Embed Size (px)
Citation preview

Polar Biol (2009) 32:529–538
DOI 10.1007/s00300-008-0547-4ORIGINAL PAPER
InXuence of weather on reproductive success of northern fulmars in the Canadian high Arctic
Mark L. Mallory · Anthony J. Gaston · Mark R. Forbes · H. Grant Gilchrist
Received: 18 June 2008 / Revised: 16 October 2008 / Accepted: 28 October 2008 / Published online: 14 November 2008© Springer-Verlag 2008
Abstract The northern fulmar (Fulmarus glacialis) is acommon seabird of the North Atlantic Ocean, with breed-ing colonies broadly dispersed between 45°N and 80°N. Athigher latitudes, breeding fulmars experience extensive sea-ice and presumably snow and low temperatures which donot aVect fulmars in the southern part of the breeding range.We studied the relationship between weather and reproduc-tive success of northern fulmars breeding at two colonies inthe Canadian high Arctic. Collectively, hatching success,Xedging success, and productivity (chicks Xedged per egglaid) were similar between our study and results from colo-nies located south of the Arctic. However, a larger propor-tion of fulmars at apparently occupied sites (AOS) in highArctic colonies appeared to forego egg-laying, resulting inlower proportions of chicks Xedged per AOS. Extremeinclement weather was the major factor inXuencing nestingsuccess, resulting in pulses of egg or chick loss during orimmediately following major storms, although the mecha-nism of eVects appeared to diVer between the two colonies.For Arctic fulmars, the risks of nest failure due to stochas-tic, deleterious weather events may be oVset by the predict-able abundance of food supplies during chick-rearing inArctic waters.
Keywords Arctic · Breeding · Fulmarus glacialis · Northern fulmar · Weather
Introduction
Many bird species exhibit phenotypic plasticity in life his-tory characteristics across their breeding range, presumablyin response to diVering physical or biological selectivepressures experienced over the gradient of environmentalconditions (Welty and Baptista 1988). For example, avianbreeding phenology, clutch size, egg hatchability, andreproductive success may vary signiWcantly with increasingaltitude or latitude (e.g. Koenig 1982; Fortescue 1999;Dunn et al. 2000). Thus, studies of the reproductive ecol-ogy of a species from one part of its range may not accu-rately reXect its ecology elsewhere, which may be the casefor the northern fulmar (Fulmarus glacialis; Mallory andForbes 2007).
The fulmar is a petrel with a circumpolar distribution,breeding across 35° of latitude (to 80°N; Hatch andNettleship 1998). It is a generalist predator and scavenger,feeding on marine invertebrates, small Wsh, carrion, andindustrial Wsheries discards (if available; Fisher 1952; Mallory2006). This long-lived seabird lays a single egg which isnot replaced if lost, and it exhibits synchronous breedingwithin colonies, but nesting dates are delayed at higher lati-tudes (Mallory and Forbes 2007). Most northern fulmarsinhabit the Low Arctic and Boreal oceanographic zonesyear round (Salomonsen 1965; Hatch and Nettleship 1998),but birds from high Arctic colonies only overwinter in thisregion.
Among seabirds, reproductive success is strongly linkedto annual productivity of the marine food web (Furness andCamphuysen 1997). This has been shown for Boreal
M. L. Mallory (&)Canadian Wildlife Service, Box 1714, Iqaluit, NU X0A 0H0, Canadae-mail: [email protected]
M. L. Mallory · M. R. ForbesDepartment of Biology, Carleton University, Ottawa, ON K1S 5B6, Canada
A. J. Gaston · H. G. GilchristNational Wildlife Research Centre, Carleton University, Raven Road, Ottawa, ON K1A 0H3, Canada
123

530 Polar Biol (2009) 32:529–538
fulmars (Ollason and Dunnet 1988; Gray et al. 2003), andmay be linked to long-term climatic and oceanographiccycles (Thompson and Ollason 2001). However, for highArctic fulmars, breeding phenology and success also maybe aVected by additional physical aspects of the marineenvironment, and speciWcally sea-ice cover. In this region,more extensive sea-ice, or ice that is delayed in breakup,constitutes a barrier to fulmar foraging, and also delays pro-duction in the marine food web (Welch et al. 1992). Inyears with late sea-ice, fulmar reproductive success isreduced (Gaston et al. 2005).
Given that high Arctic fulmars experience some addi-tional constraints on breeding compared to Boreal fulmars,we were interested in determining if the typical reproduc-tive success of high Arctic birds diVered from that of con-speciWcs nesting in the Boreal zone. Falk and Møller (1997)provided some information on fulmars breeding in one sea-son in northeast Greenland, and some summary data fornorthern Canada are found in Hatch and Nettleship (1998).There are no detailed, published accounts of the reproduc-tive ecology of this species across sites and years in thehigh Arctic, and particularly there are no accounts of howfulmars are aVected by the more extreme weather eventstypical of this region. An assessment of Arctic fulmar ecol-ogy is timely, as sea-ice and weather patterns are changingrapidly in the Arctic (ACIA 2005).
We studied the breeding ecology of northern fulmars attwo colonies in the Canadian high Arctic. Fulmars breedingat these latitudes exhibit a compressed breeding schedule(Mallory and Forbes 2007) and experience subzero temper-atures and potential snowfall throughout the nesting period.They cannot exploit industrial Wsheries during the breedingseason, making them more reliant on natural ecosystemprocesses. Collectively, these diVerences may createstresses not faced by fulmars breeding farther south. Conse-quently, we predicted that mean annual reproductive suc-cess of fulmars in the Canadian high Arctic should be lowercompared to fulmars nesting in the Low Arctic and Borealoceanographic zones. Moreover, while some fulmar nestsfail at all breeding colonies for a variety of reasons (e.g.,predation, abandonment by one member of pair), weexpected that nest failures of Arctic fulmars would be asso-ciated with particularly severe weather events.
Methods
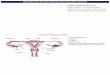
Research was conducted at two northern fulmar colonies inthe Canadian high Arctic (Fig. 1). At the Prince LeopoldIsland colony (74°N, 90°W), Weld crews observed fulmarbreeding ecology between 16 June–21 August 2001, 1June–25 August 2002, 31 May–21 August 2003, and 14June–6 August 2005 (Gaston et al. 2005). As a conse-
quence, in 2001 and 2005 observations at Prince LeopoldIsland were initiated after fulmars began laying eggs.Studies were also undertaken at the Cape Vera colony(76°15�N, 89°15�W), northern Devon Island, between 14May to 9 August 2004, and 20 April to 10 August 2005(i.e., prior to egg-laying). These marine areas are located inthe high Arctic oceanographic zone (Salomonsen 1965).
At the Cape Vera colony, nearby Jones Sound remainsice-covered from October through July of the followingyear, although there is open water immediately beside thecolony in the Hell Gate–Cardigan Strait polynya (Malloryand Forbes 2007). Prince Leopold Island is found at thejunction of Parry Channel (Lancaster Sound and BarrowStrait) and Prince Regent Inlet. Ice covers this region fromOctober through at least April of the following year, butthe extent of ice cover may vary substantially each year(Gaston et al. 2005). Both colonies are situated on extensive,sedimentary cliVs rising to 300 m or more from sea level.The Cape Vera colony supports approximately 9,000 breed-ing pairs of fulmars, while Prince Leopold Island supportsat least 16,000 breeding pairs (Gaston et al. 2006).
Weather data were recorded at Cape Vera (2003–2005)and at Prince Leopold Island (2005) using a Davis® Van-tage Pro weather station, set to record hourly. In otheryears, Prince Leopold Island weather was recorded oncedaily using a handheld weather station. Ice conditions nearboth colonies were obtained from the EnvironmentCanada—Canadian Ice Service climate archives <http://ice-glaces.ec.gc.ca>, and regional weather conditions at theclosest communities were obtained from <http://www.weatheroffice.gc.ca>.
We used “apparently occupied sites” (AOS; Lloyd et al.1991) as our assessment of breeding pairs. This methodexcludes obvious non-breeding birds on ledges unsuitablefor nesting. Northern fulmars have very high Wdelity tobreeding sites (>90% if no mortality; Hatch and Nettleship1998), and thus we assumed that observations through theseason at a nest were observations of the same pair. Long-term population monitoring plots each supporting 25–300nests were established at both Cape Vera (n = 8) and PrinceLeopold Island (n = 7) (Mallory and Gaston 2005). Nestsites were viewed using 10 £ 42 binoculars or a 60£ spot-ting scope from vantage points 30–300 m distant along thecliV top, so birds were not disturbed during counting. Dueto poor weather (below), data were not necessarily col-lected on all plots each day.
We conducted detailed, daily observations (·4 h) onselected nest sites from four of the plots to determine whichmate was attending the nest, and to identify whether an eggor chick was in the nest. It is not easy to determine if a ful-mar is on an egg, as non-breeders often appear to be incu-bating for periods of hours, but then leave the nest site(Fisher 1952; Gaston et al. 2006). We had the same personnel
123

Polar Biol (2009) 32:529–538 531
monitor the same plots as much as possible within eachyear, which allowed observers to become familiar with theindividual bird’s appearance and behaviors. Occasionallynest failures occurred during periods when we could notview the nest (e.g., fog), or when prolonged observations ofadults still could not determine if the adult bird was on anegg (although behavioral cues often helped deduce nest sta-tus). In these cases, we ascribed nest failure date as the mid-point between the previous and next conclusive assessmentof nest contents (typical error §1 day). Due to logistic con-straints, we were unable to monitor chicks in fulmar neststhrough to Xedging. The duration of chick observationspost-hatch in key years were as follows: Cape Vera 2004—16.9 § 0.4 SE days (n = 39, range 10–19 days); Cape Vera2005—15.6 § 0.6 days (n = 40, range 3–20 days); PrinceLeopold Island 2005—11.5 § 0.6 days (n = 43, range 4–17 days). Thus, we monitored most nests until both parentsceased guarding the chick (10–14 days; Hatch and Nettleship1998). Although chick survival is generally high after thepost-guard stage (Hatch 1987; Falk and Møller 1997), wecaution that our estimates of Xedging and reproductive suc-cess may be biased high for some years.
Using daily observation data, we calculated mean dailyegg or chick survival to compare among sites and years.For all calculations, dates were converted to ordinal dates,with 1 January = 001. Methodologies used to study fulmarswere similar at both sites, but more Weld assistants wereused at Cape Vera, particularly in 2005. For this reason,and because some of the Prince Leopold Island data has
been presented previously (Gaston et al. 2005, 2006), werefer principally to Cape Vera data for certain analyses, butuse data from both sites and/or all years where possible.Depending on the distribution of data, we used t tests,ANOVAs, Pearson or Spearman rank correlations to com-pare data among years or colonies (Systat Software Inc.2002). To examine relationships between nest failure andweather, we grouped data by 3 day periods (to capture thefull extent of some storms), and used Spearman rank corre-lations to compare numbers of eggs lost in that period toaverage weather conditions over that period. Means are pre-sented §SE, and all statistical tests report two-tailed proba-bilities unless otherwise noted.
Results
Weather and ice
At Cape Vera, temperatures were below ¡10°C when ful-mars arrived at the breeding cliVs (Fig. 2a). Temperatureswarmed during the breeding season (3-year mean dailymeans; r83 = 0.74, P < 0.001), but overall maximum tem-peratures during incubation and early chick-rearing (8June–11 August) were cool (3.4 § 0.4°C, n = 65 days,range ¡2.7 to 14.0°C). The mean daily maximum windspeed was 13.2 § 1.6 km/h (8.0–62.8 km/h; n = 65), andgusts exceeded 100 km/h at least once each month. Fog wasfrequent (>25% of days each year), and snowfall occurred
Fig. 1 The location of the Cape Vera and Prince Leopold Island study sites in the Canadian high Arctic. Shaded areas on lands represent glaciers
123

532 Polar Biol (2009) 32:529–538
every month, with signiWcant wet snow accumulation(>10 cm) in July of each year. At Cape Vera (and probablyPrince Leopold Island), the laying period coincided withthe point at which ambient mean daily temperatures roseabove 0°C (Fig. 2).
Weather conditions were similar at Prince LeopoldIsland (Fig. 2b), but mean daily maximum temperaturesduring incubation and chick-rearing were warmer(6.5 § 0.4°C, n = 65, range ¡0.8 to 26.3°C). Blizzardsoccurred on 18–20 August 2001, and again on 13 June, 23July, and 13 August 2003. In the breeding seasons of2001–2005, mean daily maximum wind speed was18.3 § 1.0 km/h (0–70 km/h; n = 174 days). Fog was com-mon (>25% of days) in all years, but in 2005, 22 of 51(43%) days at the Weld site were too foggy or snow was tooheavy to safely conduct fulmar observations.
In early June 2001 and 2002 (i.e., egg-laying), sea-icewas >90% solid cover from Prince Leopold Island to thenearest open water 200 km to the east, whereas in 2003 and2005 there was open water immediately beside the colony
in early June. In contrast, at Cape Vera there was a pocketof open water (polynya) located immediately beside thecolony in early June, but sea-ice extended for approxi-mately 200 km to the west in Jones Sound, which remainedthrough much of July before breaking up in August. Duringthe previous decade (1996–2005), the mean distance toopen water at Cape Vera (excluding the Hell Gate Polynya)in the second week of June was 190 § 5 km, a distance thatwas consistent across years (coeYcient of variation 9%). Incontrast, at Prince Leopold Island the mean distance was42 § 39 km and was much more variable (coeYcient ofvariation 292%; F ratio test, F18 = 50.5, P < 0.001).
Weather and timing of reproductive failure
Reproductive failure occurred throughout the breeding sea-son at both colonies (Fig. 3a). Of 346 nest failures observedat Prince Leopold Island or Cape Vera during 2001–2005,67% occurred during incubation, and 33% occurred duringchick-rearing. At Cape Vera, six of 216 (3%) nest attemptsfailed because the pair pushed the egg out of the nest cup.At Prince Leopold Island, 22 of 306 (7%) nest attemptsfailed because eggs did not hatch (i.e., eggs were observedin the nest after the latest hatch date in the colony), a pro-portion signiWcantly larger than was observed in 2 years atCape Vera [2 of 216 (1%); Fisher Exact test, P = 0.0005].
Mean daily survival rates of eggs for the typical fulmarincubation period (6 June to 24 July) were high and statisti-cally similar at both colonies (Table 1; Kruskal–Wallis test,KW = 1.96, P > 0.3). There was no evidence that consis-tently more eggs were lost per day at any stage of incuba-tion: in all three colony-years, the mean number of eggslost each day in the Wrst half of incubation was statisticallysimilar to rates in the second half of incubation (Table 1;Mann–Whitney tests, all P > 0.17).
Of the 61 eggs lost on the monitoring plots at PrinceLeopold Island in 2003, 9 (15%) were conWrmed lost duringblizzards, and a further 10 eggs (16%) were only observedafter the snow cover had melted, suggesting fulmars aban-doned nests during the storms and eggs were buried. In 2003,eggs tended to be lost during periods of low barometric pres-sure (associated with snowstorms) but not necessarily duringperiods of greater wind speed (Table 1). Furthermore, 16%of the chicks in monitoring plots died (blew oV or were dep-redated) during the snowstorm on 13 August 2003, represent-ing 58% of total chick mortality. In 2001, 26% of monitoredchicks appeared to be killed by a snowstorm on 19–20August (not all of these deaths could be veriWed before theWeld party left), representing a minimum of 45% of totalchick mortality. Barometric pressure and wind speed werenot clearly related to nest failure in 2005 (Table 1), but 21 of37 (57%) nest failures during incubation occurred during a4 day storm in late July with rain and gusts >70 km/h.
Fig. 2 Mean (§SE) daily temperature at a Cape Vera, Devon Island,2003–2005, and b Prince Leopold Island, 2002, 2003, 2005. Linesabove the temperature graph indicate the periods of arrival at the col-ony by fulmars (A), laying (L) and hatching (H) at Cape Vera, whichare similar at Prince Leopold Island
123

Polar Biol (2009) 32:529–538 533
At Cape Vera in 2005, eggs tended to be lost during peri-ods of high wind speed but not lower barometric pressure(Table 1). Three storms supporting sustained east winds>40 km/h (gusts >70 km/h) led to 62% of the 52 nest fail-ures, including one on 6 August during which 46% of mon-itored chicks disappeared (presumably all depredated).
Interestingly, a major snowstorm on 12–14 July 2005(>20 cm), with westerly winds >40 km/h, did not result insubstantial egg loss (Fig. 3b).
Mean daily survival rates of chicks at both colonies (forthe Wrst »20 days after hatch) were also high and suggestedsome diVerences among years (Table 1; KW = 7.7,P = 0.052), with chick survival signiWcantly higher atPrince Leopold Island in 2005 than in 2003 (Dunn’s Multi-ple Comparisons test, P < 0.05). In 2005, no blizzardsoccurred at Prince Leopold Island during the portion of thenestling period that we observed (but see below).
Over 2 years at Cape Vera, more chicks were lost beforethey were 10-day old (23 chicks) than between 11- and 20-day old (three chicks; Binomial test, P < 0.001; Fig. 4). In2002, 2003 and 2005 at Prince Leopold Island, 22 chickswere lost before they were 10-day old, while 11 were lostbetween day 11 to 20 (Binomial test, one-tailed P = 0.04).Higher chick survival after day 10 corresponds to the startof the rearing period when both parents may leave the chickalone while they forage (Hatch and Nettleship 1998).
Annual reproductive success
Annual reproductive success varied at Cape Vera andPrince Leopold Island (Table 2). For both sites, approxi-mately 70% of the pairs at AOS produced an egg, but annu-ally 40–81% of those eggs hatched, and 42–88% of hatchedchicks survived to the post-guard stage. The greater valuesfor chick survival in 2004 and 2005 may be related to ourWeld schedule, because chicks were monitored for up to10 days less than in the earlier years. We have no records oflocal weather at either site after our departure, and localweather diVers from conditions at the nearest weather sta-tion in Resolute Bay (M.L. Mallory, unpublished data), sowe could not model Xedging success reliably. Based onmeteorological records at Resolute Bay (170 km west) andNanisivik (200 km east), there were no major storms (gusts>70 km/h, snowfall >5 cm) for the rest of the chick-rearingperiod (August or September) after our observations ended
Fig. 3 Cumulative mortality of fulmar eggs or chicks at Cape Vera in2005 (CV05) and Prince Leopold Island in 2003 (PLI03) and 2005(PLI05). The upper panel a depicts the timing of egg losses, with puls-es of mortality noted by arrows. In the lower panel b, the thin line ismaximum daily wind speed at Cape Vera in 2005 (wind speeds >0were from the north or east, winds <0 were from the south or west), thethick lines are cumulative egg (left) or chick (right) loss, and arrowsrepresent storms with easterly wind gusts >70 km/h
Table 1 A comparison of egg and chick survival rates and weather factors potentially inXuencing nest failure of northern fulmars at PrinceLeopold Island (PLI) and Cape Vera (CV)
DSR refers to daily survival ratea Egg losses during Wrst 8 days of incubation not observed in 2005
Colony Year
Mean § SE egg DSR (n nests)
Mean § SE chick DSR (n days)
Mean § SE egg loss/day (n) early incubation
Mean § SE egg loss/day (n) late incubation
Correlations of weather and egg loss
Barometric pressure Wind speed
rs (n) P rs (n) P
PLI 2003 0.9894 § 0.0023 (99) 0.9758 § 0.0081 (20) 1.2 § 0.3 (25) 0.4 § 0.1 (24) ¡0.54 (16) 0.03 ¡0.20 (16) 0.45
PLI 2005 0.9943 § 0.0014 (74)a 0.9984 § 0.0016 (17) 0.6 § 0.2 (17)a 1.1 § 0.4 (24) ¡0.09 (15) 0.74 0.31 (15) 0.24
CV 2004 0.9739 § 0.0099 (20)
CV 2005 0.9915 § 0.0026 (102) 0.9814 § 0.0058 (19) 0.5 § 0.2 (25) 0.9 § 0.4 (24) ¡0.19 (15) 0.5 0.59 (15) 0.02
123

534 Polar Biol (2009) 32:529–538
in 2004. Thus, our assumption of little chick mortality afterour departure in 2004 was probably reasonable. However,in 2005, major storms must have aVected the Weld sites on28–29 August, and again on 10–11 September. Thus, someadditional chick mortality likely occurred after our depar-ture in 2005, so our survival rates for 2005 may be overesti-mates.
Assuming no mortality occurred after our observationsceased, fulmars produced an estimated 0.27 Xedglings/AOSin 2004 and 0.36 Xedglings/AOS in 2005 at Cape Vera. AtPrince Leopold Island in 2005, fulmars produced 0.26Xedglings/AOS. Over these Wve breeding seasons and two
sites, 231 of 901 fulmar pairs at AOS (26%) produced achick that reached the post-guard stage (or 37% of the 628fulmar pairs that laid an egg).
Discussion
The northern fulmar colonies at Cape Vera and Prince Leo-pold Island share many similarities, including their nestingstructure, body size, proportion of diVerent color morphs,and breeding schedule (Hatch and Nettleship 1998; Malloryand Forbes 2005; Gaston et al. 2006; Mallory and Forbes2007). We identiWed some additional similarities betweenthese colonies that diVer from colonies elsewhere in thespecies’ range, but discovered notable diVerences in fulmarreproductive success at these sites.
Reproductive success and weather
Consistent with our prediction, northern fulmar colonies inthe Canadian high Arctic exhibited lower annual reproduc-tive success than Boreal fulmar colonies (Tables 2, 3; notethat methodologies diVer somewhat among studies),although the pattern and timing of reproductive failure wassimilar (most losses during incubation; Dunnet et al. 1979;Hatch and Hatch 1990). Although hatching success, esti-mated Xedging success, and chick survival (Greenland,100% after 20 days; Falk and Møller 1997) in our studywas similar to previous work (Table 3), we found that only26% of AOS in the Canadian high Arctic (231 chicks of901 AOS) produced a chick that Xedged (Table 2; annualmean 0.26 § 0.03 chicks/AOS, n = 6 colony-years, range
Fig. 4 Mean (SE) daily fulmar chick survival rates at Cape Vera,pooled for 2004 and 2005. The open circle on day 1 corresponds to dayof hatching, and was 100% survival because we could not account forchicks that may have been lost on that day before our observations.Daily sample sizes (open squares, right hand Y axis) declined withincreasing age, due both to mortality, or the end of our observationperiod
Table 2 Measures of reproductive performance of northern fulmars from Prince Leopold Island (PLI) and Cape Vera (CV) in the Canadian highArctic, 2001–2005
AOS refers to apparently occupied sites. Estimates of chicks per AOS, maximum Xedging success and productivity assume that all chicks reachingthe post-guard stage survived to Xedgea Maximum Xedging (chicks observed to »20-day old)b Nests were monitored for 10 days longer at PLI in 2001–2003 than in other years; values represent chicks surviving 14.3 § 0.4 days (n = 109chicks, range 3–20 days)
Colony year
AOS Eggslaid
Chicks hatched
Median date of hatching
Chicks reaching post-guard
Chicks per AOS
Proportion (%)
Pairs that laid
Eggs that hatched
Maximum hatched chicks that could Xedge
Maximum productivity (max. chicks Xedged/egg laid)
PLI 2001 172 127 103 28 July 43a 0.25 74 81 42 34
PLI 2002 132 87 39 28 July 17a 0.13 66 45 44 45
PLI 2003 124 102 41 28 July 28a 0.22 82 40 68 40
PLI 2005 182 96 51 26 July 45b 0.25 53 53 88 47
CV 2004 154 112 54 25 July 46b 0.30 73 48 85 41
CV 2005 137 104 65 25 July 52b 0.38 76 62 80 50
Mean (SE) 0.26 (0.03) 71 (4) 55 (6) 68 (8) 43 (2)
123

Polar Biol (2009) 32:529–538 535
0.13–0.38 per year). If our estimates of chick survival arehigh (see “Methods”), then the proportion of AOS that pro-duced a Xedgling would be even lower in high Arctic colo-nies. This contrasted with higher annual reproductivesuccess reported for Alaska, Greenland, and across theUnited Kingdom (up to twofold diVerence; Table 3).
These proportional diVerences are not explained bylower success per egg laid. The mean annual number ofchicks Xedged per egg in our study (43 § 2%, n = 6 col-ony-years) did not diVer from that in Alaska (37 § 5%,n = 15 years; Hatch and Nettleship 1998) or previously atPrince Leopold Island (40 § 4%, n = 4 years; Hatch andNettleship 1998; ANOVA, F2,22 = 0.32, P = 0.7), wassomewhat higher than the 27% average for Eynhallow,Scotland, but was lower than values for Greenland in 1993(Table 3). Instead, the lower success per AOS in Canadianhigh Arctic colonies must be related to a larger proportionof birds attending sites that do not attempt to breed (assum-ing we did not miss many early nest failures).
Studies in Greenland and the Canadian high Arctic sug-gest that up to 20% more fulmar pairs attending nest sitesdid not lay an egg compared to fulmars breeding in Alaskaor the United Kingdom (Table 3). In our study, resultscould have been biased by data from Prince Leopold Islandin 2001 and 2005, where nest observations were initiatedafter the median laying date, and thus we could have under-estimated true laying attempts. However, we paid particularscrutiny to the laying period at Cape Vera in 2005, toensure that we recorded all laying attempts, and eggs laidper AOS was similar in 2005 to most other years (Table 2).Moreover, fulmars have very high nest site Wdelity (Hatchand Nettleship 1998). We tracked 33 AOS where pairs laidan egg but it or the chick was lost. Fulmars (singles orpairs) observed at those AOS post-nest failure were thesame morphs as the nesting pair for 17.0 § 1.7 days (range5–39 days), suggesting that failed pairs remained at theirnest site for some time before leaving the colony. Thus, weposit that our potential bias would have reduced our mea-sure of chicks produced per egg laid, but not productivityper AOS.
Like other fulmarine petrels breeding at high latitudes(Hodum 2002; Creuwels et al. 2008), high Arctic fulmarshave a short window of opportunity in which to breed(Mallory and Forbes 2007). When environmental conditions ortheir own physical condition are insuYcient to meet thenecessary breeding schedule, selection should favor birdsthat skip breeding rather than incurring stress that mightreduce future chances of reproduction in these long-livedbirds (Trivers 1972), particularly in late or extensive iceyears when marine production, and consequently fulmarreproductive success, are reduced (Welch et al. 1992;Gaston et al. 2005). Thus we speculate that in any year inthe high Arctic, a larger proportion of fulmars may Wnd T
able
3A
com
pari
son
of r
epro
duct
ive
perf
orm
ance
of
nort
hern
ful
mar
s fr
om v
ario
us c
olon
ies
in th
e sp
ecie
s’ r
ange
(m
etho
dolo
gies
for
ass
essi
ng s
ome
para
met
ers
diV
ered
am
ong
stud
ies)
AO
S re
fers
to a
ppar
ently
occ
upie
d si
tes.
Mar
ine
ocea
nogr
aphi
c zo
nes
refe
r to
hig
h A
rctic
(H
A)
or B
orea
l (B
)
Col
ony
Mar
ine
zone
AO
S w
ith
eggs
(%
)H
atch
ing
succ
ess
(%)
Fled
ging
su
cces
s (%
)C
hick
s pe
r eg
g la
idC
hick
s pe
r A
OS
Hat
chab
ility
(%)
Tim
e Pe
riod
Ref
eren
ce
Mal
lem
ukfj
eld,
Gre
enla
ndH
A71
7582
0.56
0.44
8419
93Fa
lk a
nd M
ølle
r (1
997)
Cap
e V
era,
Nun
avut
HA
7455
820.
450.
3498
2004
–200
5T
his
stud
y
Pri
nce
Leo
pold
Isl
and,
Nun
avut
HA
7757
750.
430.
3387
1975
–197
8L
into
n an
d N
ettl
eshi
p (1
977)
, H
atch
and
Net
tles
hip
(199
8)
Pri
nce
Leo
pold
Isl
and,
Nun
avut
HA
8157
570.
320.
2693
2001
–200
5T
his
stud
y
Sem
idi I
slan
ds, A
lask
aB
8558
700.
370.
3193
1976
–199
5H
atch
and
Net
tlesh
ip (
1998
),
Hat
ch a
nd H
atch
(19
90)
Acr
oss
Uni
ted
Kin
gdom
B54
840.
450.
4295
1986
–200
3M
avor
eta
l. (2
005)
, Mou
gin
(196
7)
Eyn
hall
ow, S
cotla
ndB
910.
270.
16–0
.52
1950
–197
7D
unne
t eta
l. (1
979)
, O
llaso
n an
d D
unne
t (19
88)
123

536 Polar Biol (2009) 32:529–538
environmental conditions unsuitable and skip breeding,compared to fulmars breeding at colonies farther south.
Hatchability of fulmar eggs (i.e., the proportion of eggsincubated to full term that hatch) varied by colony and year(Table 3). We observed anomalously high hatchability inboth years at Cape Vera (98%, higher than reported formost other species; Koenig 1982). Fulmar pairs at CapeVera have a brief period together at the colony prior todeparting for their markedly long exodus, and while allfemale fulmars store sperm (Hatch and Nettleship 1998),the phenology of breeding at Cape Vera means that femalesmust release those sperm during the exodus, whereas atother locations, females release the sperm before departing.Because females at Cape Vera may retain stored spermlonger before fertilization, we had predicted elsewhere thatthese fulmars might have lower hatchability (or higherinfertility rates; Mallory and Forbes 2007), the opposite ofwhat we observed.
In comparison to Cape Vera, egg hatchability rates werelower in Alaska, Greenland, and at Prince Leopold Island(Table 3). It seems unlikely that we failed to observe ¸4nests with unhatched eggs in each year of study at CapeVera, the amount required to give similar hatchability pro-portions as elsewhere. Observed hatchability diVerencesseem unrelated to colony size, latitude or nest initiationdate (see Koenig 1982), and likely do not reXect infertilitydiVerences within the same species at diVerent colonies(Warham 1990). If the diVerences between Cape Vera andPrince Leopold Island are the result of embryo death ratherthan infertility, we suggest two possible mechanisms thatmight account for colony-speciWc diVerences in hatchabil-ity. First, Prince Leopold Island receives more heavy snow-fall through the season, whereas Cape Vera is colder (dueto the proximity of the ice cap; Fig. 1.) In some instances,blanketing snow during incubation, or nests initiated onsnow-covered ledges (Linton and Nettleship 1977) maylead to increased egg neglect (absence by both adults fromthe nest during incubation). Despite considerable chill tol-erance in fulmar eggs (Warham 1990), inexperienced ful-mar pairs may not attend nests enough during snowstormsto provide the required incubation, and the egg may die,even though parents return after the storm to incubate, andcontinue past term before abandoning the breeding attempt.Second, exposed fulmar eggs appear to be depredated veryquickly at Cape Vera by glaucous gulls (Larus hyperboreus),presumably because these are the only seabird prey avail-able to the gulls. At Prince Leopold Island, however, gullscan also prey on thick-billed murres (Uria lomvia) andblack-legged kittiwakes (Rissa tridactyla). If the costs offoraging on these other species are lower than risking con-tamination by fulmar stomach oil (potentially lethal; Swen-nen 1974), then predation pressure may be relatively loweron fulmars at Prince Leopold Island than at Cape Vera.
Hence, neglected eggs may remain uneaten for longer,allowing absent parents to return and continue incubation.Further investigation is required to assess whether hatch-ability of fulmar eggs is consistently high at Cape Vera, andif so, what factors are responsible for higher hatchability atthis site.
Weather at high Arctic colonies may aVect hatchabilityof eggs, but more importantly it is the major factor inXu-encing nest failure at both colonies we studied, in contrastto results of Boreal fulmars in Alaska (Hatch and Hatch1990). Wind storms were associated with pulses of nestfailure at Cape Vera, as were snowstorms with peaks ofnest failure at Prince Leopold Island. In contrast to the“pulses” of nest failure observed at high Arctic colonies,failure at Boreal colonies may be more gradual unlessstorms are profoundly intense. For example, Gray et al.(2003) noted that a nine day period of bad weather resultedin 36% mortality of fulmar chicks being monitored, and53% of total chick mortality, a proportion similar to whatwe observed in storms lasting 1 or 2 days (Fig. 3).
Typical weather and sea-ice patterns may also explainsome of the diVerences between our study and the results ofFalk and Møller (1997), studying fulmars at the NortheastWater Polynya (high Arctic) near Greenland. They hadfavorable weather conditions in their study (calm with nomention of signiWcant storms during incubation or earlychick-rearing), and they recorded chick survival of 91%,higher than we estimated in our study (68 § 8%), or atPrince Leopold Island in 1976 (88%; Linton and Nettleship1977), or the range observed in the northern United King-dom (55–87%; Gray et al. 2003). Thus, the Greenland datademonstrate that not all high Arctic colonies suVer weather-related losses every year.
Sea-ice and reproductive success
Gaston et al. (2005) showed that another climate-relatedvariable, late breakup of sea-ice, also had a deleteriouseVect on fulmar reproduction at Prince Leopold Island.However, sea-ice distribution is unlikely to inXuence ful-mar reproduction at Cape Vera as strongly, because the Xoeedge is more predictable at this site (this study). Open waterbeside the latter colony is always available in the HellGate–Cardigan Strait polynya by the time the fulmarsarrive, unlike the variable sea-ice conditions in early Mayat Prince Leopold Island. Moreover, satellite telemetryindicates that active and failed breeders at Cape Vera Xy>200 km one way to feed (Mallory et al. 2008), and heli-copter transects near Cape Vera have found few fulmarsbeyond a 2 km radius from the colony during the breedingseason (ML Mallory, unpublished data). In contrast, manyfulmars apparently feed closer to Prince Leopold Islandduring the breeding season (<50 km; Nettleship and Gaston
123

Polar Biol (2009) 32:529–538 537
1978). Fulmars feeding chicks at Cape Vera typically havefew distinguishable parts in regurgitations (suggesting along retention time), whereas items in regurgitations atPrince Leopold Island can often be identiWed (Linton andNettleship 1977; Hatch and Nettleship 1998; A. J. Gastonunpublished data), again suggestive of relatively longerforaging trips by adults at Cape Vera.
We hypothesize that colony-speciWc behavioral adapta-tions may allow fulmars at Cape Vera to successfully breeddespite extensive but predictable sea-ice near the colonypersisting to the chick-rearing period, compared to PrinceLeopold Island. These adaptations may include an earlierand more synchronized breeding phenology within andacross years (Mallory and Forbes 2007), despite being far-ther north and isolated by sea-ice for longer. Future moni-toring at Cape Vera will determine if phenology andreproductive success are less variable across years com-pared to Prince Leopold Island.
Fulmar colonies in the high Arctic
Our study demonstrated that certain aspects of fulmarreproductive ecology are common across the species range.However, fulmars breeding at colonies in the high Arctichave developed a compressed breeding schedule (Malloryand Forbes 2007), and more birds attended colonies with-out breeding each year, resulting in lower chicks Xedgedper AOS, compared to Low Arctic and Boreal colonies.Extreme weather events during breeding, such as heavysnowfall or strong winds, led to considerable nest failure.
If the number of Xedglings produced per egg laid is simi-lar across the species’ range, but high Arctic fulmars experi-ence more nest weather-induced nest failure, then highArctic fulmars must experience less Xuctuation in marinefood supplies, a major factor inXuencing fulmar reproductivesuccess elsewhere (Hatch and Hatch 1990; Gray et al. 2003).At Cape Vera, fulmars forage in a large, recurrent polynyaon natural prey, and thus polynya productivity may be stableacross years. Moreover, the pulse of marine production inArctic waters around these colonies probably peaks duringchick-rearing (Welch et al. 1992; Barbraud and Chastel1998), making acquisition of food relatively easy for provi-sioning young, and perhaps oVsetting losses caused by ran-dom, catastrophic storms. In contrast, Boreal fulmars mayhave more dietary options, including Wsh and Wsheries dis-cards (Gray et al. 2003; Mallory 2006), but prey availabilitymay vary more in relation to climate cycles and economics.Given that high Arctic and Boreal fulmars overlap in winter-ing conditions in the North Atlantic Ocean (Mallory et al.2008), a simultaneous investigation of colony attendance,and the proximate causes of nest failure at colonies in bothoceanographic zones would help identify the relative impor-tance of factors inXuencing reproductive success.
Acknowledgments All research was conducted under approved per-mits as follows: animal care (2004PNR017, 2004PNR021), land use(59A/7-2-2), wildlife research (NUN-SCI-00-04, NUN-SCI-03-02,WL000714). Many collaborators made the Cape Vera and Prince Leo-pold Island projects possible in 2000–2005, and in particular J. Ake-arok, T. Arsenault, J. Berge, S. Chisholm, D. Edwards, C. Eberl, C.Ferguson, A. Fontaine, A. Moenting, B. Newton, K. O’Donovan, M.Robertson, K. Truman, and K. Woo. J. Creuwels, S. Hatch, and twoanonymous referees provided insightful reviews of the manuscript.Financial and logistic support were provided by Environment Canada(CWS, NEI), Natural Resources Canada (PCSP), the Nunavut WildlifeManagement Board, Indian and Northern AVairs Canada (NSTP), andCarleton University.
References
ACIA (2005) Arctic climate impact assessment. Cambridge UniversityPress, Cambridge
Barbraud C, Chastel O (1998) Southern fulmars molt their primaryfeathers while incubating. Condor 100:563–566
Creuwels JCS, van Franeker JA, Doust SJ, Beinssen A, Harding B,Hentschel O (2008) Breeding strategies of Antarctic petrelsThalassoica antarctica and southern fulmars Fulmarus glacialoidesin the high Antarctic and implications for reproductive success. Ibis150:160–171
Dunn PO, Thusius KJ, Kimber K, Winkler DW (2000) Geographic andecological variation in clutch size of tree swallows. Auk 117:215–221
Dunnet GM, Ollason JC, Anderson A (1979) A 28-year study of breed-ing fulmars Fulmarus glacialis in Orkney. Ibis 121:293–300
Falk K, Møller S (1997) Breeding ecology of the fulmar Fulmarusglacialis and the kittiwake Rissa tridactyla in high-arctic north-eastern Greenland, 1993. Ibis 139:270–281
Fisher J (1952) The fulmar. Collins, LondonFortescue M (1999) Temporal and spatial variation in breeding success
of the little penguin Eudyptula minor on the east coast of Austra-lia. Mar Ornithol 27:21–28
Furness RW, Camphuysen CJ (1997) Seabirds as monitors of themarine environment. ICES J Mar Sci 54:726–737
Gaston AJ, Gilchrist HG, Mallory ML (2005) Variation in ice condi-tions has strong eVects on the breeding of marine birds at PrinceLeopold Island, Nunavut. Ecography 28:331–344
Gaston AJ, Mallory ML, Gilchrist HG, O’Donovan K (2006) Status,trends and attendance patterns of the northern fulmarFulmarus glacialis in Nunavut, Canada. Arctic 59:165–178
Gray CM, Phillips RA, Hamer KC (2003) Non-random nestling mor-tality in northern fulmars: implications for monitoring marineenvironments. J Zool (Lond) 249:109–113
Hatch SA (1987) Adult survival and productivity of northern fulmarsin Alaska. Condor 89:685–696
Hatch SA, Hatch MA (1990) Components of breeding productivity ina marine bird community: key factors and concordance. Can JZool 68:1680–1690
Hatch SA, Nettleship DN (1998) Northern fulmar (Fulmarusglacialis). In: Poole A, Gill F (eds) The birds of North America,No. 361. The Birds of North America Inc., Philadelphia
Hodum PJ (2002) Breeding biology of high-latitude Antarctic fulma-rine petrels (Procellariidae). J Zool (Lond) 256:139–149
Koenig WD (1982) Ecological and social factors aVecting hatchabilityof eggs. Auk 99:526–536
Linton A, Nettleship DN (1977) Biological investigation of northernfulmars (Fulmarus glacialis) at Prince Leopold Island, NWT,summer 1976. Studies on Northern Seabirds Report No 53. Avail-able from Environment Canada, Sackville, NS
123

538 Polar Biol (2009) 32:529–538
Lloyd CS, Tasker ML, Partridge K (1991) The status of seabirds inBritain and Ireland. T&AD Poyser, London
Mallory ML (2006) The northern fulmar (Fulmarus glacialis) inArctic Canada: ecology, threats, and what it tells us about marineenvironmental conditions. Environ Rev 14:187–216
Mallory ML, Forbes MR (2005) Sex discrimination and measurementbias in northern fulmars (Fulmarus glacialis) from the CanadianArctic. Ardea 93:25–36
Mallory ML, Forbes MR (2007) Does sea-ice constrain the breedingschedules of high Arctic northern fulmars? Condor 109:895–907
Mallory ML, Gaston AJ (2005) Monitoring northern fulmars in theCanadian arctic: plot locations and counts at selected colonies.Canadian Wildlife Service Technical Report 432
Mallory ML, Akearok J, Edwards DB, O’Donovan K, Gilbert CD(2008) Autumn migration and wintering of northern fulmars(Fulmarus glacialis) from the Canadian high Arctic. Polar Biol31:745–750
Mavor RA, Parsons M, Heubeck M, Schmitt S (2005) Seabird numbersand breeding success in Britain and Ireland, 2004. Joint NatureConservation Committee, Peterborough
Mougin J-L (1967) Etude ecologique des deux especes des fulmars: lefulmar Atlantique (Fulmarus glacialis) et le fulmar Antarctique(Fulmarus glacialoides). Oiseau 37:57–103
Nettleship DN, Gaston AJ (1978) Patterns of pelagic distribution ofseabirds in western Lancaster Sound and Barrow Strait, Northwest
Territories, in August and September 1976. ESCOM Report NoAI–09, Environment Canada, Ottawa
Ollason JC, Dunnet GM (1988) Variation in breeding success infulmars. In: Clutton-Brock TH (ed) Reproductive success. Universityof Chicago Press, Chicago, pp 263–278
Salomonsen F (1965) The geographical variation of the fulmar (Fulm-arus glacialis) and the zones of marine environment in the NorthAtlantic. Auk 82:327–355
Swennen C (1974) Observations on the eVect of ejection of stomachoil by the fulmar Fulmarus glacialis on other birds. Ardea62:111–117
Systat Software Inc. (2002) Systat 10.2. Systat, RichmondThompson PM, Ollason JC (2001) Lagged eVects of ocean climate
change on fulmar population dynamics. Nature 413:417–420Trivers RL (1972) Parental investment and sexual selection. In: Camp-
bell B (ed) Sexual selection and the descent of man 1871–1971.Aldine, Chicago, pp 136–179
Warham J (1990) The petrels-their ecology and breeding systems.Academic Press, London
Welch HE, Bergmann MA, Siferd TD (1992) Energy Xow through themarine ecosystem of the Lancaster Sound region, Arctic Canada.Arctic 45:343–357
Welty JC, Baptista L (1988) The life of birds, 4th edn. HarcourtCollege Publishers, Toronto
123