Embed Size (px)
Citation preview

J. Comp. Path. 1999 Vol. 121, 265–270
SHORT PAPER
In-situ Hybridization for the Detection of PorcineCircovirus in Pigs with Postweaning Multisystemic
Wasting Syndrome
C. Choi and C. ChaeDepartment of Veterinary Pathology, College of Veterinary Medicine, Seoul National University,
Suwon 441–744, Kyounggi-Do, Republic of Korea
Summary
In-situ hybridization with a non-radioactive digoxigenin-labelled probe wasused to detect porcine circovirus (PCV) in formalin-fixed paraffin wax-embedded tissues from 10 weaned pigs with naturally occurring postweaningmultisystemic wasting syndrome (PMWS). A 530 base pair DNA fragmentfrom an open reading frame 1 was generated by the polymerase chainreaction. By in-situ hybridization, a distinct positive signal was detected inall 10 pigs. The morphology of host cells was preserved despite the relativelyhigh temperature required in parts of the incubation procedure. Positive cellstypically exhibited a dark brown to black reaction product, mainly in thecytoplasm but occasionally in the nucleus, without background staining.Particularly consistent and intense labelling for PCV occurred in the lymphnodes. Virus was also detected in macrophages of the liver, spleen, Peyer’spatches, lung and tonsil, in hepatocytes and in renal tubular epithelial cells.The abundant presence of PCV strongly suggested that PCV plays animportant role in PMWS.
1999 Harcourt Publishers Ltd
Introduction
Porcine circovirus (PCV) was first isolated in 1974 as a persistent contaminantof the continuous porcine kidney cell line, PK-15 (ATCC CCL31) (Tischer etal., 1974). This cell culture isolate was shown to possess a circular single-stranded DNA genome of 1·76 kb (Meehan et al., 1997). On the basis of itsmorphology and circular single-stranded DNA genome, PCV was classifiedin a newly recognized animal virus family, the Circoviridae (Lukert et al., 1995).Types 1 and 2 of PCV were identified by Meehan et al., (1998). Type 1 is thePK-15 contaminant referred to above. Type 2, which differs markedly, is beingfound repeatedly in pigs with postweaning multisystemic wasting syndrome(PMWS) (Meehan et al., 1998; Morozov et al., 1998).
PMWS in pigs, which is characterized clinically by poor body conditionand moderate pallor of the skin (Clark, 1997), was first identified in WesternCanada in 1991. Since then, it has been reported in the USA (Clark, 1997),France (LeCann et al., 1997) and Spain (Segales et al., 1997). The demonstration
Correspondence to: C. Chae.
0021–9975/99/070265+06 $12.00 1999 Harcourt Publishers Ltd

266 C. Choi and C. Chae
Table 1In-situ hybridization in 10 weaned pigs naturally infected with porcine circovirus
Pig no. Age In-situ hybridization results∗ in the stated location(days)
Lymph node Liver Spleen Peyer’s patches Lung Kidney Tonsil
1 60 +++ ++ ++ + − ++ +2 35 +++ ++ + + − +++ +3 50 +++ ++ + − − − +4 49 +++ ++ + + +++ − −5 37 +++ ++ ++ + ++ +++ −6 120 +++ +++ ++ ++ +++ +++ +++7 52 ++ + ++ + + ++ ++8 57 +++ ++ ++ +++ ++ ++ +++9 62 +++ ++ ++ + ++ + +10 58 ++ ++ ++ − ++ +++ ++
∗−, None;+, slight (<1 cell/high power field [×400]); ++, moderate (approximately 1 cell/high powerfield); +++, extensive (>1 cell/high power field).
of PCV antigen, closely associated with lesions, in a range of tissues fromdiseased pigs (LeCann et al., 1997; Allan et al., 1998; Ellis et al., 1998), has ledto speculation that PCV plays an aetiological role in PMWS.
In-situ hybridization is a valuable adjunct to standard DNA extraction techniquesfor evaluating gene expression in tissues and cells. The great value of in-situ hy-bridization lies in possible association of the signal with cells of particular types or withhistopathological changes. In experimentally infected pigs, PCV nucleic acids weredetected by in-situ hybridization in macrophages, lymphocytes, endothelial cells andepithelial cells fromseveralorgans (Morozov et al., 1998).However,onlya fewnaturallyinfected pigs have been examined (Allan et al., 1998). Moreover, in these two studies,either RNA probes or whole genomic probes were used, instead of DNA probes. In-situ hybridization with RNA probes is generally more sensitive than hybridizationwith DNA probes but requires more exacting conditions (Brown, 1998). If the latterare not met, false negative results may occur. However, because PCV is a singlestranded DNA virus, DNA probes are not only as sensitive as RNA probes but alsomore reliable for diagnostic purpose. The objectives of the present study were (1) todevelop a non-radioactive digoxigenin-labelled DNA probe that would detect PCV,and (2) to use such a probe to examine the distribution of PCV in formalin-fixedparaffin wax-embedded tissues from weaned pigs with PMWS.
Materials and MethodsAnimals
Samples were obtained at necropsy from pigs submitted to the Department ofVeterinary Pathology of Seoul National University from January 1997 to October1998. Ten weaned pigs (nos 1–10; Table 1) in which PCV infection was diagnosedwere used in this study.
Positive control tissue came from a 56-day-old pig in which natural infection with PCV wasconfirmed immunohistochemically by Dr E. G. Clark, University of Saskatchewan. Negativecontrol tissues were collected from 1-day-old colostrum-deprived pigs that had not beenexposed to circovirus and also from a 5-day-old conventional pig infected with porcine re-productive and respiratory syndrome virus (PRRSV).

267In-situ Hybridization for Porcine Circovirus
Probe
A 530 base pair DNA fragment from an open reading frame 1 was used as a probe.The polymerase chain reaction (PCR) was carried out as previously described (Morozovet al., 1998). The sense and antisense primers were 5′-CAGCGGCAGCACCT-CGGCAGCGTCAGT-3′ and 5′-TCCAATCACGCTGCTGCAT CTTCCCGC-3′,respectively. The PCR products were purified with Wizard PCR Preps (PromegaBiotech, Madison, WI, USA). Purified PCR product was labelled by random primingwith digoxigenin-dUTP with a commercial kit (Boehringer Mannheim, Indianapolis,IN, USA), according to the manufacturer’s instructions. Although the primers usedwere based on a PCV type 2 sequence, the probe was capable of labelling both PCVtype 1 and PCV type 2. The negative PRRSV probe was prepared as describedpreviously (Cheon and Chae, 1999).
In-situ Hybridization
Tissues from each pig were collected in 10% neutral buffered formalin, and after 1 or 2days’ fixation were dehydrated through graded alcohols and a xylene step and embeddedin paraffin wax. Sections were then cut at 4 lm, placed on Superfrost/Plus slides (FisherScientific, Pittsburgh, PA, USA), and stored at room temperature. Just before use, sectionswere dewaxed in xylene and rehydrated in phosphate-buffered saline (PBS; pH 7·4,0·01 M) for 5 min. De-proteinization was carried out in 0·2 N HCl for 20 min at roomtemperature. Tissues were then digested at 37°C for 20 min in proteinase K (Gibco BRL,Grand Island, NY, USA) 300 lg/ml in PBS and fixed in paraformaldehyde 4% in PBSfor 10 min. After rinsing with PBS twice, the slides were acetylated in 300 ml of 0·1 mMtriethanolamine-HCl buffer (pH 8·0) to which 0·75 ml of acetic anhydride (0·25%) hadbeen added. After 5 min, a further 0·75 ml of acetic anhydride was added, and 5 minlater the slides were rinsed in 2× saline sodium citrate (SSC) (1× SSC contains 50 mMNaCl and 15 mM sodium citrate, pH 7·0). Hybridization was carried out overnight at40°C. The digoxigenin-labelled probe (50 ng) was diluted in 50 ll of the standardhybridization buffer, which consisted of 2× SSC containing deionized formamide 50%,salmon sperm DNA (Oncor, Gaithersburg, MD, USA) 10 mg, sodium dodecyl sulphate(SDS) 0·02%, and dextran sulphate solution (50% concentration) 50%; it was then heatedfor 10 min in a 95°C heating block, and quenched on ice. Approximately 75 ng ofdigoxigenin-labelled probe were added to standard hybridization buffer (70 ll), whichwas then layered over the section. The fluid was held in place by a coverslip (the edgesof which were sealed with rubber cement), heated for 10 min in a 95°C heating block,and then quenched on ice. After overnight hybridization, sections were thoroughlywashed, twice in 4× SSC for 5 min at room temperature, twice in 2× SSC for 5 minat 40°C, twice in 2× SSC for 5 min at room temperature, twice in 0·2× SSC for 5 minat 40°C, twice in 0·2× SSC for 5 min, once in maleic acid buffer (100 mM maleic acidand 150 mM NaCl, pH 7·5) for 5 min and once in 1× blocking reagent (BoehringerMannheim) for 40 min at room temperature.
For detection of hybridization, sections were incubated with anti-digoxigenin conjugatedwith alkaline phosphatase (Boehringer Mannheim) diluted 1 in 500 in 1× blockingreagent (Boehringer Mannheim). After three washes in buffer, substrate consisting ofnitroblue tetrazolium (NBT) and 5-bromocresyl-3-indolylphosphate (BCIP) was layeredover the sections. Colour was allowed to develop for 1–4 h in the dark, and developmentwas then stopped by dipping the slides briefly in tri-ethylenediaminetetraacetic acid(EDTA) buffer (10 mM Tris-HCl and 1 mM EDTA, pH 8·0). Sections were counterstainedwith methyl green 0·5%, and the slides were then washed with distilled water for 1 minand dried completely.
Results
In-situ hybridization results for the 10 infected pigs are presented in Table 1.The morphology of host cells was preserved despite the relatively high
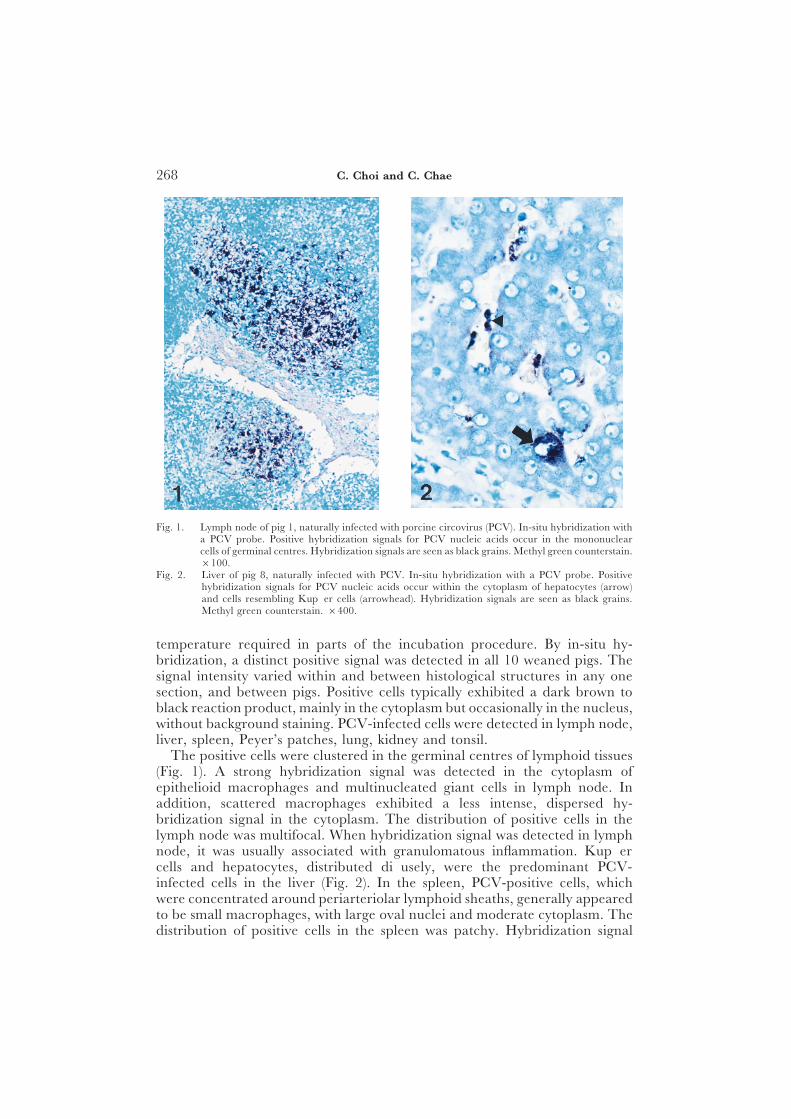
268 C. Choi and C. Chae
Fig. 1. Lymph node of pig 1, naturally infected with porcine circovirus (PCV). In-situ hybridization witha PCV probe. Positive hybridization signals for PCV nucleic acids occur in the mononuclearcells of germinal centres. Hybridization signals are seen as black grains. Methyl green counterstain.×100.
Fig. 2. Liver of pig 8, naturally infected with PCV. In-situ hybridization with a PCV probe. Positivehybridization signals for PCV nucleic acids occur within the cytoplasm of hepatocytes (arrow)and cells resembling Kupffer cells (arrowhead). Hybridization signals are seen as black grains.Methyl green counterstain. ×400.
temperature required in parts of the incubation procedure. By in-situ hy-bridization, a distinct positive signal was detected in all 10 weaned pigs. Thesignal intensity varied within and between histological structures in any onesection, and between pigs. Positive cells typically exhibited a dark brown toblack reaction product, mainly in the cytoplasm but occasionally in the nucleus,without background staining. PCV-infected cells were detected in lymph node,liver, spleen, Peyer’s patches, lung, kidney and tonsil.
The positive cells were clustered in the germinal centres of lymphoid tissues(Fig. 1). A strong hybridization signal was detected in the cytoplasm ofepithelioid macrophages and multinucleated giant cells in lymph node. Inaddition, scattered macrophages exhibited a less intense, dispersed hy-bridization signal in the cytoplasm. The distribution of positive cells in thelymph node was multifocal. When hybridization signal was detected in lymphnode, it was usually associated with granulomatous inflammation. Kupffercells and hepatocytes, distributed diffusely, were the predominant PCV-infected cells in the liver (Fig. 2). In the spleen, PCV-positive cells, whichwere concentrated around periarteriolar lymphoid sheaths, generally appearedto be small macrophages, with large oval nuclei and moderate cytoplasm. Thedistribution of positive cells in the spleen was patchy. Hybridization signal

269In-situ Hybridization for Porcine Circovirus
was detected in the cells of Peyer’s patches; these cells generally had largeoval nuclei and abundant cytoplasm. Comparison with haematoxylin andeosin-stained sections from the same block indicated that many of the positivecells in Peyer’s patches were macrophages. In the lung, hybridization signalwas occasionally detected in cells that resembled interstitial macrophages. Inthe kidney, hybridization signal was detected, although only rarely, in therenal tubular epithelial cells and interstitial macrophages.
The cDNA probes for PRRSV were consistently negative in all tissuestested. Sections from the two negative control pigs showed no hybridizationsignal for PCV.
Discussion
The results demonstrated that PCV nucleic acids can be detected in formalin-fixed, paraffin wax-embedded tissues of weaned pigs with a non-radioactivedigoxigenin-labelled probe. PCV replicated primarily within macrophages inlymph nodes, liver, spleen, Peyer’s patches, lung, kidney and tonsil. Noattempt was made to identify the cell types other than by visual examination;consequently, confirmation by electron microscopy or other methods is neededto define unequivocally the target cells of PCV. In pigs 1, 2 and 3 (Table 1)PCV nucleic acid was not detected in lung, and in pigs 4 and 5 it was notdetected in tonsil. These discrepant results may have been due to infectionwith different strains of PCV or to differences in the stage of infection, butfurther studies are needed.
PCV type 1 is non-pathogenic for pigs (Tischer et al., 1986; Allan et al.,1995). In the present study and previous studies on PMWS (Allan et al., 1998;Morozov et al., 1998), infected cells included macrophages, hepatocytes, andrenal tubular epithelial cells. This is in contrast to PCV type 1, which infectedonly cells of mononuclear phagocyte lineage (Allan et al., 1994, 1995; McNeillyet al., 1996). The abundant presence of PCV in several tissues in the presentstudy strongly suggests that PCV type 2 plays an important role in PMWS;however, additional studies are needed to clarify fully the aetiology of PMWS.
AcknowledgmentThis study was supported by the contract research fund of the Research Institute forVeterinary Science (RIVS), College of Veterinary Medicine, Seoul National University,Republic of Korea.
ReferencesAllan, G. M., McNeilly, F., Cassidy, J. P., Reilly, G. A. C., Adair, B., Ellis, J. A. and
McNulty, M. S. (1995). Pathogenesis of porcine circovirus, experimental infectionsof colostrum deprived piglets and examination of pig fetal material. VeterinaryMicrobiology, 44, 49–64.
Allan, G. M., McNeilly, F., Kennedy, S., Daft, B., Clarke, E. G., Ellis, J. A., Haines,D. M., Meehan, B. M. and Adair, B. M. (1998). Isolation of porcine circovirus-like viruses from pigs with a wasting disease in the USA and Europe. Journal ofVeterinary Diagnostic Investigation, 10, 3–10.

270 C. Choi and C. Chae
Allan, G. M., Phenix, K. V., Todd, D. and McNulty, M. S. (1994). Some biologicaland physico-chemical properties of porcine circovirus. Journal of Veterinary MedicineB, 41, 17–26.
Brown, C. (1998). In situ hybridization with riboprobes: an overview for veterinarypathologists. Veterinary Pathology, 35, 159–167.
Cheon, D.-S. and Chae, C. (1999). Distribution of a Korean strain of porcinereproductive and respiratory syndrome virus in experimentally infected pigs, asdemonstrated immunohistochemically and by in-situ hybridization. Journal ofComparative Pathology, 120, 79–88.
Clark, E. G. (1997). Post-weaning wasting syndrome. Proceedings of the American Associationof Swine Practitioners, 28, 499–501.
Ellis, J. A., Hassard, L., Clark, E., Harding, J., Allan, G., Willson, P., Strokappe, J.,Martin, K., McNeilly, F., Meehan, B., Todd, D. and Haines, D. (1998). Isolationof circovirus-like virus from lesions with post-weaning multisystemic wastingsyndrome. Canadian Veterinary Journal, 39, 44–51.
LeCann, P., Albina, E., Madec, F., Cariolet, R. and Jestin, A. (1997). Piglet wastingdisease. Veterinary Record, 141, 600.
Lukert, P., de Boer, G. F., Dale, J. L., Keese, P., McNulty, M. S., Randles, J. W.and Tischer, I. (1995). The Circoviridae. In: Virus Taxonomy, 1st Edit., F. A.Murphy, C. M. Fauquet, D. H. L. Bishop, S. A. Ghabrial, A. W. Jarvis, G. P.Martelli, M. A. Mayo and M. D. Summers, Eds, Springer-Verlag, Vienna andNew York, pp. 166–168.
McNeilly, F., Allan, G. M., Foster, J. C., Adair, B. M. and McNulty, M. S. (1996).Effect of porcine circovirus infection on porcine alveolar macrophage infection.Veterinary Immunology and Immunopathology, 49, 295–306.
Meehan, B. M., Creelan, J. L., McNulty, M. S. and Todd, D. (1997). Sequence ofporcine circovirus DNA: affinities with plant circoviruses. Journal of General Virology,78, 221–227.
Meehan, B. M., McNeilly, F., Todd, D., Kennedy, S., Jewhurst, V. A., Ellis, J. A.,Hassard, L. E., Clark, E. G., Haines, D. M. and Allan, G. M. (1998). Charac-terization of novel circovirus DNAs associated with wasting syndromes in pigs.Journal of General Virology, 79, 2171–2179.
Morozov, I., Sirinarumitr, T., Sorden, S. D., Halbur, P. G., Morgan, M. K., Yoon,K-J. and Paul, P. S. (1998). Detection of a novel strain of porcine circovirusin pigs with postweaning multisystemic wasting syndrome. Journal of ClinicalMicrobiology, 36, 2535–2541.
Segales, J., Sitjar, M., Domingo, M., Dee, S., Del Pozo, M., Noval, R., De Las Heras,A., Sacristan, C., Ferro, A. and Latimer, K. S. (1997). First report of post-weaning multisystemic wasting syndrome in Spain. Veterinary Record, 141, 600–601.
Tischer, I., Mields, W., Wolff, D., Vagt, M. and Greim, W. (1986). Studies onepidemiology and pathogenicity of porcine circovirus. Archives of Virology, 91,271–276.
Tischer, I., Rasch, R. and Tochtermann, G. (1974). Characterization of papovavirus-and piconavirus-like particles in permanent pig kidney cell lines. Zentralblatt fuerBakteriologie, Mikrobiologie und Hygiene, Series A, 226, 153–167.
CReceived, November 30th, 1998Accepted, April 12th, 1999 D





























