Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Animal Behaviour 84 (2012) 361e368
Contents lists available
Animal Behaviour
journal homepage: www.elsevier .com/locate/anbehav
Individual and social learning in tandem-running recruitment by ants
Elizabeth L. Franklin*, Nigel R. Franks 1
School of Biological Sciences, University of Bristol, U.K.
a r t i c l e i n f o
Article history:Received 5 March 2012Initial acceptance 3 April 2012Final acceptance 25 April 2012Available online 7 June 2012MS. number: A12-00174R
Keywords:antlearningsocial learningtandem runningTemnothorax albipennis
* Correspondence: E. L. Franklin, School of BioloBristol, Woodland Road, Bristol, BS8 1UG, U.K.
E-mail address: [email protected] (E. L. Fra1 E-mail address: [email protected] (N. R. Fr
0003-3472/$38.00 � 2012 The Association for the Studoi:10.1016/j.anbehav.2012.05.002
Animals may acquire information socially, but do they also need to learn independently to make effectiveuse of such information? Tandem running is a form of recruitment in certain ants in which one individualleads another to an important resource such as a better nest site. Tandem runs speed information flowbecause tandem followers typically discover new nest sites much more rapidly than through indepen-dent searching. Moreover, tandem followers often go on to lead new tandem runs, thereby recruitingothers. However, former tandem followers do not slavishly follow the route they have been shown. Here,we show that individual ants can improve upon the directness and reduce the duration of tandem runsthey subsequently lead but do so only if, between being a follower and becoming a tandem leader, theyhave explored independently. Similarly, an ant that independently discovers a new nest site, andsubsequently leads tandem runs to such a discovery, improves upon the route taken only when she hasalso explored independently after finding the nest site. Thus, we show that learning through indepen-dent exploration is important both to individuals and to their society because it augments informationgained through tandem runs and is the basis of route improvement. This will further hasten the transferof information between members of the society, causing more effective information cascades. Tandemrunning allows one ant to teach another the location of a resource, but to improve their routes, taughtants also need to learn for themselves.� 2012 The Association for the Study of Animal Behaviour. Published by Elsevier Ltd. All rights reserved.
Animals may acquire information socially, but do they also needto learn independently to make effective use of such information?Examples from the literature on animal social learning suggest thatthis is the case: individuals often learn to refine information gainedsocially through personal experience or practice. For example, adultmeerkats provide their pups with the opportunity to interact withscorpions, but it is through independent practice that the pupslearn how to kill scorpions efficiently (Thornton & McAuliffe 2006).Moreover, when certain primates handle food, there is a combina-tion of social learning followed by individual trial-and-errorlearning (Tomasello 1996). Here we ask whether certain socialinsects combine social learning with individual learning.
Organisms frequently integrate personal information, acquiredindependently, and social information, acquired from others(Valone & Templeton 2002; Dall et al. 2005; Czaczkes et al. 2011).The behaviour of actively sharing personal information with othersis known as cooperative communication (Wilson 1975). Suchcommunication is not restricted to vertebrates; moreover,
gical Sciences, University of
nklin).anks).
dy of Animal Behaviour. Published
communication and social learning are of great importance in thesocial insects, allowing them to live in large cooperative societies(Leadbeater & Chittka 2007). Indeed, the cohesive actions thatresult from their communication systems are one of the reasonsthat certain social insect colonies are referred to as superorganisms(Franks 1989; Hölldobler & Wilson 2008). Additionally, socialinsects provide opportunities to elucidate, through experiments,the behavioural mechanisms that are used to facilitate the flow ofinformation among and between individuals.
Information flow among and between individuals within socialinsect colonies allows them to achieve impressive feats of coordi-nation, for example, during foraging or house hunting (Seeley 1989;Franks et al. 2002). Social insects often use a combination of indi-vidual and cooperative social learning to acquire information(Leadbeater & Chittka 2007). One of the most famous examples ofthis is the foraging of the honeybee Apis mellifera. Honeybeeforagers find resources independently, then return to the hive toshare foraging informationwith other workers, through the code ofthe waggle dance (von Frisch 1956). The waggle dance allowsobserving bees to learn the location of the forage from the dancer’sindependent explorations. Tandem running is a behaviour thatfulfils a similar role in certain ant species. In tandem running, an antshares information about the location of a site that she foundindependently by leading a single follower to it, pausing if the
by Elsevier Ltd. All rights reserved.

E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368362
follower loses contact (Hölldobler & Wilson 1990). Temnothoraxalbipennis ants use tandem-running recruitment both for foragingand in house hunting. They nest within fragile rock crevice nests,which frequently become uninhabitable, forcing colonies to findand emigrate to new nests (Partridge et al. 1997). Thus, they are alsocontinually on the ‘look out’ for a better nest and will move intoa different nest if it is a substantial improvement over their existingnest (‘move to improve’; Dornhaus et al. 2004).
Tandem running is important in nest choice because it buildsthe ‘quorum’ (i.e. the threshold number of ants needed in a newpotential nest site to trigger recruitment by social carrying andhence acceptance of that nest site). Moreover, the quorum isassociated with the small but highly influential group of knowl-edgeable ants primarily responsible for relocating the remainingcolony members by social carrying (Pratt et al. 2002).
In an emergency emigration, moving the colony to the safety ofa new location is the priority, and even a small number of tandemruns should speed up nest discovery and any subsequent emigra-tion, thus reducing the time the colony is exposed (Pratt 2008;Franks et al. 2009). In a ‘move to improve’ situation, a particularlyhigh quorum threshold is used to help ensure that a correct deci-sion is made about a new nest site; thus, a colony only puts itself atrisk during an emigration if the new nest is much better than theold one. Here, large numbers of tandem runs are used to exceeda very high quorum threshold and to obtain a clear consensusdecision for the colony, ensuring that colonies only ‘move toimprove’ (Dornhaus et al. 2004).
In T. albipennis, a few solitary scouts from each colony find foodand new nest sites (Franks et al. 2002) mainly using visual navi-gation (Pratt et al. 2001; McLeman et al. 2002). Temnothorax albi-pennis ants probably do not use pheromone trail recruitmentbecause there are too few scouting ants at any one time to initiateand maintain pheromone trails before they degrade (Beckers et al.1989; Franks & Richardson 2006; Planqué et al. 2010). It is likelythat followers also learn navigational cues, such as landmarks,when following a tandem run or exploring independently. It alsoseems likely that these ants may use path integration (Müller &Wehner 1988; McLeman et al. 2002).
There is much more to tandem running than a leader simplygetting another ant from one location to another. IndeedT. albipennis can do this three times more quickly by social carrying.However, unlike tandem followers, carried ants seem not to be ablelearn landmarks, because they are held with their heads invertedand facing backwards, nor would they be able to use path inte-gration since they are denied an opportunity to pace out the pathfor themselves. Accordingly, carried ants typically stay where theyhave been deposited (probably because they do not know wherethe new nest is with respect to the old one) whereas tandemfollowers typically go on to explore the environment between thenewnest and the old nest and frequently subsequently lead tandemruns of their own.
Tandem running has been identified as one of the rare cases ofteaching in nonhuman animals (Franks & Richardson 2006). Theleading ant in a tandem run has also been shown to evaluate thetandem’s progress and the follower’s quality (Richardson et al.2007). Followers of tandem runs can extrapolate successfullyfrom the direction of the tandem run if they lose their leader enroute (Franks et al. 2010). Nevertheless, we do not know whatinformation is transferred to, or gained by, a follower duringa tandem run, only that, after following a tandem run, the followercan go on to lead a tandem run to the site herself (Franks &Richardson 2006). It has been hypothesized that the slowponderous nature of tandem running provides the follower withthe opportunity to learn landmarks en route to a new site (Franks &Richardson 2006). It has also been suggested that, if the route of
a tandem could be learnt by the follower, the route could beimproved on in a cumulative manner over subsequent tandem runs(Franks & Richardson 2006). However, we do not know whether itis the location of the new site or the route to it that the followingants are learning. We define route learning as having informationabout a series of cues or landmarks that can be followed to reachthe target. By contrast, by learning the location of the new nest, wemean that the ant knows the home vector (i.e. the distance anddirection of the old nest from the new one and vice versa). Suchinformation could be obtained, for example, through path inte-gration (Müller & Wehner 1988).
By examining how information is gained and exchanged, wehope to shed light on theway that information flowwithin coloniesleads to coordinated emigrations. To do this, we investigated thecontributions of social and individual learning to tandem-runningefficiency in the ant T. albipennis. We compared the durations anddirectness of tandem runs led by ants that had either been recruitedby tandem running, or found the nest independently, taking intoaccount any independent explorations they performed. We alsocompared sequential tandems where the follower of a tandemwent on to be the leader of a subsequent run. In this way, we testedwhether efficiency improves progressively over successive tandemruns, or whether independent exploration is required.
We then address the issue of what information is gained fromtandem runs. Are ants learning the route to a nest or the location ofthe target? We did this by comparing the consistency of the pathsof pairs of sequential tandem runs (i.e. where the follower ofa tandem became the leader of a subsequent one) and independenttandem runs (that had no participants in common) as well as anindividual’s last exploration and its subsequent leading of a tandemrun. We expected to see similarities in the paths of pairs ofsequential tandem runs if followers learn specific routes whenfollowing a tandem run. We also expected the paths of pairs ofsequential tandems to be more similar to one another than to pairsof independent tandem runs.
METHODS
We collected 15 T. albipennis colonies from the coast of Dorsetin October 2010. Colonies were then housed in ‘poor’ nests con-sisting of a thin cardboard perimeter (providing 0.8 mm of headroom) within a 31 � 25 mm cavity between 75 � 50 mm micro-scope slides, with a 4 mm diameter entrance hole in the centre ofthe roof. A ‘poor’-quality nest is one that is less attractive to theants, in this case because it was well lit and cramped (Franks et al.2003). The ants were fed once a week with dilute honey solutionand dead Drosophila and were provided water ad libitum.
The experiment used the ‘move to improve’ paradigm(Dornhaus et al. 2004), where ants may search for and emigrate toa nest site that is better than the nest they currently occupy. Theexperimental set-up consisted of a small arena (100 � 100 mm)containing a poor-quality nest. This small arena was connected bytwo tunnels, side by side (45 � 10 � 10 mm), to a larger arena(230 � 230 mm) containing a high-quality, more attractive nest(dark, 62 � 40 mm cavity and high roof 1.8 mm; Franks et al. 2003;Fig. 1). The arena walls were coated with Fluon � to prevent theescape of ants, and were cleaned with ethanol between experi-ments to ensure the removal of any pheromonemarkings. The new,high-quality nest’s entrance was 220 mm from the old nest and150 mm from the exit of the tunnels. The tunnels between thearenas could be blocked to control the flow of ants between arenas.The larger arena containing the new nest was back-lit to silhouettethe ants, making them easier to film and track (Fig. 1).
At the beginning of each experimental trial, a colony in its poor-quality nest was placed in the small arena. Each colony was only

Sector 1250
200
150
50
0
Sector 2 Sector 3
100
0 100 150 200 250 30050
Tooldnest
Tandem 1Tandem 2
Newnest
Figure 1. The large arena within which ants’ paths were filmed. An example of a pairof sequential paths is shown. Scales on the X and Y axes are in mm. The three sectorsare marked: sector 1 (0e50 mm), sector 2 (50e100 mm) and sector 3 (100e150 mm,new nest edge).
E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368 363
used once. The temperature of the experimental room was main-tained between 20 and 22 �C, and no objects in the room weremoved during the course of an experimental trial. A new, high-quality nest was placed in the large arena 150 mm away from theconnecting tunnel entrances. The ants were allowed to exploreuntil one ant found the new nest. At this point, we removed allother ants and placed them back in the small arena, and placeda door in the connecting tunnels to prevent other ants fromexploring. The large arena was filmed from this point on usinga digital video camera (Panasonic NV-DS55V). When the successfulant emerged from the new nest, we closely monitored her trips toand from the old nest. The ant’s movements to and from the newand old nest were tracked and recorded until she had completeda tandem run to the new nest. When the tandem reached the newnest, we removed the leader from the experiment and allowed thefollower access to the nest. When the follower emerged fromthe nest, we tracked her until she too had led a tandem run. Whenthe tandem run was complete, we removed both participantsfrom the experiment. We then opened the doorway to the tunnels,allowing a new ant to discover the nest, and repeated the process.By only allowing one ant to explore or to lead a tandem run at anyone time, we could follow the flow of the information through thecolony as it was gained individually and transferred socially.
We were particularly interested in sequential tandem runs(Fig. 2b, c), where the follower of a tandem run became the leaderof a subsequent run. Such events provide an ideal opportunity toexamine the information that the follower acquired whenfollowing a tandem run as manifest in her ability to lead a subse-quent tandem run. We were also interested in the individualexplorations of the ants. In this study, we defined an individualexploration as an extra trip from the old nest to the new nest thatwas not the trip on which the ant discovered the nest or a tandemrun. These outward journeys could then be compared directly tooutbound tandem runs.
Analysis
Complete individual and tandem paths (from the exit of thetunnels to the new nest) were recorded on a digital video camera.They were then digitized using Adobe Premier Pro 2.0 and runthrough a custom tracking program called CTRAX (version 0.1.5.6,http://dickinson.caltech.edu/ctrax) From CTRAX, we acquired the
frame-by-frame location of ants throughout their journeys. Wethen calculated the comparative efficiencies of the ants’ trajectoriesin terms of journey durations (time taken to complete a tandemrun) and directness, by measuring path tortuosity (1� (straight linedistance/actual path taken); Benhamou 2004) using MatLab(Mathworks, Natick, MA, U.S.A.). For all ants, we took the locationdata from the centroid of the ant, and in tandems, we used theleader’s centroid as the location of the tandem, as the leader andthe follower usually remain in relatively close contact throughouta tandem run (Franks & Richardson 2006).
To explore the effects of individual and social learning ontandem run efficiency and information transfer, we first comparedall tandems (Fig. 2bee) and then examined pairs of sequentialtandems (Fig. 2b, c). We compared all tandem runs (N ¼ 81) withregard to duration and tortuosity using a general linear model(GLM). We used ln transformation to normalize the residuals of theduration and tortuosity data before performing GLMs. The full GLMmodel for the analysis of duration and tortuosity included nestdiscovery (independent or recruited), independent explorations (0,1 or 2þ) and their interactions as predicting variables. The finalGLM model for duration and tortuosity only had independentexplorations as a predicting variable. The final model was obtainedby removing nonsignificant interactions and predictors. We thencompared sequential tandems (where the follower of a tandembecame the leader of a subsequent run; Fig. 2b, c) using pairedstatistical tests.
To investigatewhether target or route information is transferredduring tandem running, we compared the similarity of fourdifferent pairs of paths: independent tandems (Fig. 2a); sequentialtandems with individual explorations (Fig. 2b); sequential tandemswithout individual explorations (Fig. 2c); and an ant’s last explo-ration and its subsequent tandem run (Fig. 2d). We did this bymeasuring the maximum distance between paths at intervals of10 mmon the X axis (the straight line from the exit of the tunnels tothe new nest entrance). These measurements were grouped intothree sectors: sector 1 (0e50 mm); sector 2 (50e100 mm); sector 3(100e150 mm; new nest edge). This was done to isolate the effectsof clustering caused by the tunnels (Fig. 1, Fig. 6 inset). The tunnels’combined entrance width was only 20 mm and therefore con-stricted the separation of paths. The maximum difference betweenpaths was chosen for analysis, over the average or the minimum, astaking the average or minimum gives the appearance that convo-luted paths that double back on themselves are more similar toother paths than they actually are. All GLM fitting and otherstatistical analyses were carried out in Minitab (Minitab, Inc., StateCollege, PA, U.S.A.).
RESULTS
Individual Learning
There was significant improvement between successive tandemruns only if the former followers engaged in a bout of individualexploration before they went onto lead a tandem run. Suchimprovement was manifest in terms of both shorter durations andreduced tortuosities (univariate ANOVA: ln duration: F2,78 ¼ 6.367,P ¼ 0.003; Fig. 3a; ln tortuosity: F2,78 ¼ 6.4, P ¼ 0.003; Fig. 3b).Moreover, when we compared sequential tandem runs (where thefollower of a tandem run became the leader of a subsequenttandem run; Fig. 2b, c), only ants that followed a tandem run andthen performed individual explorations before leading a tandemrun showed significant improvement in duration and tortuosity ofthe tandems that they had initially followed (sequential tandemswith individual exploration: Wilcoxon signed-ranks test: duration:

Independent tandems
Black ant leads
grey ant in a
tandem run
Black-hashed ant
leads grey-
hashed ant in a
tandem run
Sequential tandems with independent exploration
Black ant leads
grey ant in a
tandem run
Grey ant
explores
independently
Grey ant leads
black-hashed ant
in a tandem run
Sequential tandems without independent exploration
Black ant leads
grey ant in a
tandem run
Grey ant does not explore
independently
Grey ant
immediately
leads black-
hashed ant in a
tandem run
An ant’s last exploration and its subsequent tandem run
Black ant explores
independently
Black ant does not explore
subsequently
Black ant leads
grey ant in a
tandem run
Ant performs tandem immediately after independently discovering the new nest
Black ant
discovers nest
independently
Black ant does not explore
independently
Black ant leads
grey ant in a
tandem run
(a)
(b)
(c)
(d)
(e)
Figure 2. Categories of tandem runs used by Temnothorax albipennis ants and how information may be gained in each case.
E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368364
Z ¼ �2.675, N ¼ 17, P ¼ 0.007; Fig. 4a; paired t test: tortuosity:t16 ¼ 2.55, P ¼ 0.021; Fig. 4b).
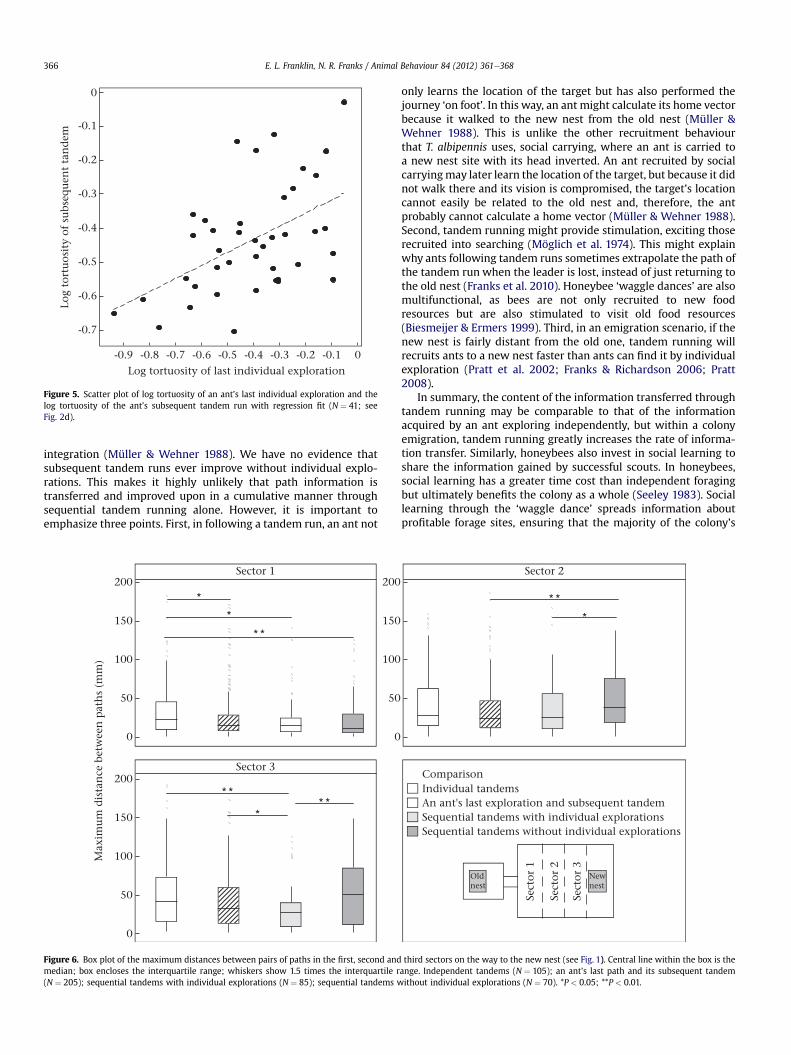
There was a positive correlation between the log tortuosity of anant’s last individual exploration and the log tortuosity of itssubsequent tandem (regression analysis: t40 ¼ 3.63, R2 ¼ 23.3%,P ¼ 0.001; Fig. 2d, Fig. 5). This correlation was not present in any ofthe other groups (i.e. sequential tandem runs with or withoutindividual explorations or independent tandem runs).
Social Learning
Most intriguingly, the duration and directness of tandem runsdid not differ significantly between individuals that had been
recruited to the nest via tandem running or that had found the nestindependently (Fig. 3). We then compared sequential tandem runs(where the follower of a tandem run became the leader ofa subsequent tandem run; Fig. 2b, c). Ants that led subsequenttandem runs showed significant improvement in duration andtortuosity of the tandem runs that they had previously followedonly when they performed individual explorations before leadingtheir own tandem run (Fig. 4).
Route or Target Learning
We compared the route consistencies of pairs of paths: (1)sequential tandems with individual explorations; (2) sequential

RecruitedIndependent nestdiscovery
RecruitedIndependent nestdiscovery
2+102+0
400
300
200
100
0Individualexplorations
Individualexplorations
2+102+1
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
(a)
(b)
500
Tim
e (s
)
1
1
Tot
uos
ity
(1−s
trai
ghtl
ine/
actu
al)
0
Figure 3. Box plots of (a) duration and (b) tortuosity of complete tandems of ants thatwere either recruited or discovered the nest independently, and the number of indi-vidual explorations that an ant performed before performing a tandem run. Horizontalline within the box is the median; box encloses the interquartile range; whiskers show1.5 times the interquartile range; asterisks indicate outliers. Independent discoveries:0 explorations, N ¼ 19 (Fig. 2e); 1 exploration, N ¼ 19 (Fig. 2d); 2 or more explorations,N ¼ 12, (Fig. 2b). Recruited: 0 explorations, N ¼ 15 (Fig. 2c); 1 exploration, N ¼ 10(Fig. 2b); 2 or more explorations, N ¼ 7 (Fig. 2b).
Subsequenttandem led
Tandemfollowed
Subsequenttandem led
400
300
200
100
0
Tim
e (s
)
1
0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
Without individualexplorations
With individualexplorations
Sequential tandems
Tandemfollowed
Subsequenttandem led
Tandemfollowed
Subsequenttandem led
Tandemfollowed
(a)* *
*(b)
500
Tor
tuos
ity
(1−s
trai
ght
lin
e/ac
tual
)
Figure 4. Box plots of the (a) duration (b) tortuosity of pairs of sequential tandems(the tandem run followed by an ant and the subsequent tandem run she led), with(Fig. 2b) and without (Fig. 2c) at least one individual exploration. Horizontal linewithin the box is the median; box encloses the interquartile range; whiskers show 1.5times the interquartile range; asterisks denote outliers. Without individual explora-tions, N ¼ 14; with individual explorations, N ¼ 17. Group differences calculated witha paired t test: *P < 0.05; **P < 0.01.
E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368 365
tandems without individual explorations; (3) pairs of independenttandems from within an emigration; and (4) an ant’s last inde-pendent exploration and its subsequent tandem (Fig. 2aed).We didthis by measuring the maximum distance between paths. In thefirst sector, independent tandems (Fig. 2a) were significantly lesssimilar than the other groups (KruskaleWallis test: H3 ¼ 10.30,P ¼ 0.016; Fig. 6). In the second sector, an ant’s last individualexploration and its subsequent tandem (Fig. 2d) were more similarthan either sequential tandem pairs with or without individualexplorations (H3 ¼ 9.15, P ¼ 0.027; Fig. 6). In the third sector,sequential tandem pairs with individual exploration (Fig. 2b) weresignificantly more similar than other pairs (H3 ¼ 12.72, P ¼ 0.005;Fig. 6).
DISCUSSION
Our results show that the duration and directness of tandemruns improved only when ants performed at least one bout ofexploration before leading a new tandem run, irrespective ofwhether the ants had been recruited by a tandem run or haddiscovered the new nest independently (Figs 3, 4). This implies thattandem-running recruitment provides the following ant with thesame information it would acquire if it had found the new nestindependently. Ants following a tandem run do not know theultimate target of the tandem they are following until they reach it,just as in individual exploration. Our results suggest that theinformation transferred through a tandem run is comparable to anant finding the nest herself and learning the return route bylandmarks (Pratt et al. 2001; McLeman et al. 2002) or path
0-0.1-0.2-0.3-0.4-0.5-0.6-0.7-0.8-0.9
0
-0.1
-0.2
-0.3
-0.4
-0.5
-0.6
-0.7
Log tortuosity of last individual exploration
Log
tort
uos
ity
of s
ubs
equ
ent
tan
dem
Figure 5. Scatter plot of log tortuosity of an ant’s last individual exploration and thelog tortuosity of the ant’s subsequent tandem run with regression fit (N ¼ 41; seeFig. 2d).
E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368366
integration (Müller & Wehner 1988). We have no evidence thatsubsequent tandem runs ever improve without individual explo-rations. This makes it highly unlikely that path information istransferred and improved upon in a cumulative manner throughsequential tandem running alone. However, it is important toemphasize three points. First, in following a tandem run, an ant not
150
100
50
0
200
150
100
50
0
Max
imu
m d
ista
nce
bet
wee
n p
ath
s (m
m)
150
100
50
0
Sector 1
Sector 3
*
*
* *
* ** *
*
200200
Figure 6. Box plot of the maximum distances between pairs of paths in the first, second andmedian; box encloses the interquartile range; whiskers show 1.5 times the interquartile r(N ¼ 205); sequential tandems with individual explorations (N ¼ 85); sequential tandems w
only learns the location of the target but has also performed thejourney ‘on foot’. In this way, an ant might calculate its home vectorbecause it walked to the new nest from the old nest (Müller &Wehner 1988). This is unlike the other recruitment behaviourthat T. albipennis uses, social carrying, where an ant is carried toa new nest site with its head inverted. An ant recruited by socialcarryingmay later learn the location of the target, but because it didnot walk there and its vision is compromised, the target’s locationcannot easily be related to the old nest and, therefore, the antprobably cannot calculate a home vector (Müller & Wehner 1988).Second, tandem running might provide stimulation, exciting thoserecruited into searching (Möglich et al. 1974). This might explainwhy ants following tandem runs sometimes extrapolate the path ofthe tandem run when the leader is lost, instead of just returning tothe old nest (Franks et al. 2010). Honeybee ‘waggle dances’ are alsomultifunctional, as bees are not only recruited to new foodresources but are also stimulated to visit old food resources(Biesmeijer & Ermers 1999). Third, in an emigration scenario, if thenew nest is fairly distant from the old one, tandem running willrecruits ants to a new nest faster than ants can find it by individualexploration (Pratt et al. 2002; Franks & Richardson 2006; Pratt2008).
In summary, the content of the information transferred throughtandem running may be comparable to that of the informationacquired by an ant exploring independently, but within a colonyemigration, tandem running greatly increases the rate of informa-tion transfer. Similarly, honeybees also invest in social learning toshare the information gained by successful scouts. In honeybees,social learning has a greater time cost than independent foragingbut ultimately benefits the colony as a whole (Seeley 1983). Sociallearning through the ‘waggle dance’ spreads information aboutprofitable forage sites, ensuring that the majority of the colony’s
Individual tandemsAn ant's last exploration and subsequent tandemSequential tandems with individual explorationsSequential tandems without individual explorations
Comparison
Sect
or 1
Sect
or 2
Sect
or 3
Sector 2
Oldnest
Newnest
* *
*
third sectors on the way to the new nest (see Fig. 1). Central line within the box is theange. Independent tandems (N ¼ 105); an ant’s last path and its subsequent tandemithout individual explorations (N ¼ 70). *P < 0.05; **P < 0.01.

E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368 367
foragers are visiting them (Seeley 1983). The major contrastbetween T. albipennis and honeybee recruitment strategies is mostlikely due to colony size. Temnothorax albipennis ants live in smallcolonies where they can only spare a few scouts for exploration.Consequently, they need to ensure that information is not lost intransmission. Hence, T. albipennis invest in slow, one-on-one,tandem running rather than the less directed broadcasting typi-cally used by larger social insect colonies (Beckers et al. 1989;Franks & Richardson 2006).
Tandem running is a form of social learning where the leaderactively facilitates the learning of the follower. It is a robust(Franklin et al. 2011), sophisticated communication system (Franks& Richardson 2006) with evaluation (Richardson et al. 2007) andfail-safes (Franks et al. 2010) that recruits the ants needed to reacha consensus and to emigrate the colony (Franks et al. 2009).Tandem running can be performed by young and inexperiencedants in T. albipennis, but it can also be refined through experience(Franklin et al. 2012). The role of tandem running in an emigrationis to increase the information flow within colony and ensure thatinformation is transferred securely.
Our results also show that only one bout of individual explora-tion is needed to improve a subsequent tandem’s duration andstraightness significantly and that performing extra explorationsbeyond this does not improve efficiency further (Figs 3e6). Inaddition, successful, efficient and independent exploration waspositively correlated with subsequent accurate leadership oftandem runs (Fig. 5). These results may explain why tandemleaders, if they lose their follower, typically continue to the newnest rather than returning to the old one to find another follower(Franks et al. 2010). The leader, by continuing to the new nest, maybe able to improve her route efficiency before beginning to recruitagain, which may help her perform a more direct tandem runsubsequently. She may also be checking to see whether the quorumhas been met in the nest so that she can switch her recruitmenteffort to social carrying, which is much quicker (Pratt et al. 2002;Pratt 2008).
When we compared the consistency of pairs of paths, we foundthat no pairs were consistently more similar than others. Pairs ofindependent tandem runsmight be expected to be less similar thanpairs that share information (either independently or socially), butthis was apparent only in the first sector. This finding combinedwith the evidence that the ants did not use similar routes duringtheir own explorations or during the tandems they followed, leadsus to suggest that the information transferred in tandem running istarget oriented rather than route based (Fig. 6). The small yetsignificant differences that we detected in the second and thirdsector may be associated with path improvement. If paths areimproved towards the shortest route down the centre of the arena,then more accurate paths will ultimately be more similar.
In summary, both social and individual learning are used byT. albipennis in the discovery and recruitment of others to a newnest site. Tandem runs speed up the rate of information transferwithin the colony during emigration by directing scouts so thatthey find the target more rapidly than through individual explo-ration (Pratt et al. 2002; Pratt 2008). This is beneficial both inhelping to expedite the building of a consensus in favour of a betternest site in a ‘move to improve’ situation and in ensuring thatenough ants know the location of suitable new nest sites so thatthey can take an active role in a rapid emigration (Franks et al.2009). Intriguingly, it is individual exploration that improves theefficiency of routes and which in turn may lead to more efficientrecruitment. As the ants learn progressively shorter routes throughindividual exploration, they will be able to produce shorter tandemruns. These shorter tandem runs should be more reliable becausethere will be less opportunity for tandem leaders and followers to
lose one another. By maximizing information flow within thecolony, through more efficient recruitment, T. albipennis is able toachieve its impressive feats of coordination, facilitating well-informed decisions about new nests and coordinating theemigration of the whole colony.
Our study suggests that in other fields of animal behaviour thatinvolve social learning (Tomasello 1996) and especially those withteaching (Thornton & McAuliffe 2006; Thornton & Raihani 2008),attention should be paid to the possibility that information gainedsocially is built upon by independent learning by individuals.Indeed, this principle is so familiar in human teaching that it isrecognized by the cliché ‘practice makes perfect’.
Acknowledgments
We thank Ana Sendova-Franks for her advice on statisticalanalyses, and Elizabeth Langridge, Carolina Doran and DaphnaGottlieb for their helpful comments on the manuscript. We alsothank Andy and Lynn Franklin and Duncan Bates for all theirsupport and comments.
References
Beckers, R., Goss, S., Deneubourg, J. L. & Pasteels, J. M. 1989. Colony size,communication and ant foraging strategy. Psyche, 96, 239e256.
Benhamou, S. 2004. How to reliably estimate the tortuosity of an animal’s path:straightness, sinuosity, or fractal dimension? Journal of Theoretical Biology, 229,209e220.
Biesmeijer, J. C. & Ermers, M. C. W. 1999. Social foraging in stingless bees: howcolonies of Melipona fasciata choose among nectar sources. Behavioral Ecologyand Sociobiology, 46, 129e140.
Czaczkes, T. J., Grüter, C., Jones, S. M. & Ratnieks, F. L. W. 2011. Synergy betweensocial and private information increases foraging efficiency in ants. BiologyLetters, 7, 521e524.
Dall, S. R. X., Giraldeau, L.-A., Olsson, O., McNamara, J. M. & Stephens, D. W. 2005.Information and its use by animals in evolutionary ecology. Trends in Ecology &Evolution, 20, 187e193.
Dornhaus, A., Franks, N. R., Hawkins, R. M. & Shere, H. N. S. 2004. Ants move toimprove: colonies of Leptothorax albipennis emigrate whenever they finda superior nest site. Animal Behaviour, 67, 959e963.
Franklin, E. L., Richardson, T. O., Sendova-Franks, A. B., Robinson, E. J. H. &Franks, N. R. 2011. Blinkered teaching: tandem running by visually impairedants. Behavioral Ecology and Sociobiology, 65, 569e579.
Franklin, E. L., Robinson, E. J. H., Marshall, J. A. R., Sendova-Franks, A. B. &Franks, N. R. 2012. Do ants need to be old and experienced to teach? Journal ofExperimental Biology, 215, 1287e1292.
Franks, N. R. 1989. Army ants: a collective intelligence. American Scientist, 77,138e145.
Franks, N. R. & Richardson, T. 2006. Teaching in tandem running ants. Nature,439, 153.
Franks, N. R., Pratt, S. C., Mallon, E. B., Britton, N. F. & Sumpter, D. J. T. 2002.Information flow, opinion polling and collective intelligence in house-huntingsocial insects. Philosophical Transactions of the Royal Society of London, SeriesB, 357, 1567e1583.
Franks, N. R., Mallon, E. B., Bray, H. E., Hamilton, M. J. & Mischler, T. C. 2003.Strategies for choosing between alternatives with different attributes: exem-plified by house-hunting ants. Animal Behaviour, 65, 215e223.
Franks, N. R., Dechaume-Moncharmont, F.-X., Hanmore, E. & Reynolds, J. K.2009. Speed versus accuracy in decision-making ants: expediting politics andpolicy implementation. Philosophical Transactions of the Royal Society of London,Series B, 364, 845e852.
Franks, N. R., Richardson, T. O., Keir, S., Inge, S. J., Bartumeus, F. & Sendova-Franks, A. B. 2010. Ant search strategies after interrupted tandem runs. Journalof Experimental Biology, 213, 1697e1708.
von Frisch, K. 1956. Bees: Their Vision, Chemical Senses, and Language. Ithaca, NewYork: Cornell University Press.
Hölldobler, B. & Wilson, E. O. 1990. The Ants. Cambridge, Massachusetts: HarvardUniversity Press.
Hölldobler, B. & Wilson, E. O. 2008. The Superorganism: the Beauty, Elegance, andStrangeness of Insect Societies. London: W.W. Norton.
Leadbeater, E. & Chittka, L. 2007. Social learning in insects: from miniature brainsto consensus building. Current Biology, 17, 703e713.
McLeman, M. A., Pratt, S. C. & Franks, N. R. 2002. Navigation using visual land-marks by the ant Leptothorax albipennis. Insectes Sociaux, 49, 203e208.
Möglich, M., Maschwit, U. & Hölldobler, B. 1974. Tandem calling: new kind ofsignal in ant communication. Science, 186, 1046e1047.
Müller, M. & Wehner, R. 1988. Path integration in desert ants, Cataglyphis fortis.Proceedings of the National Academy of Sciences, U.S.A., 85, 5287e5290.

E. L. Franklin, N. R. Franks / Animal Behaviour 84 (2012) 361e368368
Partridge, L. W., Partridge, K. A. & Franks, N. R. 1997. Field survey of a monogy-nous leptothoracine ant (Hymenoptera, Formicidae) evidence of seasonalpolydomy? Insectes Sociaux, 44, 75e83.
Planqué, R., van den Berg, J. B. & Franks, N. R. 2010. Recruitment strategies andcolony size in ants. PLoS One, 5, e11664.
Pratt, S. C. 2008. Efficiency and regulation of recruitment during colony emigrationby the ant Temnothorax curvispinosus. Behavioral Ecology and Sociobiology, 62,1369e1376.
Pratt, S. C., Brooks, S. E. & Franks, N. R. 2001. The use of edges in visual navigationby the ant Leptothorax albipennis. Ethology, 107, 1125e1136.
Pratt, S. C., Mallon, E. B., Sumpter, D. J. T. & Franks, N. R. 2002. Quorum sensing,recruitment, and collective decision-making during colony emigration by theant Leptothorax albipennis. Behavioral Ecology and Sociobiology, 52, 117e127.
Richardson, T. O., Sleeman, P. A., McNamara, J. M., Houston, A. I. & Franks, N. R.2007. Teaching with evaluation in ants. Current Biology, 17, 1520e1526.
Seeley, T. D. 1983. Division of labor between scouts and recruits in honeybeeforaging. Behavioral Ecology and Sociobiology, 12, 253e259.
Seeley, T. D. 1989. The honey bee as a superorganism. American Scientist, 77,546e553.
Thornton, A. & McAuliffe, K. 2006. Teaching in wild meerkats. Science, 313,227e229.
Thornton, A. & Raihani, N. J. 2008. The evolution of teaching. Animal Behaviour, 75,1823e1836.
Tomasello, M. 1996. Do apes ape? In: Social Learning in Animals: the Roots of Culture(Ed. by C. M. Heyes & B.G. Galef Jr), pp. 319e347. London: Academic Press.
Valone, T. J. & Templeton, J. J. 2002. Public information for the assessment ofquality: a widespread social phenomenon. Philosophical Transactions of theRoyal Society of London, Series B, 357, 1549e1557.
Wilson, E. O. 1975. Sociobiology: the New Synthesis. Cambridge, Massachusetts:Belknap Press.





























