Embed Size (px)
Citation preview

Induction of vacuolar apical compartments in the Caco-2 intestinal
epithelial cell line
THIERRY GILBERT* and ENRIQUE RODRIGUEZ-BOULANT
Department of Cell Biology and Anatomy, Cornell University Medical College, New York, NY 10021, USA
*Present address: INSERM U.319, University Paris 7, 75005 Paris, France t Author for correspondence
Summary
Complete disruption of the microtubular network by colchicine or nocodazole in Caco-2 intestinal epi- thelial cells results in the appearance of basolateral microvilli and brush border-containing intracellular vacuoles (vacuolar apical compartment: VAC). These vacuoles are surrounded by a terminal web, express apical markers and exclude basolateral markers. The vacuoles do not originate from internalized apical or basolateral plasma membrane and their development is blocked by protein synthesis inhibi- tors, suggesting that they are newly synthesized. After removal of the microtubule inhibitors, VACs
are usually degraded and/or released into the lateral intercellular space. Rarely was fusion with the apical membrane observed. These experiments support a role for microtubules in the biogenesis of the apical surface and indicate that, under some conditions, apical plasma membrane assembly may occur in the cytoplasm, as observed in some human pathological states.
Key words: VAC, Davidson's disease, epithelial polarity, Caco-2 cells.
Introduction
Transporting epithelial cells exhibit two distinct plasma membrane domains, apical and basolateral, separated by tight junctions. The polarized surface distribution of transporting systems between these two surfaces under- lies the many vectorial functions of epithelial cells (Simons and Fuller, 1985; Rodriguez-Boulan and Nelson, 1989). Considerable advances have been recently made on the biogenesis of epithelial cell polarity. In the model kidney-derived cell line MDCK, the biosynthetic path- ways of apical and basolateral proteins appear to diverge in the trans Golgi network (intracellular sorting; Griffiths and Simons, 1986; Matlin and Simons, 1984; Misek et al. 1984; Rindler et al. 1984). Consistent with these obser- vations, two distinct transport vesicle populations have been identified that are thought to mediate the delivery of plasma membrane proteins to the apical and basolateral domains (Wandinger-Ness et al. 1990). On the other hand, in other systems, such as intestinal and liver epithelial cells (including the human colon carcinoma cell line Caco- 2), sorting of many proteins takes place a t the basolateral surface (Massey et al. 1987; Bartles et al. 1987; Le Bivic et al. 1990; Matter et al. 1990).
The Caco-2 cell line undergoes a spontaneous enterocy- tic differentiation upon in vitro culture. These cells express a well-developed brush border and many hydro- lases specific for the small intestine or the fetal colon (Pinto et al. 1983; Chantret et al. 1988). Recent work explored the role of microtubules in targeting processes in Caco-2 cells. Nocodazole treatment was found to retard both direct and indirect apical pathways with no effect on the final distribution of apical or basolateral plasma
membrane proteins (Matter et al. 1990) whereas colchicine treatment led to a completely non-polarized delivery of some apical markers (Gilbert et al. 1991). In vivo, colchicine treatment is known to interfere with the apical pathway (Quaroni et al. 1979; Achler et al. 1989) and to induce ultrastructural modifications like basolateral mic- rovilli and intracytoplasmic brush border vacuoles (Pavelka et al. 1983; Achler et al. 1989). A familial enteropathy, microvillus inclusion disease or Davidson's disease, is characterized by a highly reduced apical brush border and the appearance of large intracellular vacuoles with inwardly facing microvilli in enterocytes (Cutz et al. 1989; Davidson et al. 1978). Attempts to obtain an in vitro model for this disease were made using primary cultures of human fetal intestine but only partial results were reported (Carruthers et al. 1986).
We report here that microtubule-disrupting agents induce the appearance of basolateral brush border and apical-like intracellular vacuoles in Caco-2 monolayers. Using laser scanning confocal microscopy and electron microscopy, we investigated the formation and origin of these vacuoles and their behavior upon removal of the microtubule inhibitors. We discuss these observations as a possible in vitro model for Davidson's disease.
Materials and methods
Reagents Cell culture reagents were purchased from Gibco Laboratories (Grand Island, NY). Sulfosuccinimidyl 6-(biotinamido) hexa- nonate (NHS-LC-biotin) and Texas red(TR)-avidin were from Pierce (Rockford, IL). Fluorescein (F1TC)-conjugated goat anti-
Journal of Cell Science 100, 451-458 (1991) Printed in Great Britain @The Company of Biologists Limited 1991

mouse or anti-rabbit were from Jackson Immunoresearch (WestGrove, PA). All other reagents were purchased from Sigma (StLouis, MO).
Cell culture and drug treatmentCaco-2 human intestinal cells were obtained from A. Zweibaum(INSERM U.178, Villejuif, France) and grown in DME sup-plemented with 10 % FBS, 1 % non-essential amino acids, withoutantibiotics. For experiments, cells were plated on polycarbonatefilters with 0.4/on pores (Transwell no. 3412, Costar Inc.,Cambridge, MA) and grown for 2-3 weeks after confluency topermit complete differentiation of the cells (Pinto et al. 1983).
All drugs were prepared in stock solutions at lOmgml"1 inDMSO (dimethyl sulfoxide) and stored frozen at -20°C. Prior toeach experiment, samples were diluted in cold normal mediumand drugs used at the following concentrations: colchicine (10 or20 im), and nocodazole (10 or 33JJM). After transepithelialresistance (TER) measurement, cells were washed twice with ice-cold PBS containing O.lmgmr1 CaCl2 and l rngmP 1 MgCl2(PBS/CM), then incubated in cold normal medium containing thedrug for 30 min at 4°C and warmed up at 37 °C in the presence ofthe drug for 3 or 6 h. Control experiments were performed in thepresence of 0.04% DMSO. In some experiments, filter-grown cellswere biotinylated from either the apical or the basolateral side asdescribed (Sargiacomo et al. 1989) and then exposed to the drug.For inhibition of protein synthesis, cycloheximide was used at
x
AntibodiesRabbit polyclonal antibodies against human placental alkalinephosphatase were from Accurate (Westbury, NY) and a mono-clonal antibody against human transferrin receptor (TFR) wasfrom Boerhinger (Mannheim, FRG). Mouse monoclonal anti-bodies against sucrase (HSI 9), isomaltase (HSI 14), aminopepti-dase N (HBB2/45), and dipeptidylpeptidase IV (DA07/219) havebeen described (Beaulieu et al. 1989; Quaroni and Isselbacher,1985) and were a generous gift from A. Quaroni. Monoclonalantibody 525-5-4 against Ag 525 and Mab 517 against antigen517 have been previously characterized (Le Bivic et al. 1987,1988a) and were a gift from A. Le Bivic. Actin filaments wererevealed by using FITC-phalloidin at l^gml"1.
Indirect immunofluorescence and laser scanning confocalmicroscopy (LSCM)Cells grown on filters were fixed, processed and observed bysimple indirect immunofluorescence as already described (Gilbertet al. 1991). For double indirect immunofluorescence, combinationof FITC-conjugated second antibobies and TR-avidin was used.Epifluorescence examination was performed with a NikonOptiphot microscope using a xlOO lens (1.4 numerical aperture).LSCM was obtained by a Phoibos 1000 unit attached to themicroscope (Sarastro, Molecular Dynamics, Sunnyvale, CA). Todetect simultaneously FITC- and TR-labeled antigens, sampleswere excited at 514 nm and emitted light between 525 and 555 nmwas recorded for FITC and above 630 nm for TR. The cellmonolayer was optically sectioned in horizontal (x-y) or vertical(x-z) planes. Images generated by the Personal Iris graphicsworkstation (Silicon Graphics, Mountain View, CA) and Sarastrosoftware were photographed from the screen using a Minoltax500 camera and a 200 mm objective. Kodak Tmax 100 and Ektar125 films were used.
Electron microscopyControl and drug-treated cells grown on filters were processed forultrastructural observation as described (Gilbert et al. 1991). Insome experiments, to detect any compartment in communicationwith the apical or intercellular space, cells were postfixed fromthe apical or basolateral side of the filter in 2% lanthanumnitrate, 1% OsO4 in 0 . 1 M S-collidine buffer at pH7.2 for 2haccording to Revel and Karnovsky (1967). This staining was also
used to enhance the basolateral plasma membrane contrast.Samples were then dehydrated and embedded in Epon. Ultrathinsections were cut perpendicular to the filter and viewed with aJEM electron microscope (JEOL) at 80 kV.
Results
Induction of VACs by microtubule-disrupting drugsUnder normal monolayer culture conditions, the humanintestinal Caco-2 cells spontaneously differentiate intocolumnar enterocyte-like cells with apical brush borderand hydrolases (Pinto et al. 1983; Chantret et al. 1988).When confluent filter-grown Caco-2 cells were treatedwith colchicine at low concentrations (10/um for 6h or20 jian for 3h), the microtubular network virtually disap-peared (Gilbert et al. 1991). A striking feature ofcolchicine-treated monolayers was the development oflarge intracellular vacuoles, easily detected by indirectimmunofluorescence and LSCM. Vertical optical sectionsgenerated by LSCM revealed that these vacuoles werelabeled with FITC-phalloidin as strongly as the apicalbrush border (Fig. IB). A weak basolateral staining wasobserved in control and colchicine-treated cells due to thesubmembranous actin network and seemed to indicatethat these vacuoles are intracellular. Basolateral markerswere not expressed in these vacuoles (Fig. ID). Thesestructures were brightly labeled with antibodies againstmost peptidases known to be expressed on the apicalplasma membrane of Caco-2 cells, e.g. sucrase-isomaltase(Fig. 2A, B and D; Table 1). In addition, apical markerswere also observed in numerous smaller vesicles (Figs 2Cand 3C) located throughout the cytoplasm. Vertical opticalsectioning through the vacuoles confirmed their locationwithin the monolayer (Fig. 2B, C and D). To demonstratethe intracellular (as opposed to intercellular) location ofthese vacuoles, basolateral plasma membranes werelocalized at the end of the colchicine treatment by biotinlabelling and Texas-red-conjugated avidin (Fig. 3). Combi-nation of horizontal (i.e. parallel to the filter) and vertical(i.e. perpendicular to the monolayer) serial sectionsconfirmed their intracellular location and showed noconnections with apical or basolateral plasma membranes.Thus, the colchicine-induced large (8-10 f.im) intracellularvacuoles are equivalent to the vacuolar apical compart-ment (VAC) that was previously described in MDCK cells(Vega Salas et al. 1987). VACs are preferentially located atthe lower half of the cell (see Figs 2 and 3) but mayoccasionally occupy the entire cell height (Fig. 1). VAC
Table 1. Expression of apical and basolateral markersin VACs induced by microtubule-disrupting drugs
Presence in VAC
Apical markers:
Basolateral markers:
Actin*Alkaline phosphataseAminopeptidase NDipeptidylpeptidase IVIsomaltaseMab.517Sucrase
Ag.525Transferrin receptorUvomorulin
All the apical markers tested were present in microvillar inclusions.* Actin, also present along the lateral walls, is considered here an
apical marker because of the much stronger phalloidin apical stainingdue to well-developed brush borders.
452 T. Gilbert and E. Rodriguez-Boulan

Fig. 1. Induction of VAC in Caco-2 intestinal epithelial cells. Control (A and C) and 6h colchicine-treated (B and D) monolayers offilter grown Caco-2 cells were fixed and processed for immunofluorescence. Labeling with FITC-phalloidin (A and B) andimmunostaining of Ag.525 (C and D) were performed and viewed by LSCM on vertical optical sections perpendicular to the filter.Two large vacuoles brightly stained with FITC-phalloidin were present in this slice of the colchicine-treated monolayer (B). Thelateral cellular walls were also labeled, indicating an intracellular location for the VACs. The basolateral marker Ag 525 wasnever found in colchicine-induced VACs, as shown in D. Bar, 10/an.
structures staining with both FITC-phalloidin and apicalmarker antibodies were observed in ~5 % of the cells.
To characterize these VACs further, we examinedcolchicine-treated cells by electron microscopy. Fig. 4shows a large VAC (diameter ~6^m), lined with acomplete brush border and a terminal web, in thecytoplasm of a Caco-2 cell. VAC microvilli are 2,um inlength, as are those facing the external medium. Theinternal lumen is filled with a flocculant slightly electron-dense material and some small vesicle-like structures.Cells containing VACs usually have a very reduced apicalsurface, with few microvilli still present above the tight-junctional complexes, which results in a pyramidal shape.We also observed ectopic microvilli with well-developedcore filaments along the lateral walls of the cells, but theirlength rarely exceeded 1 ,um (Fig. 4, top inset). Further-more, numerous areas containing glycocalyx-like materialwere seen along the lateral plasma membranes (bottominset).
The two different colchicine concentrations we used leadto VAC formation in Caco-2 cells. VAC induction wasdetected within 3 h when cells were treated with 20 JXM ofthe drug. A 10 fiM treatment for the same period of timeinduced the formation of only basolateral brush borderand microvilli; 6h incubation with 10 JAM was necessary todetect VACs. Nocodazole (33 /m, 6h) also induced theformation of VACs and the expression of apical markers onlateral membranes and in intracellular vesicles (data notshown). Upon examination following a 3h treatment at33 [IM, we observed few intracellular and basolateral areaslabeled with apical markers. With a higher dose (50 ^M),we did not see an earlier induction of VAC in Caco-2 cells.
Colchicine therefore seemed to be more effective in VACinduction than nocodazole in these cells.
Origin of VACsExperiments were designed to test whether VACs werederived from internalized plasma membranes or werenewly synthesized. To examine the first hypothesis weused the following protocol. Filter-grown Caco-2 cells werelabelled from the apical or basolateral side with a biotinanalog at 4°C (Sargiacomo et al. 1989), incubated in thepresence of microtubule-disrupting drug for 6 h to induceVACs, and processed for double indirect immunofluor-escence with an apical marker/FITC-second antibody andTR-avidin (Fig. 5). Apical or basolateral biotinylationprior to drug treatment never resulted in the detection ofbiotin within VACs (Fig. 5B and D) or the numeroussmaller intracytoplasmic vesicles expressing apicalmarkers (Fig. 5A and C). We conclude that VACs are notformed by internalization of apical or basolateral plasmamembranes. These experiments provided additional evi-dence that VAC-expressing cells have reduced (Fig. 5D) orpracticaly absent (Fig. 5C) free apical surfaces. However,as proposed by Achler et al. (1989) using an in vivo system,newly synthesized basolateral brush border might becomeinternalized in brush border vacuoles during colchicinetreatment. To check this possibility, Caco-2 cells weretreated for 3h with colchicine (10 /JM), incubated foranother 3h with the drug in presence of sucrase-isomaltase antibodies added to either the apical orbasolateral media. Cells were then processed for doubleindirect immunofluorescence with Texas-red-conjugatedgoat anti-mouse and FITC-phalloidin. VACs induced
VAC in Caco-2 cells 453
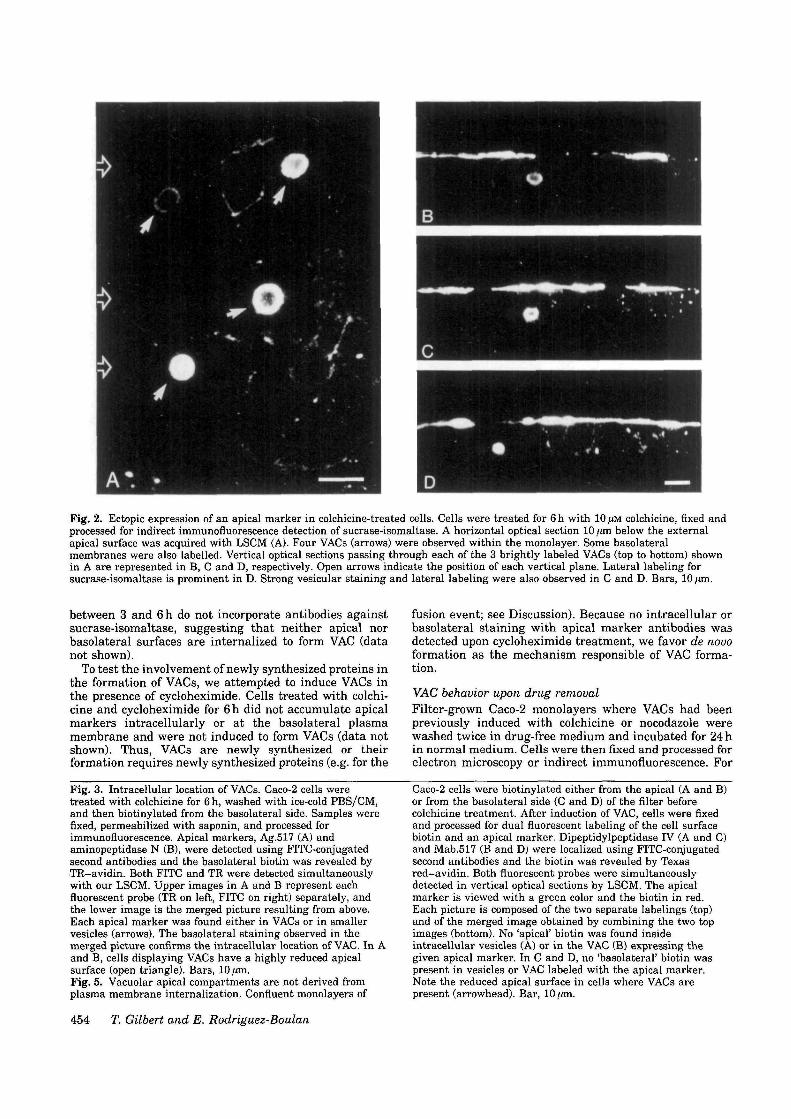
Fig. 2. Ectopic expression of an apical marker in colchicine-treated cells. Cells were treated for 6 h with 10 ,UM colchicine, fixed andprocessed for indirect immunofluorescence detection of sucrase-isomaltase. A horizontal optical section 10 /on below the externalapical surface was acquired with LSCM (A). Four VACs (arrows) were observed within the monolayer. Some basolateralmembranes were also labelled. Vertical optical sections passing through each of the 3 brightly labeled VACs (top to bottom) shownin A are represented in B, C and D, respectively. Open arrows indicate the position of each vertical plane. Lateral labeling forsucrase-isomaltase is prominent in D. Strong vesicular staining and lateral labeling were also observed in C and D. Bars, 10 /an.
between 3 and 6h do not incorporate antibodies againstsucrase-isomaltase, suggesting that neither apical norbasolateral surfaces are internalized to form VAC (datanot shown).
To test the involvement of newly synthesized proteins inthe formation of VACs, we attempted to induce VACs inthe presence of cycloheximide. Cells treated with colchi-cine and cycloheximide for 6h did not accumulate apicalmarkers intracellularly or at the basolateral plasmamembrane and were not induced to form VACs (data notshown). Thus, VACs are newly synthesized or theirformation requires newly synthesized proteins (e.g. for the
fusion event; see Discussion). Because no intracellular orbasolateral staining with apical marker antibodies wasdetected upon cycloheximide treatment, we favor de novoformation as the mechanism responsible of VAC forma-tion.
VAC behavior upon drug removalFilter-grown Caco-2 monolayers where VACs had beenpreviously induced with colchicine or nocodazole werewashed twice in drug-free medium and incubated for 24 hin normal medium. Cells were then fixed and processed forelectron microscopy or indirect immunofluorescence. For
Fig. 3. Intracellular location of VACs. Caco-2 cells weretreated with colchicine for 6 h, washed with ice-cold PBS/CM,and then biotinylated from the basolateral side. Samples werefixed, permeabilized with saponin, and processed forimmunofluorescence. Apical markers, Ag.517 (A) andaminopeptidase N (B), were detected using FITC-conjugatedsecond antibodies and the basolateral biotin was revealed byTR-avidin. Both FITC and TR were detected simultaneouslywith our LSCM. Upper images in A and B represent eachfluorescent probe (TR on left, FITC on right) separately, andthe lower image is the merged picture resulting from above.Each apical marker was found either in VACs or in smallervesicles (arrows). The basolateral staining observed in themerged picture confirms the intracellular location of VAC. In Aand B, cells displaying VACs have a highly reduced apicalsurface (open triangle). Bars, 10 /on.Fig. 5. Vacuolar apical compartments are not derived fromplasma membrane internalization. Confluent monolayers of
Caco-2 cells were biotinylated either from the apical (A and B)or from the basolateral side (C and D) of the filter beforecolchicine treatment. After induction of VAC, cells were fixedand processed for dual fluorescent labeling of the cell surfacebiotin and an apical marker. Dipeptidylpeptidase IV (A and C)and Mab.517 (B and D) were localized using FITC-conjugatedsecond antibodies and the biotin was revealed by Texasred-avidin. Both fluorescent probes were simultaneouslydetected in vertical optical sections by LSCM. The apicalmarker is viewed with a green color and the biotin in red.Each picture is composed of the two separate labelings (top)and of the merged image obtained by combining the two topimages (bottom). No 'apical' biotin was found insideintracellular vesicles (A) or in the VAC (B) expressing thegiven apical marker. In C and D, no 'basolateral' biotin waspresent in vesicles or VAC labeled with the apical marker.Note the reduced apical surface in cells where VACs arepresent (arrowhead). Bar, 10 /an.
454 T. Gilbert and E. Rodriguez-Boulan



\
\
\
\
\
v\
\\
Fig. 4. Electron micrographs of filter-grown Caco-2 monolayers treated with colchicine for 6 h. In the cell delimited by the brokenlines, a large vacuolar apical compartment (VAC) is present and has an infranuclear location. VACs are characterized by thepresence of a well-developed brush border facing an internal lumen (*), and are surrounded by a terminal web where microvilli areanchored (arrowheads). This VAC has a diameter of about 6,um. The apical external surface of the cell exhibiting a VAC almostdisappeared (arrow, upper left) but the junctional complexes were still visible. Open arrows indicate the interface between the filterand the basal plasma membrane. Numerous microvilli along the lateral cellular walls (top inset) and glycocalyx-like structures(bottom inset) were found protruding into the intercellular space. Bars: 2 ^m; 1 /on, top inset; and 0.5 ,um, bottom inset.
VAC in Caco-2 cells 455

6A
• i "
B
456 T. Gilbert and E. Rodriguez-Boulan

Fig. 6. Drug removal induced lost of VAC expression.Nocodazole- (A) or colchicine- (B) treated Caco-2 cells werewashed twice in drug-free medium and incubated for 24 h inregular medium. Cells were then processed for electronmicroscopy or indirect immunofluorescence. As shown in A, theintercellular space was filled with numerous heterogeneousmaterials (arrows). Examined at higher magnification, some ofthis material resembled remnants of microvilli (inset). The cellin the middle is still in contact with the external apicalmedium despite a compressed apical web (arrowheads). Nearby,a small vesicle containing microvilli was present close to theapical surface (open arrow). In B, an extracellular tracer,lanthanum nitrate, was added from the basolateral side duringthe postfixation step. The lateral intercellular space was filledwith flocculant material, and aggregates of vesicle-likestructures and was clearly dilated in some areas (*) but thejunctional complexes remained tight (arrowhead). Somestructure appeared to be degenerate VACs that had fused withthe lateral space (arrow). After a 12 h recovery period, indirectimmunofluorescence for sucrase-isomaltase (inset) showedbright apical staining and discontinuous internal labeling,suggesting a fusion event between a VAC and the lateralplasma membrane. Bars: 2.5 ;<m A and B; and 5;tm, each inset.
both drugs, the microtubular network reappeared and nomore VACs were observed after a one-day recovery period.
After nocodazole treatment and removal, electron-microscopic examination revealed that the intercellularspace was filled with numerous vesicle-like structures andheterogeneous material (Fig. 6A). Some of these struc-tures resembled degraded microvilli (Fig. 6A, inset). InFig. 6A, the cell in the center has a narrow apicalcytoplasm and a dramatically reduced apical surface,seemingly compressed by two adjacent cells, but withjunctional complexes still present at the apex.
After colchicine treatment, fixed monolayers werepostfixed in the presence of lanthanum nitrate added fromthe basolateral side of the filter to enhance the lateralplasma membrane contrast. Large dilatations of thelateral intercellular space were detected (Fig. 6B). Withinthese lateral lumina, electron-dense materials and vacuo-lar structures were detected. No tracer was found on theapical surface, indicating that the monolayer remainedtight. Confocal microscopy studies performed after a 12 hrecovery period suggested that VACs might be fusing withthe lateral membrane (Fig. 6B, inset).
Discussion
The occurrence of intracellular lumina has been reportedin many malignant epithelial cells but is rare in normalcells (for review, see Remy, 1986). An example of thisprocess has been described in vitro in the epithelial cellline HT-29 derived from a human adenocarcinoma (Remyet al. 1984). This system was used to study epithelialdifferentiation and polarity. Briefly, undifferentiated HT-29 cells (grown in the presence of glucose) form multi-layers and develop intracellular and intercellular cystslined with brush border (Zweibaum et al. 1985; Pinto et al.1988). Upon replacement of glucose with galactose, HT-29cells differentiate, lose their VACs, and acquire a polarizedphenotype (Remy et al. 1984; Zweibaum et al. 1985; LeBivic et al. 19886). On the other hand, the human coloncarcinoma cell line Caco-2, when grown in glucose-supplemented medium, display a differentiated phenotypeand no intracellular lumina (VACs) or intercellular cysts(Pinto et al. 1983; Chantret et al. 1988). In vivo, transient
secondary intercellular lumina were also described duringdifferentiation of the fetal rat colon epithelium, but nointracellular lumina were observed (Colony and Neutra,1983).
No clues exist on the biogenesis of these largemicrovillar inclusions. The results of this report usingCaco-2 cells suggest that VAC is a newly synthesizedcompartment, rather than a compartment derived fromthe pre-existing brush border. A similar origin wasproposed (with no experimental evidence) for the brushborder vesicles observed in Davidson's disease (Cutz et al.1989).
In vivo treatment of rats with colchicine has also beenshown to induce the formation of brush border vacuoles inintestinal cells (Achler et al. 1989). The proposed mechan-ism in this case was the internalization of newlysynthesized brush borders misplaced to the basolateralsurface due to microtubular disruption and consequentmissorting of apical proteins. In our in vitro system, weclearly demonstrated, using prebiotinylated monolayers,that internalization of neither apical nor basolateralmembranes was responsible for VAC formation. Further-more, antibodies against an apical marker (sucrase-isomaltase) added continuously to the apical or basolateralmedium at the time of VAC production did not result inlabelling of VACs by the antibody. Thus, in cultured Caco-2 cells, apical components appear to be incorporated intoVACs without prior surface appearance. Another differ-ence between native intestinal cells and Caco-2 cells isthat, in vivo, VACs fused with the apical surface whereas,in vitro, VACs appear to be degraded. Perhaps, thesedifferences may be attributed to the different experimen-tal protocols utilized in vivo and in vitro (drug wasadministered to animals by stomach gavage, by theprocedure used by Achler et al. (1989) and, therefore, thelevels were constantly changing during the time of theexperiment) or, alternatively, to differences in behavior inthe response of native and cultured cells. Experimentsperformed on human tissues indicated that not onlymicrotubular disruption but also microfilament disassem-bly induced similar microvillar inclusions (Carruthers etal. 1986). In this case, collapse of the straight brush borderinto folds is believed to be the initial step leading to theformation of brush border vacuoles within the cytoplasm(Carruthers et al. 1986). Preliminary results in Caco-2cells indicated that cytochalasine D treatment cangenerate small VAC-like structures (data not shown).Their location close to the apical surface seemed to supportthis hypothesis.
Recent studies of the biogenetic pathways of plasmamembranes proteins in Caco-2 cells have shown that about10 % of several newly synthesized apical proteins failed toreach the cell surface during colchicine treatment (Gilbertet al. 1991). The results of this morphological studysuggest that these proteins are delivered to VACs orintracellular vesicles rather than to the apical surface.This is consistent with the de novo origin of the VAC weproposed. VAC formation may involve fusion betweenpost-Golgi vesicles. It is interesting to note that Caco-2VACs expressed all apical markers previously detected inthis cell line (Matter et al. 1990; Le Bivic et al. 1990;Gilbert et al. 1991), independently of whether thesemarkers follow direct (e.g. sucrase-isomaltase) or indirect(e.g. dipeptidylpeptidase IV) pathways to the cell surface.
Only 5% of colchicine-treated Caco-2 cells expressedVACs. The reason for this low percentage of cellularresponse to colchicine is unclear. One possibility is that
VAC in Caco-2 cells 457

the ability to form VACs is expressed permanently by onlya small percentage of cells, in spite of the uniformdisruption of the microtubular network caused by colchi-cine. If this is the case, it should be possible to clone 'VAC-forming cells'; we are currently attempting to do this.Alternatively, the VAC-forming ability may be a stochas-tic phenomenon for a given cell population. The avail-ability of a model system in which brush-border-contain-ing vesicles may be developed in controlled in vitroexperiments opens the way towards an understanding ofthe biogenesis of the apical domain in normal cells and inpathological states, such as Davidson's disease.
The authors thank Dr David Hanzel for assistance in LSCMand Dr Sharon K. Powell for critical reading of the manuscript.We also thank Drs Andre Le Bivic and Andrea Quaroni forproviding antibodies. This work was supported by NIH grantsGM-34017 and HL-37675, and a grant from the American CancerSociety.
References
ACHLER, C, FILMER, D., MERTE, C. AND DRENCKHAHN, D. (1989). Role ofmicrotubules in polarized delivery of apical membrane proteins to thebrush border of the intestinal epithelium. J. Cell Biol. 109, 179-189.
BARTLES, J. R., FERRACCI, H. M., STIEGER, B. AND HUBBARD, A. L.(1987). Biogenesis of the rat hepatocyte plasma membrane in vivo:comparison of the pathways taken by apical and basolateral proteinsusing subcellular fractionation. J. Cell Biol. 105, 1241-1251.
BEAULIEU, J. F., NICHOLS, B. AND QUARONI, A. (1989). Posttranslationalregulation of sucrase-isomaltase expression in intestinal crypt andvillus cells. J. biol. Chem. 264, 20000-20011.
CARRUTHERS, L., DOURMASHKIN, R. AND PHILLIPS, A. (1986). Disorders ofthe cytoskeleton of the enterocyte. Clin. Gastroenterol. 15, 105-120.
CHANTRET, I., BARBAT, A., DUSSAULX, E., BRATTAIN, M. G. ANDZWEIBAUM, A. (1988). Epithelial polarity, villin expression, andenterocytic differentiation of cultured human colon carcinoma cells: Asurvey of twenty cell lines. Cancer Res. 48, 1936-1942.
COLONY, P. C. AND NEUTRA, M. R. (1983). Epithelial differentiation inthe fetal rat colon. I. Plasma membrane phosphatase activities. DeviBiol. 97, 349-363.
CUTZ, E., RHOADS, M., DRUMM, B., SHERMAN, P. M., DURIE, P. R. ANDFORSTNER, G. G. (1989). Microvillus inclusion disease: an inheriteddefect of brush-border assembly and differentiation. New Engl. J.Med. 320, 646-651.
DAVIDSON, G. P., CUTZ, E., HAMILTON, J. R. AND GALL, D. G. (1978).Familial enteropathy: A syndrome of protracted diarrhea from birth,failure to thrive, and hypoplastic villus atrophy. Gastroenterology 75,783-790.
GILBERT, T., LE BIVIC, A., QUARONI, A. AND RODRIGUEZ-BOULAN, E.(1991). Microtubular organization and its involvement in thebiogenetic pathways of plasma membrane proteins in Caco-2intestinal epithelial cells. J. Cell Biol. 113, 275-288.
GRIFFITHS, G. AND SIMONS, K. (1986). The trans Golgi network: sortingat the exit site of the Golgi complex. Science 234, 438-443.
LE BIVIC, A., BOSC-BIERN, I. AND REGGIO, H. (1988a). Characterization ofa glycoprotein expressed on the basolateral membrane of humanintestinal epithelial cells and cultured colonic cell lines. Eur. J. CellBiol. 46, 113-120.
LE BIVIC, A., HIRN, M. AND REGGIO, H. (1987). Apical membrane markeris expressed early in colonic epithelial cells. Biol. Cell 60, 209-216.
LE BIVIC, A., HIRN, M. AND REGGIO, H. (19886). HT-29 cells are an invitro model for the generation of cell polarity in epithelia duringembryonic differentiation. Proc. natn. Acad. Sci. U.S.A. 85, 136-140.
LE BIVIC, A., QUARONI, A., NICHOLS, B. AND RODRIGUEZ-BOULAN, E.(1990). Biogenic pathways of plasma membrane proteins in Caco-2, ahuman intestinal epithelial cell line. J. Cell. Biol. I l l , 1351-1361.
MASSEY, D., FERRACCI, H., GORVEL, J. P., RIGAL, A., SOULIE, J. M. ANDMAROUX, S. (1987). Evidence for the transit of aminopeptidase N
through the basolateral membrane before it reaches the brush borderof enterocytes. J. Membr. Biol. 96, 19-25.
MATLIN, K. S. AND SIMONS, K. (1984). Sorting of an apical plasmamembrane glycoprotein occurs before it reaches the cell surface incultured epithelial cells. J. Cell Biol. 99, 2131-2139.
MATTER, K., BRAUCHBAR, M., BUCHER, K. AND HAURI, H. P. (1990).Sorting of endogenous plasma membrane proteins occurs from twosites in cultured human intestinal epithelial cells (Caco-2). Cell 60,429-437.
MATTER, K., BUCHER, K. AND HAURI, H. P. (1990). Microtubuleperturbation retards both the direct and the indirect apical pathwaybut does not affect sorting of plasma membrane proteins in intestinalepithelial cells (Caco-2). EMBO J. 9, 3163-3170.
MISEK, D. E., BARD, E. AND RODRIGUEZ-BOULAN, E. (1984). Biogenesis ofepithelial cell polarity: intracellular sorting and vectorial exocytosis ofan apical plasma membrane glycoprotein. Cell 39, 537-546.
PAVELKA, M., ELLINGER, A. AND GANGL, A. (1983). Effect of colchicine onrat small intestinal absorptive cells. J. Ultrastructure Res. 85,249-259.
PINTO, M., APPAY, M. D., SIMON-ASSMANN, P., CHEVALIER, G.,DRACOPOLI, N., FOGH, J. AND ZWEIBAUM, A. (1988). Enterocyticdifferentiation of cultured human colon cancer cells by replacement ofglucose by galactose in the medium. Cancer Res. 48, 1936-1942.
PINTO, M., ROBINE-LEON, S., APPAY, M. D., KEDINGER, M., TRIADOU, N.,DUSSAULX, E., LACROIX, B., SIMON-ASSMAN, P., HAFFEN, K., FOGH, J.AND ZWEIBAUM, A. (1983). Enterocyte-like differentiation andpolarization of the human colon carcinoma cell line Caco-2 in culture.Biol. Cell 47, 323-330.
QUARONI, A. AND ISSELBACHER, K. J. (1985). Study of intestinal celldifferentiation with monoclonal antibodies to intestinal cell surfacecomponents. Devi Biol. I l l , 267-279.
QUARONI, A., KIRSCH, K. AND WEISER, M. M. (1979). Synthesis ofmembrane glycoproteins in rat small intestinal villus cells. Biochem.J. 182, 213-221.
REMY, L. (1986). The intracellular lumen: origin, role and implicationsof a cytoplasmic neostructure. Biol. Cell 56, 97-106.
REMY, L., MARVALDI, J., RUA, S., SECCHI, J. AND LECHENE DE LA PORTE,P. (1984). The role of intracellular lumina in the repolarizationprocess of a colonic adenocarcinoma cell line. Virchows Arch. B.Zellpath. 46, 297-305.
REVEL, J. P. AND KARNOVSKY, M. J. (1967). Hexagonal array of subunitsin intercellular junctions of the mouse heart and liver. J. Cell Biol.33, C7-C12.
RINDLER, M. J., IVANOV, I. E., PLESKEN, H., RODRIGUEZ-BOULAN, E. ANDSABATINI, D. D. (1984). Viral glycoproteins destined for apical orbasolateral plasma membrane domains traverse the same Golgiapparatus during their intracellular transport in doubly infectedMadin-Darby canine kidney cells. J. Cell Biol. 98, 1304-1319.
RODRIGUEZ-BOULAN, E. AND NELSON, W. J. (1989). Morphogenesis of thepolarized epithelial cell phenotype. Science 245, 718-725.
SARGIACOMO, M., LISANTI, M., GRAEVE, L., LE BIVIC, A. AND RODRIGUEZ-BOULAN, E. (1989). Integral and peripheral protein compositions of theapical and basolateral membrane domains in MDCK cells. J. Membr.Biol. 107, 277-286.
SIMONS, K. AND FULLER, S. D. (1985). Cell surface polarity in epithelia.A. Rev. Cell Biol. 1, 243-288.
VEGA SALAS, D. E., SALAS, P. J. AND RODRIGUEZ-BOULAN, E. (1987).Modulation of the expression of an apical plasma membrane proteinof Madin-Darby canine kidney epithelial cells: cell-cell interactionscontrol the appearance of a novel intracellular storage compartment.J. Cell Biol. 104, 1249-1259.
WANDINGER-NESS, A., BENNETT, M. K., ANTONY, C. AND SIMONS, K.(1990). Distinct transport vesicles mediate the delivery of plasmamembrane proteins to the apical and basolateral domains of MDCKcells. J. Cell Biol. I l l , 987-1000.
ZWEIBAUM, A., PINTO, M., CHEVALIER, G., DUSSAULX, E., TRIADOU, N.,LACROIX, B., HAFFEN, K., BRUN, J. L. AND ROUSSET, M. (1985).Enterocytic differentiation of a subpopulation of the human colontumor cell line HT-29 selected for growth in sugar-free medium andits inhibition by glucose. J. cell. Physiol. 122, 21-29.
(Received 26 June 1991 - Accepted 12 August 1991)
458 T. Gilbert and E. Rodriguez-Boulan





![Vacuolar Transporters – Companions on a Longtime … · Vacuolar Transporters – Companions on a Longtime Journey[OPEN] Enrico Martinoia1 Department of Plant and Microbial Biology,](https://img.dokumen.tips/doc/110x75/603fbba48d3fd353b308f80e/vacuolar-transporters-a-companions-on-a-longtime-vacuolar-transporters-a-companions.jpg)























