Embed Size (px)
Citation preview

1
Coordinated ATP - hydrolysis by the Hsp90 dimer
Klaus Richter, Paul Muschler, Otmar Hainzl and Johannes Buchner*
Institut für Organische Chemie und Biochemie
Technische Universität München
Lichtenbergstr.4, 85747 Garching
* To whom correspondence should be adressed
Phone: 0049-89-28913340
Fax: 0049-89-28913345
E-Mail: [email protected]
Running Title: Coordinated ATP - hydrolysis of Hsp90
Keywords: Hsp90, ATPase, dimerization, heterodimers, gyrase
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on July 5, 2001 as Manuscript M103832200 by guest on February 21, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2
Abstract
The Hsp90 dimer is a molecular chaperone with an unusual N-terminal ATP binding site. The
structure of the ATP binding site makes it a member of a new class of ATP hydrolysing
enzymes, known as the GHKL-family. While for some of the family members structural data
on conformational changes occuring after ATP binding are available, these are still lacking for
Hsp90. Here we set out to investigate the correlation between dimerization and ATP
hydrolysis by Hsp90. The dimerization constant of wt Hsp90 was determined to be 60nM.
Heterodimers of wt Hsp90 with fragments lacking the ATP binding domain form readily and
exhibit dimerization constants similar to full length Hsp90. However, the ATPase activity of
these heterodimers was significantly lower than that of the wild type protein, indicating
cooperative interactions in the N-terminal part of the protein, that lead to the activation of the
ATPase activity. To further address the contribution of the N-terminal domains to the ATPase
activity, we used an Hsp90 point mutant which is unable to bind ATP. As heterodimers
between the wt protein and this mutant showed wt ATPase activity, this mutant, though
unable to bind ATP, still has the ability to stimulate the activity in its wt partner domain. Thus
contact formation between the N-terminal domains might not depend on ATP bound to both
domains. Together these results suggests a mechanism for coupling the hydrolysis of ATP to
the opening - closing movement of the Hsp90 molecular chaperone.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

3
Abbreviations
GA, Geldanamycin; HPLC, high pressure liquid chromatography; Hsp90, heat shock protein
90; Hsp90 - 262C, yeast Hsp90 fragment ranging from amino acid 262 to 709; Hsp90 - 527C,
fragment from amino acid 527 to 709 of Hsp90; Hsp90 - D79N, the D79N - point mutant of
Hsp90; Hsp90 - N210, fragment 1 to 210; Hsp90 - N272, fragment 1 to 272; Hsp90 - N529,
fragment 1 to 529; Hsp90 - N599, fragment 1 to 599; IPTG, Isopropyl-thiogalactosid; Kd,
dissociation constant for the Hsp90 dimer; Kd,app apparent dissociation constant; PAGE,
polyacrylamide gel electrophoresis; PCR, polymerase chain reaction; SDS, sodium-
dodecylsulfate; SEC, size exclusion chromatography; wt, wild type.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

4
Introduction
Hsp90 is an abundant cytosolic protein which belongs to the class of molecular chaperones.
Interaction with non-native and destabilised proteins has been shown in vitro (1, 2, 3, 4, 5). In
vivo an ever increasing number of proteins have been found to be associated with Hsp90 (6,
7). Most of the known in vivo substrates are involved in signal transduction pathways, like
tyrosin and serine/threonine kinases, steroid hormone receptors, helix-loop-helix transcription
factors and tumour supressor proteins (6, 7, 8). Other proteins, like reverse transcriptase (9) or
telomerase (10) were found to be dependent on Hsp90 action as well. Consistent with the
importance of the substrate proteins, Hsp90 was found to be an essential protein in yeast (11).
Although the mechanism of Hsp90 is still far from being understood, Hsp90 is thought to
maintain an otherwise unstable conformation of the substrate (7). ATP binding and hydrolysis
were found to be essential for the function of Hsp90 (12, 13) and competitive inhibitors for
ATP-binding, like geldanamycin were shown to be potent anti-proliferative agents (14, 15).
First indications that Hsp90 conformation is influenced by ATP were reported by Csermely et
al. (16) and Grenert et al. (17). Experiments using spin-labeled ATP showed that Hsp90 binds
ATP weakly (18). The nucleotide binding site was identified in the crystal structure of the N-
terminal domain of yeast Hsp90 in the presence of ADP (19, 20). This binding site exhibits a
new type of fold similar to that of DNA gyrase B and MutL (21). The nucleotide is bound in
an unusual kinked conformation with the adenosine base and the ribose buried inside the
protein in a cleft. The β-phosphate and probably even more the γ-phosphate of ATP are
solvent accessible in the crystal structure. Interestingly, the crystal structures of the domain in
the presence or absence of nucleotide were almost identical (19). The kinetic analysis of the
ATPase cycle of Hsp90 revealed that after ATP binding, a conformational change occurs in
Hsp90 which traps the ATP molecule (22). This trapped ATP molecule proved to be
committed to hydrolysis, as it was impossible to exchange it for unbound ATP. This
conformational change requires parts of the protein that are C-terminal to the binding site. A
conformational change was also observed in the crystal structure of the homologous protein
MutL (23, 24). MutL traps ATP by contacts between the γ-phosphate of ATP and a domain
further C-terminal from the ATP binding site. Conformational changes of Hsp90 upon ATP
binding were also reported by Prodromou et al. (25). Here, cross-linking data indicate that the
N-terminal domains associate in the presence of AMP-PNP, which had been suggested
previously based on electron microscopic data (26). This seems to be a prerequisite for the
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

5
association of Hsp90 with the co-chaperone p23, which is known to occur after ATP-binding
(27, 28, 29). In the studies of Weikl et al. (22) and Prodromou et al. (25) fragments of Hsp90
lacking C-terminal domains were found to be considerably less active, than wild-type Hsp90.
These fragments are thought to be monomeric, in contrast to full length Hsp90, which is a
dimer. The dimerization site was shown to reside in the very C-terminal domain of Hsp90 (30,
31).
To gain further insight into the ATPase mechanism of Hsp90 we investigated the importance
of dimer formation for the ATPase activity. To this end we used a set of Hsp90 deletion
mutants and analysed their quaternary structure as well as their ATPase activities. Hsp90 as
well as C-terminal fragments were shown to be dimeric with dissociation constants of
0.06µM. Analysis of the ATPase activity of these fragments demonstrated that the presence of
two N-terminal domains is required to stimulate the ATPase activity, as heterodimers formed
between C-terminal fragments and the wt protein had diminished ATPase activities.
Surprisingly, the stimulation of hydrolysis does not require ATP bound on both sides of the
dimer. The presence of two N-terminal domains is sufficient to activate ATP-hydrolysis.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

6
Material and Methods
Materials
Geldanamycin (GA) was a kind gift of the NIH, NCI Experimental Drug Division, Bethesda,
Maryland, USA.
Hsp90 constructs
Fragments of yeast Hsp90 were constructed using the plasmid pET28-Hsp82, containing the
full length Hsp82 gene of S. cerevisiae with an N-terminal His-Tag as a template. All the
PCR-fragments were cloned into the Qiagen vectors pQE30, resulting in the constructs
pQE30-N529, pQE30-N599 and pQE30-527C. In addition, fragments constructed by Scheibel
et al. (1998) were used. The mutagenesis leading to the D79N mutation in full length Hsp90
was done by overlap extention PCR in N210 using two primers conatining the mutation (5’-
CCAATACCAGAGTTTCTGATTTCC-3’ and 5’-GGAAATCAGAAACTCTGGTATT GG-
3’) and subsequent insertion of this fragment into the full-length gene, using an N-terminal
BamHI - restriction site and the internal XbaI site. The identity of all constructs was
confirmed by DNA sequencing.
Protein expression and purification
His-Hsp90 was expressed in the strain BL21 (DE3) cop+ (Stratagene, La Jolla, USA) in LBKan
and induced with 1mM IPTG. For all other constructs the strain M15 prep and 2mM IPTG
were used. Cells were lysed using a Cell Disruption System (Constant Systems, Warwick,
UK). The fragments and the His-tagged full-length protein were purified on a Chelating
Sepharose Column (Amersham Pharmacia Biotech, Uppsala, Sweden) preloaded with 100mM
NiSO4. Cell lysis and loading of the proteins was done in loading buffer (40mM KPO4, pH
8.0, 400mM KCl, 6mM imidazole). The column was washed with washing buffer (40mM
KPO4, pH 8.0, 400mM KCl, 20mM imidazole) before elution was performed in a step
gradient with washing buffer containing 300mM imidazole. A Resource Q column
(Amersham Pharmacia Biotech, Uppsala, Sweden) was used to further purify the proteins. The
protein was loaded in 50mM Tris, pH 8.0, 20mM KCl and eluted with a gradient from 20mM
to 1000mM KCl. As a final purification step, a Superdex 200 HiLoad (Amersham Pharmacia
Biotech, Uppsala, Sweden) or Superdex 75 HiLoad (Amersham Pharmacia Biotech, Uppsala,
Sweden), depending on protein size was run in 40mM HEPES, pH 7.5, 300mM KCl, 5%
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

7
glycerol. Proteins were stored in 40mM HEPES, pH 7.5, 20mM KCl, 5% glycerol at
concentrations of 1.5mg/ml to 6mg/ml at -70°C.
Protein structure and stability
Far-UV CD-Spectroscopy was used to confirm the secondary - structure of the fragments.
Measurements were performed in 40mM potassium phosphate, pH 7.0 at protein
concentrations of 200µg/ml and 20°C. CD-spectra were collected between 195nM and 250nM
on a J-715 spetropolarimeter (Jasco, Groß-Umstadt, Germany).
Urea transitions were performed with 20µg/ml protein and urea concentrations ranging from
0M to 7.5M in 40mM HEPES, pH 7.5, 20mM KCl. Changes in tertiary structure were
detected by fluorescence measurements on a FluoroMax-2 (Spex, Edison, USA) at 25°C. The
excitation wavelength was set to 280nM, while emission spectra were collected from 300nM
to 400nM. The midpoint of the urea transition was obtained from a plot of the fluorescence
signal at 328nM against the urea concentration.
ATPase activity
ATPase activities were measured using the EnzCheck phosphate detection kit (Molecular
Probes, Leiden, Netherlands). The assays were performed in 120µl cuvettes and the
production of phosphate was detected by the increase of adsorbance at 360nm, using a
Pharmacia 40/60 spectrophotometer. The temperature was set to 37°C. Assays were
performed in 40mM HEPES, pH 7.5, 150mM KCl, 5mM MgCl2 and 2mM ATP. Typical
protein concentrations were 2.5µM for Hsp90 and up to 50µM for the less active N-terminal
fragments. To subtract Hsp90 contaminating ATPase activities that usually co-purify with
Hsp90, GA was added at concentrations of 20µM for Hsp90 and 100µM for the N-terminal
fragments. The remaining ATPase activity was Hsp90 independent.
ATPase - Competition experiments
For the ATPase assays Hsp90 concentrations were 2.5µM, while the concentrations of
fragments were varied between 1µM and 16µM. To allow equilibration between homodimers
and heterodimers, the samples were incubated for 10 minutes at 37°C, which was sufficient to
reach equilibrium before ATP was added. Again, GA was added later on to ensure, that the
detected ATPase activity is due to ATP hydrolysis by Hsp90. The data analysis of the titration
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

8
relied on the assumption that the binding of fragments to each other is as effective as the
binding of fragments to Hsp90 and the binding of Hsp90 to itself. To correct for potential
mistakes in the concentration of one of the proteins, a stoichiometry factor n was incorporated
that should, in case that all the concentrations are correct, be close to one. n-values smaller
than one could also indicate that the fragments bind with a lower affinity to Hsp90 than Hsp90
to itself. The resulting equation was
[ ][ ] [ ]
[ ][ ] [ ]Activity Act
Hsp
Hsp n FragAct
Hsp
Hsp n Fraghet= ⋅
+ ⋅+ ⋅ −
+ ⋅
homo
90
901
90
90 (equation 1)
Acthomo is the activity of the homodimer and Acthet is the activity of the heterodimer.
Cross-linking
Cross-linking was performed with glutaraldehyde (Merck, Darmstadt, Germany). 20µl of a
2.4µM Hsp90 solution containing different amounts of Hsp90 - 527C were incubated for 10
minutes at room temperature. 1µl of a 1:10 solution of 25% glutaraldeyde in reaction buffer
(40mM HEPES, pH 7.5, 150mM KCl, 5mM MgCl2) was added for two minutes. 2mM AMP-
PNP was included optional in the reaction buffer. Then the reaction was stopped by adding
5µl of 1M Tris, pH 8.0. After addition of 5µl of 5x Laemmli-buffer, the proteins were
separated on precast 4-12% Polyacrylamide gels (Invitrogen, Groningen, Netherlands) and
stained with Coomassie Blue (Sigma-Aldrich, St.Louis, USA). Evaluation of the stained gels
was performed with the programs LabScan and ImageMaster (Amersham Pharmacia Biotech).
Size -exclusion HPLC and data analysis
Gelfiltration HPLC was performed on a PU-1580 system (Jasco, Groß-Umstadt, Germany)
with a Superdex 200HR - column (Amersham Pharmacia Biotech, Uppsala, Sweden) in
40mM HEPES, pH 7.5, 150mM KCl. The Hsp90 concentrations applied to the column were
varied between 12µM and 9nM. Protein fluorescence was detected using a FP 1520-S
fluorescence detector (Jasco, Victoria, Canada) with excitation at 280nm and emission at
328nm. The gain of the photomultiplier was varied to be able to record the signal over a
concentration range of three magnitudes. No interaction of monomeric and dimeric species
with the column matrix was observed (with the exception of 527C) as the peak shapes were
similar for the dimeric and the monomeric species. The analysis of the dissociation curves was
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

9
based on the assumption that at any time monomeric and dimeric species were in equilibrium.
Data were fitted using the Scientist program (MicroMath, Salt Lake City, USA) and equation
2:
ET ET ET ETotein
otein Kmono mono erd app
= − − ⋅+
( )[Pr ]
[Pr ]dim,
(equation 2)
where ET is the elution time, ETmono the elution time of the monomeric species and ETdimer the
elution time of the dimeric species. The obtained dissociation konstant Kd,app is only an
aproximation, as the protein is diluted severalfold during the course of the experiment. To
determine the dilution factor the peak area was analysed. At the time of injection, the width of
the peak was 0.2 minutes, which corresponds to 0.1ml injected volume. Dividing the peak
area by 0.2 minutes gave a signal, that is about seven times as high as the maximum of the
originally observed peak. This factor was used as the dilution factor for the calculation of the
”real” Kd by dividing Kd,app with the dilution factor. As the dilution factors were found to be
similar for the different Hsp90 fragments they allowed to directly compare their association
behaviours. All values in the following text are already corrected for dilution.
Analytical ultracentrifugation
Data for sedimentation equilibrium runs were collected on a Beckman XL-A analytical
ultracentrifuge using a Ti60 rotor. All runs were performed at 4°C for 48 hours with the
addition of protease inhibitors (complete, Roche Molecular Biochemicals, Mannheim,
Germany) to avoid degradation. The protein gradient was detected at 280nm. Data analysis
was done with the programme Origin (Beckman, Fullerton, USA).
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

10
Results
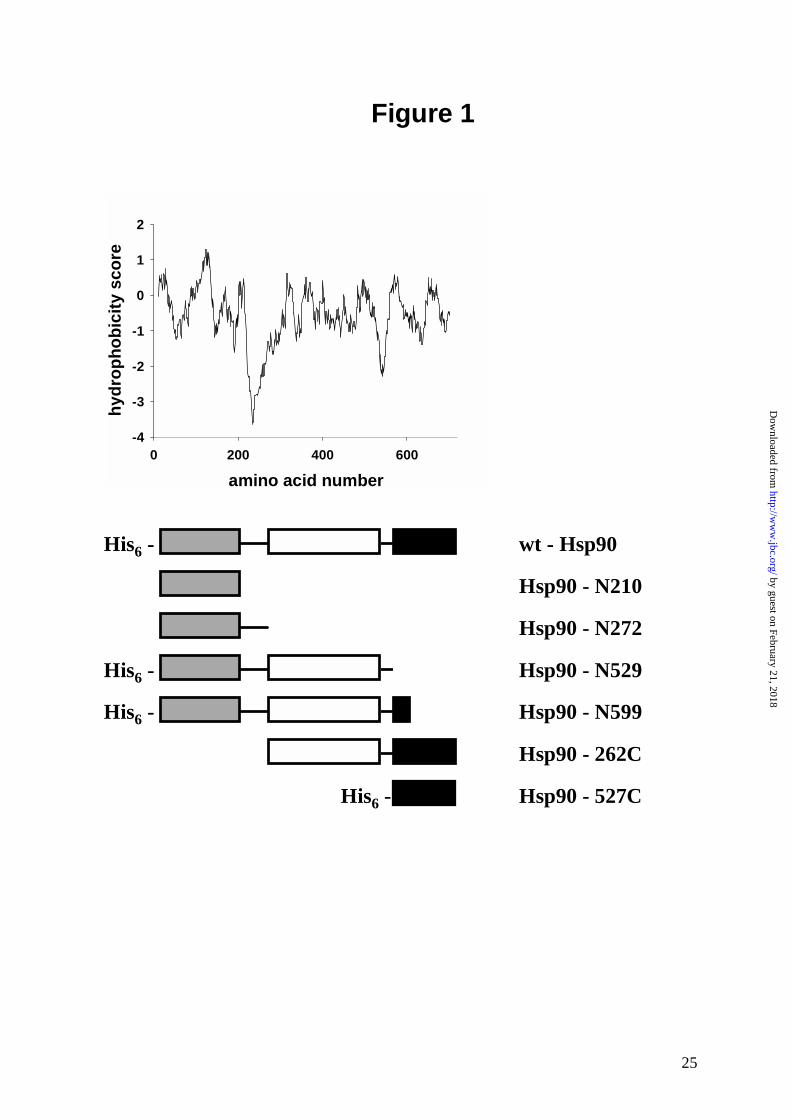
N-terminal fragments of yeast Hsp90 show reduced ATPase activity
Earlier studies showed that truncation mutants of Hsp90 which contain the N-terminal ATP-
binding site, but lack C-terminal regions are considerably less active in ATP-hydrolysis than
the wt-protein (5, 25, 22). We were interested in further defining the requirements for full
ATPase activity of Hsp90. For this purpose, we designed a set of Hsp90 fragments based on
proteolytic digests (5), sequence alignments with homologous proteins and hydropathy plots
(Figure 1). The fragments, varing in size from 210 amino acids (1 - 210) to 599 amino acids (1
- 599), were purified and their structures and stabilities were analysed using CD-spectroscopy
and urea transitions. The midpoint of the unfolding transition were between 3M and 5M urea
for all fragments tested. This compares to the stability of the wt protein. All domains used
were thus shown to be folded and stable (data not shown). As reported previously, Hsp90 -
N210 and Hsp90 - N272 showed extremely weak ATPase activities (0.05µmole ATP per
µmole protein per minute), while Hsp90 - N529 and, more pronounced, Hsp90 - N599
exhibited increased ATPase activities (0.12 and 0.18 µmole ATP per µmole protein per
minute, respectively). However, these activities are still by a factor of six lower than the
ATPase activity of wild-type Hsp90, which was 1.1 µmole ATP per µmole protein per minute
(Figure 2A).
We were unable to detect any concentration dependence of the ATPase activity for Hsp90 -
N210 and for the full-length protein. However, Hsp90 - N529 and Hsp90 - N599 showed a
reproducable increase of activity with protein concentration (Figure 2B).
C-terminal fragments and wt - Hsp90 share similar dimerization properties
To determine the dissociation constant (Kd) for Hsp90 we decided to employ size-exclusion
HPLC (SEC - HPLC) with fluorescence detection at different protein concentrations. Using
Hsp90 concentrations of 2nM to 1µM, only one peak was observed at every concentration
tested. However, the elution times differed depending on the protein concentration used
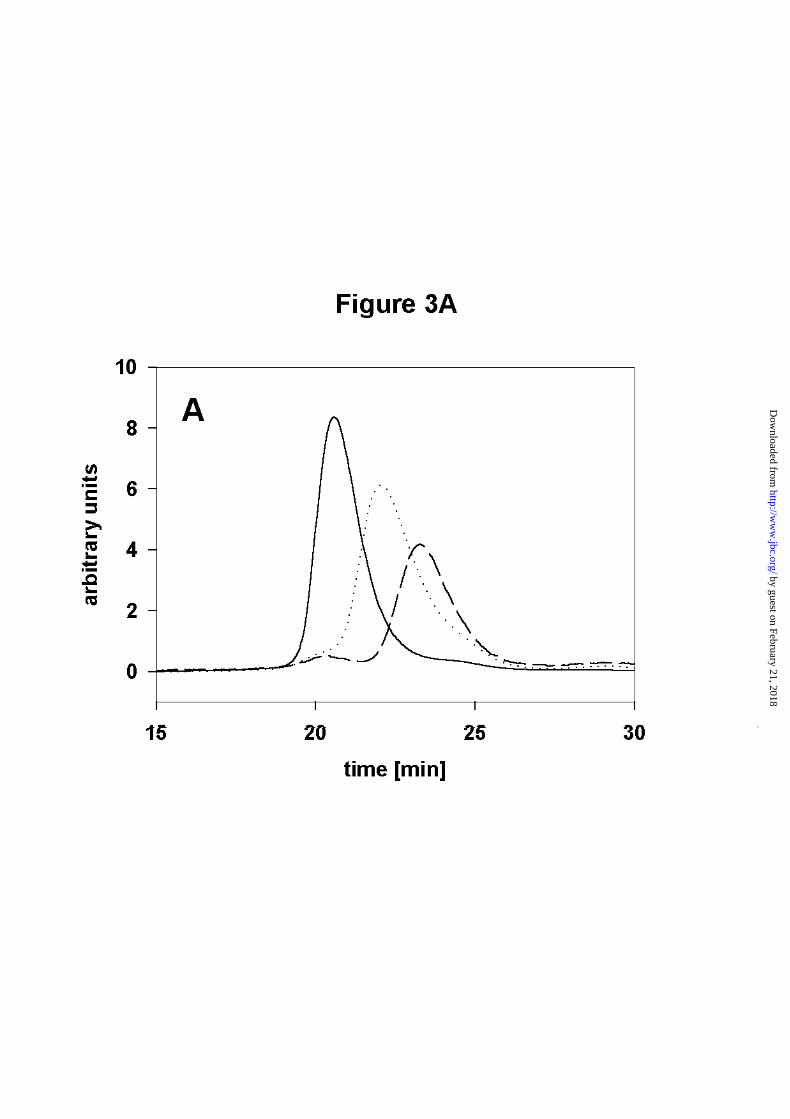
(Figure 3A). A shift could be observed starting from an elution time of 20 minutes (high
protein concentration) to an elution time of 23 minutes (low protein concentration). The
presence of one peak at every concentration is consistent with fast equilibration compared to
the time scale of the experiment. Using a model which assumes a monomer-dimer equilibrium
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

11
at every time point of the experiment, we fitted the data points and obtained a Kd of 60nM +/-
12nM (Figure 3B).
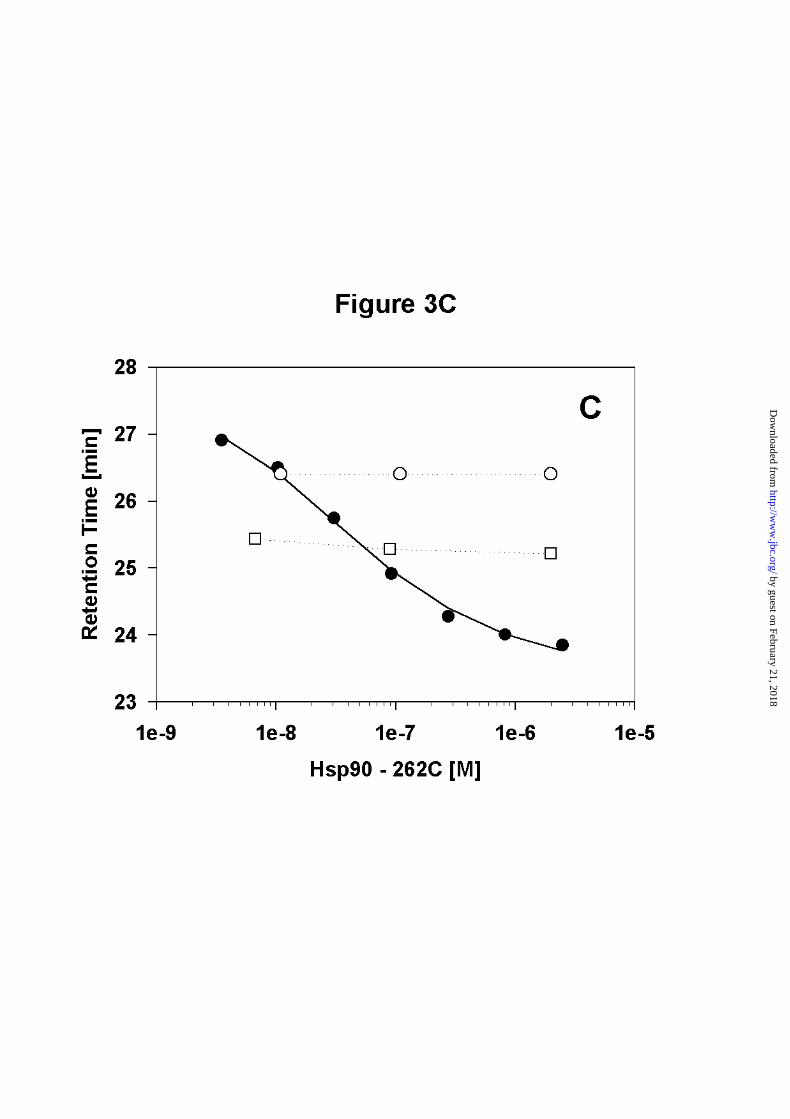
Having calculated the Kd for wt - Hsp90, we were interested to investigate whether C-terminal
fragments containing the dimerization site show similar dimerization characteristics. Two
fragments were constructed, one ranging from amino acids 262 to 708 (Hsp90 - 262C) and the
other containing amino acids 527 to 709 (Hsp90 - 527C). Hsp90 - 262C gave a transition
curve similar to Hsp90 (Figure 3C). The Kd obtained for Hsp90 - 262C was 45nM +/- 12nM.
For Hsp90 - 527C no titration curve was obtained, as this protein interacted with the
gelfiltration matrix, resulting in significant peak broadening.
These data clearly show, that Hsp90 and its C-terminal fragments are dimeric under
conditions used for the ATPase assay, where protein concentrations usually are by at least one
order of magnitude higher than the dimerization constant of Hsp90.
N-terminal Hsp90 fragments are monomeric
To analyse the quaternary structure of N-terminal fragments the same experimental setup was
used. For Hsp90 - N529, no changes in the elution time were observed at concentrations
ranging from 0.04µM to 5µM. This, together with equilibrium sedimentation data from
analytical ultracentrifugation collected for Hsp90 - N529 (data not shown) led us to conclude,
that Hsp90 - N529 is monomeric within the concentration range tested. Similarly SEC - HPLC
experiments were performed with Hsp90 - N599. Here no change in quaternary structure was
observed within the concentration range tested (0.04µM to 2µM) as well (Figure 3C).
Next we investigated, whether the oligomerization properties of the N-terminal fragments
change in the presence of ATP. Hsp90 - N529 as well as Hsp90 - N599 showed a slight
concentration dependence in SEC - HPLC experiments (data not shown). In agreement with
the ATPase assays (Fig. 2B) which were performed at concentrations between 2 and 10µM for
Hsp90 - N599, these data indicate that this could be the beginning of a monomer - dimer
transition curve, but still be far away from the actual dimerization constant.
Taken together, these data suggest that the fragments lacking the C-terminal domain are
monomeric and that ATP does not alter the oligomerization behaviour of these proteins in the
concentration range tested. At higher concentrations, as used in the ATPase assays (Figure
2B), they dimerize with dimerization constants in the range of 10µM for Hsp90 - N599 and
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

12
about 70µM for Hsp90 - N530, as obtained by analysis of the concentration dependent
ATPase assays.
Heterodimers can be formed between Hsp90 and C-terminal Hsp90 fragments
Having established that the dimerization properties for Hsp90 and its C-terminal fragment
262C are similar, we were interested whether heterodimers can be obtained by mixing Hsp90
with C-terminal fragments. To detect dimers we used glutaraldehyde which is known to
efficiently crosslink dimeric Hsp90 (32). Crosslinking is not complete since about 20%
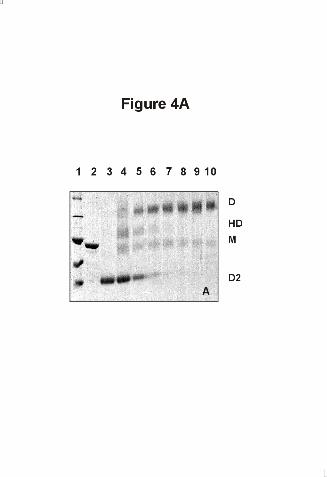
monomers can be detected. (Figure 4). Uncomplete glutaraldehyde - crosslinking of Hsp90
has been observed previously (32). Hsp90 - 527C was added to wt - Hsp90. In the presence of
Hsp90 - 527C, a decrease in the intensity of the dimeric wt - Hsp90 band was visible on SDS-
PAGE, while an additional band appeared, consisting of one molecule Hsp90 - 527C and one
molecule Hsp90 (Figure 4A). Thus, it is possible to form heterodimers between Hsp90 and its
C-terminal fragments in a concentration - dependent manner. Even though the concentration
of the C-terminal fragment was much higher than the concentration of wt - Hsp90 in these
experiments, still the sample seems to contain a fraction of homodimers of wt - Hsp90. A
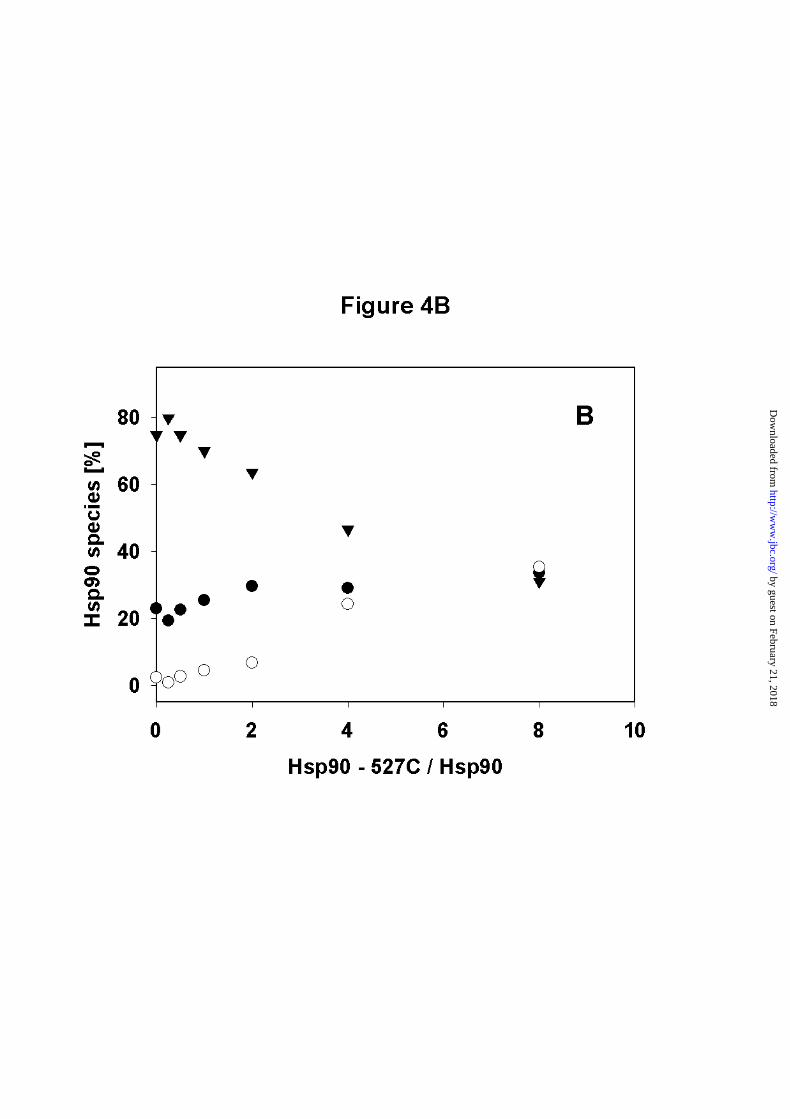
quantitative densitometric analysis of the scanned SDS-gels showed that the heterodimer band
increases with increased concentrations of Hsp90 – 527C. At the highest concentration of
Hsp90 - 527C the cross-linked sample contains about equal concentrations of monomeric
Hsp90, dimeric Hsp90 and heterodimeric Hsp90 (Figure 4B). This ratio does not necessarily
reflect the amount present under equilibrium conditions, especially as it is known from the
dissociation constant, that dimerization of Hsp90 is complete at the concentrations used. Thus,
this result might reflect a higher crosslinking efficiency of the homodimer compared to the
heterodimer and the C-terminal fragment. Crosslinks made in the absence and presence of
AMP-PNP did not show detectable differences in the intensity of the heterodimer band (data
not shown). Thus the N-terminal association induced by AMP-PNP does not contribute
significantly to the ability of 527C - Hsp90 to disrupt Hsp90 homodimers.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

13
Heterodimers of Hsp90 and C-terminal Hsp90 fragments show reduced ATPase activity
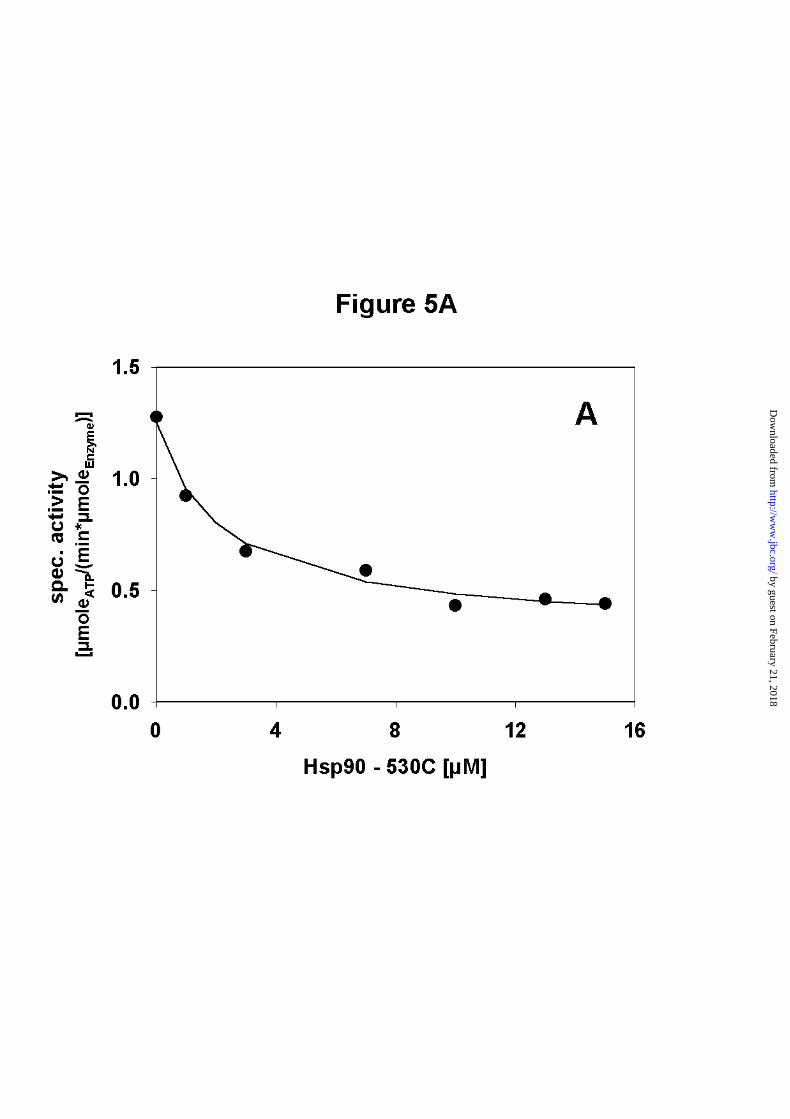
Next, ATPase assays were performed with Hsp90 and Hsp90 - 527C. Hsp90 - 527C itself was
found to have no ATPase activity. When the concentration of Hsp90 - 527C was increased in
the presence of a constant amount of Hsp90, a decrease in ATPase activity was observed (Fig.
5A). This result suggests that the heterodimers have reduced ATPase activity. A statistical
model which assumes equal probabilities for the formation of homodimers and heterodimers
of Hsp90 and Hsp90 - 527C gave an excellent fit for the data points (Figure 5A).
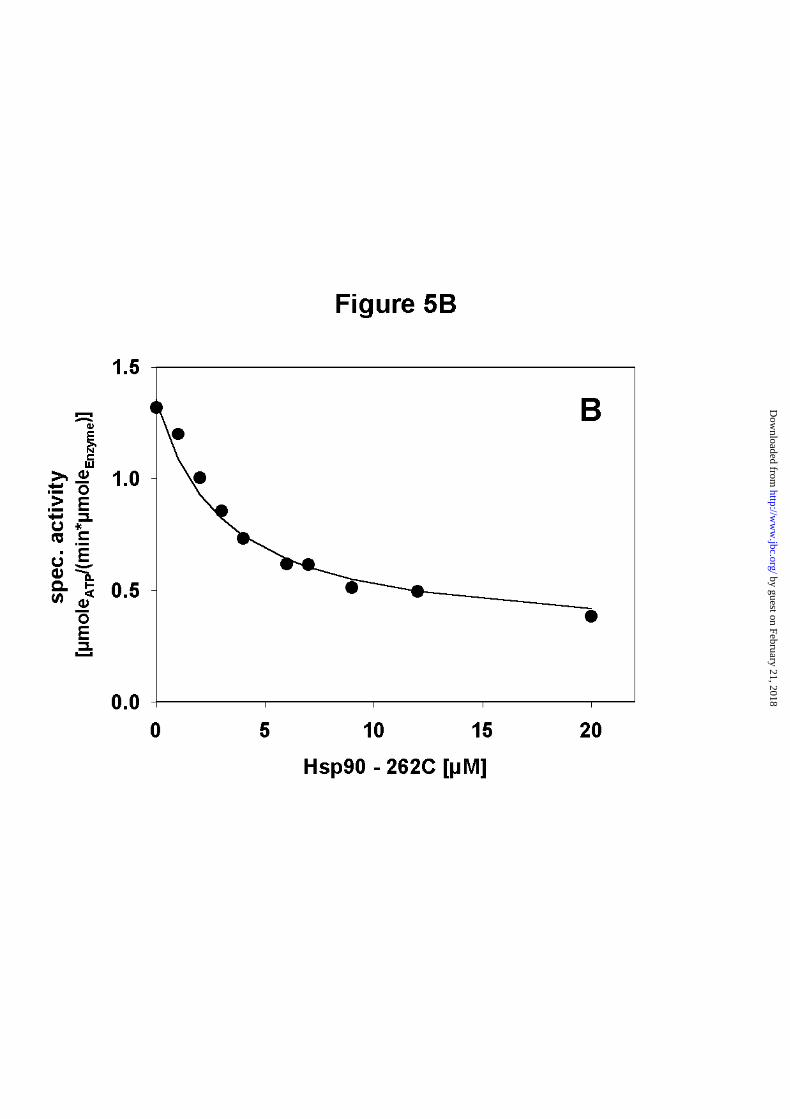
Addition of longer C-terminal fragments containing the middle domain in addition to the C-
terminal domain (amino acid 262 - 709, cf. Fig.1) also led to decreased ATPase activities
(Figure 5B). The inhibition curves were very similar to that of Hsp90 - 527C and again the
statistical model gave a good fit to the data points. The model used to fit the data points also
provides information on the ATPase activity of the heterodimer. The calculated activity of the
heterodimer is one third of the wt activity. This indicates that the ATPase is not completely
inhibited in the heterodimer, at least not the same extend as in the isolated N-terminal
domains. The fact that further increase in the concentrations of N - terminal truncated mutants
did not result in additional decrease of the ATPase activity suggests that the endpoint of the
titration is reached as expected based on the concentrations of the two proteins. This suggests
that beside the N-terminal dimerization, C-terminal regions seem to be important for efficacy
of the ATPase cycle. Also, fast dissociation and reassociation of dimers could result in
formation of homodimers, which might be sufficient to stimulate the ATPase activity and thus
prevent complete inhibition.
Taken together, the data suggest that the presence of two N-terminal domains is required to
activate the ATPase activity in wt Hsp90. Additionally, they confirm that the dimerization
properties of Hsp90 - 262C and Hsp90 - 527C are very similar to that of wild-type Hsp90,
indicating that interactions in the N-terminal part of the protein do not contribute significantly
to the dissociation constant of the wild-type protein.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

14
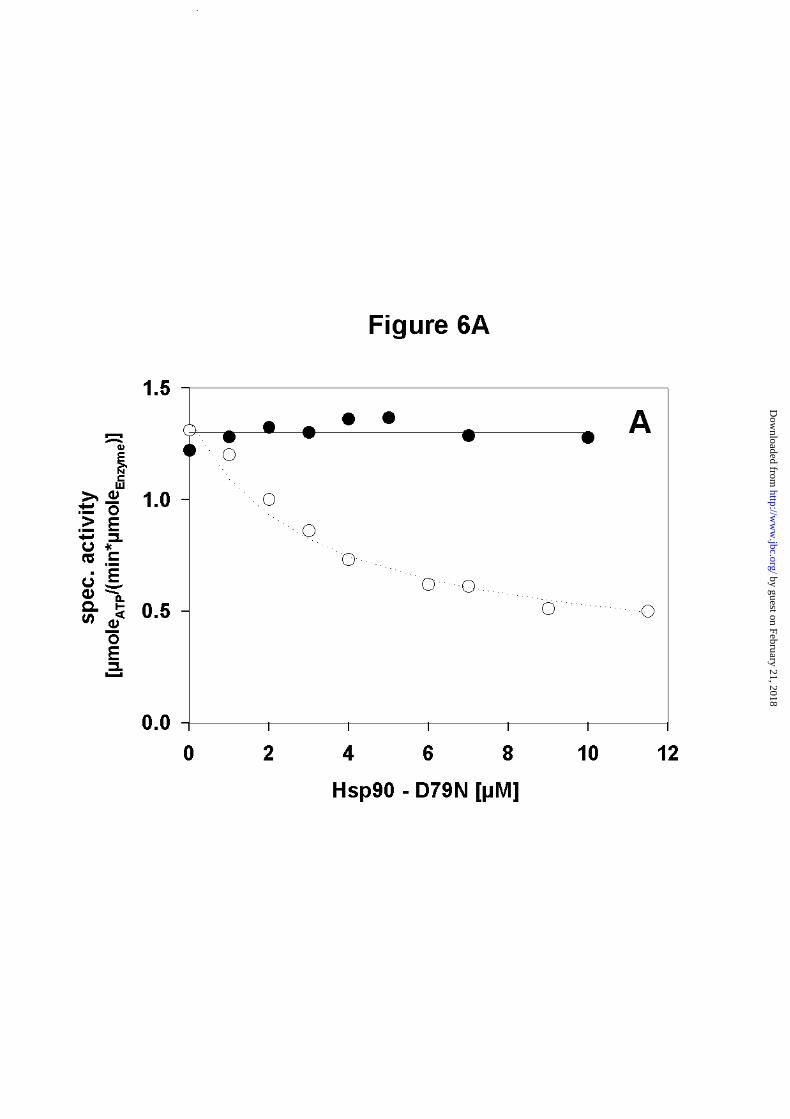
Hsp90 - D79N does not inhibit the ATPase activity of Hsp90
Next we wanted to know, whether ATP binding is required in both N-terminal domains of the
homodimer to achieve wild-type activity. To test this, we used a point mutant of Hsp90,
Hsp90 - D79N, which does not bind nor hydrolyse ATP (12, 13).
Competition experiments performed with increasing concentrations of Hsp90 - D79N and a
fixed amount of Hsp90 showed that Hsp90 - D79N did not affect the ATPase activity of
Hsp90, even if an eightfold excess of the Hsp90 mutant was used (Figure 6A). This implies
that the activity of an individual wt-monomer in the dimer context is the same, independent of
whether the N-terminal partner domain is active or inactive. Thus a defect in ATP binding
does not compromise stimulation of ATP hydrolysis in the partner domain.
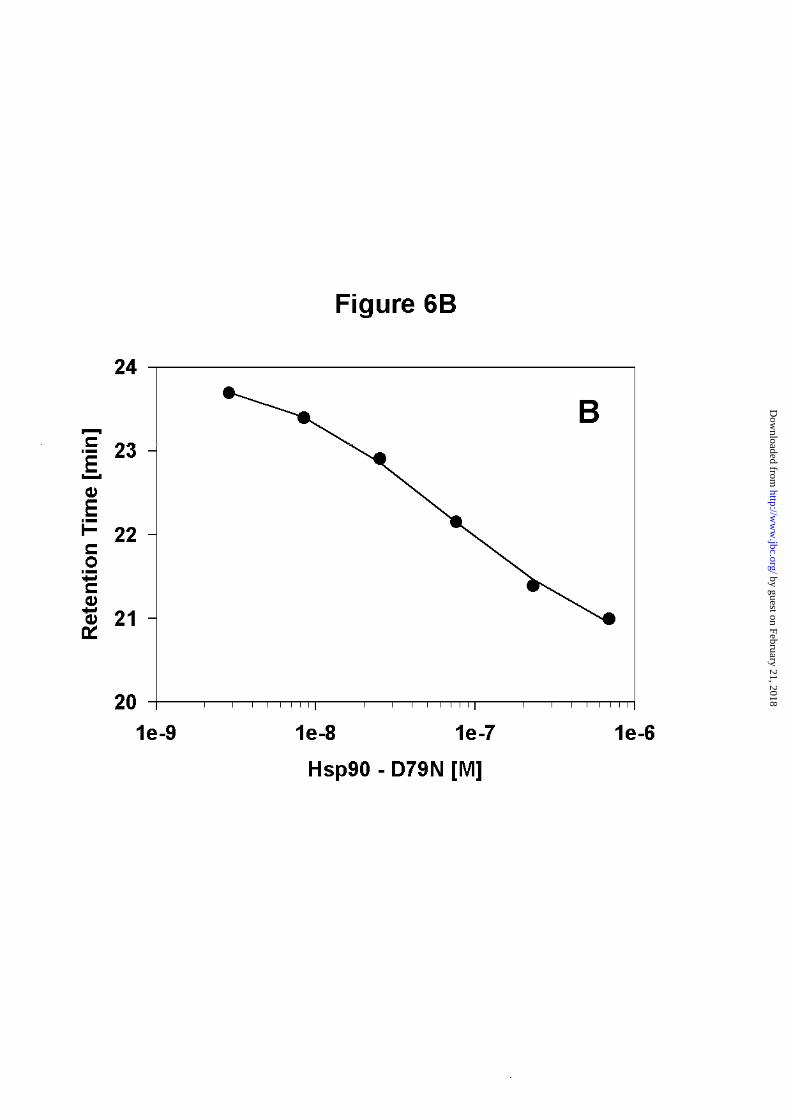
As we could not detect any change in ATPase activity, we wanted to rule out the possibility,
that formation of heterodimers did not occur in the competition experiment. Hsp90 is known
to be very sensitive to C-terminal proteolysis, and the loss of even a small fragment from the
C-terminal end affects the dimerization properties significantly (data not shown). Thus, we
determined the dimerization constant for Hsp90 - D79N by SEC-HPLC. Here we obtained a
dimerization constant of 80nM +/- 20nM (Figure 6B). This value indicates, that the
dimerization of Hsp90 - D79N does not differ significantly from that of wt - Hsp90.
Dimerization of Hsp90 is a dynamic process
We used the inhibiton of the ATPase activity of Hsp90 by formation of heterodimers to obtain
insight into the dynamics of the dimerization reaction. To this end, we added Hsp90 - 262C to
the ATPase reaction while monitoring the progress of ATP hydrolysis. As expected, the
formation of heterodimers resulted in a pronounced decrease in ATP hydrolysis immediately
after mixing (Figure 7A).
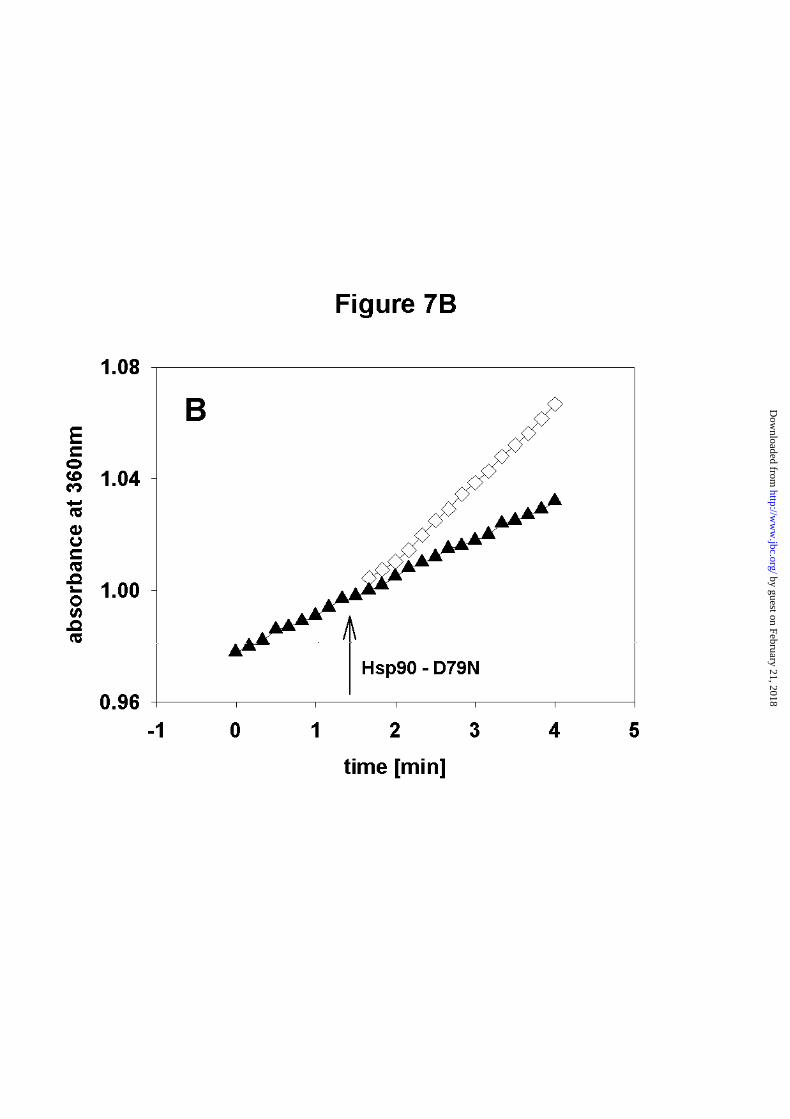
Additionally, we monitored the kinetics of formation of active dimers by adding Hsp90 -
D79N to an ATPase-suppressed heterodimeric form of Hsp90 and Hsp90 - 262C. This
experiment gave the expected increase in activity (Figure 7B), as the addition of Hsp90 -
D79N leads to the formation of Hsp90/Hsp90 - D79N - heterodimers, in which the intact N-
terminal domain exhibits wild type ATPase activity (cf. Figure 6A). Again, the increase in
activity was obtained within the first seconds of the experiment.
These results indicate, that the Hsp90 dimer is a highly dynamic structure.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

15
Discussion
The X-ray structure of an N-terminal fragment of Hsp90 in complex with ADP showed a
nucleotide binding site which buries the ribose backbone and the adenine base inside a cleft
but leaves the β-phosphate pointing towards the surface of the molecule (19). It is obvious
from this structure, that for ATP the γ-phosphate would be completely solvent-exposed. More
importantly, no conformational changes were observed in the crystal structure of the N-
terminal domain upon binding of ADP or GA (19, 20). Subsequent studies showed that
conformational changes occur in other parts of the protein, which trap the ATP molecule and
commit it to hydrolysis (22). In addition, the N-terminal domains come in close contact (25).
The functional consequences of these changes are still largely unclear. One of the few
functional correlations of these rearrangements is that binding of the cofactor p23 requires
ATP bound to Hsp90 (28) and the dimeric form of the N-terminal domains (22, 29).
In this study, the Hsp90 dimer was shown to exhibit a dissociation constant of about 60nM.
The major dimerization site resides in the C-terminal region, as an Hsp90 fragment consisting
of amino acids 527 - 709 formed heterodimers with a affinity comparable to the native dimer.
This is in agreement with previous qualitative studies, using truncated fragments or deletion
mutants of Hsp90. For human Hsp90, Jibard et al. (31) mapped sites important for
dimerization to amino-acids 548-567, 661-677 and 679-728. These sites can be found in
fragment 527C of yeast Hsp90 indicating that this domain is responsible for dimerization. In
addition to this, our results suggest that much weaker (Kd > 70µM) interaction sites are
present within amino acid 1 - 529.
Our approach to obtain the dimerization constant of Hsp90 differs from other methods, in that
it does not require labelling. The quantitative analysis of gelfiltration allows a wide
concentration range to be covered using sensitive fluorescence detection. Previously
dimerization constants for homodimers were obtained for enzymes like lactate
dehydrogenases using enzymatic activity, as the probe for the native tetrameric state (33, 34).
The value obtained is 1nM. A dissociation constant of 60nM for Hsp90 may reflect the
requirement for a dynamic monomer/dimer equilibrium. Most interestingly, the dissociation
constant for the dimer of CheA, a histidine kinase with an ATP-binding site homologous to
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

16
Hsp90 was determined by enzymatic assays and found to be in the range of 200 to 400nM
(35). Dissociation and association in the Hsp90 dimer occurs fast, as we were unable to
measure the rates of subunit-exchange in experiments with a deadtime of about 20 seconds.
This shows, that the dynamic of the dimerization of Hsp90 is at least in the same range as
ATP hydrolysis, leading to the interesting possibility, that Hsp90 might be able to dissociate
during the ATPase cycle.
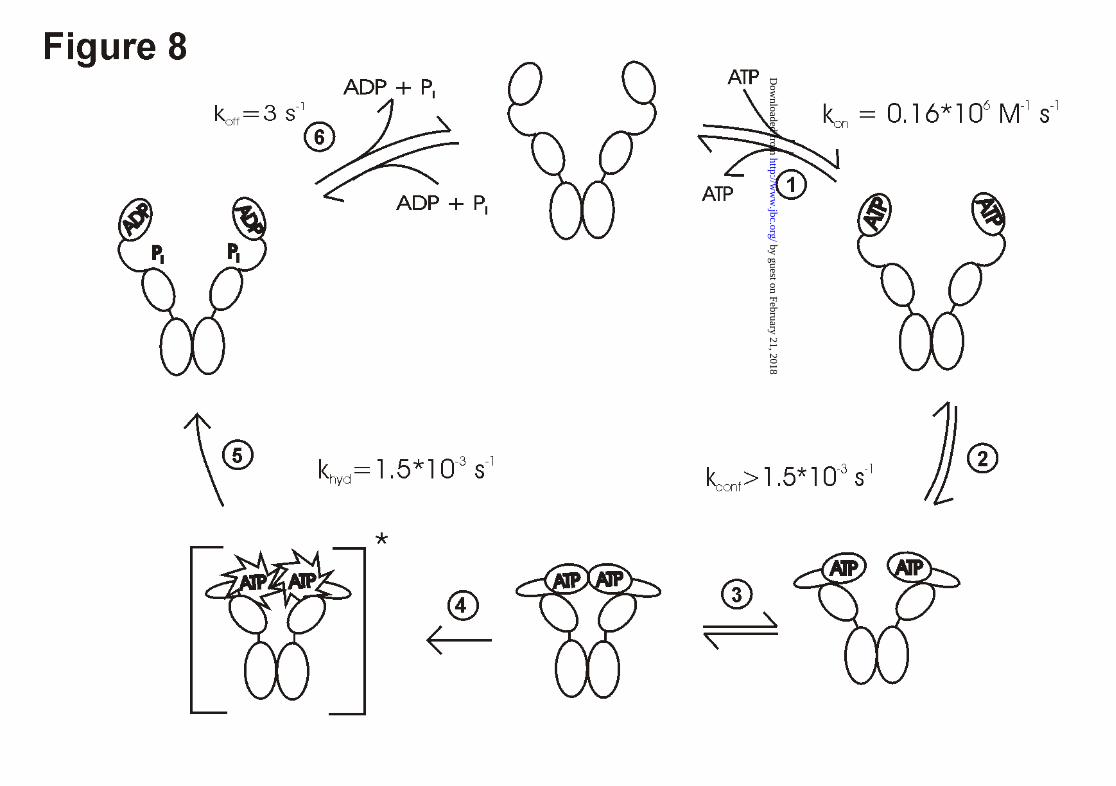
The data of this study, combined with earlier data (22, 25) allow us to define the key steps of
the Hsp90 ATPase as summarized below (Fig. 8).
In the first step of our model, ATP binding occurs independently at both N-terminal domains
of the dimer in a fast reaction (Fig. 8, step 1). Cooperativity does not seem to be involved in
the binding reaction, since the binding constant for AMP-PNP to wt - Hsp90 is identical to
that of Hsp90 - N529 or Hsp90 - N210 (data not shown).
Following the binding of ATP, a conformational change occurs in Hsp90 (Fig. 8, step 2),
which requires amino-acids 1 - 451 and was suggested to use Lys-342 as the acceptor for the
γ-phosphate of the ATP (22). These movements seem to be a prerequisite for a functional N-
terminal dimerization reaction (25), that we consider to be the next step (Fig. 8, step 3).
This dimerization reaction was shown to be possible for Hsp90 - N529, but much more
efficient for Hsp90 - N599, indicating that major interaction sites exist between amino acids
529 and 599. Our results show that the N-terminal dimerization is the prerequisite for an
efficient ATP hydrolysis reaction. Formation of heterodimers with fragments that compete for
the dimerization site on the wt-protein leads to a significant inhibition of the Hsp90 ATPase
activity. A heterodimer comprising one wild - type protein and one fragment lacking the first
261 amino acids shows reduced ATPase - activity suggesting that the two N - terminal
fragments hydrolyse their ATP-molecules in a cooperative manner (Fig. 8, step 4 and 5). This
is reminescent of the ATP-dependent chaperone GroEL, which is known to hydrolyse its
ATP-molecules in a highly regulated way (36, 37) and where the hydrolysis of ATP is
preceeded by conformational changes, that trap the ATP molecule and commit it to hydrolysis
(36).
In the case of Hsp90, surprisingly, addition of Hsp90 - D79N did not change the activity of the
wild-type protein, indicating that the wt - monomer in complex with Hsp90 - D79N is as
active as the wt - monomer in the homodimer. This implies that Hsp90 - D79N fulfills all the
requirements needed to activate the ATPase activity of wt - protein, although its part is only a
passive one. Thus, in principle, Hsp90 is able to work even with only one ATP bound to the
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

17
dimer, although this case might be unphysiological given the high ATP concentrations inside
the cell and the fact that ATP binding is known to be very fast compared to the following
steps in the hydrolysis cycle (22). After hydrolysis, Hsp90 presumably opens and releases the
ADP molecule (Fig. 8, step 6), which brings Hsp90 back to the conformation competent to
bind ATP.
Additional evidence for this model comes from studies of homologous proteins, known to
share a structurally related ATP-binding site with Hsp90. N-terminal fragments of gyrase B
show a concentration-dependent ATPase-activity, that closely resembles that observed for
Hsp90. Most striking is the crystal structure of an N-terminal fragment of gyrase B (38). Here,
the structural organization seems to be very similar to that of Hsp90. The model proposed for
the ATPase mechanism of gyrase involves the transient association of the N-terminal domains
in their ATP-bound state (39). This may lead to changes in the active center of the enzyme,
which are the basis for the cooperative hydrolysis of ATP (40). A cooperative mechanism of
this kind would guarantee, that ATP hydrolysis is closely coupled to the coordinated
movement of the two N-terminal domains. It might be envisioned, that Hsp90 this way
couples the energy of ATP hydrolysis to the coordinated movement of domains.
It remains to be seen, how these movements are influenced by the partner proteins of Hsp90
and how they affect the conformational processing of client proteins.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

18
Literature
1. Wiech, H., Buchner, J., Zimmermann, R. & Jakob, U. (1992) Nature 358, 169-170.
2. Miyata, Y. & Yahara, I. (1992) J. Biol. Chem. 267, 7042-7047.
3. Freeman, B. C. & Morimoto, R. I. (1996) EMBO J. 15, 2969-2979.
4. Jakob, U., Lilie, H., Meyer, I. & Buchner, J. (1995) J. Biol. Chem. 270, 7288-7294.
5. Scheibel, T., Weikl, T. & Buchner, J. (1998), Proc. Nat. Acad. Sci. U.S.A. 95, 1495-
1499.
6. Pratt, W. B. (1998) Proc. Soc. Exp. Biol. Med. 217, 420-434.
7. Buchner, J. (1999) Trends. Biochem. Sci. 24, 136-141.
8. Richter, K. & Buchner, J. (2001) J. Cell. Physiol., in press.
9. Hu, J. & Seeger, C. (1996) Proc. Natl. Acad. Sci. U.S.A. 93, 1060-1064.
10. Holt, S. E., Aisner, D. L., Baur, J., Tesmer, V. M., Dy, M., Ouellette, M., Trager, J. B.,
Morin, G. B., Toft, D. O., Shay, J. W., Wright, W. E. & White, M. A. (1999) Genes
Dev. 13, 817-826.
11. Nathan, D. F. & Lindquist, S. (1995) Mol. Cell. Biol. 15, 3917-3925.
12. Obermann, W. M. J., Sondermann, H., Russo, A. A., Pavlevitch, N. P. & Hartl, F. U.
(1998) J. Cell. Biol. 143, 901-910.
13. Panaretou, B., Prodromou, C., Roe, S. M., O’Brien, R., Ladbury, J. E., Piper, P. W. &
Pearl, L. H. (1998) EMBO J. 17, 4829-4836.
14. Whitesell, L., Mimnaugh, E. G., DeCosta, B., Myers, C. E. & Neckers, L. M. (1994)
Proc. Natl. Acad. Sci. U.S.A. 91, 8324-8328.
15. An, W. G., Schnur, R. C., Neckers, L. & Blgosklonny, M. V. (1997) Cancer Chemother.
Pharmacol. 40, 60-64.
16. Csermely, P., Kajtar, J., Hollosi, M., Jalsovszky, G., Holly, S., Kahn, C. R., Gergely, P.,
Soti, C., Mihaly, K. & Somogyi, J. (1993) J Biol Chem. 268, 1901-1907.
17. Grenert, J. P., Sullivan, W. P., Fadden, P, Haystead, T. A., Clark, J., Mimnaugh, E.,
Krutzsch, H., Ochel, H. J., Schulte, T. W., Sausville, E., Neckers, L. M. & Toft, D.O.
(1997) J. Biol. Chem. 272, 23832-23850.
18. Scheibel, T., Neuhofen, S., Weikl, T., Mayr, C., Reinstein, J., Vogel, P. D. & Buchner,
J. (1997) J. Biol. Chem. 272, 18608-18613.
19. Prodromou, C., Roe, S. M., O’Brien, R., Ladbury, J. E., Piper, P. W. & Pearl, L. H.
(1997) Cell 90, 65-75.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

19
20. Stebbins, C. E., Russo, A. A., Schneider, C., Rosen, N., Hartl, F. U. & Pavletich, N. P.
(1997) Cell 89, 239-250.
21. Dutta, R. & Inouye, M. (2000) Trends. Biochem. Sci. 25, 24-28.
22. Weikl, T., Muschler, P., Richter, K., Veit, T., Reinstein, J. & Buchner, J. (2000) J. Mol.
Biol. 303, 583-592.
23. Ban, C. & Yang, W. (1998) Cell 95, 541-552.
24. Ban, C., Junop, M. & Yang, W. (1999) Cell 97, 85-97.
25. Prodromou, C., Panaretou, B., Chohan, S., Siligardi, G., O’Brien, R., Ladbury, J. E.,
Roe, S. M., Piper, P. W. & Pearl, L. H. (2000) EMBO J. 16, 4383-4392.
26. Maruya M., Sameshima M., Nemoto T. & Yahara I. (1999) J. Mol. Biol. 285, 903-907.
27. Johnson, J. L. & Toft, D. O. (1994) J. Biol. Chem. 269, 24989-24993.
28. Johnson, J. L., Corbisier, R., Stensgard, B. & Toft, D. O. (1996) J. Steroid. Biochem.
Mol. Biol. 56, 31-37.
29. Chadli, A., Bouhouche, I., Sullivan, W., Stensgard, B., McMahon, N., Catelli, M. G. &
Toft, D. O. (2000) Proc. Natl. Acad. Sci. USA 97, 12524-12529.
30. Nemoto, T., Ohara-Nemoto, Y., Ota, M., Takagi, T. & Yokoyama, K. (1995) Eur. J.
Biochem. 233, 1-8.
31. Jibard, N., Meng, X., Leclerc, P., Rajkowski, K., Fortin, D., Schweizer-Groyer, G.,
Catelli, M. G., Baulieu, E. E. & Cadepond, F. (1999) Exp. Cell. Res. 247, 461-474.
32. Jakob, U., Meyer, I., Bügl, H., Andre, S., Bardwell, J. C. & Buchner, J. (1995) J. Biol.
Chem. 270, 14412-14419.
33. Bartholmes, P., Durchschlag, H. & Jaenicke, R. (1973) Eur. J. Biochem. 39, 101-108.
34. Berr, K., Wassenberg, D., Lilie, H., Behlke, J. & Jaenicke, R. (2000) Eur. J. Biochem.
267, 5413-5420.
35. Surette, M. G., Levit, M., Liu, Y., Lukat, G., Ninfa, E. G., Ninfa, A. & Stock, J. B.
(1996) J. Biol. Chem. 271, 939-945.
36. Todd, M. J., Viitanen, P. V. & Lorimer, G. H. (1994) Science 265, 659-666.
37. Weissman, J. S., Rye, H. S., Fenton, W. A., Beechem, J. M. & Horwich, A. L. (1996)
Cell 84, 481-490.
38. Wigley, D. B., Davies, G. J., Dodson, E. J., Maxwell, A. & Dodson, G. (1991) Nature
351, 624-629.
39. Kampranis, S. C. & Maxwell, A. (1998) J. Biol. Chem. 273, 26305-26309.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

20
40. Brino, L., Urzhumtsev, A., Mousli, M., Bronner, C., Mitschler, A., Oudet, P. & Moras,
D. (2000) J. Biol. Chem. 275, 9468-9475.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

21
Acknowledgements
The authors would like to thank Dr. Stefan Walter for stimulating discussions and Alex Frenzl
and Martin Haslbeck for help with the artwork. J.B. was supported by grants from the
Deutsche Forschungs Gemeinschaft and the Fonds der chemische Industrie.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

22
Figure Legends
Figure 1.
Fragments of Hsp90 used in this study. The ATP binding site is located in the N - terminal
domain (■ ), while the dimerization site is in the C - terminal domain (■ ). The middle part of
Hsp90 (❑ ) may consist of several domains. The hydropathyblot was calculated using the
Expasy-Tool ProtScale software.
Figure 2. ATPase activity of Hsp90 fragments
ATPase assays were performed at 37°C in a buffer containing 40mM HEPES, 150mM KCl,
5mM MgCl2, 2mM ATP, pH 7.5.
A) ATPase activity of N-terminal Hsp90 fragments. All values were corrected for background
activity using geldanamycin. Protein concentrations were 20µM for Hsp90 – N210 and Hsp90
– N272, 5µM for Hsp90 – N530 and 3µM for Hsp90 – N599 and wt-Hsp90.
B) The concentration dependence of the ATPase activity of N-terminal Hsp90 fragments. (▼)
Hsp90 - N599, (❍ ) Hsp90 - N529, (● ) Hsp90 - N210. Lines represent the fits of the data to the
estimated end point of the wt - Hsp90 activity of 1.2µM phosphate / min*µM Hsp90
Figure 3. Analysis of Hsp90 dimerization by SEC - HPLC
A) Size-exclusion HPLC of Hsp90. The running buffer contained 40mM HEPES, 150mM
KCl, pH 7.5 at 20°C. Protein was detected by fluorescence with an excitation wavelength of
280nm and an emission wavelength of 328nm. The shift of the peaks from 20min to 24min
elution time represents the dissociation of Hsp90. ( ____ ) 1µM Hsp90, (.......) 0.03µM Hsp90,
(------) 5nM Hsp90.
B) Dissociation curve of the Hsp90 dimer. The data points of the SEC analysis were fitted
according to equation 1 (see materials and methods section). The resulting dissociation
constant is 60nM +/- 12nM.
C) Dissociation curve of the C-terminal fragment Hsp90 - 262C (● ) and Hsp90 - N529 (❍ )
and Hsp90 – N599 (❑ ) The dissociation constant for Hsp90 - 262C is 45nM +/- 12nM. The
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

23
elution time of Hsp90 - N529, which is suggested to be monomeric does not change over a
wide range of concentrations, as does the elution time of Hsp90 – N599.
Figure 4. Demonstration of heterodimer formation between Hsp90 and Hsp90 - 527C by
crosslinks
A) SDS-PAGE of crosslinked Hsp90 complexes in a buffer containing 40mM HEPES, pH
7.5, 150mM KCl, 5mM MgCl2 and 2mM AMP-PNP. Hsp90 - 527C was added at a
stoichiometry of 8:1, 4:1, 2:1, 1:1, 1:2 and 1:4 (lanes 4-9) to Hsp90. Crosslinking was
performed for 2 minutes at 30°C with glutaraldehyde. Lane 1: high molecular weight standard,
lane 2: Hsp90 without cross-linking, lane 3 and lane 10: crosslinking of Hsp90 - 527C and wt
- Hsp90 respectively. D: Dimer, M: Monomer, HD: Heterodimer, D2: Dimer of Hsp90 -
527C.
B) Densitometric analysis of the SDS-PAGE. (▼) wt - Hsp90 dimer, (● ) wt - Hsp90
monomer, (❍ ) heterodimer of wt - Hsp90 and Hsp90 - 527C.
Figure 5. Influence of heterodimer formation on the ATPase activity of Hsp90
To form heterodimers during the ATPase assay the concentration of the C - terminal fragment
was varied between 1µM and 20µM and the wt - Hsp90 concentration was kept constant at
2.5µM. ATPase assays were performed as described in Material and Methods.
A) Addition of Hsp90 - 527C to Hsp90. The decrease can be fitted to a statistical model. The
n-value was obtained as 0.8, suggesting similar association constants for the homodimer and
the heterodimer.
B) Addition of Hsp90 - 262C to Hsp90. The specific activity of 2.5µM wt-Hsp90 decreases
with increasing concentrations of Hsp90 - 262C. The n-value obtained from fitting the data
points to a statistical model was 0.7.
Figure 6. Influence of an inactive N - terminal domain on the ATPase activity of Hsp90
heterodimers
Hsp90 - D79N was added with increasing concentrations to 2.5µM wt - Hsp90. ATPase
assays was performed as described in Material and Methods.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

24
A) The specific activity of wt - Hsp90 is not influenced by the presence of increasing amounts
of Hsp90 - D79N, which is unable to bind ATP (● ). These values are compared to the
inhibition obtained with Hsp90 - 262C (❍ ) (cf. Fig. 5B).
B) The dissociation curve of Hsp90 - D79N shows its ability to form dimers. The experiment
was performed as described in Figure 3. The dissociation constant is 85nM +/- 20nM.
Figure 7. Kinetics of subunit exchange in the Hsp90 dimer
A) ATPase inhibition of Hsp90 by heterodimer formation with Hsp90 - 262C. The ATPase
activity of Hsp90 was analysed as described. Starting with Hsp90 (❍ ), Hsp90 - 262C was
added to the ATPase assay after 2 minutes (see arrow) (▲). After roughly 20 seconds of
mixing, the ATPase assay was continued. The ATPase activity levelled off after about 30
seconds, indicating that subunit exchange occurs very efficient.
B) Restoration of ATPase activity by adding Hsp90 - D79N to a solution containing
Hsp90/Hsp90 - 262C heterodimers. The specific activity increases after addition of Hsp90 -
D79N (see arrow). Again subunit exchange is fast compared to the deadtime of the
measurement. (▲) ATPase assay without addition of Hsp90 - D79N, (❑ ) ATPase assay after
addition of Hsp90 - D79N.
Figure 8. Model of the ATPase cycle of Hsp90
The kinetic values are from Weikl et al. (22). These measurements were performed at 25°C
under buffer conditions similar to the ATPase - assays in this study. The model does not
reflect data from Maruya et al., (26), that indicate an antiparallel organization of the Hsp90
dimer in its ATP free state. The N-terminal interaction, as demonstrated in this study,
stimulates the ATP hydrolysis reaction in both domains, leading to the activated species
painted in brackets. After ATP hydrolysis, the phosphate and the ADP are likely to be released
after opening, as the ATP had been trapped inside the protein before hydrolysis occured.
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from
25
Figure 1
amino acid number
0 200 400 600
hy
dro
ph
ob
icit
y s
co
re
-4
-3
-2
-1
0
1
2
His6 -
His6 -
His6 -
His6 -
wt - Hsp90
Hsp90 - N210
Hsp90 - N272
Hsp90 - N529
Hsp90 - N599
Hsp90 - 262C
Hsp90 - 527C
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from

Klaus Richter, Paul Muschler, Otmar Hainzl and Johannes BuchnerCoordinated ATP hydrolysis by the Hsp90 dimer
published online July 5, 2001J. Biol. Chem.
10.1074/jbc.M103832200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on February 21, 2018http://w
ww
.jbc.org/D
ownloaded from