Embed Size (px)
Citation preview

1
Daisuke Igarashi
Institute of Life Sciences, Ajinomoto Co., Inc., Kawasaki 210-8681, Japan
Tel. +81-44-244-5643
Fax +81-44-244-9617
E-mail [email protected]
Area Bioenergetics and Photosynthesis
Plant Physiology Preview. Published on September 1, 2006, as DOI:10.1104/pp.106.085514
Copyright 2006 by the American Society of Plant Biologists
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

2
Glutamate:glyoxylate aminotransferase (GGAT) modulates amino acid contents
during photorespiration
Running head: Photorespiratory GGAT modulates amino acid contents
Daisuke Igarashi*, Hiroko Tsuchida, Mitsue Miyao, Chieko Ohsumi
Institute of Life Sciences, Ajinomoto Co., Inc., Kawasaki 210-8681, Japan (D.I., C.O.)
National Institute of Agrobiological Sciences, Tsukuba 305-8602, Japan (H.T., M.M.)
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

3
Financial source
Part of this work was performed in the project “Development of Fundamental
Technologies for Controlling the Production of Industrial Materials by Plants,” which
was supported by the New Energy and Industrial Technology Development Organization
(NEDO), Japan.
*Corresponding author:
E-mail [email protected]
Tel. +81-44-244-5643
Fax +81-44-244-9617
Institute of Life Sciences, Ajinomoto Co., Inc., 1-1 Suzuki-cho Kawasaki-shi 210-8681,
Japan
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

4
ABSTRACT
In photorespiration, peroxisomal glutamate:glyoxylate aminotransferase (GGAT)
catalyzes the reaction of glutamate and glyoxylate to produce 2-oxoglutarate and glycine.
Previous studies demonstrated that alanine aminotransferase-like protein functions as a
photorespiratory GGAT. Photorespiratory transamination to glyoxylate, which is
mediated by GGAT and serine glyoxylate aminotransferase (SGAT), is believed to play
an important role in the biosynthesis and metabolism of major amino acids. To better
understand its role in the regulation of amino acid levels, we produced 42 GGAT1
overexpression lines that express different levels of GGAT1 mRNA. The levels of free
serine, glycine, and citrulline increased markedly in GGAT1 overexpression lines
compared with levels in the wild-type, and levels of these amino acids were strongly
correlated with the levels of GGAT1 mRNA and with the GGAT activity in the leaves.
This accumulation began soon after exposure to light and was repressed under high levels
of CO2. Light and nutrient conditions both affected the amino acid profiles;
supplementation with NH4NO3 increased the levels of some amino acids compared with
the controls. The results suggest that the photorespiratory aminotransferase reactions
catalyzed by GGAT and SGAT are both important regulators of amino acid contents.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

5
INTRODUCTION
Photorespiration is a metabolic pathway in which glycolate-2-phosphate is produced by
the oxygenase activity of ribulose-1,5-bisphosphate carboxylase/oxygenase (rubisco;
Leegood et al., 1995). At current atmospheric concentrations of O2 and CO2,
photorespiration in C3 plants dissipates more than 25% of the carbon fixed by means of
photosynthesis (Sharkey, 1988). Thus, photorespiration is often considered to be a
wasteful process. However, photorespiration may be a mechanism for mitigation of the
photoinhibition that can occur under high light levels, drought, and salt stress (Kozaki
and Takeba 1996; Hoshida et al., 2000; Wingler et al., 2000).
The photorespiratory nitrogen cycle contributes to the metabolism of certain amino
acids (Gln, Glu, Ser, and Gly). Nitrogen is incorporated into the photorespiratory C2
cycle through transamination of glyoxylate to glycine. Peroxisomal glyoxylate
aminotransferases play a central role within the photorespiratory pathway. The reactions
in this pathway are catalyzed by GGAT and SGAT. In Arabidopsis, it was reported that
alanine 2-oxoglutarate aminotransferase (AOAT)-like protein (Igarashi et al., 2003;
Liepman and Olsen, 2003) and alanine glyoxylate aminotransferase (AGT)-like protein
(Liepman and Olsen, 2001) function as photorespiratory GGAT and SGAT, respectively.
In the conversion of glycine to serine, ammonium and CO2 are released. Glutamine
synthetase 2 (GS-2) and ferredoxin-dependent (Fd-) GOGAT effectively refix the
released ammonium in plastids. It has been estimated that the production of ammonium
by photorespiration is an order of magnitude greater than the primary assimilation of
nitrogen resulting from nitrate reduction (Keys et al., 1978). That is, photorespiration
appears to be an essential process in the control of amino acid biosynthesis and
metabolism.
Serine and glycine are involved in protein biosynthesis and serve as precursors in a
variety of important biosynthetic pathways, including phospholipid synthesis (serine) and
purine formation (glycine). Serine and glycine are synthesized through two main
pathways. The first pathway involves transamination of glyoxylate into glycine, which is
in turn converted into serine. A source of glyoxylate is provided by oxidation of glycolate
during the photorespiratory cycle or the conversion of isocitrate to succinate during the
glyoxylate cycle involved in the degradation of fatty acids. The second pathway is
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

6
connected with glycolysis and leads to serine formation from 3-phosphoglycerate.
Glycine is obtained from serine through a serine hydroxymethyltransferase (SHMT)
catalyzed reaction (Bourguignon et al., 1999). In photosynthetic tissues, photorespiration
is assumed to be a major pathway for serine and glycine synthesis because of the high
flux of carbon through this cycle (Gerbaud and Andre, 1979; Tolbert 1980; Somerville
and Somerville, 1983). In nonphotosynthetic tissues (e.g., seeds or roots), serine and
glycine are synthesized mainly though glycolysis and the glyoxylate cycle. However,
detailed regulation mechanisms of biosynthesis of these amino acids are unclear.
In the present study, we investigated photorespiratory transamination using GGAT1
overexpression and knockout lines, and estimated the function of GGAT in the regulation
of amino acid metabolism linked to photorespiration.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

7
RESULTS
Expression analysis
Genomic sequence information indicated that there are four alanine aminotransferase-
like genes in Arabidopsis (Igarashi et al., 2003). Two of them (GGAT1; At1g23310 and
GGAT2; At1g70580) that encode proteins that contain peroxisome targeting signal 1
(PTS1) were identified as encoding photorespiratory GGAT proteins (Igarashi et al.,
2003; Liepman and Olsen, 2003). To better characterize the function of these genes, we
examined the tissue specific expression patterns of two GGAT genes (GGAT1 and
GGAT2) using promoter-GUS system. We also examined the diurnal changes in mRNA
levels.
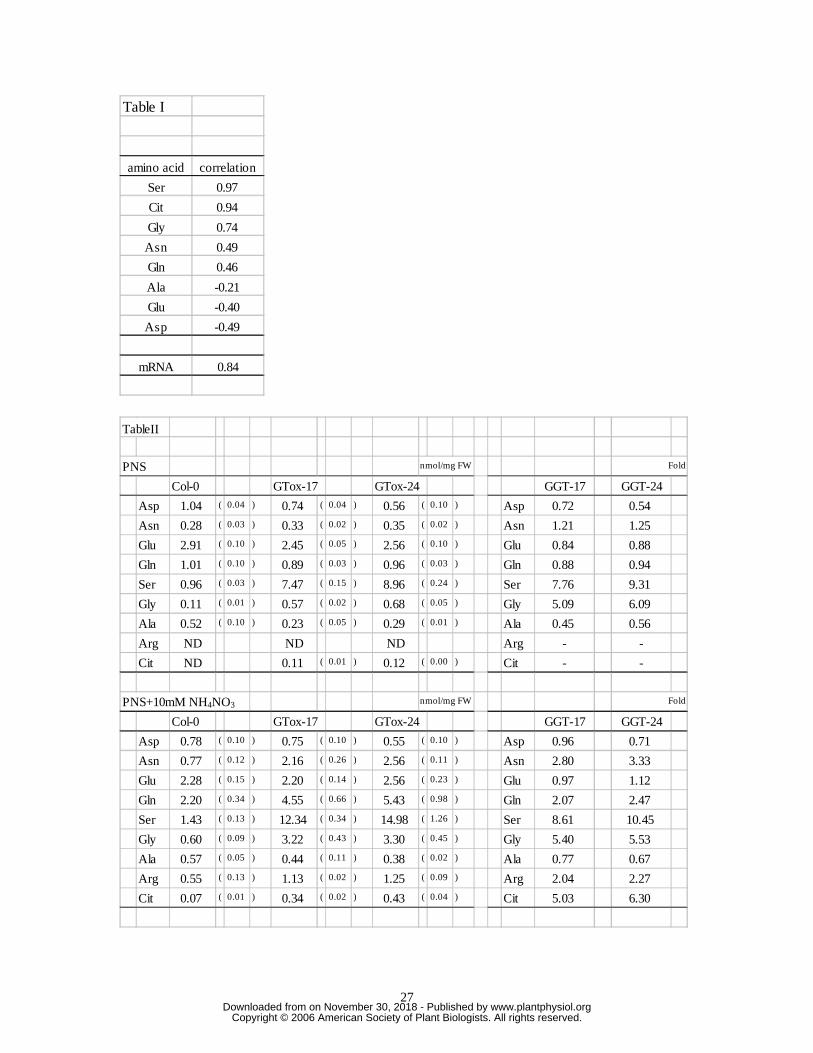
The GUS stain was strongly detected in leaves both of GGAT1- and GGAT2–GUS
seedlings (Fig. 1A), which suggests that GGAT1 and GGAT2 function as
photorespiratory GGAT enzymes. In GGAT1–GUS seedlings, GUS signals were also
strongly detected in roots (Fig. 1A), suggesting that the role of GGAT1 is not restricted to
photorespiration. The function of root peroxisomes has not been identified, but in
nonphotorespiratory tissue (roots) and in darkness, GGAT1 may catalyze the
aminotransferase reaction. At night, serine contents increased in a GGAT1-knockout line
(Igarashi et al., 2003), supporting this hypothesis.
After light exposure, the levels of GGAT1 mRNA decreased and those of GGAT2
mRNA increased rapidly (Fig. 1B). We previously reported that GGAT1 knockout line
(ggat1-1) cannot grow normally in air and recovered under high CO2 conditions, and
concluded that GGAT1 is a major (Igarashi et al., 2003). However the enzymatic activity
of GGAT remained constant during the day (Fig. 2A), suggesting that GGAT1 and
GGAT2 may function redundantly in leaf photorespiration.
Generation of GGAT1 overexpression lines
Detailed analysis of GGAT1 overexpression and knockout plants is necessary for
characterization of the regulatory system for amino acid metabolism that depends on
photorespiration. We previously isolated a GGAT1 knockout line (ggat1-1) from tag-
insertion lines (Igarashi et al., 2003). In the present study, we generated 42 GGAT1
overexpression lines by introducing the GGAT1 genomic region. To characterize the
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

8
relationship between GGAT1 activity and amino acid metabolism, it is essential to isolate
many GGAT1 overexpression lines that have different GGAT activities. We selected the
native GGAT1 promoter rather than a constitutive promoter to control GGAT1
expression. Furthermore, by transferring the 5'-region of GGAT1 to the left border side
of T-DNA, we successfully isolated many transgenic lines that expressed various levels
of GGAT1 mRNA. Levels of GGAT1 mRNA and GGAT activities in the GTox-17
GGAT1 overexpression line were higher than those in the wild-type (Fig. 2A,B). The
diurnal changes in mRNA levels (Fig. 1) remained apparent (Fig. 2B). Diurnal GGAT
activities remained stable in both the wild-type and the GTox-17 line (Fig. 2A).
Correlation of amino acid contents, GGAT activities, and GGAT1 mRNA levels
We selected nine transgenic lines that expressed a wide range of GGAT1 mRNA levels
and determined their amino acid contents, GGAT activities, and GGAT1 mRNA levels.
As shown in Figure 3, there was a strong correlation between GGAT activity and the
level of GGAT1 mRNA (R=0.84). In addition, the serine (R=0.97), glycine (R=0.74), and
citrulline (R=0.94) contents were also strongly correlated with GGAT activity. Table I
presents the correlation values between GGAT1 activity and amino acid contents. Other
than serine, glycine, and citrulline, no significant correlations were observed. The results
suggest that GGAT activity is the limiting factor for synthesis of serine, glycine and
citrulline
Amino acid accumulation
All 42 GGAT1 overexpression lines accumulated serine, glycine, and citrulline (Fig. 4).
Most of the increase in total amino acid level compared with the wild-type was accounted
for by increased serine in the GGAT1 overexpression lines and by increased glutamate in
the knockout line (ggat1-1) because the ratios of serine and glutamate to total amino acid
in the wild-type were 0.1 and 0.2, respectively. Following two results strongly suggest
that the alteration of amino acid contents resulted directly from overexpression of the
GGAT1 genes. First, there were strong correlations between GGAT activity and the levels
of GGAT1 mRNA, serine, glycine, and citrulline (Table I). Second, all transgenic lines
accumulated serine, glycine, and citrulline (Fig. 4). Furthermore, the accumulation of
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

9
serine suggested that active conversion of glycine to serine is occurring and that SGAT
activity is limited. The increased total amino acid contents in the overexpression lines
reveal the possibility of improving nitrogen use efficiency by creating GGAT1
overexpression lines.
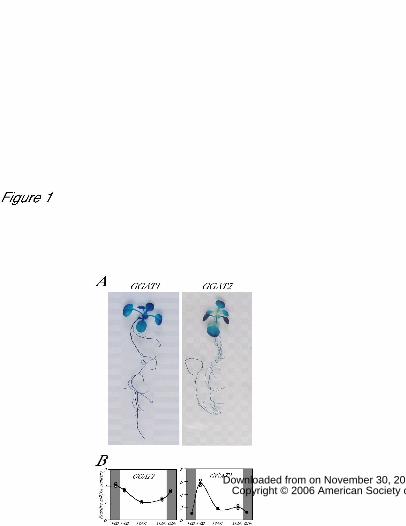
Serine accumulation in the GGAT1 overexpression line of rice
We generated transgenic rice that expressed the Arabidopsis GGAT1 cDNA. The
expression of the GGAT1 cDNA under the control of the Arabidopsis GGAT1 promoter
did not lead to overproduction of GGAT1 in transgenic rice (data not shown). Instead,
the use of the rice Cab promoter, which is known to direct high levels of expression in
photosynthetic tissues (Sakamoto et al., 1991) successfully increased GGAT1 mRNA
levels and serine contents in the leaves, as was the case in Arabidopsis (Fig. 5).This
suggests that basic mechanisms for amino acid metabolic regulation by means of
photorespiration processes may be similar between dicots and monocots.
Photorespiration and amino acid accumulation
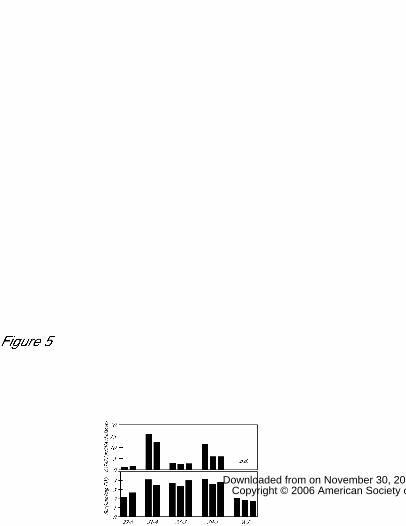
To confirm that the increased levels of serine, glycine, and citrulline in GGAT1
overexpression lines resulted directly from alterations in the photorespiration pathways,
we analyzed the amino acid contents of transgenic plants grown under conditions that
would suppress photorespiration (3,000 ppm). GGAT1 overexpression line GTox-17
accumulated serine, glycine, and citrulline at 13.4, 3.2, and 2.4 times the respective levels
observed in the wild-type (Col-0) under normal CO2 conditions, versus 5.2, 1.9, and 1.9
times the respective levels in the wild-type under high CO2 conditions (Fig. 6). The
reduction of accumulation levels under high CO2 conditions in GGAT1 overexpression
lines also observed, indicating that accumulation of serine, glycine and citrulline
depended on photorespiration. Furthermore serine was not accumulated in GGAT1
knockout line, suggesting that almost of serine is synthesized through photorespiration.
Diurnal changes in amino acid levels
To investigate the relation of amino acid metabolisms with photorespiration, we
analyzed diurnal changes in amino acid.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

10
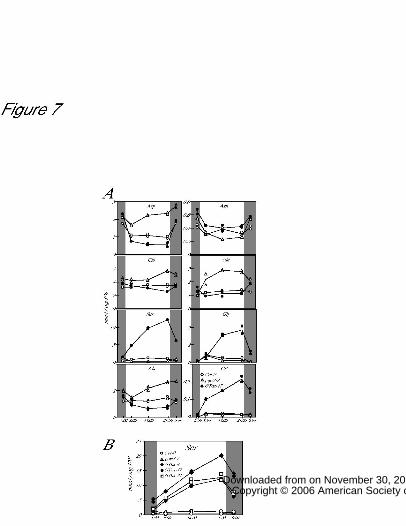
The levels of serine, glycine, and citrulline increased during the morning, remained
relatively constant during the afternoon, and then declined after sunset in the wild-type
(Fig. 7A). In contrast, the levels of these amino acids continued to increased throughout
the daytime in the GGAT1 overexpression lines. Serine increased most dramatically, to
20 times the level in the wild-type. The increase in serine depended on GGAT activity in
the GGAT1 overexpression line during the day (Fig. 3, 4). Therefore, it is possible that
GGAT activity is a limiting factor for serine biosynthesis in the wild-type.
Since GGAT activity did not change during the daytime (Fig. 2A), it is concluded that
the substrate supply is the limiting factor for serine biosynthesis in the GGAT1
overexpression lines. This hypothesis is supported by the observation that overexpression
line GTox-4, which expressed the highest level of GGAT1 mRNA, and two other lines
(GTox-17 and GTox-24) with levels nearly as high, showed a similar rate of increase in
serine (Fig. 7B).
Glutamate and alanine decreased in the GGAT1 overexpression lines and increased in
the knockout line (ggat1-1), suggesting that these amino acids as amine donors for the
GGAT reaction. Glutamine and aspartate accumulation in the knockout line would result
from excess amino groups that were transferred into pools of these amino acids.
Differences in nitrogen source affect the amino acid profiles
Amino acid profiles in higher plants are modulated by the composition of the nitrogen
source exogenously supplied to the growth medium (Raab and Terry, 1995). We
therefore compared the amino acid contents between the wild-type and the GGAT1
overexpression lines grown in different nutrient solutions.
As shown in Table II, the GGAT1 overexpression lines grown in PNS medium or in
PNS + NH4NO3 medium both accumulated serine, glycine, and citrulline compared with
the wild-type. Glutamine and asparagine accumulated in plants grown in the PNS +
NH4NO3 medium, but not in plants grown in the PNS medium. Although arginine was
not detected in plants grown in PNS medium, it accumulated in the GGAT1
overexpression lines grown in PNS + NH4NO3 medium. Other than these amino acids,
the different nitrogen sources did not appear to affect the amino acid contents. These
modulations in the amino acid profiles appear to reflect the efficiency of ammonium use.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

11
Amino acid accumulation in sink tissues
We investigated the effects of GGAT1 overexpression on amino acid profiles in the sink
tissues. In the GGAT1 overexpression lines, serine and glycine both accumulated much
more in sink tissues than was the case in the wild-type and knockout lines (ggat1-1; Fig.
8). However, serine and glycine levels in the ggat1-1 appeared to be comparable to those
in the wild-type in both stmes and silique. Although the mechanism is unclear, it appears
that alteration of metabolic regulation by photorespratory GGAT improved nutrient levels
in the sink tissues.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

12
DISCUSSION
Accumulation of serine, glycine, and citrulline in GGAT1 overexpression lines
More than 25% of the carbon fixed during photosynthesis may be used up by
photorespiration (Sharkey, 1988). In addition, leaf mitochondria generate ammonium at
an extraordinary rate during photorespiration, as much as 50 times the rate of nitrate
reduction (Ogren, 1984). This toxic ammonium is efficiently reassimilated into glutamine
and glutamate by GS-2 and Fd-GOGAT, and these are then used to produce nitrogenous
compounds needed for plant growth. Thus, photorespiratory amino acid metabolic
pathways appear to be important for nitrogen metabolism in plants.
GGAT1 overexpression lines accumulated high levels of serine, glycine, and citrulline
compared with the wild-type. High positive correlations between levels of these amino
acids, GGAT1 expression, and GGAT enzymatic activities were observed under
photosynthetic conditions (Fig. 3). These amino acids accumulated after transfer of the
plants into the light (Fig. 7). In contrast, the rate of accumulation decreased under high
CO2 (Fig. 6). These results demonstrate that GGAT1 overexpression lines accumulate
these amino acids at rates determined by the rate of photorespiration. Although the level
of GGAT1 mRNA declined from morning to midday in both the wild-type and the GTox-
17 GGAT1 overexpression line (Fig. 2B), the decrease was much larger in GTox-17 and
GGAT activity did not change in either group of plants. This might result from GGAT2
accumulation after light irradiation and the stability of GGAT proteins. Throughout the
day, serine, glycine, and citrulline increased in the GTox-17 GGAT1 overexpression line
(Fig. 7A) even though GGAT activity did not increase (Fig. 2A), suggesting that shortly
after light exposure, the limiting factor for accumulation of these amino acids is not
GGAT activity but rather the supply of substrate for the GGAT reactions during
photorespiration. Thus, when the rate of photorespiration is high, GGAT activity may be
a limiting factor in the photorespiratory cycle.
SGAT also plays a major role in controlling the flux of carbon toward glycine, and in
supplying the substrate for serine synthesis (Somerville and Ogren, 1980). In this way
GGAT and SGAT could function cooperatively in the production of the photorespiratory
amino acid glycine.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

13
Conversion of glycine into serine and energy consumption
The positive correlation between serine content and GGAT activity (Fig. 3) was
stronger than that for glycine, which is synthesized directly from the GGAT reaction. The
absolute serine content was also much higher than that of glycine (Table II). The
conversion of glycine into serine is catalyzed by glycine decarboxylase (GDC) and
SHMT and is accompanied by the generation of NH3, CO2, and NADH (Oliver and
Raman, 1995). GDC is extremely abundant in mitochondria isolated from leaves (Douce
and Neuburger, 1999). Therefore, the conversion of glycine into serine should be a stable
and active reaction. NADH generated by this reaction might be translocated to the
peroxisomes, where it would serve as a substrate for the reduction of hydroxypyruvate to
glycerate by hydroxypyruvate reductase (Raghavendra et al., 1998). In addition, recent
research has suggested that ammonium and NADH generated during glycine conversion
for glutamine and citrulline synthesis is catalyzed by GS-2, carbamoylphosphate
synthetase, and ornithine carbamoyltransferase in the mitochondria (Taira et al., 2004;
Linka and Weber, 2005). Glycine-driven oxidative phosphorylation should yield roughly
three ATP per glycine consumed. In the conversion of ornithine to citrulline, one ATP is
consumed for glutamine synthesis and two ATP are consumed for carbamoylphosphate
synthesis, allowing tight coupling of the phosphorylation and synthesis reactions (Taira et
al., 2004). Citrulline is converted into arginine (Ludwig, 1993). Because the citrulline
contents increased greatly after transfer of plants into the light in GGAT1 overexpression
lines and in the wild-type (Fig. 7A), the arginine content in GGAT1 overexpression line
grown in a medium with supplemental ammonium was higher than that in the wild-type
(Table II). Our results are thus consistent with reported pathways for the metabolism of
nitrogen released by photorespiration. The absolute contents of citrulline and arginine
that accumulated in the GGAT1 overexpression line were low compared with those of
glutamine and glutamate, demonstrating that mitochondrial reassimilation of ammonium
is a minor pathway.
Analysis of the function of GGAT in monocot
Transgenic rice that expressed Arabidopsis GGAT1 also accumulated serine. Rice also
has predicted GGAT-like genes in both the O. japonica (BAC20747) and O. indica
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

14
(AAO84040) cultivars (International Rice Genome Sequencing Project, 2005). The
amino-acid sequence of the protein encoded by Arabidopsis GGAT1 is 80% similar to
that of the rice GGAT1-like proteins. These two groups of proteins contain a
serine/arginine/methionine group at the C-terminus that is a conserved PTS1. These
sequence characteristics support the hypothesis that the rice GGAT1-like proteins
function as photorespiratory GGATs. This suggests that the regulation of
photorespiratory glyoxylate aminotransferase is similar in a dicot (Arabidopsis) and a
monocot (rice). However, the degree of serine accumulation in rice was not remarkable
compared with that in Arabidopsis, and other than serine, we detected no marked
differences in amino acid accumulation between the wild-type and the GGAT1
overexpression line of rice. This could result from instability in GGAT1 originating in a
heterologous expression system, slight differences in the metabolite flux during
photorespiration, and small absolute levels of GGAT1 mRNA.
Photorespration and nitrogen
Nitrogen, an essential nutrient for plant growth, is primarily taken up by roots in the
form of nitrate and is reduced to ammonium by nitrate reductase and nitrite reductase. In
most higher plants, ammonium is assimilated into glutamine through the cooperative
activity of GOGAT and GS. These reactions represent the major pathway for the
assimilation of ammonium into amino acids. At elevated CO2 concentrations,
photorespiration is limited and the assimilation of nitrogen is inhibited in the shoots of
dicotyledons (e.g., Arabidopsis) and monocotyledons (e.g., wheat; Bloom et al., 2002;
Rachmilevitch et al., 2004). Thus, under normal conditions, photorespiration is assumed
to contribute to effective use of nitrogen.
Our results demonstrated that in an ammonium-rich medium, levels of Glutamine,
asparagine, and arginine increased in the both of the wild-type and the GGAT1
overexpression lines (Table II), suggesting that these amino acids form a nitrogen pool
under nitrogen-rich conditions. However, the increase in levels of these amino acids in
the GGAT1 overexpression line in response to supplemental ammonium was much higher
than the increase in the wild-type. This suggests that increased release of ammonia during
the conversion of glycine into serine and uptake of ammonium by the roots, ammonium
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

15
increases to excessive levels and would be assimilated into less-toxic glutamine,
asparagine, and arginine. The high accumulation of total amino acids in the GGAT1
overexpression line may show that improvement in GGAT activity in this line can
enhance photorespiration and the efficiency of nitrate and ammonium assimilation.
Substrates for GGAT reaction
The analysis of GGAT- and SGAT-deficient mutants and of recombinant proteins
indicated that alanine is a substrate for both GGAT and SGAT (Murray et al., 1987;
Liepman and Olsen, 2001, 2003; Igarashi et al., 2003). GGAT catalyzes an AGT reaction
(Noguchi and Hayashi, 1981; Nakamura and Tolbert, 1983; Yu et al., 1984, Igarashi et
al., 2003; Liepman and Olsen, 2003). However, a high Km value of SGAT for alanine
(101.2 mM) indicates that alanine may not be a physiological substrate for SGAT
(Liepman and Olsen, 2001).
The diurnal glutamate and alanine profiles differed between the knockout line (ggat1-1)
and the overexpression line (Fig. 7A). From morning (8:00) to night (0:00), these amino
acids increased in ggat1-1, and decreased in the overexpression line. Both glutamate and
alanine are used as amine donors for transamination to glyoxylate in the GGAT reaction.
Estimates based on whole-leaf labeling analysis suggested that alanine contributed three
times more amino groups to photorespiratory glycine formation than glutamate did
(Betsche, 1983), whereas another study based on feeding metabolites to purified
peroxisomes concluded that glutamate contributed more to glyoxylate transamination
than alanine did (Yu et al., 1984). The cooperative change of glutamate and alanine in the
ggat1-1 knockout line and the GGAT1 overexpression line suggests that both amino acids
contribute to transamination for glycine formation.
Amino acid accumulation in sink tissues
We showed that serine and glycine contents increased in sink tissues (Fig. 8). Although
this may be a result of photorespiration in stem and silique, accumuration level of serine
was higher than that of in leaves, suggesting that amino acids in the source tissues would
be directly transported to sink tissues and concentrated. Because photorespiration is
involved in photosynthesis and metabolisms of nitrogen and amino acid, and protection
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

16
against stress, it is a proposed target for genetic engineering to improve crop productivity.
However, further analysis of the flux of photorespiratory metabolites and of the transport
and accumulation mechanisms between source and sink tissues will be necessary to
permit effective improvement of crop production by engineering of the genes involved in
photorespiration.
In summary, our analysis of the ggat1-1 knockout and GGAT1 overexpression lines
suggested that GGAT activity is directly involved in the regulation of glycine and serine
levels. Furthermore, the total amino acid content and levels of other major amino acids
differed markedly between these two lines, suggesting that GGAT may regulate the
biosynthesis and metabolism of fundamental amino acids.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

17
MATERIALS AND METHODS
Plant materials and growth conditions
Arabidopsis thaliana L. (Col-0) cultures were grown at photosynthetically active
radiation (PAR) of 70 µmol m-2 s-1 on a 16:8-hr L:D photoperiod at 22 °C unless
otherwise indicated. A growth camber (EYELA FLI-301NH-CL) was used for the
analysis of plants grown under high CO2 concentrations. Plants were grown on sterile
plant nutrients and sugar (PNS) medium (Haughn and Somerville, 1986) in petri dishes
containing 0.8% (w/v) agar.
To compare the amino acid contents between the wild-type and the GGAT1
overexpression lines grown under different nitrogen sources, these plants were grown on
PNS medium with or without supplementation with 10 mM NH4NO3. PNS medium
contains 5 mM KNO3 and 2 mM Ca(NO3)2 as nitrogen sources and an additional 10 mM
NH4NO3 almost doubled nitrogen in the medium. The plants were grown for 2 weeks on
PNS medium or on PNS + NH4NO3 medium, and the aboveground tissues were collected
for amino acid analysis.
To determine the amino acid contents in the plants grown under high CO2 conditions or
in the siliques and stems, the plants were grown in soil watered with PN medium at PAR
of 120µmol m-2 s-1. Arabidopsis siliques (9 to 11 days after flowering), stems (cut 15 cm
above the roots),and fully expanded leaves were harvested for analysis.
Rice (Oryza sativa L. cv ‘Kitaake’) was planted in soil watered with 1,000-fold diluted
Hyponex (Hyponex, Osaka) and grown under natural light conditions on a 14:10-hr L:D
photoperiod at 23°C day/18 °C night cycle in a greenhouse (Tsuchida et al., 2001).
Expression analysis of GGAT
For reverse-transcriptase PCR (RT-PCR), total RNA was isolated from leaves using an
RNeasy Plant Mini Kit (Qiagen). First-strand cDNA synthesis was conducted with an
oligo dT 12-18 primer (GE Healthcare) and reverse transcriptase, superscript II
(Invitrogen). Real-time PCR reactions were performed using an Applied Biosystems
Prism 7700 Sequence Detector. Specific primers for GGAT1 (5'-
TTCTTCTTCTGAACGACTATTGTG-3' and 5'-
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

18
GAATAGGGCAAAGAGAAAGAGTG-3') and for GGAT2 (5'-
TCACTTCTTCTTCTTTCACGACA-3' and 5'-GAGTTTGACACAGAGTAGGACCA-
3') were used.
For rice analysis, specific primers for GGAT1 (5'-TGAAAGCAAGGGGATTCTTG-3'
and 5'-GACGTTTTTGCAGCTGTTGA-3') were used for quantification of the
introduced gene.
Real-time PCR reactions were performed in duplicate using 0.9 µM of each primer and
1 x SYBR green PCR master mix (Applied Biosystems) in a 50-µL volume. Relative
differences were determined using the deltaCt method described by the manufacturer.
Construction of plasmids and transformation of Arabidopsis
The promoter regions were isolated by means of PCR using specific primers for GGAT1
(5'-CAATAACAATGCAAAGTTAAGATTCGGATC-3' and 5'-
GACGGATCCCATTTATGTTCACTCTGAACC-3') and for GGAT2 (5'-
GACAAGCTTTGAGTTCGGACACGGCTCTA-3' and 5'-
GACGGATCCCATTTCCAGGATTTACTTTTC-3'). The amplified fragments were
digested by BamHI (GGAT1) or HindIII/BamHI (GGAT2) and the fragments (1843 bp for
GGAT1 and 2k bp for GGAT2) were cloned into the 5'-end of the GUS gene in vector
pBI101.
For generation of a GGAT1 overexpression line of Arabidopsis, a 5089-bp genomic
fragment of GGAT1, that extends 5'-upstream (1843 bp) from the translational initiation
and 3'-downstream (612 bp) from the point of translational termination, was isolated by
means of PCR using specific primers (5'-
CAATAACAATGCAAAGTTAAGATTCGGATC-3' and 5'-
GCTTCTTCTCAACCATCGTCACC-3'). The amplified fragment was cloned into vector
pBI121 between the HindIII and EcoRI sites and introduced into the wild-type (Col-0) ,
or an GGAT1 knockout line (ggat1-1) by means of Agrobacterium tumefaciens–mediated
transformation. Two of 42 lines are ggat1-1 background and others are wild-type
background.
Generation of a GGAT1 overexpression line of rice
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

19
Arabidopsis GGAT1 cDNA was isolated by means of RT-PCR using specific primers
(5'-GGCTCTAGATGGCTCTCAAGGCATT-3' and 5'-
GCCGAGCTCTCACATTTTCGAATAA-3'). The amplified fragment was fused to the
Cab promoter (Sakamoto et al., 1991) and cloned into the vector pIG121Hm. The
resultant plasmid was introduced into calli derived from rice by means of A. tumefaciens–
mediated transformation.
Histochemical staining for GUS activity
Histochemical staining for GUS activity was carried out as described by Jefferson et al.
(1987), but with the following modifications. After transgenic plants had been grown for
2 weeks on the PNS–agar plates, plants were harvested and incubated overnight in GUS
staining solution [100 mM sodium phosphate (pH 7.0), 10 mM EDTA, 0.1% (v/v) triton
X-100, 1 mM potassium ferricyanide, 1 mM potassium ferrocyanide and 1 mM X-Gluc]
at 37 °C. After staining, samples were treated with 70% ethanol to remove chlorophyll
from the tissues.
Enzyme assays
Aminotransferase activity was assayed as described by Hørder and Rej (1983), but with
the following modifications. The aboveground tissues of seedlings grown on PNS–agar
plates at PAR of 70 µmol m-2 s-1 were harvested, frozen, and ground into a fine powder
using an MM 300 mixer (Qiagen). The ground powder was added to 0.5 mL of grinding
buffer [100 mM Tris-HCl (pH 7.3) and 10 mM dithiothreitol] and gently mixed to ensure
complete suspension. The suspension was then centrifuged (13,000 x g for 10 min at 4
°C) to remove insoluble material. The resulting supernatant (0.5 mL) was applied to a
UFV5BGC00 Ultrafree filter (Millipore) for concentration and desalination. The total
protein concentration was assayed using a Bio-Rad protein assay system
GGAT activity was measured by coupling the reduction of 2-oxoglutarate by NADH in
a reaction catalyzed by GDH. The reaction was assayed in a mixture containing 100 mM
Tris-HCl (pH 7.3), 20 mM glutamate, 1 mM glyoxylate, 0.18 mM NADH, 0.11 mM
pyridoxyal 5-phosphate, 83 mM NH4Cl, and 0.3 U GDH in a final volume of 0.6 mL.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

20
Activity was assayed spectrophotometrically by monitoring NADH oxidation at 340 nm.
The GGAT assays used 50 µg of crude extract. All reactions were performed at 30 °C.
Measurement of amino acid contents
Amino acids were extracted from the aboveground tissues of the plants in 80% ethanol
at 80 °C. After evaporation, dried samples were dissolved in 0.02 N HCl. Amino acid
contents were determined using an L-8800 amino acid analyzer (Hitachi). Briefly, amino
acids, separated by cation-exchange chromatography, were detected
spectrophotometrically after postcolumn reaction with ninhydrin reagent (Noguchi et al,
2006).
ACKNOWLEDGMENTS
We thank Kiyoshi Miwa, Yutaka Ishiwatari, Hiroyuki Kiyohara, and Ai Koga for
helpful discussion and technical assistances during this work.
LITERATURE CITED
Betsche T (1983) Aminotransfer from alanine and glutamate to glycine and serine during
photorespiration in oat leaves. Plant Physiol 71: 961-965
Bloom AJ, Smart DR, Nguyen DT, Searles PS (2002) Nitrogen assimilation and
growth of wheat under elevated carbon dioxide. Proc Natl Acad Sci USA 99: 1730-
1735
Bourguignon J, Rebeille F, Douce R (1999) Serine and glycine metabolism in higher
plants. In BK Singh, ed, Plant Amino Acids: Biochemistry and Biotechnology.
Marcel Dekker, New York, pp 111-146
Douce R, Neuburger M (1999) Biochemical dissection of photorespiration. Curr Opin
Plant Biol. 2: 214-222
Gerbaud A, Andre M (1979) Photosynthesis and photorespiration in whole plants of
wheat. Plant Physiol 64: 735-738
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

21
Haughn, GW, Somerville C (1986) Sulfonylurea-resistant mutants of Arabidopsis
thaliana. Mol Gen Genet 204: 430-434
Hørder M, Rej R (1983) Alanine aminotransferase. In HU Bergmeyer, J Bergmeyer, M
Grabl, eds, Methods of Enzymatic Analysis, Ed 3, Vol 3. Verlag Chemie, Weinheim,
pp 444-456
Hoshida H, Tanaka Y, Hibino T, Hayashi Y, Tanaka A, Takabe T, Takabe T (2000)
Enhanced tolerance to salt stress in transgenic rice that overexpresses chloroplast
glutamine synthetase. Plant Mol Biol 43: 103-111
Igarashi D, Miwa T, Seki M, Kobayashi M, Kato T, Tabata S, Shinozaki K, Ohsumi
C (2003) Identification of photorespiratory glutamate:glyoxylate aminotransferase
(GGAT) gene in Arabidopsis. Plant J 33: 975-987
International Rice Genome Sequencing Project. (2005) The map-based sequence of
the rice genome. Nature 436: 793-800
Jefferson RA, Kavanagh TA, Bevan MW (1987) GUS fusions: beta-glucuronidase as a
sensitive and versatile gene fusion marker in higher plants. EMBO J. 20: 3901-3907
Keys AJ, Bird IF, Cornelius MJ, Lea PJ, Wallsgrove RM, Miflin BJ (1978)
Photorespiratory nitrogen cycle. Nature 275: 741-743
Kozaki A, Takeba G (1996) Photorespiration protects C3 plants from photooxidation.
Nature 384: 557-560
Leegood RC, Lea PJ, Adcock MD, Hausler RE (1995) The regulation and control of
photorespiration. J Exp Bot 46: 1397-1414
Liepman AH, Olsen LJ (2001) Peroxisomal alanine:glyoxylate aminotransferase
(AGT1) is a photorespiratory enzyme with multiple substrates in Arabidopsis
thaliana. Plant J 25: 487-498
Liepman AH, Olsen LJ (2003) Alanine aminotransferase homologs catalyze the
glutamate:glyoxylate aminotransferase reaction in peroxisomes of Arabidopsis. Plant
Physiol 131: 215-227
Linka M, Weber AP (2005) Shuffling ammonia between mitochondria and plastids
during photorespiration. Trends Plant Sci 10: 461-465
Ludwig RA (1993) Arabidopsis chloroplasts dissimilate L-arginine and L-citrulline for
use as N source. Plant Physiol 101: 429-435
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

22
Murray AJS, Blackwell RD, Joy KW, Lea PJ (1987) Photorespiratory N donors,
aminotransferase specificity and photosynthesis in a mutant of barley deficient in
serine:glyoxylate aminotransferase activity. Planta 172: 106-113
Nakamura Y, Tolbert NE (1983) Serine:glyoxylate, alanine:glyoxylate, and
glutamate:glyoxylate aminotransferase reactions in peroxisomes from spinach leaves.
J Biol Chem 258: 7631-7638
Noguchi T, Hayashi S (1981) Plant leaf alanine:2-oxoglutarate aminotransferase
Peroxisomal localization and identity with glutamate:glyoxylate aminotransferase.
Biochem J 195: 235-239
Noguchi Y, Zhang QW, Sugimoto T, Furuhata Y, Sakai R, Mori M, Takahashi M,
Kimura T (2006) Network analysis of plasma and tissue amino acids and the
generation of an amino index for potential diagnostic use. Am J Clin Nutr 83: 513S-
519S
Ogren WL (1984) Photorespiration: pathways, regulation, and modification. Annu Rev
Plant Biol 35: 415-442
Oliver DJ, Raman R (1995) Glycine decarboxylase: protein chemistry and molecular
biology of the major protein in leaf mitochondria. J Bioenerg Biomembr 27: 407-414.
Raab TK, Terry N (1995) Carbon, nitrogen, and nutrient interactions in Beta vulgaris L.
as influenced by nitrogen source, NO3- versus NH4
+. Plant Physiol 107: 575-584
Rachmilevitch S, Cousins AB, Bloom AJ (2004) Nitrate assimilation in plant shoots
depends on photorespiration. Proc Natl Acad Sci USA 101: 11506-11510
Raghavendra AS, Reumann S, Heldt HW (1998) Participation of mitochondrial
metabolism in photorespiration Reconstituted system of peroxisomes and
mitochondria from spinach leaves. Plant Physiol 116: 1333-1337
Sakamoto M, Sanada Y, Tagiri A, Murakami T, Ohashi Y, Matsuoka M (1991)
Structure and characterization of a gene for light-harvesting Chl a/b binding protein
from rice. Plant Cell Physiol 32: 385–393
Sharkey TD (1988) Estimating the rate of photorespiration in leaves. Physiol Plant 73:
147-152
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

23
Somerville CR, Ogren WL (1980) Photorespiration mutants of Arabidopsis thaliana
deficient in serine-glyoxylate aminotransferase activity. Proc Natl Acad Sci USA 77:
2684-2687
Somerville SC, Somerville CR (1983) Effect of oxygen and carbon dioxide on
photorespiratory flux determined from glycine accumulation in a mutant of
Arabidopsis thaliana. J Exp Bot 34: 415-424
Taira M, Valtersson U, Burkhardt B, Ludwig RA (2004) Arabidopsis thaliana GLN2-
encoded glutamine synthetase is dual targeted to leaf mitochondria and chloroplasts.
Plant Cell 16: 2048-2058
Tolbert NE (1980) Photorespiration. In DD Davis, ed, The Biochemistry of Plants. Vol 2
Academic Press, New York, pp 487-523
Tsuchida H, Tamai T, Fukayama H, Agarie S, Nomura M, Onodera H, Ono K,
Nishizawa Y, Lee BH, Hirose S, Toki S, Ku MS, Matsuoka M, Miyao M (2001)
High level expression of C4-Specific NADP-malic enzyme in leaves and impairment
of photoautotrophic growth in a C3 plant, rice. Plant Cell Physiol 42: 138-145
Wingler A, Lea PJ, Quick WP, Leegood RC (2000) Photorespiration: metabolic
pathways and their role in stress protection. Philos Trans Roy Soc London B 355:
1517-1529
Yu C, Liang Z, Huang AHC (1984) Glyoxylate transamination in intact leaf
peroxisomes. Plant Physiol 75: 7-12
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

24
FIGURE LEGENDS
Figure 1
Expression of GGAT1 and GGAT2 in Arabidopsis.
A, Analysis of tissue-specific expression by means of histochemical staining of 2-week-
old seedlings harboring promoter–GUS constructs.
B, Diurnal changes in mRNA contents
Time-points were 2, 8, and 15 hours after exposure to light and 2 and 7 hours after
darkness resumed. Relative amounts of mRNA were measured by means of RT-PCR and
calculated using the deltaCt method. Seedlings grown on two independent plates were
used for samples. Values represent means (black circles) and maximum and minimum
values (white circles). Within each graph, values are normalized by expressing them as a
multiple of the lowest value recorded during the day.
Figure 2
Diurnal changes in GGAT enzymatic activities and mRNA levels in the GGAT1
overexpression line of Arabidopsis.
Seedlings of Col-0 and the GTox-17 GGAT1 overexpression line were grown on PNS–
agar plates for 2 weeks. Aboveground tissues were harvested at each sampling time.
A, Durnal change in GGAT enzymatic activities. The methods of enzyme assay is
described in the text.
B, Relative amounts of mRNA were measured by means of real-time PCR and calculated
using the deltaCt method same as described in legend of figure 1.
Figure 3
Correlations between amino acid contents, GGAT activities, and GGAT1 mRNA levels.
Nine independent GGAT1 overexpression line of Arabidopsis that exhibited a wide range
of levels of GGAT1 mRNA were grown on PNS–agar plates for 2 weeks. Aboveground
tissues were harvested at midday and amino acid contents, GGAT activities, and GGAT1
mRNA levels were determined. The assay methods are described in the text. Data are
only presented for correlations that were significant in Table I. Values represent the
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

25
relative amounts compared with the value for the wild-type (Col-0). R means correlation
coefficient.
Figure 4
Amino acid contents in the GGAT1 overexpression line of Arabidopsis.
Forty-two independent GGAT1 overexpression lines that expressed various levels of
GGAT1 mRNA, as well as a knockout line (ggat1-1) and the wild-type (Col-0), were
grown on PNS–agar plates for 2 weeks. Aboveground tissues were harvested at midday
and amino acid contents were determined. In the graph of total amino acid contents, dark
and white bars indicate serine and other protein composition amino acid with citrulline,
respectively.
Figure 5
Levels of GGAT1 mRNA and serine accumulation in a GGAT1 overexpression line of
rice.
Ten independent T2 transgenic plants derived from four independent GGAT1
overexpression lines (lines 27-6, 31-4, 35-3, and 39-3) and the wild-type were grown on
soil for 6 weeks. The middle sections of the leaves were harvested at midday and GGAT1
mRNA levels and serine contents were determined. Relative amounts of mRNA were
measured by means of real-time PCR and calculated using the deltaCt method. Within
each graph, values are normalized by expressing them as a multiple of the lowest value
(line 27-6) recorded in GGAT1 overexpression line of rice. The n.d. means not detected.
Figure 6
Effects of CO2 concentration on the accumulation of amino acids.
Arabidopsis plants of the wild-type (Col-0), GGAT1 knockout line (ggat1-1), and GGAT1
overexpression line (GTox-17) were grown in soil under normal (3,000 ppm) or high
(300 ppm) CO2 levels for 3 weeks. Aboveground tissues were harvested and amino acid
contents were determined.
Figure 7
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

26
Diurnal changes in amino acid contents.
Arabidopsis seedlings of the wild-type (Col-0), GGAT1 knockout line (ggat1-1), and
GGAT1 overexpression line (GTox-17) were grown on PNS–agar plates for 2 weeks.
Values represent the means from two dependent samples from dishes (line) and the
maximum and minimum values (each symbol).
A, Aboveground tissues were harvested at each time and amino acid contents were
determined. B, Serine levels in the wild-type, in the knockout line (ggat1-1), and in three
independent overexpression lines (GTox-4, -17 and -24).
Figure 8
Serine and glycine accumulation in stem and silique.
Arabidopsis plants of the wild-type (Col-0), GGAT1 knockout line (ggat1-1), and GGAT1
overexpression line (GTox-17) were grown in soil for 6 weeks. Stems were harvested 15
cm from the base. Siliques were harvested 9 to 11 days after flowering. All samples
including leaves were harvested at midday.
Table I
Correlations between GGAT activity and each amino acid that was assayed, as well as
between GGAT activity and the levels of GGAT1 mRNA.
Col-0 (wild-type) and GGAT1 overexpression lines (GTox) were grown and amino acid
contents and mRNA levels were determined using the methods described in the text.
Table II
Amino acids pool size in GGAT1 overexpression lines
Each line was grown on PNS medium or on PNS + 10mM NH4NO3 medium on agar
plates for 2 weeks. Aboveground tissues were harvested at midday and amino acid
contents were determined. Seedling were grown in 4 separate plates. Value represent the
average for each experiment, with the S.D. in parentheses.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.
27
Table I
amino acid correlation
Ser 0.97
Cit 0.94
Gly 0.74
Asn 0.49
Gln 0.46
Ala -0.21
Glu -0.40
Asp -0.49
mRNA 0.84
TableII
PNS nmol/mg FW Fold
Col-0 GTox-17 GTox-24 GGT-17 GGT-24
Asp 1.04 ( 0.04 ) 0.74 ( 0.04 ) 0.56 ( 0.10 ) Asp 0.72 0.54
Asn 0.28 ( 0.03 ) 0.33 ( 0.02 ) 0.35 ( 0.02 ) Asn 1.21 1.25
Glu 2.91 ( 0.10 ) 2.45 ( 0.05 ) 2.56 ( 0.10 ) Glu 0.84 0.88
Gln 1.01 ( 0.10 ) 0.89 ( 0.03 ) 0.96 ( 0.03 ) Gln 0.88 0.94
Ser 0.96 ( 0.03 ) 7.47 ( 0.15 ) 8.96 ( 0.24 ) Ser 7.76 9.31
Gly 0.11 ( 0.01 ) 0.57 ( 0.02 ) 0.68 ( 0.05 ) Gly 5.09 6.09
Ala 0.52 ( 0.10 ) 0.23 ( 0.05 ) 0.29 ( 0.01 ) Ala 0.45 0.56
Arg ND ND ND Arg - -
Cit ND 0.11 ( 0.01 ) 0.12 ( 0.00 ) Cit - -
PNS+10mM NH4NO3 nmol/mg FW Fold
Col-0 GTox-17 GTox-24 GGT-17 GGT-24
Asp 0.78 ( 0.10 ) 0.75 ( 0.10 ) 0.55 ( 0.10 ) Asp 0.96 0.71
Asn 0.77 ( 0.12 ) 2.16 ( 0.26 ) 2.56 ( 0.11 ) Asn 2.80 3.33
Glu 2.28 ( 0.15 ) 2.20 ( 0.14 ) 2.56 ( 0.23 ) Glu 0.97 1.12
Gln 2.20 ( 0.34 ) 4.55 ( 0.66 ) 5.43 ( 0.98 ) Gln 2.07 2.47
Ser 1.43 ( 0.13 ) 12.34 ( 0.34 ) 14.98 ( 1.26 ) Ser 8.61 10.45
Gly 0.60 ( 0.09 ) 3.22 ( 0.43 ) 3.30 ( 0.45 ) Gly 5.40 5.53
Ala 0.57 ( 0.05 ) 0.44 ( 0.11 ) 0.38 ( 0.02 ) Ala 0.77 0.67
Arg 0.55 ( 0.13 ) 1.13 ( 0.02 ) 1.25 ( 0.09 ) Arg 2.04 2.27
Cit 0.07 ( 0.01 ) 0.34 ( 0.02 ) 0.43 ( 0.04 ) Cit 5.03 6.30
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.
www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.

www.plantphysiol.orgon November 30, 2018 - Published by Downloaded from Copyright © 2006 American Society of Plant Biologists. All rights reserved.



![Bile Acid Sodium Symporter BASS6 Can Transport Glycolate ...fluorescence (Badger et al., 2009; Sun et al., 2009). When illu-minated in low [CO 2], photorespiration-deficient mutants](https://img.dokumen.tips/doc/110x75/5e84656e050e0863441b9bd8/bile-acid-sodium-symporter-bass6-can-transport-glycolate-iuorescence-badger.jpg)

























