Embed Size (px)
Citation preview

Pharmacology & Therapeutics xxx (2014) xxx–xxx
JPT-06669; No of Pages 7
Contents lists available at ScienceDirect
Pharmacology & Therapeutics
j ourna l homepage: www.e lsev ie r .com/ locate /pharmthera
Associate editor: O. Binah
Induced pluripotent stem cell-derived cardiomyocytes for drugdevelopment and toxicity testing
Daniel Sinnecker a, Karl-Ludwig Laugwitz a,b, Alessandra Moretti a,b,⁎a Klinikum rechts der Isar— Technische Universität München, I. Medical Department— Cardiology, Ismaninger Strasse 22, 81675 Munich, Germanyb DZHK (German Centre for Cardiovascular Research) — Partner Site, Munich Heart Alliance, Munich, Germany
⁎ Corresponding author at: Klinikum rechts der Isar— TI. Medical Department — Cardiology, Ismaninger StrasseTel.: +49 89 41406907; fax: +49 89 41404900.
E-mail address: [email protected] (A. Moretti).
http://dx.doi.org/10.1016/j.pharmthera.2014.03.0040163-7258/© 2014 Elsevier Inc. All rights reserved.
Please cite this article as: Sinnecker, D., et alPharmacology & Therapeutics (2014), http://
a b s t r a c t
a r t i c l e i n f oKeywords:
iPSCDrug screeningHigh-throughput screeningQT prolongationDrug toxicityInduced pluripotent stem cell (iPSC) technology is creating exciting new opportunities for cardiovascular re-search by providing platforms to study themechanisms of disease pathogenesis that could lead to new therapiesor reveal drug sensitivities. In this review, the potential usefulness of iPSC-derived cardiomyocytes in drug devel-opment as well as in drug toxicity testing is discussed, with a focus on the achievements that have been alreadymade in this regard. Moreover, the crucial steps that have to be taken before this technology can be broadly usedin drug discovery and toxicology assessments are highlighted.
© 2014 Elsevier Inc. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 02. Possible applications of induced pluripotent stem cell-derived cardiomyocytes in drug development and toxicity testing . 03. Generation of induced pluripotent stem cell-derived cardiomyocytes — considerations on reprogramming and
differentiation protocols . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 04. Pharmacology of induced pluripotent stem cell-derived cardiomyocytes — lessons from disease modeling studies . 05. Exploiting the potential of induced pluripotent stem cell-derived cardiomyocytes for pharmacological and
toxicological screening — phenotype-based assays . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 06. Critical prospect . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Conflict of interest statement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 01. Introduction
The discovery that somatic cells can be reprogrammed to pluripo-tent stem cells (induced pluripotent stem cells, iPSC; Takahashi &Yamanaka, 2006), capable of differentiating to all cell types present inthe adult organism, and especially the rapid adaptation of the technolo-gy to human cells (Takahashi et al., 2007; Yu et al., 2007) has generatedlarge expectations concerning the possible applications. The technologyhas a particularly strong appeal for disciplines such as cardiovascularmedicine, which deal with cell types (e.g., cardiomyocytes) that cannotbe easily obtained from human probands or patients. Among the
echnischeUniversitätMünchen,22, 81675 Munich, Germany.
., Induced pluripotent stem cedx.doi.org/10.1016/j.pharmth
possible applications of iPSC technology in the cardiovascular field, thepotential usefulness in drug development as well as in drug toxicitytesting has been already highlighted in the initial reports on the gener-ation of human iPSC (Takahashi et al., 2007; Yu et al., 2007). The aim ofthis review is to define the possible role of iPSC-derived cardiomyocytesin this context, to point out the achievements that have been alreadymade in this regard, and to discuss the crucial steps that have to betaken before this technology can be broadly used in drug developmentand toxicity testing.
2. Possible applications of induced pluripotent stem cell-derivedcardiomyocytes in drug development and toxicity testing
The identification and characterization of potential drug targets, thescreening of compound libraries for drugs with a desired effect, as well
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004
2 D. Sinnecker et al. / Pharmacology & Therapeutics xxx (2014) xxx–xxx
as the evaluation of drug candidates for possible adverse effectsall require reliable test systems. Such test systems can beengineered based on primary cells, immortalized cell lines, or ani-mal models; however, cardiovascular pharmacology suffers fromseveral drawbacks of the currently-used test systems based oncardiomyocytes.
Primary human cardiomyocytes are not easily obtained and cannotbe kept in culture for prolonged time periods or expanded in vitro. Im-mortalized human cardiomyocyte cell lines that faithfullymodel impor-tant aspects of cardiac physiology such as action potentials are notavailable. Alternatively, human cell cultures derived from embryonicsources, such as human embryonic kidney (HEK) lines, can be used togenerate overexpression systems of the potential drug target molecule(Goineau et al., 2012). This allows studying the effects of a drug on aspecific gene or molecular mechanism, but fails to provide informationon the compound's overall cellular (cardiomyocyte) outcome. Thus,currently, much of the research in this field depends on animal models.For example, genetically-modifiedmice are frequently used to study thephysiology that underlies humanheart disease. However, species differ-ences are a relevant problem if cardiomyocytes from laboratory animalsare used to model features of human cardiovascular disorders. For ex-ample, murine hearts beat about 6–10 times faster than their humancounterparts, with markedly shorter action potentials that are shapedby different ion channels. In humans, mutations of the KCNQ1 geneencoding the ion channel responsible for the IKs current may cause thelong-QT syndrome type 1, a disease characterized by a prolonged QT in-terval in the ECG and a susceptibility to potentially lethal arrhythmias(Lehnart et al., 2007). However, genetic ablation of KCNQ1 in mice didnot consistently lead to a cardiac phenotype in accordance with thatof long-QT syndrome patients (Lee et al., 2000; Casimiro et al., 2001).
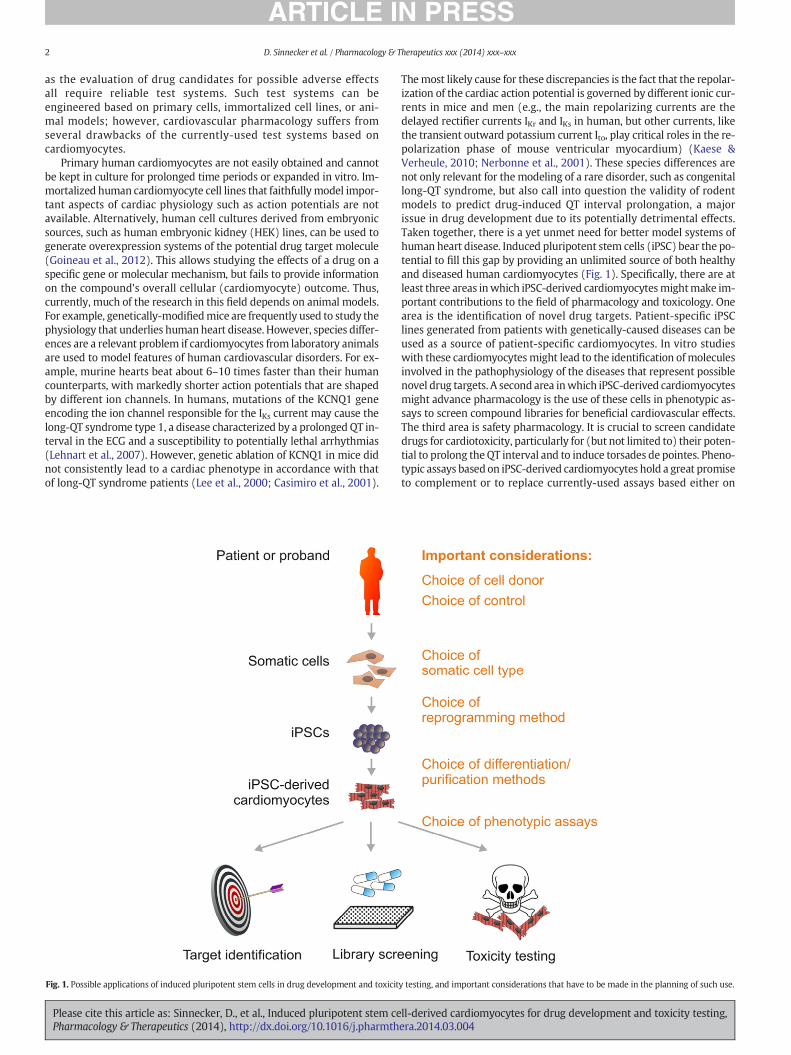
Fig. 1. Possible applications of induced pluripotent stem cells in drug development and toxicit
Please cite this article as: Sinnecker, D., et al., Induced pluripotent stem cePharmacology & Therapeutics (2014), http://dx.doi.org/10.1016/j.pharmth
Themost likely cause for these discrepancies is the fact that the repolar-ization of the cardiac action potential is governed by different ionic cur-rents in mice and men (e.g., the main repolarizing currents are thedelayed rectifier currents IKr and IKs in human, but other currents, likethe transient outward potassium current Ito, play critical roles in the re-polarization phase of mouse ventricular myocardium) (Kaese &Verheule, 2010; Nerbonne et al., 2001). These species differences arenot only relevant for themodeling of a rare disorder, such as congenitallong-QT syndrome, but also call into question the validity of rodentmodels to predict drug-induced QT interval prolongation, a majorissue in drug development due to its potentially detrimental effects.Taken together, there is a yet unmet need for better model systems ofhuman heart disease. Induced pluripotent stem cells (iPSC) bear the po-tential to fill this gap by providing an unlimited source of both healthyand diseased human cardiomyocytes (Fig. 1). Specifically, there are atleast three areas inwhich iPSC-derived cardiomyocytesmightmake im-portant contributions to the field of pharmacology and toxicology. Onearea is the identification of novel drug targets. Patient-specific iPSClines generated from patients with genetically-caused diseases can beused as a source of patient-specific cardiomyocytes. In vitro studieswith these cardiomyocytesmight lead to the identification ofmoleculesinvolved in the pathophysiology of the diseases that represent possiblenovel drug targets. A second area inwhich iPSC-derived cardiomyocytesmight advance pharmacology is the use of these cells in phenotypic as-says to screen compound libraries for beneficial cardiovascular effects.The third area is safety pharmacology. It is crucial to screen candidatedrugs for cardiotoxicity, particularly for (but not limited to) their poten-tial to prolong theQT interval and to induce torsades de pointes. Pheno-typic assays based on iPSC-derived cardiomyocytes hold a great promiseto complement or to replace currently-used assays based either on
y testing, and important considerations that have to be made in the planning of such use.
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004

3D. Sinnecker et al. / Pharmacology & Therapeutics xxx (2014) xxx–xxx
primary cardiomyocytes from laboratory animals or on cell lines over-expressing ion channels.
3. Generation of induced pluripotent stem cellerivedcardiomyocytes — considerations on reprogramming anddifferentiation protocols
3.1. Reprogramming
The first reportedmethod for the generation of iPSC used integratingretroviruses to deliver four transcription factors (Oct3/4, Sox2, c-Mycand Klf-4) to fibroblasts (Takahashi & Yamanaka, 2006). Since then,rapid progress has been made in reprogramming various somatic celltypes usingmany variants of the original cocktail of reprogramming fac-tors, including chromatin-modifying small molecules. Moreover, nu-merous methods for delivering the reprogramming factors to the cells,such as non-integrating genomic approaches (Gonzalez et al., 2011)have been applied. Consequently, it is now possible to use easily acces-sible somatic cells from a patient (e.g., skin fibroblasts, hair follicles, orwhole blood) to generate iPSC lines that are free from viral transgenesand genetically identical to the patient (Jia et al., 2010; Seki et al.,2010; Yakubov et al., 2010; Narsinh et al., 2011).
3.2. Differentiation
To use iPSC-derived cardiomyocytes for drug development or safetypharmacology, the availability of reliable protocols to differentiate iPSCto cardiomyocytes is crucial. The major aim of cardiac differentiationprotocols is to provide the differentiating cells with the specific cuesnecessary to direct their development towards a cardiomyocytic fate.Therefore, it is important to understand the regulation of cardiac differ-entiation, an area in which substantial progress has been made duringthe past decade (Laflamme et al., 2007; Yang et al., 2008; Kattmanet al., 2011; Burridge et al., 2012). Differentiation of iPSC to cardiovascu-lar cells is a multistep process tightly regulated by developmental sig-nals, epigenetic programs and extracellular microenvironments(reviewed in Moretti et al., 2013). This process involves diverse path-ways such as BMP, TGF-b/activin/nodal,WNT/b-catenin, and FGF signal-ing. Developments in order to more efficiently direct the developingcells towards the cardiomyocyte lineage include co-culture with END-2 stromal cells (Mummery et al., 2003) and ascorbic acid, which en-hances collagen synthesis and promotes proliferation of cardiac progen-itors (Cao et al., 2012). Microenvironmental factors that are potentiallyimportant for cardiac differentiation are not limited to paracrine factors,but may also include other factors present during embryonic heart de-velopment, such as mechanical stretch and rhythmic electric activity.Consequently, biomatrices with defined elastic properties (Engleret al., 2008) and electrical stimulation (Nunes et al., 2013) have been ap-plied to promote cardiac differentiation of pluripotent stem cell-derivedcardiomyocytes.
3.3. Maturity of induced pluripotent stem cell-derived cardiomyocytes
A key problem so far unresolved is posed by the fact thatcardiomyocytes generated frompluripotent stemcellswith the current-ly available protocols are immature compared to their adult counter-parts. In many aspects, the cells are more similar to fetal than to adultcardiomyocytes. Morphologically, the cells lack a fully-developed trans-verse tubule system (Lieu et al., 2009). Functionally, the cells are fre-quently characterized by spontaneous contractions, which are notfound in adult ventricular cardiomyocytes. The maximum diastolicmembrane potential is less negative than that in adult cardiomyocytes,and the action potential upstroke velocities and amplitudes are similarto those of the 10-week-old embryonic hearts (Mummery et al.,2003). Conflicting data exist regarding thematurity of the calcium han-dling system in pluripotent stem cell-derived cardiomyocytes, although
Please cite this article as: Sinnecker, D., et al., Induced pluripotent stem cePharmacology & Therapeutics (2014), http://dx.doi.org/10.1016/j.pharmth
there is evidence that at least basic components of the calcium cyclingmachinery and excitation–contraction coupling are functional (Satinet al., 2008; Itzhaki et al., 2011b; Jung et al., 2012; Novak et al., 2012).The transcriptional profiles of iPSC-derived cardiomyocytes are alsosimilar to those of fetal cardiomyocytes (Gupta et al., 2010).
3.4. Heterogeneity of induced pluripotent stem cell-derived cardiomyocytes
The cardiomyocytes generated by current differentiation protocolsare a mixture of cells belonging to all three major cardiomyocyte sub-types: cells with atrial-, ventricular- and nodal-like phenotypes. Whilethis can be considered an advantage due to the possibility to assessphysiological properties in all these cell types, it also holds thedisadvan-tage that changes that occur only in one subpopulation of cells may bediluted if the readout is taken from all cells. Particularly, this problemis likely to arise in assays that do not record the action potential of singlecells, which is the most straightforward method to classify each cell asatrial-, ventricular- or nodal-like. It is thus important to understandthe mechanisms of cardiac subtype specification, and significant effortshave been made to develop protocols that reduce heterogeneity ofhuman pluripotent stem cell-derived cardiomyocytes. For example, in-hibition of NRG-1b/ERBB signaling has been shown to enhance the pro-portion of nodal-like cells (Zhu et al., 2010), and retinoid signalsenhance atrial versus ventricular specification during cardiac hESC dif-ferentiation (Zhang et al., 2011).
3.5. Purification of induced pluripotent stem cell-derived cardiomyocytes
Especially for high-throughput applications, it is essential to be ableto generate populations of iPSC-derived cardiomyocytes or even cardio-myocyte subtypes of a high purity. Consequently, many research effortshave been undertaken in order to reach this goal. The identification ofmarkers expressed specifically on cardiomyocytes, including EMILIN2(Van Hoof et al., 2010), SIRPA (Dubois et al., 2011; Elliott et al., 2011)and VCAM (Elliott et al., 2011; Uosaki et al., 2011) has made it possibleto isolate highly enriched populations of these cells from pluripotentstem cells by FACS or magnetic bead sorting. However, specific surfacemarkers for iPSC-derived cardiomyocyte subtypes (ventricular-, atrial-and nodal-like cells) have not yet been discovered. Meanwhile, theintroduction of ectopic reporters or the targeting of endogenous loci isbecoming a very useful tool for cell-type-specific lineage marking.Selectable markers, such as fluorescent markers or antibiotic resistancegenes, under the control of lineage-specific promoters allow forpurification of precise cell populations by either FACS or selectionwith antibiotics. Using this approach, the human α-myosin heavy-chain (α-MHC) promoter coupled to a bicistronic reporter (GFP and pu-romycin) has been used to achieve high purity of cardiomyocytes (90%)from differentiating human embryonic stem cells (Anderson et al.,2007). Ventricular-like cardiomyocytes can be marked by lentiviral de-livery of a selectable human myosin light chain-2v (MLC2v) enhancerdriving EGFP expression (Huber et al., 2007). Generation of reportersfor the other cardiomyocyte subtypes has been so far hampered bythe fact that, in contrast to adult cardiomyocytes, iPSC-derived cardio-myocyte subtypes are not explicitly characterized by subtype-specificexpression of proteins, such as the atrial isoform of MLC2 (MLC2a) foratrial or the hyperpolarization-activated cyclic nucleotide-gated potas-sium channel 4 (HCN4) for conduction system cells.
4. Pharmacology of induced pluripotent stem cell-derivedcardiomyocytes — lessons from disease modeling studies
Soon after the initial description of human iPSC, the first studiesaimed at modeling cardiac diseases with patient-specific stem cellshave been published. A summary of these disease modeling studies isprovided in Table 1. The first diseases that have been investigatedusing this approach weremonogenic channelopathies, such as different
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004

Table 1Modeling heart disease with patient-specific iPSC.
Disease Reference Genetic basis Tested drugs
Long-QT syndromeLQT1 Moretti et al., 2010 KCNQ1 c.569GNA; p.R190Q Propranolol, isoprenaline
Egashira et al., 2012 KCNQ1 c.1893delC; p.P631fs/33LQT2 Itzhaki et al., 2011a KCNH2 c.1841CNT; p.A614V Nifedipine, pinacidil, ranolazine
Matsa et al., 2011 KCNH2 c.1681GNA; p.A561T Propranolol, isoprenaline, nicorandil, PD-118057,Matsa et al., 2013 KCNH2 c.1681GNA; p.A561T Allele-specific siRNALahti et al., 2012 KCNH2 c.526CNT; p.R176W Erythromycin, sotalol, cisapride
LQT3, Brugada Syndrome Davis et al., 2012 SCN5A c.5387_5389ins TGA; p.1795insDLQT8, Timothy Syndrome Yazawa et al., 2011 CACNA1C c.1216GNA; p.G406R Roscovitine
Catecholaminergic polymorphic ventricular tachycardiaCPVT1 Fatima et al., 2011 RYR2 c.7447TNA; p.F2483I Isoprenaline, forskolin
Jung et al., 2012 RYR2 c.1217CNT; p.S406L DantroleneItzhaki et al., 2012 RYR2 c.12056TNG; p.M4109R Flecainide, thapsigargin
CPVT2 Novak et al., 2012 CASQ2 c.1183GNC; p.D307H
CardiomyopathiesDCM Sun et al., 2012 TNTT2 p.R173W Metoprolol, norepinephrineHCM Lan et al., 2013 MYH7 c.1988GNA; p.R663H Propranolol, verapamil, nifedipin, diltiazem, lidocaine,
mexiletine, ranolazineLEOPARD syndrome Carvajal-Vergara et al., 2010 PTPN11 c.140CNT; p.T468MARVC Ma et al., 2013 PKP2 c.1841TNC; p.L614P Nifedipine, isoprenaline
Kim et al., 2013 PKP2 c.2484CNT; r2483_2489del c.2013delC; p.R672fsX683Caspi et al., 2013 PKP2 c.972InsT/N; p.A324fs335X PKP2 c.148_151delACAG/N;
p.T50SfsX110Isoproterenol, carbamylcholine, BIO
4 D. Sinnecker et al. / Pharmacology & Therapeutics xxx (2014) xxx–xxx
subtypes of long-QT syndrome or catecholaminergic polymorphic ven-tricular tachycardia (CPVT). The cell-autonomous pathophysiology ofthese disorders allows the investigation of disease-specific phenotypesin single cells. Accordingly, single-cell methodologies such as patchclamp electrophysiology, fluorometric calcium imaging using calcium-sensitive dyes and single cell RT-PCR were used in these studies. Mostof these studies have already investigated the effect of drugs onthe patient-specific iPSC-derived cardiomyocytes (Table 1). Drugs al-ready known to have an effect on the disease phenotype in patients(e.g., beta blockers in long-QT syndrome) were used, mainly to demon-strate that the in vitro disease models recapitulate key features of thedisease. However, in some studies, novel pharmacological conceptshave been evaluated. For example, it was shown that dantrolene, adrug clinically used for the treatment of malignant hyperthermia, wasable to revert the arrhythmogenic phenotype in cardiomyocytes affect-ed by CPVT caused by a mutation of the cardiac ryanodine receptor cal-cium channel (Jung et al., 2012).
Patient-specific iPSC-derived cardiomyocytes have been even usedto investigate a novel RNA-based treatment strategy for long-QT syn-drome (Matsa et al., 2013; Sinnecker et al., 2013). In cardiomyocytes af-fected by long-QT syndrome type 2 caused by an A516Tmutation in theKCNH2 gene encoding the hERG potassium channel, it was shown thatthe mutation exerted a dominant-negative effect on the hERG-mediated IKr current: A hERG potassium channel is formed by fourKCNH2 subunits. In patient cardiomyocytes, which are heterozygousfor the A516T mutation, wild type and mutated KCNH2 subunits arepresumably synthetized in equal amounts. The mutated subunits areable to co-assemble withwild type subunits, but the resulting ion chan-nel tetramers are not correctly incorporated into the plasmamembraneif they contain one or more mutated subunits. Consequently, in patientcardiomyocytes, the IKr current is reduced not only to 50% (whichwouldindicate a haploinsufficiency mechanism of the mutation), but to lessthan 10%, leading to a marked prolongation of the action potential. Tak-ing advantage of this dominant-negativemechanism, the authors of thestudy designed an allele-specific small interfering RNA that specificallyled to a knockdown of the mutated, but not of the wild type KCNH2mRNA. This strategy reduced the amount of mutated KCNH2 subunitsinterfering with membrane trafficking of the wild type subunits,
Please cite this article as: Sinnecker, D., et al., Induced pluripotent stem cePharmacology & Therapeutics (2014), http://dx.doi.org/10.1016/j.pharmth
increasing the IKr level and thereby significantly reducing the action po-tential duration.
4.1. Choice of control lines
An important consideration that has to be made in the planning ofsuch disease modeling studies is the choice of the experimental control.In many of the early studies (e.g., Moretti et al., 2010; Itzhaki et al.2011a; Fatima et al., 2011; Yazawa et al., 2011), control cells werecardiomyocytes generated from iPSC lines obtained from unrelatedhealthy donors. While this approach is straightforward, it bears therisk that phenotypic differences between patient and control cells donot arise from the disease-causing mutation, but by differences in thegenetic background. This problem becomes especially relevant whendiseases are studied in which the suspected phenotypic effect of thedisease-causing mutation is weak, or in which the suspected effect ofthe genetic background is strong (e.g., diseases known for incompletepenetrance). Also in cases in which the evidence for the causation of aphenotype by a specific mutation is weak (e.g., in cases of newly-identifiedmutations in genes not implicated before in a specific pathol-ogy), it is important to rule out genetic background variability as thecause of the phenotype observed in iPSC-derived cardiomyocytes.
To reduce the genetic background variability between patient andcontrol cells, iPSC lines generated from family members of the patientunaffected by the disease have been used (Carvajal-Vergara et al.,2010; Sun et al., 2012; Lan et al., 2013). However, it should be notedthat this approach can only limit, but not abolish the genetic back-ground variability, given that even siblings or parent–child pairs onlyhave 50% genetic overlap.
A promising approach to control background variability is to genet-ically modify pluripotent cell lines by means of gene targeting (Bellinet al., 2013) to generate pairs of isogenic lines that genetically differonly in themutation of interest. Such amodification can consist of eithercorrecting the disease-causingmutation in a patient-derived cell line or,as a complementary approach, of engineering a known disease-causingmutation into a cell line derived from a healthy person. With this ap-proach, even subtle differences in action potential duration between
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004

5D. Sinnecker et al. / Pharmacology & Therapeutics xxx (2014) xxx–xxx
healthy and diseased cells could be reliably measured and unequivocal-ly attributed to a KCNH2mutation (Bellin et al., 2013).
5. Exploiting the potential of induced pluripotent stem cell-derivedcardiomyocytes for pharmacological andtoxicological screening — phenotype-based assays
In contrast to other systems frequently used in drug development,such as cell lines overexpressing specific ion channels, iPSC-derivedcardiomyocytes bear the potential to be used in phenotype-based as-says. In such assays, the readout is not drug effect on a known targetstructure (e.g., the currentmediated by a specific ion channel), but rath-er a complex phenotype, such as the beating rate, action potential dura-tion, or the occurrence of arrhythmias. An important advantage of suchphenotypic assays is that it is possible to evaluate the effect of drugs thatdo not interact with known target molecules. For example, the cardiacaction potential is shaped by the complex gating behavior of morethan a handful of ion channels, many of which are composed of severalsubunits encoded by different genes. While the effects of a compoundon a single ion channel gating can be studied in immortalized celllines, such assays may not always answer the question which effectthe compound will have on a cardiomyocyte. Many drugs that prolongthe QT interval do so by inhibiting hERG activity, which can be assessedusing hERG-overexpressing cell lines; other drugs (e.g., alfuzosin(Lacerda et al., 2008)) do so by a different mechanism of action. TheQT-prolonging (and thus hazardous) potential of these drugs is there-fore likely to be overlooked if relying solely on a hERG assay. On theother hand, there are drugs, such as verapamil, that block hERG indoses near to therapeutic plasma concentrations (Redfern et al.,2003), but do not prolong the QT interval in patients (Nademanee &Singh, 1988). Thus, similarly-behaving novel compounds might notenter the clinic nowadays due to being sorted out early in the drug de-velopment process due to their action on hERG.
Assays that measure the action potential duration in iPSC-derivedcardiomyocytes might provide a solution to these two problems. Thishas been demonstrated by measuring action potential durations in thewhole cell patch clamp configuration in iPSC-derived cardiomyocytes(Liang et al., 2013): Alfuzosin, but not verapamil significantly prolongedaction potential duration in therapeutic concentrations, in contrast tothe effects on hERG elicited by the two drugs. These observationscould be also reproduced in an assay that used microelectrode arrays(MEA) tomeasure the field potential duration,which is closely correlat-ed to the action potential duration of single cardiomyocytes (Navarreteet al., 2013). The use ofMEA instead of single-cell patch clampmeasure-ments is an important step towards increasing the throughput of suchassays.
To bring the potential of iPSC-derived cardiomyocytes for pharma-cological and toxicological screening to fruition, the incorporation ofthese cells into reliable assays that can be scaled up to medium- orhigh-throughput applications would be desirable. Several attempts toreach this goal have been already made. Drug-induced cardiotoxicitycan manifest as cardiomyocyte cell death. Using high-content automat-ed microscopy in a 96-well format in conjunction with live cell stainingas well as immunofluorescence, the effect of drugs on a panel of celldeath-related abnormalities, such as nuclear shape change and frag-mentation, DNA degradation, caspase activation, mitochondrial outermembrane permeabilization, and cell detachment was assessed iniPSC-derived cardiomyocytes (Mioulane et al., 2012). This approachbears the possibility to obtain specific cardio-toxicity profiles of drugcandidates in vitro, which might be then compared to those of sub-stances with proven cardiotoxic potential, such as doxorubicin.
Another step towards high-throughput application of iPSC-derivedcardiomyocytes is an assay thatmeasures the beating rate based on cal-cium transients imagedwith fluorescent indicators, which has been im-plemented in a 96- and 384-well format (Sirenko et al., 2013). Byapplying drugs in different concentrations using automated liquid
Please cite this article as: Sinnecker, D., et al., Induced pluripotent stem cePharmacology & Therapeutics (2014), http://dx.doi.org/10.1016/j.pharmth
handling, dose–response curves for several drugs that affect the beatingrate could be recorded with good reproducibility. Moreover, the assayalso allows for the analysis of arrhythmias based on morphologicalchanges of the calcium transients such as prolongations that werefound with some drugs known to prolong the QT interval. However, ithas not been systematically investigated whether such alterations in-deed correlate with established measures of arrhythmogenesis, suchas action potential prolongation, early afterdepolarizations, or triggeredactivity.
A phenotypic assay for drugs interfering with cardiac hypertrophyhas been also adapted for use in a 384-well format (Carlson et al.,2013). Treatment of human iPSC-derived cardiomyocytes withendothelin-1 reproducibly led to actin remodeling, an increase in cellsize and increased expression of brain natriuretic peptide (BNP). Theendothelin-induced increase in intracellular BNPwas imaged by immu-nofluorescence using automated microscopy and image analysis. Thistechnology allowed the investigation of the dose-dependent effect ofseveral drugs on this surrogate parameter for the cardiac hypertrophicresponse.
6. Critical prospect
It is widely recognized that iPSC-derived cardiomyocytes have ahuge potential to advance drug development and toxicity testing. How-ever, in order to fulfill this potential, several important issues will haveto be resolved. Standardization of methods for iPSC generation, cardiacdifferentiation and quality control will be necessary if these cells areto be incorporated into standardized assays. The commercial availabilityof well-characterized iPSC-derived cardiomyocytes (Ma et al., 2011)might represent one important step towards this goal. The advance-ment of differentiation protocols in order to generate cells that aremore similar to adult cardiomyocytes is another important step. De-pending on the planned application, the immature phenotype ofcardiomyocytes generated with current protocols might represent amore or less relevant problem. The most important question that hasto be answered for each proposed assay is to which extent the pheno-type observed in iPSC-derived cardiomyocytes treated with a specificdrug correlates with the clinical findings in patients treated with thesame drug. This will have to be systematically investigated – by testingasmany drugs as possible – before the usefulness of the assay can be re-liably predicted.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Acknowledgments
The authors would like to acknowledge and thank the German Re-search Foundation, the German Ministry for Education and Research,and the European Research Council for their ongoing support for the re-search. KLL and AM also acknowledge the Munich Heart Alliance, amember of the German Centre for Cardiovascular Research.
References
Anderson, D., Self, T., Mellor, I. R., Goh, G., Hill, S. J., & Denning, C. (2007). Transgenicenrichment of cardiomyocytes from human embryonic stem cells. Mol Ther 15,2027–2036.
Bellin, M., Casini, S., Davis, R. P., D'Aniello, C., Haas, J., Ward-van Oostwaard, D., et al.(2013). Isogenic human pluripotent stem cell pairs reveal the role of a KCNH2 muta-tion in long-QT syndrome. EMBO J 32, 3161–3175.
Burridge, P. W., Keller, G., Gold, J.D., & Wu, J. C. (2012). Production of denovo cardiomyocytes: Human pluripotent stem cell differentiation and directreprogramming. Cell Stem Cell 10, 16–28.
Cao, N., Liu, Z., Chen, Z., Wang, J., Chen, T., Zhao, X., et al. (2012). Ascorbic acid enhancesthe cardiac differentiation of induced pluripotent stem cells through promoting theproliferation of cardiac progenitor cells. Cell Res 22, 219–236.
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004

6 D. Sinnecker et al. / Pharmacology & Therapeutics xxx (2014) xxx–xxx
Carlson, C., Koonce, C., Aoyama, N., Sinhorn, S., Fiene, S., Thompson, A., et al. (2013). Phe-notypic screening with human iPS cell-derived cardiomyocytes: HTS-compatible as-says for interrogating cardiac hypertrophy. J Biomol Screen 18, 1203–1211.
Carvajal-Vergara, X., Sevilla, A., D'Souza, S. L., Ang, Y. S., Schaniel, C., Lee, D. F., et al. (2010).Patient-specific induced pluripotent stem-cell-derived models of LEOPARD syn-drome. Nature 465, 808–812.
Casimiro, M. C., Knollmann, B. C., Ebert, S. N., Vary, J. C., Greene, A. E., Franz, M. R., et al.(2001). Targeted disruption of the Kcnq1 gene produces a mouse model of Jervelland Lange–Nielsen syndrome. Proc Natl Acad Sci U S A 98, 2526–2531.
Caspi, O., Huber, I., Gepstein, A., Arbel, G., Maizels, L., Boulos, M., et al. (2013). Modeling ofarrhythmogenic right ventricular cardiomyopathy with human induced pluripotentstem cells. Circ Cardiovasc Genet 6, 557–568.
Davis, R. P., Casini, S., van den Berg, C. W., Hoekstra, M., Remme, C. A., Dambrot, C., et al.(2012). Cardiomyocytes derived from pluripotent stem cells recapitulate electro-physiological characteristics of an overlap syndrome of cardiac sodium channel dis-ease. Circulation 125, 3079–3091.
Dubois, N. C., Craft, A.M., Sharma, P., Elliott, D. A., Stanley, E. G., Elefanty, A. G., et al. (2011).SIRPA is a specific cell-surface marker for isolating cardiomyocytes derived fromhuman pluripotent stem cells. Nat Biotechnol 29, 1011–1018.
Egashira, T., Yuasa, S., Suzuki, T., Aizawa, Y., Yamakawa, H., Matsuhashi, T., et al. (2012).Disease characterization using LQTS-specific induced pluripotent stem cells.Cardiovasc Res 95, 419–429.
Elliott, D. A., Braam, S. R., Koutsis, K., Ng, E. S., Jenny, R., Lagerqvist, E. L., et al. (2011).NKX2-5(eGFP/w) hESCs for isolation of human cardiac progenitors andcardiomyocytes. Nat Methods 8, 1037–1040.
Engler, A. J., Carag-Krieger, C., Johnson, C. P., Raab, M., Tang, H. Y., Speicher, D. W., et al.(2008). Embryonic cardiomyocytes beat best on a matrix with heart-like elasticity:Scar-like rigidity inhibits beating. J Cell Sci 121, 3794–3802.
Fatima, A., Xu, G., Shao, K., Papadopoulos, S., Lehmann, M., Arnaiz-Cot, J. J., et al. (2011). Invitromodeling of ryanodine receptor 2 dysfunction using human induced pluripotentstem cells. Cell Physiol Biochem 28, 579–592.
Goineau, S., Legrand, C., & Froget, G. (2012). Whole-cell configuration of the patch-clamptechnique in the hERG channel assay to predict the ability of a compound to prolongQT interval. Curr Protoc Pharmacol 10, 10.15. http://dx.doi.org/10.1002/0471141755.ph1015s57.
Gonzalez, F., Boue, S., & Izpisua Belmonte, J. C. (2011). Methods for making induced plu-ripotent stem cells: Reprogramming a la carte. Nat Rev Genet 12, 231–242.
Gupta, M. K., Illich, D. J., Gaarz, A., Matzkies, M., Nguemo, F., Pfannkuche, K., et al. (2010).Global transcriptional profiles of beating clusters derived from human induced plu-ripotent stem cells and embryonic stem cells are highly similar. BMC Dev Biol 10, 98.
Huber, I., Itzhaki, I., Caspi, O., Arbel, G., Tzukerman, M., Gepstein, A., et al. (2007). Identi-fication and selection of cardiomyocytes during human embryonic stem cell differen-tiation. FASEB J 21, 2551–2563.
Itzhaki, I., Maizels, L., Huber, I., Gepstein, A., Arbel, G., Caspi, O., et al. (2012). Modeling ofcatecholaminergic polymorphic ventricular tachycardia with patient-specific humaninduced pluripotent stem cells. J Am Coll Cardiol 60, 990–1000.
Itzhaki, I., Maizels, L., Huber, I., Zwi-Dantsis, L., Caspi, O., Winterstern, A., et al. (2011a).Modelling the long QT syndrome with induced pluripotent stem cells. Nature 471,225–229.
Itzhaki, I., Rapoport, S., Huber, I., Mizrahi, I., Zwi-Dantsis, L., Arbel, G., et al. (2011b). Calci-um handling in human induced pluripotent stem cell derived cardiomyocytes. PLoSOne 6, e18037.
Jia, F., Wilson, K. D., Sun, N., Gupta, D.M., Huang, M., Li, Z., et al. (2010). A nonviralminicircle vector for deriving human iPS cells. Nat Methods 7, 197–199.
Jung, C. B., Moretti, A., Mederos y Schnitzler, M., Iop, L., Storch, U., Bellin, M., et al. (2012).Dantrolene rescues arrhythmogenic RYR2 defect in a patient-specific stem cell modelof catecholaminergic polymorphic ventricular tachycardia. EMBO Mol Med 4, 180–191.
Kaese, S., & Verheule, S. (2010). Cardiac electrophysiology in mice: A matter of size. FrontPhysiol 3. http://dx.doi.org/10.3389/fphys.2012.00345.
Kattman, S. J., Witty, A.D., Gagliardi, M., Dubois, N. C., Niapour, M., Hotta, A., et al. (2011).Stage-specific optimization of activin/nodal and BMP signaling promotes cardiac dif-ferentiation of mouse and human pluripotent stem cell lines. Cell Stem Cell 8, 228–240.
Kim, C., Wong, J., Wen, J., Wang, S., Wang, C., Spiering, S., et al. (2013). Studying arrhyth-mogenic right ventricular dysplasia with patient-specific iPSCs. Nature 494, 105–110.
Lacerda, A. E., Kuryshev, Y. A., Chen, Y., Renganathan, M., Eng, H., Danthi, S. J., et al. (2008).Alfuzosin delays cardiac repolarization by a novel mechanism. J Pharmacol Exp Ther324, 427–433.
Laflamme, M.A., Chen, K. Y., Naumova, A. V., Muskheli, V., Fugate, J. A., Dupras, S. K., et al.(2007). Cardiomyocytes derived from human embryonic stem cells in pro-survivalfactors enhance function of infarcted rat hearts. Nat Biotechnol 25, 1015–1024.
Lahti, A. L., Kujala, V. J., Chapman, H., Koivisto, A. P., Pekkanen-Mattila, M., Kerkela, E., et al.(2012). Model for long QT syndrome type 2 using human iPS cells demonstrates ar-rhythmogenic characteristics in cell culture. Dis Model Mech 5, 220–230.
Lan, F., Lee, A. S., Liang, P., Sanchez-Freire, V., Nguyen, P. K., Wang, L., et al. (2013). Abnor-mal calcium handling properties underlie familial hypertrophic cardiomyopathy pa-thology in patient-specific induced pluripotent stem cells. Cell Stem Cell 12, 101–113.
Lee, M. P., Ravenel, J.D., Hu, R. J., Lustig, L. R., Tomaselli, G., Berger, R. D., et al. (2000).Targeted disruption of the Kvlqt1 gene causes deafness and gastric hyperplasia inmice. J Clin Invest 106, 1447–1455.
Lehnart, S. E., Ackerman, M. J., Benson, W., Brugada, R., Clancy, C. E., Donahue, J. K., et al.(2007). Inherited arrhythmias. A national heart, lung, and blood institute and officeof rare diseases workshop consensus report about the diagnosis, phenotyping,molecular mechanisms, and therapeutic approaches for primary cardiomyopathiesof gene mutations affecting ion channel function. Circulation 116, 2325–2345.
Liang, P., Lam, F., Lee, A. S., Gong, T., Sanchez-Freire, V., Wang, Y., et al. (2013). Drugscreening using a library of human induced pluripotent stem cell-derived
Please cite this article as: Sinnecker, D., et al., Induced pluripotent stem cePharmacology & Therapeutics (2014), http://dx.doi.org/10.1016/j.pharmth
cardiomyocytes reveals disease-specific patterns of cardiotoxicity. Circulation 127,1677–1691.
Lieu, D. K., Liu, J., Siu, C.W., McNerney, G. P., Tse, H. F., Abu-Khalil, A., et al. (2009). Absenceof transverse tubules contributes to non-uniform Ca(2+) wavefronts in mouse andhuman embryonic stem cell-derived cardiomyocytes. Stem Cells Dev 18, 1493–1500.
Ma, J., Guo, L., Fiene, S. J., Anson, B.D., Thomson, J. A., Kamp, T. J., et al. (2011). High purityhuman-induced pluripotent stem cell-derived cardiomyocytes: Electrophysiologicalproperties of action potentials and ionic currents. Am J Physiol Heart Circ Physiol301, H2006–H2017.
Ma, D., Wei, H., Lu, J., Ho, S., Zhang, G., Sun, X., et al. (2013). Generation of patient-specificinduced pluripotent stem cell-derived cardiomyocytes as a cellular model of arrhyth-mogenic right ventricular cardiomyopathy. Eur Heart J 34, 1122–1133.
Matsa, E., Dixon, J., Medway, C., Georgiou, O., Patel, M., Morgan, K., et al. (2013). Allele-specific RNA interference rescues the long-QT syndrome phenotype in human in-duced pluripotent stem cell cardiomyocytes. Eur Heart J. http://dx.doi.org/10.1093/eurheartj/eht067.
Matsa, E., Rajamohan, D., Dick, E., Young, L., Mellor, I., Staniforth, A., et al. (2011). Drugevaluation in cardiomyocytes derived from human induced pluripotent stem cellscarrying a long QT syndrome type 2 mutation. Eur Heart J 32, 952–962.
Mioulane, M., Foldes, G., Ali, N. N., Schneider, M.D., & Harding, S. E. (2012). Developmentof high content imaging methods for cell death detection in human pluripotent stemcell-derived cardiomyocytes. J Cardiovasc Transl Res 5, 593–604.
Moretti, A., Bellin, M.,Welling, A., Jung, C. B., Lam, J. T., Bott-Flugel, L., et al. (2010). Patient-specific induced pluripotent stem-cell models for long-QT syndrome. N Engl J Med363, 1397–1409.
Moretti, A., Laugwitz, K. L., Dorn, T., Sinnecker, D., & Mummery, C. (2013). Pluripotentstem cell models of human heart disease. Cold Spring Harb Perspect Med 3, a014027.
Mummery, C., Ward-van Oostwaard, D., Doevendans, P., Spijker, R., van den Brink, S.,Hassink, R., et al. (2003). Differentiation of human embryonic stem cells tocardiomyocytes: Role of coculture with visceral endodermlike cells. Circulation 107,2733–2740.
Nademanee, K., & Singh, B. N. (1988). Control of cardiac arrhythmias by calcium antago-nists. Ann N Y Acad Sci 522, 536–552.
Narsinh, K. H., Jia, F., Robbins, R. C., Kay, M.A., Longaker, M. T., & Wu, J. C. (2011). Gener-ation of adult human induced pluripotent stem cells using nonviral minicircle DNAvectors. Nat Protoc 6, 78–88.
Navarrete, E. G., Liang, P., Lan, F., Sanchez-Freire, V., Simmons, C., Gong, T., et al. (2013).Screening drug-induced arrhythmia events using human induced pluripotent stemcell-derived cardiomyocytes and low-impedance microelectrode arrays. Circulation128(Suppl. 1), S3–S13.
Nerbonne, J. M., Nichols, C. G., Schwarz, T. L., & Escande, D. (2001). Genetic manipulationof cardiac K+ channel function in mice. What we have learned, and where do we gofrom here? Circ Res 89, 944–956.
Novak, A., Barad, L., Zeevi-Levin, N., Shick, R., Shtrichman, R., Lorber, A., et al. (2012).Cardiomyocytes generated from CPVTD307H patients are arrhythmogenic in re-sponse to beta-adrenergic stimulation. J Cell Mol Med 16, 468–482.
Nunes, S. S., Miklas, J. W., Liu, J., Aschar-Sobbi, R., Xiao, Y., Zhang, B., et al. (2013). Biowire:A platform for maturation of human pluripotent stem cell-derived cardiomyocytes.Nat Methods 10, 781–787.
Redfern, W. S., Carlsson, L., Davis, A. S., Lynch, W. G., MacKenzie, I., Palethorpe, S., et al.(2003). Relationships between preclinical cardiac electrophysiology, clinical QT inter-val prolongation and torsade de pointes for a broad range of drugs: Evidence for aprovisional safety margin in drug development. Cardiovasc Res 58, 32–45.
Satin, J., Itzhaki, I., Rapoport, S., Schroder, E. A., Izu, L., Arbel, G., et al. (2008). Calcium han-dling in human embryonic stem cell-derived cardiomyocytes. Stem Cells 26, 1961–1972.
Seki, T., Yuasa, S., Oda, M., Egashira, T., Yae, K., Kusumoto, D., et al. (2010). Generation ofinduced pluripotent stem cells from human terminally differentiated circulating Tcells. Cell Stem Cell 7, 11–14.
Sinnecker, D., Moretti, A., & Laugwitz, K. L. (2013). Negating the dominant-negativeallele: A new treatment paradigm for arrhythmias explored in human inducedpluripotent stem cell-derived cardiomyocytes. Eur Heart J. http://dx.doi.org/10.1093/eurheartj/eht130.
Sirenko, O., Crittenden, C., Callamaras, N., Hesley, J., Chen, Y. -W., Funes, C., et al. (2013).Multiparameter in vitro assessment of compound effects on cardiomyocyte physiol-ogy using iPSC cells. J Biomol Screen 18, 39–53.
Sun, N., Yazawa, M., Liu, J., Han, L., Sanchez-Freire, V., Abilez, O. J., et al. (2012). Patient-specific induced pluripotent stem cells as a model for familial dilated cardiomyopa-thy. Sci Transl Med 4, 130ra147.
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., et al. (2007). In-duction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell131, 861–872.
Takahashi, K., & Yamanaka, S. (2006). Induction of pluripotent stem cells frommouse em-bryonic and adult fibroblast cultures by defined factors. Cell 126, 663–676.
Uosaki, H., Fukushima, H., Takeuchi, A., Matsuoka, S., Nakatsuji, N., Yamanaka, S., et al.(2011). Efficient and scalable purification of cardiomyocytes from human embryonicand induced pluripotent stem cells by VCAM1 surface expression. PLoS One 6, e23657.
Van Hoof, D., Dormeyer, W., Braam, S. R., Passier, R., Monshouwer-Kloots, J., Ward-vanOostwaard, D., et al. (2010). Identification of cell surface proteins for antibody-based selection of human embryonic stem cell-derived cardiomyocytes. J ProteomeRes 9, 1610–1618.
Yakubov, E., Rechavi, G., Rozenblatt, S., & Givol, D. (2010). Reprogramming of human fi-broblasts to pluripotent stem cells using mRNA of four transcription factors.Biochem Biophys Res Commun 394, 189–193.
Yang, L., Soonpaa, M. H., Adler, E. D., Roepke, T. K., Kattman, S. J., Kennedy, M., et al.(2008). Human cardiovascular progenitor cells develop from a KDR+ embryonic-stem-cell-derived population. Nature 453, 524–528.
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004
7D. Sinnecker et al. / Pharmacology & Therapeutics xxx (2014) xxx–xxx
Yazawa, M., Hsueh, B., Jia, X., Pasca, A.M., Bernstein, J. A., Hallmayer, J., et al. (2011). Usinginduced pluripotent stem cells to investigate cardiac phenotypes in Timothy syn-drome. Nature 471, 230–234.
Yu, J., Vodyanik, M.A., Smuga-Otto, K., Antosiewicz-Bourget, J., Frane, J. L., Tian, S., et al.(2007). Induced pluripotent stem cell lines derived from human somatic cells.Science 318, 1917–1920.
Please cite this article as: Sinnecker, D., et al., Induced pluripotent stem cePharmacology & Therapeutics (2014), http://dx.doi.org/10.1016/j.pharmth
Zhang, Q., Jiang, J., Han, P., Yuan, Q., Zhang, J., Zhang, X., et al. (2011). Direct differentiationof atrial and ventricular myocytes from human embryonic stem cells by alternatingretinoid signals. Cell Res 21, 579–587.
Zhu, W. Z., Xie, Y., Moyes, K. W., Gold, J.D., Askari, B., & Laflamme, M.A. (2010).Neuregulin/ErbB signaling regulates cardiac subtype specification in differentiatinghuman embryonic stem cells. Circ Res 107, 776–786.
ll-derived cardiomyocytes for drug development and toxicity testing,era.2014.03.004





























