3b).
Alnus hirsuta Alnus maximowiczii
High-P Low-PWet WetDry Dry
High-P Low-PWet WetDry Dry
AR
A(µ
mol
C2H
4 g
nodu
le-1
hr
-1)
0
5
10
15
20
25
30
0
5
10
15
20
25
30W 0.079CO2xW 0.098
CO2xPxW 0.087(a) (b)
Fig. 1 Acetylene reducing activity per unit mass of nodule (ARA) inAlnus hirsuta a and Alnus maximowiczii b. Values shown are means +SE (n=6). Ambient CO2, open columns; elevated CO2, closedcolumns. High-P, high-P treatment; Low-P, low-P treatment; Wet,well-watered treatment; Dry, drought treatment. Results of three-wayANOVA are shown when P<0.15. W, water treatment
Alnus hirsuta
High-P Low-PWet WetDry Dry
High-P Low-PWet WetDry DryN
odul
e bi
omas
s (g
pla
nt-1
)
0
0.2
0.4
0.6
0.8
1
1.2
0
0.5
1
1.5
2
2.5
3
3.5P < 0.0001W 0.001CO2xP 0.027WxP 0.0006
P 0.0004W 0.013CO2xP 0.131WxP 0.007
(a) (b)Alnus maximowiczii
Fig. 2 Nodule biomass per plant in Alnus hirsuta a and Alnusmaximowiczii b. Values shown are means + SE (n=6). Ambient CO2,open columns; elevated CO2, closed columns. High-P, high-Ptreatment; Low-P, low-P treatment; Wet, well-watered treatment;Dry, drought treatment. Results of three-way ANOVA are shownwhen P<0.15. W, water treatment
62 H. Tobita et al.
3.4 Biomass allocation
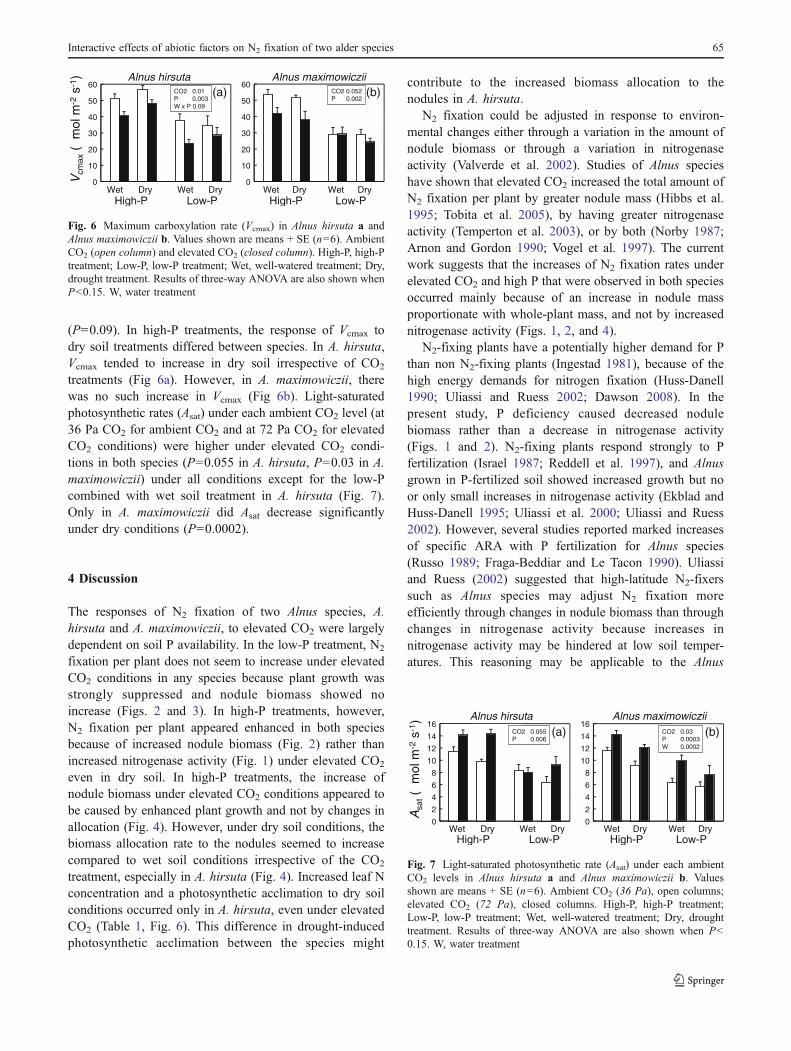
The relationship between whole-plant biomass andwhole-root biomass showed no clear differences amongtreatments in both species (Fig. 4a, b). However, theslope of the relationships between nodule biomass andwhole-plant biomass tended to be greater under dry thanwet conditions irrespective of CO2 treatment in A. hirsuta,as confirmed by the whole-plant biomass × watertreatment interaction (ANCOVA; P<0.0001) (Fig. 4c).This trend was less apparent because of the large variationin A. maximowiczii (Fig. 4d). In A. hirsuta, the meanvalues of nodule biomass to whole-plant biomass (NWR;nodule weight ratio, g g−1) for each treatment showed theinteractive effects between water treatments and soil Pavailability (P=0.006) (Fig. 4e). The NWR was about 1.6times under high-P treatment when wet (ca. 1.2%) and dry(ca. 1.9%) conditions are compared for A. hirsuta. In A.maximowiczii, there was a significant effect of soil Pavailability on NWR (P=0.003; Fig. 4f).
Leaf area (LA) per tree was smaller in low-P thanhigh-P treatments in both species (Fig. 5a, b). The slopeof the relationships between LA and whole-plant biomasstended to be less steep in elevated CO2 than ambient CO2
under dry soil conditions in both species, as confirmed bythe whole-plant biomass × CO2 treatment interaction(ANCOVA; P=0.0024 in A. hirsuta, P=0.013 in A.maximowiczii). In the high-P treatment, the response ofthe averaged leaf area ratio (LAR) to dry soil treatmentswas dependent on CO2 conditions and species (Fig. 5c, d).Under ambient CO2 treatments, LAR increased in drycompared to wet conditions in both species. Underelevated CO2, there was no increase in LAR in dryconditions in A. hirsuta (Fig. 5c). On the contrary, LAR ofA. maximowiczii tended to decrease under elevated CO2
treatments (Fig. 5d).
3.5 Leaf N and P concentration
The concentration of N in foliage decreased under elevatedCO2 conditions in both species (P=0.013 in A. hirsuta, P=0.015 in A. maximowiczii; Table 1). In A. hirsuta, there weresignificant interactive effects between soil P availability andwater treatments (P=0.007). Leaf N increased under dry soilconditions in high-P but not in low-P. In A. maximowiczii,there were significant interactive effects between CO2 andsoil P treatments (P<0.0001). There was no significant effectof soil water treatments in A. maximowiczii.
In A. hirsuta, the concentration of P in foliage decreasedunder elevated CO2 conditions (P=0.009; Table 1), though
Who
le-p
lant
bio
mas
s(g
pla
nt-1
)
High-P Low-PWet WetDry Dry
High-P Low-PWet WetDry Dry
Alnus hirsuta Alnus maximowiczii
0
20
40
60
80
100
0
20
40
60
80
100CO2 0.026 P < 0.0001W 0.001CO2xP 0.145WxP < 0.0001
P < 0.0001W 0.0001CO2xW 0.039WxP < 0.0001CO2xPxW 0.05
(a) (b)
Fig. 3 Whole-plant biomass in Alnus hirsuta a and Alnus maximo-wiczii b. Values shown are means + SE (n=6). Ambient CO2, opencolumns; elevated CO2, closed columns. High-P, high-P treatment;Low-P, low-P treatment; Wet, well-watered treatment; Dry, droughttreatment. Results of three-way ANOVA are shown when P<0.15. W,water treatment
Roo
t bio
mas
s (g
pla
nt-1
)
0
10
20
30
40
50
0 20 40 60 80 1000
10
20
30
40
50
0 20 40 60 80 100
Whole-plant biomass (g plant-1)
Alnus hirsuta Alnus maximowiczii
(a) (b)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
0 20 40 60 80 100Nod
ule
biom
ass
(g p
lant
-1)
Whole-plant biomass (g plant-1)
Alnus hirsuta Alnus maximowiczii
0
1
2
3
4
5
6
7
8
0 20 40 60 80 100
(c) (d)
0
0.01
0.02
0.03
0.04
0.05
0.06
0
0.005
0.01
0.015
0.02Alnus hirsuta Alnus maximovictii
High-P Low-PWet WetDry Dry
High-P Low-PWet WetDry Dry
NW
R (
g g-
1 )
P 0.030W 0.006WxP 0.006
P 0.003CO2xPxW 0.137
(e) (f)
Fig. 4 The relationships between whole-plant biomass, whole-rootbiomass a, b, and nodule biomass c, d) in Alnus hirsuta a, c and Alnusmaximowiczii b, d. Open symbols, ambient CO2; closed symbols,elevated CO2. high-P, wet (squares); high-P, dry (circles); low-P, wet(triangles); low-P, dry (diamonds). Nodule biomass to whole-plantbiomass ratio (NWR; nodule weight ratio) in Alnus hirsuta (e) andAlnus maximowiczii (f). Values shown are means + SE (n=6).Ambient CO2, open columns; elevated CO2, closed columns. High-P, high-P treatment; Low-P, low-P treatment; Wet, well-wateredtreatment; Dry, drought treatment. Results of three-way ANOVA areshown when P<0.15. W, water treatment
Interactive effects of abiotic factors on N2 fixation of two alder species 63
there were significant interactive effects between soil P andCO2 treatment (P<0.0001). There were also significantinteractive effects between soil P and water treatments(P<0.0001). In A. maximowiczii, there were significantinteractive effects between CO2 and soil P treatments (P=0.023) and soil P and water treatments (P=0.035). In high-P, the differences between CO2 or water treatments werelarger than in low-P.
There was no significant effect of CO2 treatments on theN/P ratio of leaves in both species (Table 1). There weresignificant interactive effects between water and soil Ptreatments in both species (P=0.001 in A. hirsuta, P=0.029in A. maximowiczii). In high-P, leaf N/P ratio tended todecrease under dry soil conditions in both species. In low-P,however, the effects of water conditions on leaf N/P ratiodiffered between species.
3.6 Photosynthetic properties
Maximum carboxylation rate, Vcmax, decreased under low-Pconditions compared to high-P conditions in both species(P=0.003 in A. hirsuta, P=0.002 in A. maximowiczii), andalso decreased under elevated CO2 conditions (P=0.01 inA. hirsuta, P=0.052 in A. maximowiczii; Fig. 6). However,there were slight interactive effects between soil Pavailability and soil water treatments on Vcmax of A. hirsuta
Table 1 Leaf N concentration (mg g−1), leaf P concentration (mg g−1), and leaf N/P ratio of Alnus hirsuta and Alnus maximowiczii. Values shownare means ± SE (n=6). Analysis of variance (F value and P value) for CO2, P, and water regime, and their interactive effects on each parameter. n.s. = not significant at P>0.15
Alnus hirsuta Alnus maximowiczii
Leaf N (mg g−1) Leaf P (mg g−1) Leaf N/P Leaf N (mg g−1) Leaf P (mg g−1) Leaf N/P
High-P
Ambient CO2-Wet 15.6±0.96 1.01±0.03 15.7±1.32 16.7±0.61 0.74±0.10 24.6±3.54
Elevated CO2-Wet 13.8±0.44 0.90±0.06 15.5±0.92 11.5±1.12 0.51±0.13 29.7±6.59
Ambient CO2-Dry 20.8±1.19 1.82±0.20 12.1±1.44 18.7±0.97 1.38±0.17 14.3±1.32
Elevated CO2-Dry 15.8±0.92 1.17±0.15 14.2±1.32 12.6±0.70 0.88±0.06 14.5±0.82
Low-P
Ambient CO2-Wet 14.4±0.65 0.80±0.05 18.2±0.91 14.6±0.86 0.69±0.04 21.8±1.93
Elevated CO2-Wet 12.5±0.84 0.67±0.04 18.8±1.01 13.0±0.54 0.67±0.01 19.4±0.56
Ambient CO2-Dry 15.0±0.94 0.65±0.09 24.6±2.96 14.2±0.87 0.85±0.05 16.8±1.11
Elevated CO2-Dry 11.7±0.78 0.50±0.06 25.5±3.39 13.6±1.09 0.84±0.13 17.0±1.07
Source of variance F P F P F P F P F P F P
CO2 15.8 0.013 18.5 0.009 2.0 n.s. 16.8 0.015 9.2 0.038 0.1 n.s.
P 15.1 0.014 180.8 <0.0001 31.2 0.003 45.1 0.002 5.2 0.084 0.8 n.s.
P×CO2 1.2 n.s. 12.4 0.009 0.1 n.s. 241.2 <0.0001 12.7 0.023 0.8 n.s.
Water 8.7 0.036 7.3 0.048 0.2 n.s. 0.6 n.s. 25.8 0.007 15.7 0.017
Water×CO2 2.4 n.s. 5.5 0.072 1.2 n.s. 0.0 n.s. 0.9 n.s. 0.0 n.s.
Water×P 8.5 0.007 22.7 <0.0001 15.3 0.0005 2.1 0.149 4.9 0.035 5.3 0.029
Water×P×CO2 1.3 n.s. 3.8 0.060 0.6 n.s. 0.8 n.s. 0.7 n.s. 1.0 n.s.
Table 1 Leaf N concentration (mg g−1), leaf P concentration (mg g−1),and leaf N/P ratio of Alnus hirsuta and Alnus maximowiczii. Valuesshown are means ± SE (n=6). Analysis of variance (F value and P
value) for CO2, P, and water regime, and their interactive effects oneach parameter. n.s. = not significant at P>0.15
0
0.1
0.2
0.3
0.4
0.5
0 20 40 60 80 1000
0.1
0.2
0.3
0.4
0.5
0 20 40 60 80 100
Leaf
are
a (m
2 pl
ant-1
)
Whole-plant biomass (g plant-1)
Alnus hirsuta Alnus maximowiczii
(a) (b)
LAR
(m
2 g-
1 )
0
0.001
0.002
0.003
0.004
0.005
0.006
0.007
High-P Low-PWet WetDry Dry
Alnus hirsuta Alnus maximowiczii
CO2 0.067P 0.002CO2xP 0.036CO2xW 0.083WxP 0.018
0
0.001
0.002
0.003
0.004
0.005
0.006
0.007
CO2 0.018P 0.003W 0.041CO2xW 0.065WxP 0.054
(c) (d)
High-P Low-PWet WetDry Dry
Fig. 5 The relationships between whole-plant biomass and leaf areaof Alnus hirsuta a and Alnus maximowiczii b. Open symbols, ambientCO2; closed symbols, elevated CO2. High-P, wet (squares); high-P,dry (circles); low-P, wet (triangles); low-P, dry (diamonds). Leaf areato whole-plant biomass ratio (LAR; leaf area ratio) in Alnus hirsuta cand Alnus maximowiczii d. Values shown are means + SE (n=6).Ambient CO2, open columns; elevated CO2, closed columns. High-P,high-P treatment; Low-P, low-P treatment; Wet, well-watered treat-ment; Dry, drought treatment. Results of three-way ANOVA areshown when P<0.15. W, water treatment
64 H. Tobita et al.
(P=0.09). In high-P treatments, the response of Vcmax todry soil treatments differed between species. In A. hirsuta,Vcmax tended to increase in dry soil irrespective of CO2
treatments (Fig 6a). However, in A. maximowiczii, therewas no such increase in Vcmax (Fig 6b). Light-saturatedphotosynthetic rates (Asat) under each ambient CO2 level (at36 Pa CO2 for ambient CO2 and at 72 Pa CO2 for elevatedCO2 conditions) were higher under elevated CO2 condi-tions in both species (P=0.055 in A. hirsuta, P=0.03 in A.maximowiczii) under all conditions except for the low-Pcombined with wet soil treatment in A. hirsuta (Fig. 7).Only in A. maximowiczii did Asat decrease significantlyunder dry conditions (P=0.0002).
4 Discussion
The responses of N2 fixation of two Alnus species, A.hirsuta and A. maximowiczii, to elevated CO2 were largelydependent on soil P availability. In the low-P treatment, N2
fixation per plant does not seem to increase under elevatedCO2 conditions in any species because plant growth wasstrongly suppressed and nodule biomass showed noincrease (Figs. 2 and 3). In high-P treatments, however,N2 fixation per plant appeared enhanced in both speciesbecause of increased nodule biomass (Fig. 2) rather thanincreased nitrogenase activity (Fig. 1) under elevated CO2
even in dry soil. In high-P treatments, the increase ofnodule biomass under elevated CO2 conditions appeared tobe caused by enhanced plant growth and not by changes inallocation (Fig. 4). However, under dry soil conditions, thebiomass allocation rate to the nodules seemed to increasecompared to wet soil conditions irrespective of the CO2
treatment, especially in A. hirsuta (Fig. 4). Increased leaf Nconcentration and a photosynthetic acclimation to dry soilconditions occurred only in A. hirsuta, even under elevatedCO2 (Table 1, Fig. 6). This difference in drought-inducedphotosynthetic acclimation between the species might
contribute to the increased biomass allocation to thenodules in A. hirsuta.
N2 fixation could be adjusted in response to environ-mental changes either through a variation in the amount ofnodule biomass or through a variation in nitrogenaseactivity (Valverde et al. 2002). Studies of Alnus specieshave shown that elevated CO2 increased the total amount ofN2 fixation per plant by greater nodule mass (Hibbs et al.1995; Tobita et al. 2005), by having greater nitrogenaseactivity (Temperton et al. 2003), or by both (Norby 1987;Arnon and Gordon 1990; Vogel et al. 1997). The currentwork suggests that the increases of N2 fixation rates underelevated CO2 and high P that were observed in both speciesoccurred mainly because of an increase in nodule massproportionate with whole-plant mass, and not by increasednitrogenase activity (Figs. 1, 2, and 4).
N2-fixing plants have a potentially higher demand for Pthan non N2-fixing plants (Ingestad 1981), because of thehigh energy demands for nitrogen fixation (Huss-Danell1990; Uliassi and Ruess 2002; Dawson 2008). In thepresent study, P deficiency caused decreased nodulebiomass rather than a decrease in nitrogenase activity(Figs. 1 and 2). N2-fixing plants respond strongly to Pfertilization (Israel 1987; Reddell et al. 1997), and Alnusgrown in P-fertilized soil showed increased growth but noor only small increases in nitrogenase activity (Ekblad andHuss-Danell 1995; Uliassi et al. 2000; Uliassi and Ruess2002). However, several studies reported marked increasesof specific ARA with P fertilization for Alnus species(Russo 1989; Fraga-Beddiar and Le Tacon 1990). Uliassiand Ruess (2002) suggested that high-latitude N2-fixerssuch as Alnus species may adjust N2 fixation moreefficiently through changes in nodule biomass than throughchanges in nitrogenase activity because increases innitrogenase activity may be hindered at low soil temper-atures. This reasoning may be applicable to the Alnus
Asa
t (µm
ol m
-2 s
-1)
0
2
4
6
8
10
12
14
16CO2 0.03P 0.0003W 0.0002
0
2
4
6
8
10
12
14
16CO2 0.055P 0.006
High-P Low-PWet WetDry Dry
High-P Low-PWet WetDry Dry
Alnus hirsuta Alnus maximowiczii
(a) (b)
Fig. 7 Light-saturated photosynthetic rate (Asat) under each ambientCO2 levels in Alnus hirsuta a and Alnus maximowiczii b. Valuesshown are means + SE (n=6). Ambient CO2 (36 Pa), open columns;elevated CO2 (72 Pa), closed columns. High-P, high-P treatment;Low-P, low-P treatment; Wet, well-watered treatment; Dry, droughttreatment. Results of three-way ANOVA are also shown when P<0.15. W, water treatment
Vcm
ax (
µmol
m-2
s-1
)
0
10
20
30
40
50
60
0
10
20
30
40
50
60CO2 0.01P 0.003W x P 0.09
CO2 0.052P 0.002
High-P Low-PWet WetDry Dry
High-P Low-PWet WetDry Dry
Alnus hirsuta Alnus maximowiczii(a) (b)
Fig. 6 Maximum carboxylation rate (Vcmax) in Alnus hirsuta a andAlnus maximowiczii b. Values shown are means + SE (n=6). AmbientCO2 (open column) and elevated CO2 (closed column). High-P, high-Ptreatment; Low-P, low-P treatment; Wet, well-watered treatment; Dry,drought treatment. Results of three-way ANOVA are also shown whenP<0.15. W, water treatment
Interactive effects of abiotic factors on N2 fixation of two alder species 65

seedlings grown at relatively high-latitude in the presentstudy.
Sufficient P is required for nodule initiation (Huss-Danell 1997) and Alnus are dependent on an adequate Psupply to establish the symbiosis with Frankia (Wall et al.2000). In the current study, we could not establish whetherthe decreased nodule biomass in P deficient soil was causedby an inhibition of nodulation or limited plant growth.However, we speculate that nodule growth was limited dueto the suppression of plant growth by low photosyntheticrates and decreased leaf area even under elevated CO2
(Figs. 2, 4, 5, and 6). Under elevated CO2, an additional Psupply seems to be needed to enhance N2 fixation of thetwo Alnus species.
One important way for plants to control actinorhizalsymbioses may be the regulation of the proportion ofsymbiotic tissue in the plant, in relation to plant biomassallocation (Wall and Berry 2008). There are two levels offeedback regulation of nodulation (Wall 2000); autoregula-tion of nodulation during early nodule formation (Wall andHuss-Danell 1997), and fixed-N regulation of the suscep-tibility of roots to infection and nodulation (Parsons et al.1993). The number of nodules in Alnus is determined bythe plant in the autoregulation process that is modulated bynutrients such as N and P (Huss-Danell 1997; Wall et al.2000). In the current experiment, the averaged ratio ofnodule biomass to whole-plant biomass (NWR) tended tobe higher at high-P than at low-P in both species, but theresponse was more pronounced in the drought treatments,especially in A. hirsuta (Fig. 4). As the results, the biomassallocation rate to the nodules seemed to increase under drysoil conditions compared to wet soil conditions irrespectiveof the CO2 treatment, especially in A. hirsuta (Fig. 4).However, we could not verify whether changes in nodulenumber or nodule mass caused the increased rate.
Since nodule formation was detected at the beginning ofthe current experiment, fixed-N regulation could contributeto the regulation of nodulation. Fixed-N regulation ofnodulation depends on the external N/P ratio that is sensedby the plant (Ekblad and Huss-Danell 1995; Wall et al.2000; Gentili and Huss-Danell 2003; Wall and Berry 2008).In the current study, only one tenth of the standard Nconcentration of the nutrient solution was applied to theseedlings to avoid inhibition of nodule formation by N(Tobita et al. 2005). The N/P ratio in the fertilizer was 1.5under high-P and 15 under low-P treatment. The low N/Pratio under well-watered and high-P conditions suggeststhat there was no regulation of nodulation by an excessavailability of N.
Foliar N/P ratio can be used as an indicator of nutrientlimitation at community level and ecosystem processes(Koerselman and Meuleman 1996). Multiple publishedstudies propose that leaf N/P ratios <14 should indicate N
limitation, while N/P ratios >16 likely reflect P limitation(Koerselman and Meuleman 1996; Townsend et al. 2007).In an actinorhizal plants, Discaria trinervis, the proportionof nodule biomass was negatively correlated with leaf N/Pratio, and showed lower values at leaf N/P>16 (Valverde etal. 2002). In the current study, there was no significantrelationship between leaf N/P ratio and NWR, except forunder high-P treatments in A. hirsuta (Table 1, Fig. 4).Though the nutrient balance indicated by leaf N/P ratiosmight also contribute to the increased biomass allocation tothe nodules under high-P – dry condition in A. hirsuta(Fig. 4), the leaf N/P ratios could not explain the responsesof NWR for the other treatments.
The leaf N/P ratios exhibited no significant change underelevated CO2 in both species, though soil water conditionsand soil P availability affected the values (Table 1). Thisresult suggests that nutrient balance of leaves might not beaffected by elevated CO2. On the contrary, the plant N/Pratios in herbaceous N2-fixers, Trifolium repens, wereaffected by elevated CO2 (Edwards et al. 2006). Foliar N/P ratios are not simply a function of soil availability. It maybe necessary to consider that various factors can regulatefoliar chemistry (Wright et al. 2001).
High rates of nodule growth and a maintenance of highrates of nitrogen fixation were observed in Alnus species inearly autumn (Kaelke and Dawson 2003). In the currentstudy, nodule biomass was evaluated once in the secondsummer of the experimental period. At that stage, the plantshad already experienced autumn once. Therefore, althoughthe absolute nodule biomass may be different betweensummer and autumn, the differences of nodule biomassbetween treatments could be considered to representspecific responses to each treatment.
Assimilated carbon is required for the formation, growth,and maintenance of nodule tissues and Frankia cells, forthe process of N2 fixation, and for N-assimilation andtransfer (Huss-Danell 1997). Actinorhizal plants on N-limited soils can maintain high net CO2 assimilation withminimal negative adjustment of photosynthetic capacity,Vcmax, following exposure to elevated CO2 (Dawson 2008).While Vcmax tended to decrease under elevated CO2 in bothspecies (Fig. 6), photosynthetic rate was higher when CO2
was elevated (Fig. 7). This increase in photosynthetic ratesunder elevated CO2 will compensate for the decline inLAR, especially under dry soil conditions at high-P(Fig. 5), and might indicate an increase of photosyntheticcapacity per plant under elevated CO2 conditions. Alnusglutinosa grown at doubled ambient CO2 showed a 16%greater rate of CO2 fixation, and a 46% increase innitrogenase activity of detached nodules relative to controlskept at ambient levels of CO2 (Vogel and Curtis 1995).
Only under high-P conditions, LAR increased in bothspecies in dry soil compared to watered soil under ambient
66 H. Tobita et al.

CO2; photosynthetic acclimation to drought, as indicated bythe increase in Vcmax, occurred in A. hirsuta irrespective ofthe CO2 treatment (Figs. 5 and 6). Similar photosyntheticacclimation to dry soil was reported for Betula platyphylla(Kitao et al. 2007) and other species (Qaderi et al. 2006)under elevated CO2. These photosynthetic responses todrought probably alleviate the negative adjustment ofphotosynthetic capacity to elevated CO2. More photo-synthates than required to support plant growth may beproduced due to increased photosynthetic capacity on drysoil, and the allocation of assimilates to nodules mightincrease. In this way, the photosynthetic acclimation todrought might contribute to an increased biomass allocationto nodules in A. hirsuta.
The leaves of Alnus maximowiczii contain largeramounts of defense chemicals such as condensed tannin,than those of A. hirsuta (Agari et al. 2007). Because thesynthesis of these chemicals is ultimately based on photo-synthates, we speculate that the amount of carbohydratesthat may be allocated to nodules is smaller in A.maximowiczii than in A. hirsuta. As a result, photosyntheticacclimation and the increase of biomass allocation tonodules under dry soil conditions might be not aspronounced in A. maximowiczii as it is in A. hirsuta (Figs. 4and 6). This hypothesis will be tested in future studies.
We measured nitrogenase activity after watering the rootsystems. Thus, what we determined actually was thepotential nitrogenase activity of the nodules. Nitrogenaseactivity is sensitive to drought in the short term (Seiler andJohnson 1984; Huss-Danell 1997). Alnus incana ssp.incana showed declining nitrogenase activity with decreas-ing water potential from -0.6 to -1.4 MPa, which was thelevel at which stomatal closure occurred and plants startedto wilt (Sundström and Huss-Danell 1987). In the currentstudy, the leaf water potential at predawn decreased downto about -1.2 MPa during the driest periods of the weeklywatering cycle in the drought treatment (data not shown). Inthese phases, nitrogenase activity may decline to someextent. The leaf water potential at predawn, however,recovered to about -0.1 MPa after scheduled irrigation,implying that nitrogenase activity will not always be lowerthan the potential activity we have determined (Fig. 1).
Yamanaka et al. (2003) reported that the presence ofmycorrhizae on nodulated alders in a tripartite symbiosisimproved phosphate uptake by Alnus tenuifolia, enhancinggrowth and nitrogen fixation of the host plant. In thecurrent study, we did not focus on the possible effects ofmycorrhizae, but we observed ectomycorrhizal roots on allplants (data not shown). We have to await future research toevaluate the effects of elevated CO2 on such tripartiteassociations.
In conclusion, soil P supply proved more important toN2 fixation than the soil moisture level in this experiment.
In P deficient soil, elevated CO2 could not increase N2
fixation per plant because of the suppression of growth. Inhigh-P conditions, elevated CO2 increased N2 fixation dueto increased nodule biomass even under drought.
Acknowledgement This study was supported in part by the Programfor Promotion of Basic and Applied Researches for Innovations inBio-oriented Industry (BRAIN).
References
Agari T, Matsuki S, Tobita H, Sasa K, Koike T (2007) The effects ofelevated CO2 and soil fertility on the defense capacity againstherbivore in two species of alder seedlings. Transactions of theMeeting in Hokkaido Branch of the Japanese Forest Society55:56–58 (in Japanese)
Ainsworth EA, Rogers A (2007) The response of photosynthesis andstomatal conductance to rising [CO2]: mechanisms and environ-mental interactions. Plant Cell and Environ 30:258–270
Arnon JA III, Gordon JC (1990) Effect of nodulation, nitrogenfixation and CO2 enrichment on the physiology, growth and drymass allocation of seedlings of Alnus rubra Bong. New Phytol116:55–66
Asher CJ, Edwards DG (1983) Modern solution culture techniques.In: Läuchli A, Bieleski RL (eds) Encyclopedia of PlantPhysiology, NS. vol 15A, Inorganic Plant Nutrition. Springer-Verlag, Berlin, pp 94–119
Bernacchi CJ, Singsaas EL, Pimentel C, Portis AR Jr, Long SP (2001)Improved temperature response functions for models of Rubisco-limited photosynthesis. Plant Cell and Environ 24:253–259
Dawson JO (1983) Dinitrogen fixation in forest ecosystems. Can JMicrobiol 29:979–992
Dawson JO (2008) Ecology of actinorhizal plants. In: Pawlowski K,Newton WE (eds) Nitrogen-fixing Actinorhizal Symbioses.Springer, Dordrecht, pp 119–234
Edwards EJ, McCaffery S, Evans JR (2006) Phosphorus availabilityand elevated CO2 affect biological nitrogen fixation and nutrientfluxes in a clover-dominated sward. New Phytol 169:157–167
Ekblad A, Huss-Danell K (1995) Nitrogen fixation by Alnus incana andnitrogen transfer from A. incana to Pinus sylvestris influenced bymacronutrients and ectomycorrhiza. New Phytol 131:453–459
Farqhar GD, von Caemmerer S, Berry JA (1980) A biochemicalmodel of photosynthetic acclimation in leaves of C3 species.Planta 149:78–90
Flexas J, Medrano H (2002) Drought-inhibition of photosynthesis inC3 plants: stomatal ad non-stomatal limitations revisited. AnnBot 89:183–189
Fraga-Beddiar A, Le Tacon F (1990) Interactions between a VAmycorrhizal fungus and Frankia associated with alder (Alnusglutinosa (L.) Gaerten.). Symbiosis 9:247–258
Gentili F, Huss-Danell K (2003) Local and systemic effects ofphosphorus and nitrogen on nodulation and nodule function inAlnus incana. J Exp Bot 54:2757–2767
Geßler A, Keitel C, Kreuzwieser J, Matyssek R, Seiler W, RennenbergH (2007) Potential risks for European beech (Fagus sylvatica L.)in a changing climate. Trees 21:1–11
Gress SE, Nichols TD, Northcraft CC, Peterjohn WT (2007) Nutrientlimitation in soil exhibiting differing nitrogen availabilities: whatlies beyond nitrogen saturation? Ecology 88(1):119–130
Hardy RWF, Burns RC, Holsten RD (1973) Application of theacetylene-ethylene assay for the measurement of nitrogenfixation. Soil Biol Biochem 5:47–81
Interactive effects of abiotic factors on N2 fixation of two alder species 67

Hibbs DE, Chan SS, Castellano M, Niu CH (1995) Response of redalder seedlings to CO2 enrichment and water stress. New Phytol129:569–577
Houghton JT, Ding Y, Griggs DJ, Noguer M, van des Linden PJ, DaiX, Maskell K, Johnson CA (2001) Climate change 2001: thescientific basis. Intergovernmental panel on climate change(IPCC). Cambridge University, Cambridge, p 881
Hungate BA, Dukes JT, Shaw MR, Luo Y, Field CB (2003) Nitrogenand climate change. Science 302:1512–1513
Hungate BA, Johnson DW, Dijkstra P, Hymus G, Stiling P, MegonigalJP, Pagel AL, Moan JA, Day F, Li J, Hinkle CR, Drake BG(2006) Nitrogen cycle during 7 years of atmospheric CO2
enrichment in a scrub oak woodland. Ecology 87(1):26–40Huss-Danell K (1978) Nitrogenase activity measurements in intact
plants of Alnus incana. Physiol Plant 43:372–376Huss-Danell K (1990) The physiology of actinorhizal nodules. In:
Schwintzer CR, Tjepkema JD (eds) The biology of Frankia andactinorhizal plants. Academic, Tokyo, pp 129–156
Huss-Danell K (1997) Actinorhizal symbioses and their N2 fixation.New Phytol 136:375–405
Hyvönen R, Ågren GI, Linder S, Persson T, Cotrufo F, Ekblad A,Freeman M, Grelle A, Janssens IA, Jarvis PG, Kellomäki S,Lindroth A, Loustau D, Lundmark T, Norby RJ, Oren R,Pilegaard K, Ryan MG, Sigurdsson BD, Stromgren M, van OijenM, Wallin G (2007) The likely impact of elevated [CO2],nitrogen deposition, increased temperature and management oncarbon sequestration in temperate and boreal forest ecosystems: aliterature review. New Phytol 173:463–480
Ingestad T (1981) Nutrition and growth of birch and grey alderseedlings in low conductivity solutions and at varied relativerates of nutrient addition. Physiol Plant 52:454–466
Israel DW (1987) Investigation of the role of phosphorus in symbioticnitrogen fixation. Plant Physiol 84:835–840
Johnson DW (2006) Progressive N limitation in forest: review andimplications for long-term responses to elevated CO2. Ecology87(1):64–75
Kaelke CM, Dawson JO (2003) Seasonal flooding regimes influencesurvival, nitrogen fixation, and the partitioning of nitrogen andbiomass in Alnus incana ssp. rugosa. Plant Soil 254:167–177
Kikuzawa K (1983) Leaf survival of woody plants in deciduousbroad-leaved forests. 1. Tall trees. Can J Bot 61:2133–2139
Kitao M, Lei TT, Koike T, Tobita H, Maruyama Y (2003) Higherelectron transport rate observed at low intercellular CO2
concentration in long-term drought-acclimated leaves of Japanesemountain birch (Betula ermanii Cham.). Physiol Plant 118:406–413
Kitao M, Lei TT, Koike T, Kayama M, Tobita H, Maruyama Y (2007)Interaction of drought and elevated CO2 on photosynthetic down-regulation and susceptibility to photoinhibition in Japanese whitebirch (Betula platyphylla var. japonica) seedlings grown underlimited N availability. Tree Physiol 27(5):727–735
Koerselman W, Meuleman AFM (1996) The vegetation N:P ratio: anew tool to detect the nature of nutrient limitation. J Appl Ecol33(6):1441–1450
Kogawara S, Norisada M, Tange T, Yagi H, Kojima K (2006)Elevated atmospheric CO2 concentration alters the effects ofphosphate supply on growth of Japanese red pine (Pinusdensiflora) seedlings. Tree Physiol 26:25–33
Körner C (2006) Plant CO2 responses: an issue of definition, time andresource supply. New Phytol 172:393–411
Long SP, Ainsworth EA, Rogers A, Ort DR (2004) Ridingatmospheric carbon dioxide: plants FACE the future. Annu RevPlant Physiol 55:591–628
Luo Y, Su B, Currie WS, Dukes JS, Finzi A, Hartwig U, Hungate B,McMurtrie RE, Oren R, Parton WJ, Pataki DE, Shaw MR, ZakDR, Field CB (2004) Progressive nitrogen limitation of ecosys-
tem responses to rising atmospheric carbon dioxide. Bioscience54(8):731–739
Murphy J, Riley JP (1962) A modified single solution method for thedetermination of phosphate in natural waters. Anal Chim Acta27:31–36
Naumburg E, Loik ME, Smith SD (2004) Photosynthetic responses ofLarrea tridentata to seasonal temperature extremes underelevated CO2. New Phytol 162:323–330
Norby RJ (1987) Nodulation and nitrogenase activity in nitrogen-fixing woody plants stimulated by CO2 enrichment of theatmosphere. Physiol Plant 71:77–82
Norby RJ, Iversen CM (2006) Nitrogen uptake, distribution, turnover,and efficiency of use in a CO2-enriched sweetgum forest.Ecology 87(1):5–14
Norby RJ, Wullschleger SD, Gjunderson CA, Johnson DW,Ceulemans R (1999) Tree responses to rising CO2 in fieldexperiments: implications for the future forest. Plant CellEnviron 22:683–714
Nowak RS, Ellsworth DS, Smith SD (2004) Functional responses ofplants to elevated atmospheric CO2-do photosynthetic andproductivity data from FACE experiments support early predic-tions? New Phytol 162:253–280
Parsons R, Stanforth A, Raven JA, Sprent JI (1993) Nodule growthand activity may be regulated by a feedback mechanisminvolving phloem nitrogen. Plant Cell Environ 16:125–136
Qaderi MM, Kurepin LV, Reid DM (2006) Growth and physiologicalresponses of canola (Brassica napus) to three components ofglobal climate chanre: temperature, carbon dioxide and drought.Physiol Plant 128:710–721
Reddell P, Yun Y, Shipron WA (1997) Do Casuarina cunninghamianaseedlings dependent on symbiotic N2 fixation have higherphosphorus requirements than those supplied with adequatefertilizer nitrogen? Plant Soil 189:213–219
Reich PB, Hobbie SE, Lee T, Ellsworth DS, West JB, Tilman D,Knops JMH, Naeem S, Trost J (2006) Nitrogen limitationconstrains sustainability of ecosystem response to CO2. Nature440:922–925
Russo RO (1989) Evaluating alder-endophyte (Alnus acuminate-Frankia-Mycorrhizae) interactions. Plant Soil 118:151–155
SAS Institute (2003) JMP: statistics and graphics guide, version 5.1.SAS, Cary, p 792
Schoenau JJ, O’Halloran IP (2008) Sodium bicarbonate-extractablephosphate. In: Carter MR, Gregorich EG (eds) Soil sampling andmethods of analysis. 2nd edn. CRC, pp 89–94
Schwintzer CR, Tjepkema JD (1994) Factors affecting the acetylene to15 N2 conversion ratio in root nodules of Myrica gale L. PlantPhysiol 106:1041–1047
Seiler JR, Johnson JD (1984) Growth and acetylene reduction of blackalder seedlings in response to water stress. Can J For Res14:477–480
Somego M (1985) Cytogenetic studies in the subgenus Alnuster ofthe genus Alnus. Japan Forest Tree Breeding Association,Tokyo, p 132
Sundström K-R, Huss-Danell K (1987) Effects of water stress onnitrogenase activity in Alnus incana. Physiol Plant 70:342–348
Temperton VM, Grayston SJ, Jackson G, Millard P, Jarvis PG (2003)Effects of elevated carbon dioxide concentration on growth andnitrogen fixation in Alnus glutinosa in a long-term fieldexperiment. Tree Physiol 23:1051–1059
Tobita H, Kitao M, Koike T, Maruyama Y (2005) Effects of elevatedCO2 and Nitrogen availability on nodulation of Alnus hirsutaTurcz. Phyton 45:125–131
Townsend AR, Cleveland CC, Asner GP, Bustamante MMC (2007)Controls over foliar N: P ratios in tropical rain forests. Ecology88(1):107–118
68 H. Tobita et al.

Uliassi DD, Ruess RW (2002) Limitation to symbiotic nitrogenfixation in primary succession on the Tanana river floodplain.Ecology 83:88–103
Uliassi DD, Huss-Danell K, Ruess RW, Doran K (2000) Biomassallocation and nitrogenase activity in Alnus tenuifolia: responsesto successional soil type and phosphorus availability. Ecoscience7:73–79
Valverde C, Ferrari A,Wall LG (2002) Phosphorous and the regulation ofnodulation in the actinorhizal symbiosis between Discaria trinervis(Rhamnaceae) and Frankia BCU110501. New Phytol 153:43–52
Vitousek PM, Howarth RW (1991) Nitrogen limitation on land and inthe sea: how can it occur? Biogeochemistry 13:87–115
Vitousek PM, Aber JD, Howarth RW, Likens GE, Matson PA,Schindler DW, Schlesinger WM, Tilman DG (1997) Humanalteration of the global nitrogen cycle: sources and consequences.Ecol Appl 7(3):737–750
Vogel CS, Curtis PS (1995) Leaf gas and nitrogen dynamics of N2-fixing field-grown Alnus glutinosa under elevated atmosphericCO2. Glob Chang Biol 1:55–61
Vogel CS, Curtis PS, Thomas RB (1997) Growth and nitrogenaccretion of dinitrogen-fixing Alnus glutinosa (L.) Gaertn. underelevated carbon dioxide. Plant Ecol 130:63–70
Wall LG (2000) The actinorhizal symbiosis. J Plant Growth Regul19:167–182
Wall LG, Berry AM (2008) Early interactions, infection andnodulation in actinorhizal symbiosis. In: Pawlowski K, Newton
WE (eds) Nitrogen-fixing Actinorhizal Symbioses. Springer,Dordrecht, pp 147–166
Wall LG, Huss-Danell K (1997) Regulation of nodulation in AlnusFrankia symbiosis. Physiological Plantarum 99:594–600
Wall LG, Hellsten A, Huss-Danell K (2000) Nitrogen, phosphorous,and the ratio between them affect nodulation in Alnus incana andTrifolium pratense. Symbiosis 29:91–105
Wallace ZP, Lovett GM, Hart JE, Machona B (2007) Effects ofnitrogen saturation on the tree growth and death in a mixed-oakforest. For Ecol Manag 243:210–218
Watanabe Y, Tobita H, Kitao M, Maruyama Y, Choi D, Sasa K,Funada R, Koike T (2008) Effects of elevated CO2 and nitrogenon wood structure related to water transport in seedlings of twodeciduous broad-leaved tree species. Trees 22(4):403–411
Wheeler CT, Cameron EM, Gordon JC (1978) Effects of handling andsurgical treatments on nitrogenase activity in root nodules ofAlnus glutinosa, with special reference to the application ofindole-acetic acid. New Phytol 80:175–178
Wright IJ, Reich PB, Westoby M (2001) Strategy shifts in leafphysiology, structure and nutrient content between species ofhigh- and low-rainfall and high- and low-nutrient habitats. FunctEcol 15(4):423–434
Yamanaka T, Li C-Y, Bormann BT, Okabe H (2003) Tripartiteassociations in alder: Effects of Frankia and Alpova diplophloeuson the growth, nitrogen fixation and mineral acquisition of Alnustenuifolia. Plant Soil 254:179–186
Interactive effects of abiotic factors on N2 fixation of two alder species 69





























