Embed Size (px)
Citation preview

of August 26, 2018.This information is current as
Myeloid DifferentiationIkaros Isoform X Is Selectively Expressed in
Payne and Gay M. CrooksZhu, Natasha S. Barteneva, Lora W. Barsky, Marvin A. Kimberly J. Payne, Grace Huang, Eva Sahakian, Judy Y.
http://www.jimmunol.org/content/170/6/3091doi: 10.4049/jimmunol.170.6.3091
2003; 170:3091-3098; ;J Immunol
Referenceshttp://www.jimmunol.org/content/170/6/3091.full#ref-list-1
, 34 of which you can access for free at: cites 53 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2003 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Ikaros Isoform X Is Selectively Expressed in MyeloidDifferentiation 1
Kimberly J. Payne,2* Grace Huang,* Eva Sahakian,† Judy Y. Zhu,* Natasha S. Barteneva,*Lora W. Barsky,* Marvin A. Payne, † and Gay M. Crooks*
The Ikaros gene is alternately spliced to generate multiple DNA-binding and nonbinding isoforms that have been implicated asregulators of hematopoiesis, particularly in the lymphoid lineages. Although early reports ofIkaros mutant mice focused onlymphoid defects, these mice also show significant myeloid, erythroid, and stem cell defects. However, the specific Ikaros proteinsexpressed in these cells have not been determined. We recently described Ikaros-x (Ikx), a new Ikaros isoform that is the pre-dominant Ikaros protein in normal human hematopoietic cells. In this study, we report that the Ikx protein is selectively expressedin human myeloid lineage cells, while Ik1 predominates in the lymphoid and erythroid lineages. Both Ik1 and Ikx proteins areexpressed in early human hematopoietic cells (Lin�CD34�). Under culture conditions that promote specific lineage differentiation,Ikx is up-regulated during myeloid differentiation but down-regulated during lymphoid differentiation from human Lin �CD34�
cells. We show that Ikx and other novel Ikaros splice variants identified in human studies are also expressed in murine bonemarrow. In mice, as in humans, the Ikx protein is selectively expressed in the myeloid lineage. Our studies suggest that Ikarosproteins function in myeloid, as well as lymphoid, differentiation and that specific Ikaros isoforms may play a role in regulatinglineage commitment decisions in mice and humans.The Journal of Immunology, 2003, 170: 3091–3098.
T he Ikaros protein (also designated Lyf-1) (1) and gene (2)were first identified using strategies designed to detecttranscription factors that regulate lymphoid-specific
genes, leading to the conclusion that Ikaros was a lymphoid-spe-cific transcription factor. Subsequent studies provided evidencethat the role of Ikaros proteins in normal hematopoiesis extendedbeyond the lymphoid lineages (3–6). In addition, Ikaros proteinswere shown to act as both positive (5, 7–12) and negative (5,13–21) regulators of transcription.
The murine Ikaros gene was initially reported to contain sevenexons (7) that were alternately spliced to generate transcripts formultiple DNA-binding and nonbinding Ikaros isoforms (Fig. 1A)(3, 7, 22, 23). Human and murine Ikaros were shown to be highlyhomologous, and isoforms corresponding to murine Ik1 throughIk6 (Fig. 1A) were detected in human cells (23). This suggestedthat Ikaros mRNA splicing patterns might be identical in mice andhumans. However, subsequent studies of human Ikaros expressionin leukemia samples (24–26) and in normal hematopoietic cells
(27–29) identified transcripts for multiple Ikaros splice variantsthat had not been reported in murine studies. Our studies showedthat Ikaros splice variants with a 60-base insertion following exon2 (Fig. 1B) were expressed at the protein level in normal humanhematopoietic cells (29). We also detected transcripts for Ikaros-x(Ikx; Fig. 1C), a new Ikaros isoform not previously described inmice or humans. We found Ikx to be the predominant Ikaros pro-tein in normal human umbilical cord blood (CB) and bone marrow(BM) cells, although we observed little Ikx in T and B lymphoidcell lines (29). Murine hematopoietic cells have not been examinedfor expression of Ikx and the other novel Ikaros isoforms identifiedin human studies.
Studies of mutant mice implicated Ikaros proteins as regulatorsof critical events in hematopoietic differentiation and proliferation,particularly in the lymphoid lineages. Ikaros-null mice fail to pro-duce B cells and generate T cells only postnatally (30). Mice witha defect in the Ikaros DNA-binding domain show more severelymphoid defects, including a complete loss of T, B, and NK cells(31). In addition, mice with reduced Ikaros DNA-binding activityshow abnormal T cell proliferation and differentiation (32–34).
Although the initial reports of Ikaros mutant mice focused onlymphoid defects, (30, 31), these mice display multiple hemato-poietic abnormalities (4, 35, 36), including myeloid and hemato-poietic stem cell (HSC) defects (4, 30, 31, 36) and aberrant eryth-ropoiesis (6, 30, 31). Ikaros proteins have now been shown to actas the DNA binding component of the PYR chromatin remodelingcomplex that functions in globin switching (5, 6). A recent studyshows that Ikaros is important at early stages in granulocyte dif-ferentiation (37). These findings are consistent with erythroid andmyeloid defects exhibited by Ikaros mutant mice (4, 30, 31).
Reports of Ikaros isoform expression in primary cells outsidethe lymphoid lineages are limited to a few studies that examinedIkaros mRNA expression by RT-PCR (3, 8, 27). However, ourstudies have shown a poor correlation between the expression ofIkaros proteins and Ikaros mRNA, as assessed by RT-PCR (29).The expression of Ikaros isoforms, at the protein level, has not
*Division of Research Immunology/Bone Marrow Transplantation, Childrens Hos-pital Los Angeles, Los Angeles, CA 90027; and †Department of Chemistry and Bio-chemistry, La Sierra University, Riverside CA 92515
Received for publication October 9, 2002. Accepted for publication January 9, 2003.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work is supported by National Institutes of Health Grants R01DK54567,2P50HL54850, and P01CA59318 (to G.M.C.), National Research Service Award1F32DK10101 (to K.J.P.), a fellowship from Childrens Hospital Los Angeles Re-search Institute (to K.J.P.), and a grant from the College of Arts and Sciences, LaSierra University (to M.A.P.). G.M.C. is a Scholar of the Leukemia and LymphomaSociety.2 Address correspondence and reprint requests to Dr. Kimberly J. Payne, ChildrensHospital Los Angeles, Division of Research Immunology/Bone Marrow Transplan-tation, Mail Stop #62, 4650 Sunset Boulevard, Los Angeles, CA 90027. E-mail ad-dress: [email protected] Abbreviations used in this paper: Ik, Ikaros isoform; BM, bone marrow; CB, um-bilical cord blood; HSC, hematopoietic stem cell; CTS, C-terminal sequence; NTS,N-terminal sequence.
The Journal of Immunology
Copyright © 2003 by The American Association of Immunologists, Inc. 0022-1767/03/$02.00
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

been examined in primary cells outside the lymphoid lineages, andlittle is known of the role of Ikaros proteins in the myeloid lineage.
The lack of Ikx in human lymphoid cell lines led us to ask whatcells might be responsible for the abundance of Ikx proteins thatwe detected in human CB and BM (29). One possibility was thatIkx was expressed by erythroid and/or myeloid lineage cells. Thepredominance of Ikx that we observed in normal human hemato-poietic cells also raised the question of whether Ikx and othernovel Ikaros splice variants identified in human studies might beexpressed in the mouse. This is particularly important because ex-periments to determine the mechanisms of Ikaros function haverelied almost exclusively on the murine model. Neither murine norhuman studies have examined differential expression of Ikaros pro-teins between normal primary cells of the various hematopoieticlineages or at different points in differentiation. Consequently,questions concerning potential roles for specific Ikaros proteins inhematopoietic lineage commitment have not been addressed.
Materials and MethodsCell sources and preparation
Human CB and BM were collected according to guidelines approved bythe Childrens Hospital Los Angeles Committee on Clinical Investigation(Investigational Review Board) and mononuclear cells were isolated aspreviously described (38).
Nucleated erythroid lineage cells were isolated from mononuclear cellsthat had been treated with PharM Lyse ammonium chloride lysing solution(BD PharMingen, San Diego, CA) to lyse any remaining erythrocytes.CD66B� granulocytes were isolated (within 6 h of delivery) from fresh CBcells located below the buffy layer after Ficoll-Hypaque separation. Theremaining human hematopoietic cells were isolated from total CB or BMmononuclear cells.
Murine BM, spleen, and thymus were obtained from C57BL/6 mice(The Jackson Laboratory, Bar Harbor, ME), aged 6–12 wk, and rested forat least 5 days after arrival before organs were harvested.
The murine S17 stromal cell line was a generous gift from Dr. K. Dorshkind(University of Southern California, Los Angeles, CA). The Jurkat E6-1cell line was obtained from American Type Tissue Culture Collection(Manassas, VA).
Isolation of hematopoietic populations
Murine and human hematopoietic cells were incubated with Abs (see be-low) for 20–30 min on ice, washed, and then cell populations were isolatedby FACS sorting or magnetic separation based on surface Ag expression.
For RT-PCR, cells were isolated by FACS (FACSVantage; BD Bio-sciences, San Jose, CA) based on forward scatter and side scatter gatescharacteristic of living mononuclear cells and surface Ag staining as indi-cated below (typical purity of 99% or greater).
All HSC and progenitor populations were FACS sorted from CD34�-enriched mononuclear cells. CD34� enrichment was performed using theMiniMacs CD34 progenitor isolation kit (Miltenyi Biotec, Auburn, CA)according to the manufacturer’s instructions.
For immunoblot analysis, mature cells of the hematopoietic lineageswere isolated using the MiniMACS magnetic separation system (MiltenyiBiotec). Murine and human B lineage cells were isolated using respective,species-specific, CD19 MicroBeads (Miltenyi Biotec) to a purity of �99%.The remaining lineages were isolated by positive selection (typical purityof 95% or greater) using anti-FITC or anti-PE MicroBeads (Miltenyi Bio-tec) following staining with FITC- or PE-conjugated Abs as describedbelow.
Abs used to isolate cells from human hematopoietic lineages were thefollowing: T cells, anti-CD3 FITC or anti-CD3 PE (clone Sk7); B cells,anti-CD19 PE (clone 4G7); NK cells, anti-CD56 PE (clone NCAM16.2);myeloid cells, anti-CD14 FITC (clone M0P9), anti-CD15 FITC (cloneMMA), anti-CD66B FITC (clone G10F5), and anti-CD11B PE (cloneD12) (all from BD Biosciences); and erythroid cells, anti-glycophorin AFITC or anti-glycophorin A PE (clone KC16; Immunotech, Westbrook,ME). Abs used to isolate cells from murine hematopoietic lineages werethe following: T cells, anti-CD3 FITC (clone 17A2); B cells, anti-CD19 PE(1D3); and myeloid cells, anti-CD11B PE (M1/70) (all from BD Bio-sciences). Abs used to isolated CD34� subsets were the following: anti-CD34 allophycocyanin (clone 8G12; BD Biosciences) and biotinylated anti-CD38 (clone HIT2; Caltag Laboratories, Burlingame, CA), visualized withstreptavidin red 613 (Invitrogen, Carlsbad, CA). Lin�CD34� cells were
isolated using a mixture of Abs directed against glycophorin A, CD3,CD19, CD56, CD14, CD15, and CD66B (Ab clones and sources listedabove).
Culture conditions
Freshly sorted Lin�CD34� cells were cultured in vitro under conditionsthat promote lymphoid differentiation or myeloid differentiation (39). Lym-phoid supporting conditions were as follows: coculture on the murine S17stromal cell line (a generous gift from Dr. K. Dorshkind) in R10 medium(RPMI 1640 medium (Irvine Scientific, Santa Ana, CA) with 5% heat-inactivated FCS, 0.5% penicillin/streptomycin, 1% L-glutamine, and 2-ME(Sigma-Aldrich, St. Louis, MO)) supplemented with IL-3 at 5 ng/ml (R&DSystems, Minneapolis, MN) and Flt-3 ligand at 50 ng/ml (Immunex, Se-attle, WA) for the first 3 days, then with Flt-3 ligand at 50 ng/ml alone forthe remaining culture period. Myeloid supporting conditions were as fol-lows: coculture on the murine S17 stromal cell line in BBMM medium(IMDM (BioWhittaker, Walkersville, MD) with 20% heat-inactivatedFCS, 10% BSA, 0.5% penicillin/streptomycin, 1% L-glutamine, 10�4
mol/L 2-ME, and 10�6 mol/L hydrocortisone (to prevent lymphoid prolif-eration; Sigma-Aldrich)), supplemented with IL-3 at 5 ng/ml, IL-6 at 5ng/ml, and stem cell factor at 25 ng/ml (R&D Systems).
RNA extraction and cDNA preparation
Total RNA was obtained using STAT 60 (Tel-Test, Friendswood, TX) asper manufacturer’s directions. cDNA was prepared using oligo(dT) primers(Pharmacia Biotech, Uppsala, Sweden) and the Omniscript kit (Qiagen,Valencia, CA) as per manufacturer’s directions. cDNA from equivalentcell numbers was used for RT-PCR analysis of human Ikaros expression inhematopoietic lineages and in progenitor populations. Using primers andconditions described previously (40), integrity of cDNA was assessed byRT-PCR amplification of �2-microglobulin from the same cDNA mastermix used to assess Ikaros expression.
RT-PCR analysis
cDNA was subjected to PCR using HotStarTaq (Qiagen) as per manufac-turer’s directions. Homologous primer pairs were designed to amplify se-lected murine and human Ikaros transcripts (29), including splice formswith the 60-base insertion following exon 2.
Primers and conditions for amplification of human Ikaros cDNAs wereas described previously (29). Primers for amplification of mouse IkaroscDNAs were the following: forward, 5�-TGAGCCCATGCCTGTCCCTGAG-3�, and reverse, 5�-GGTCTTCTGCCATCTCGTTGTGGTTA-3�.PCR conditions for mouse primers were as follows: 15-min, 95° hot startfollowed by 35 cycles of 1 min and 30 s at 95°, 30 s at 68°, and 3 min at72°, followed by a final 10-min, 72° extension and cooling to 4°. To pre-vent heteroduplex formation between PCR products generated from dif-ferent Ikaros isoforms as primer concentrations decreased, 20 additionalpicomoles each of forward and reverse primers were added two cyclesbefore completion of the PCR.
Sequencing of RT-PCR products
Individual PCR products for sequencing were obtained by cutting individ-ual bands from gel and purifying using UltraClean GelSpin DNA purifi-cation kit (MO BIO Laboratories, Solana Beach, CA) or by generatingsingle PCR products in a second PCR (identical with above, except run for30 cycles) performed on samples aspirated by syringe from individualbands. Sequencing was performed by the University of Southern Califor-nia/Norris Comprehensive Cancer Center Microchemical Core Facility atthe Keck School of Medicine (Los Angeles, CA) or the Core Facility at theCenter for Molecular Biology and Gene Therapy (Loma Linda University,Loma Linda, CA).
Immunoblots
For preparation of cell lysates, washed cells were frozen at �80° andthawed, on ice, in cold universal immunoprecipitation buffer (50 mM Tris,150 mM NaCl, 2 mM EDTA, 2 mM EGTA, 25 mM NaF, 25 mM �-glyc-erol phosphate, and 0.3% IGEPAL in water), brought up to 1 mM PMSF,1 �g/ml leupeptin, and 1 �g/ml aprotinin immediately before use. Sus-pended cells were sonicated on ice for �10 s, checked for complete lysisby microscope and then centrifuged at 14,000 rpm for 10 min at 4° to pelletmembranes. Supernatants were diluted in NuPage sample buffer (Invitro-gen) and immediately run on gel or stored at �80°. Lysates were run alongwith Seeblue m.w. markers on a NuPage 10% Bis-Tris gel with MOPsbuffer using the XCell SureLock system (Invitrogen) and transferred toImmobilon-P membrane (Millipore, Bedford, MA) using the XCell II BlotModule (Invitrogen), all performed as per manufacturer’s instructions.
3092 Ikx IS SELECTIVELY EXPRESSED IN MYELOID DIFFERENTIATION
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Ikaros was detected using Ikaros M-20 and Ikaros E-20 (polyclonal goatAbs specific to the N and C termini, respectively, common to all Ikarosisoforms (Santa Cruz Biotechnology, Santa Cruz, CA)), anti-IkH (poly-clonal rabbit Ab specific to residues encoded by the 60-base insert follow-ing Ikaros exon 2), or the mouse mAb 2A9 (specific to exon 3 of the Ikarosprotein) (generous gifts kindly provided by the laboratory of S. Smale(Howard Hughes Medical Institute, University of California, Los Angeles,School of Medicine, Los Angeles CA)). Blots were developed using theECL�Plus Western blotting detection system and accompanying Abs(Amersham, Arlington Heights, IL) or anti-goat-HRP (Santa Cruz Biotech-nology) as per manufacturer’s protocol, with the exception that blockingreagent was present continually during Ab incubation.
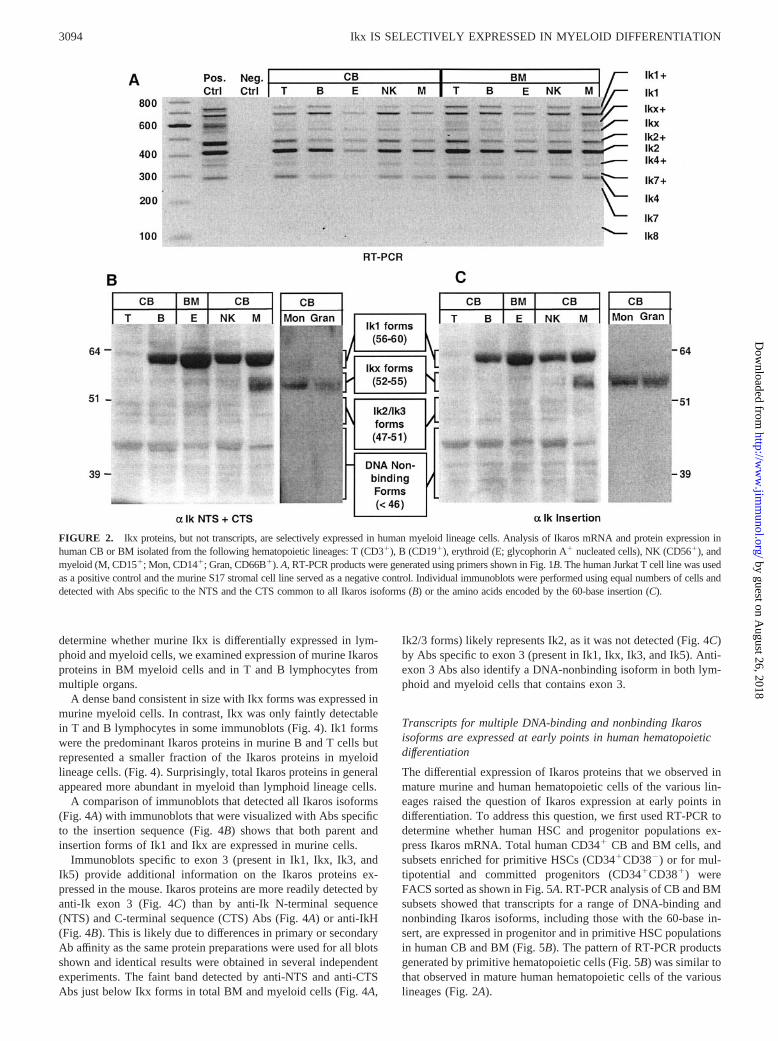
ResultsIkx proteins, but not transcripts, are selectively expressed inhuman myeloid lineage cells
Our previous studies showed that Ikx (Fig. 1C) is the predominantIkaros protein in human CB and BM, but not in human lymphoidcell lines (29). One explanation for these observations is that Ikxis selectively expressed in nonlymphoid lineages. To determinewhether this explanation is correct, we examined expression ofIkaros mRNA and protein in human lymphoid, myeloid, and nu-cleated erythroid lineage cells.
Multilineage RT-PCR analysis of Ikaros mRNA expression inhuman CB and BM showed a similar pattern of products for all ofthe hematopoietic lineages (Fig. 2A). PCR primers (Fig. 1B) gen-erated products representing multiple DNA binding and nonbind-ing Ikaros isoforms, including many with the 60-base insertion, inFACS-sorted cells from the T, B, NK, erythroid, and myeloid lin-eages (Fig. 2A). Isoforms with the 60-base insertion (insertionforms) were designated as Ik1�, Ikx�, Ik2�, etc. The pattern ofRT-PCR products generated in the individual lineages matched thepattern that we had previously observed in total human BM andCB (29).
Contrary to what was observed in RT-PCR analysis of mRNAexpression, immunoblots showed that Ikaros isoforms are differ-entially expressed at the protein level in the hematopoietic lin-eages. Ik1 proteins are predominant in B, NK, and nucleated ery-
throid lineage cells (Fig. 2B). Human T cells isolated from CBshowed only very faint bands representing multiple DNA-bindingand nonbinding Ikaros proteins (Fig. 2B). Ikx proteins are selec-tively expressed in myeloid lineage cells (Fig. 2B). The CD15�
cells isolated from CB include monocytic and granulocytic subsets(41); however, the CD15 Ag is a differentiation and tumor-asso-ciated Ag that has been detected on nonhematopoietic cells (42,43). To more precisely define Ikaros protein expression within themyeloid lineages, we isolated CD14� (primarily monocytes andmacrophages (44)) and CD66B� (granulocytes (45, 46)) cells fromCB. Ikx predominates in both the CD14� and CD66B� myeloidpopulations (Fig. 2B).
Ik1 predominated in the B, NK, and nucleated erythroid lineagecells we examined. However, cells from these three lineages com-bined generally comprised �20% (and in some cases �10%) ofmononuclear cells in CB and only about a third of those in BM(data not shown). This explains the predominance of Ikx that weobserved in our previous studies of total human CB and BM mono-nuclear cells (29).
Immunoblots specific to the amino acids encoded by the 60-baseinsertion show that Ikaros insertion forms follow a pattern of lin-eage-specific expression that mirrors that observed for parentIkaros isoforms: Ikx� is selectively expressed in myeloid cellswhile Ik1� predominates in B, NK, and erythroid lineage cells(Fig. 2C).
These data show that the Ikx and Ikx� proteins, previously re-ported to predominate in human BM and CB cells (29), are selec-tively expressed in myeloid lineage cells. This pattern of proteinexpression was not predicted based on RT-PCR assessment ofIkaros mRNA.
Murine hematopoietic cells express transcripts for Ikx
Given the predominance of Ikx in human hematopoietic cells, wewanted to know whether murine hematopoietic cells express Ikxand the novel Ikaros insertion forms identified in human studies.Murine and human primer pairs were designed to amplify selectedIkaros transcripts with the 60-base insertion following exon 2 (Fig.1B). DNA sequencing confirmed the identity of murine RT-PCRproducts consistent in size with Ik1, Ik2, Ik4, Ik8, and the newIkaros isoform, Ikx. (Fig. 3A). RT-PCR products consistent in sizewith Ikx� and, in some cases, Ik1� were also detected (Fig. 3A).However, we were unable to isolate RT-PCR products from mu-rine insertion forms for DNA sequencing.
Ikx proteins predominate in mouse BM
Immunoblots specific to both murine and human Ikaros proteinsshow a similar pattern of Ikaros expression in mouse and humanBM (Fig. 3B). The predominant Ikaros protein is detectable as abroad dense band consistent in size with Ikx forms (Fig. 3B). Afainter band representing Ik1 forms is also detectable in both mu-rine and human cells (Fig. 3B). Although we were unable to iden-tify RT-PCR products for murine Ikaros insertion forms by DNAsequencing, Abs specific to the insert show that the Ikaros insertionforms are expressed at the protein level in mouse BM cells (Fig.3C). These data show that mice, like humans, express proteinsrepresenting the new Ikaros isoform, Ikx, and Ikaros splice formswith the 60-base insertion. Furthermore, Ikx forms are the pre-dominant Ikaros proteins in mouse, as well as human, BM.
Murine Ikx proteins are differentially expressed in lymphoid andmyeloid lineage cells
Previous studies of Ikaros protein expression in the mouse havefocused on the lymphoid lineage and failed to identify Ikx. To
FIGURE 1. Diagram of Ikaros isoforms and placement of RT-PCRprimers. A, Alternate splicing of exons 1–7 of the Ikaros gene gives rise toisoforms Ik1 through Ik8. Zinc fingers are represented as vertical bars. B,Placement of mouse and human primers designed to detect Ik1, Ikx, Ik2,Ik4, Ik7, Ik8, and variants of these isoforms (designated Ik1�, Ikx�, Ik2�,etc.) with the 60-base insertion following exon 2. C, Diagram of the newIkaros isoform, Ikx.
3093The Journal of Immunology
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
determine whether murine Ikx is differentially expressed in lym-phoid and myeloid cells, we examined expression of murine Ikarosproteins in BM myeloid cells and in T and B lymphocytes frommultiple organs.
A dense band consistent in size with Ikx forms was expressed inmurine myeloid cells. In contrast, Ikx was only faintly detectablein T and B lymphocytes in some immunoblots (Fig. 4). Ik1 formswere the predominant Ikaros proteins in murine B and T cells butrepresented a smaller fraction of the Ikaros proteins in myeloidlineage cells. (Fig. 4). Surprisingly, total Ikaros proteins in generalappeared more abundant in myeloid than lymphoid lineage cells.
A comparison of immunoblots that detected all Ikaros isoforms(Fig. 4A) with immunoblots that were visualized with Abs specificto the insertion sequence (Fig. 4B) shows that both parent andinsertion forms of Ik1 and Ikx are expressed in murine cells.
Immunoblots specific to exon 3 (present in Ik1, Ikx, Ik3, andIk5) provide additional information on the Ikaros proteins ex-pressed in the mouse. Ikaros proteins are more readily detected byanti-Ik exon 3 (Fig. 4C) than by anti-Ik N-terminal sequence(NTS) and C-terminal sequence (CTS) Abs (Fig. 4A) or anti-IkH(Fig. 4B). This is likely due to differences in primary or secondaryAb affinity as the same protein preparations were used for all blotsshown and identical results were obtained in several independentexperiments. The faint band detected by anti-NTS and anti-CTSAbs just below Ikx forms in total BM and myeloid cells (Fig. 4A,
Ik2/3 forms) likely represents Ik2, as it was not detected (Fig. 4C)by Abs specific to exon 3 (present in Ik1, Ikx, Ik3, and Ik5). Anti-exon 3 Abs also identify a DNA-nonbinding isoform in both lym-phoid and myeloid cells that contains exon 3.
Transcripts for multiple DNA-binding and nonbinding Ikarosisoforms are expressed at early points in human hematopoieticdifferentiation
The differential expression of Ikaros proteins that we observed inmature murine and human hematopoietic cells of the various lin-eages raised the question of Ikaros expression at early points indifferentiation. To address this question, we first used RT-PCR todetermine whether human HSC and progenitor populations ex-press Ikaros mRNA. Total human CD34� CB and BM cells, andsubsets enriched for primitive HSCs (CD34�CD38�) or for mul-tipotential and committed progenitors (CD34�CD38�) wereFACS sorted as shown in Fig. 5A. RT-PCR analysis of CB and BMsubsets showed that transcripts for a range of DNA-binding andnonbinding Ikaros isoforms, including those with the 60-base in-sert, are expressed in progenitor and in primitive HSC populationsin human CB and BM (Fig. 5B). The pattern of RT-PCR productsgenerated by primitive hematopoietic cells (Fig. 5B) was similar tothat observed in mature human hematopoietic cells of the variouslineages (Fig. 2A).
FIGURE 2. Ikx proteins, but not transcripts, are selectively expressed in human myeloid lineage cells. Analysis of Ikaros mRNA and protein expression inhuman CB or BM isolated from the following hematopoietic lineages: T (CD3�), B (CD19�), erythroid (E; glycophorin A� nucleated cells), NK (CD56�), andmyeloid (M, CD15�; Mon, CD14�; Gran, CD66B�). A, RT-PCR products were generated using primers shown in Fig. 1B. The human Jurkat T cell line was usedas a positive control and the murine S17 stromal cell line served as a negative control. Individual immunoblots were performed using equal numbers of cells anddetected with Abs specific to the NTS and the CTS common to all Ikaros isoforms (B) or the amino acids encoded by the 60-base insertion (C).
3094 Ikx IS SELECTIVELY EXPRESSED IN MYELOID DIFFERENTIATION
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Ik1 and Ikx protein levels are differentially modulated duringmyeloid and lymphoid differentiation
Our data demonstrate that the ability to generate transcripts for arange of DNA-binding and nonbinding Ikaros isoforms is present
very early in hematopoietic differentiation. However, we have ob-served a poor correlation between the expression of Ikaros tran-scripts and proteins. Therefore, to determine which of these tran-scripts are expressed as proteins, we examined Ikaros proteinexpression in early hematopoietic cells and the progeny they gen-erate during lymphoid and myeloid differentiation.
FACS-sorted CB cells that lack markers of lineage-committedcells (CD3, CD19, CD56, glycophorin A, CD15, CD14, andCD66B) but express CD34 (Lin�CD34�; Fig. 6A) were used toprepare lysates for immunoblot analysis or placed into culture con-ditions that selectively support either lymphoid or myeloid lineage
FIGURE 4. Murine Ikx and Ikx� are differentially expressed in lymphoid and myeloid lineage cells. Immunoblots performed using equal numbers ofcells from mouse BM, spleen, or thymus (Thy). B, T, and myeloid (M) lineage cells were isolated from indicated tissues based on expression of CD19,CD3, or CD11B, respectively. Ikaros proteins were detected with Abs specific to the NTS and the CTS common to all Ikaros isoforms (A), the amino acidsencoded by the 60-base insertion (B), or exon 3 common to Ik1, Ikx, Ik3, and Ik5 (C).
FIGURE 3. Transcripts and proteins for novel Ikaros isoforms, includ-ing Ikx, are expressed in mouse BM. A, RT-PCR analysis of Ikaros mRNAexpression in human (Hu) and mouse (M) BM (see Fig. 1B for primerlocation). Immunoblots performed using equal numbers of murine or hu-man BM cells detected with Abs specific to the NTS and the CTS commonto all Ikaros isoforms (B) or the amino acids encoded by the 60-base in-sertion (C).
FIGURE 5. Transcripts for multiple DNA-binding and nonbindingIkaros isoforms are expressed at early points in human hematopoietic dif-ferentiation. A, Gates used for FACS sorting human progenitor and HSC-enriched populations. B, RT-PCR analysis of FACS-sorted populationsfrom CB or BM (see Fig. 1B for primer placement).
3095The Journal of Immunology
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

differentiation (Fig. 6B). After 18 days in culture, lymphoid andmyeloid progeny were harvested for immunoblot analysis.
Unlike total CB cells which predominantly express Ikx, freshlyisolated Lin�CD34� cells express both Ik1 and Ikx (Fig. 6C).When Lin�CD34� cells were placed in culture conditions thatselectively support myeloid differentiation, Ikx expression was up-regulated in their progeny (Fig. 6C). In contrast, progeny generatedin culture conditions that promote lymphoid differentiation showeddown-regulation of Ikx (Fig. 6C). Thus, both Ik1 and Ikx are ex-pressed in early hematopoietic cells and the expression of Ikx is mod-ulated as cells undergo lineage commitment and differentiation.
DiscussionIn this report, we describe a comprehensive analysis of IkarosmRNA and protein expression during normal human hematopoi-esis. Our studies show that Ikaros protein expression follows alineage- and stage-specific pattern of expression that correlatespoorly with Ikaros mRNA expression as assessed by RT-PCR.Both Ik1 and Ikx proteins are expressed early in hematopoiesis.Thereafter, Ikx is selectively expressed in the myeloid lineage,while Ik1 predominates in erythroid and most lymphoid lineagecells. The expression pattern of murine Ikaros proteins, includingIkx, in lymphoid and myeloid cells is similar to that observed forhuman Ikaros.
In contrast to expression at the protein level, RT-PCR analysisof human Ikaros mRNA shows a similar pattern of expression forDNA-binding and nonbinding Ikaros isoforms in all of the hema-topoietic lineages and in HSC and progenitor populations. Onepossible explanation for the poor correlation between RT-PCRproducts and Ikaros proteins is that the expression of Ikaros isregulated after transcripts for the various isoforms have been gen-erated by RNA processing. A poor correlation between mRNA andprotein expression has been reported in a number of other cases.These include genes expressed in hematopoietic cells (47) andgenes that, like Ikaros, are alternately spliced to generate multipleisoforms (48).
Alternatively, simultaneous RT-PCR amplification of tran-scripts for multiple isoforms may not provide an accurate indicatorof the relative levels of each transcript i.e., amplification of sometranscripts may be more efficient than others. A difference in abil-ity to compete for primers may seem unlikely given that primersbind an identical sequence in all transcripts. However, particulartranscripts may be at a selective disadvantage for primer binding orprimer extension due to secondary structures conferred by specificexons. For example, Ikaros isoforms that lack exon 3 (Ik2 and Ik4)are disproportionately overrepresented among RT-PCR productsas compared with their representation at the protein level. In con-trast, isoforms that contain exon 3 (Ik1 and particularly Ikx) areunderrepresented among RT-PCR products as compared with pro-tein expression (Figs. 2, 3, 5, and 6).
Given the predominance of Ikx that we observe in normal hu-man and murine hematopoietic cells, it might seem surprising thatIkx was not identified in early studies. However, the Ikaros genegives rise to a number of splice forms, many of which were notidentified in the initial reports (7, 22). Ikaros expression has mostfrequently been examined by RT-PCR and products consistent insize with Ikx were detected in a number of murine (3, 23) andhuman studies (23, 27, 49, 50). However, these products were notidentified and, in some cases, were thought to be artifacts. Thecomparatively low frequency of Ikx among RT-PCR products didnot suggest that Ikx proteins might be abundantly expressed. Inaddition, there was no indication that the unidentified RT-PCRproduct (consistent in size with Ikx) might correspond to a my-eloid-specific Ikaros protein, because the lineage-specific expres-sion of Ikx proteins is not observed for Ikx transcripts (Fig. 2).
Due to the initial link between Ikaros and the lymphoid lineages,most studies of Ikaros protein expression have been performed onlymphoid cells which express little, if any, Ikx. Ikx was not iden-tified in these studies, although a protein product consistent in sizewith Ikx was detectable in some cases in both murine (22) andhuman (25–27, 49, 50) cells. Thus, the identity of Ikx and itsabundance and almost exclusive expression in the myeloid lineagewere not known before our studies.
Murine HSC and progenitor populations have been shown toexpress Ikaros mRNA (3, 8). As with mature human cells of the
FIGURE 6. Ik1 and Ikx proteins are differentially modulated duringmyeloid and lymphoid differentiation. A, CD34�-enriched CB mononu-clear cells that fall within a forward scatter and side scatter gate (R1)characteristic of lymphoid cells are shown. Lin�CD34� cells were FACSsorted based on R1, R2 and R3 gates. B, Sorted Lin�CD34� cells werefrozen for later immunoblot analysis or cultured under conditions that se-lectively support lymphoid or myeloid differentiation (see Materials andMethods). C, Total CB mononuclear cells, FACS-sorted Lin�CD34� cells,and their progeny generated under lymphoid (L) or myeloid (M) cultureconditions were assessed by immunoblot analysis for total Ikaros proteinexpression using Abs specific to the NTS and the CTS common to allIkaros isoforms.
3096 Ikx IS SELECTIVELY EXPRESSED IN MYELOID DIFFERENTIATION
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

various lineages (Fig. 2), we found that highly purified, FACS-sorted human HSC (CD34�CD38�) and progenitor (CD34�
CD38�) populations generate transcripts for multiple DNA-bind-ing and nonbinding Ikaros isoforms (Fig. 5). Our data are consis-tent with a study by Nakayama et al. (27) that detected transcriptsfor DNA-binding and nonbinding Ikaros isoforms in CD34�-en-riched human CB cells and in culture-generated myeloid and ery-throid cells.
The rarity of CD34�CD38� cells (�0.05% of CB and BMmononuclear cells (38)) precluded immunoblot analysis of proteinexpression in this population. Therefore, for protein analysis, weisolated CD34� CB cells that lacked markers of lineage commit-ment (�0.75% of mononuclear cells). Both Ik1 and Ikx proteinsare detectable in the Lin�CD34� population. However, this pop-ulation is heterogeneous, including primitive HSCs, the CB mul-tilymphoid progenitors (40), and potentially a common myeloidprogenitor. It is possible that Ik1 and Ikx are expressed in differentsubsets of the Lin�CD34� population, rather than coexpressed inthe entire population. Thus, our studies do not provide an analysisof differential Ikaros protein expression in subpopulations of HSCsand early progenitors. However, they do provide the first report ofIkaros protein expression in early hematopoietic cells and demon-strate that Ik1 and Ikx proteins are expressed very early in normalhuman hematopoietic differentiation.
Among the human hematopoietic lineages, Ik1 proteins predom-inate in BM nucleated erythroid lineage cells and in mature NKand B cells isolated from CB. Surprisingly, CB T cells expressonly low levels of multiple DNA-binding and nonbinding Ikarosproteins. However, it is possible that the pattern of Ikaros expres-sion we observed is unique to naive human T cells, because CB Tcells have not yet encountered Ag. In the murine hematopoieticlineages, as in human, Ikx proteins predominate in BM and areselectively expressed in myeloid lineage cells. In contrast to hu-man CB T cells, murine T cells, isolated from spleen and thymus,express Ik1 similarly to B cells. Surprisingly, total Ikaros proteinsappear to be more abundant in myeloid than in lymphoid lineagecells, particularly in the mouse.
Using a mouse erythroid leukemia cell line, O’Neill et al. (5)identified two Ikaros proteins (�65 and 55 kDa in size) that co-purified with the PYR complex that functions in globin switching.Based on size, these proteins were identified as Ik1 and Ik2, re-spectively. Although the 65-kDa protein does correspond to formsof murine Ik1 identified in our studies, the 55-kDa protein is con-sistent in size with murine Ikx proteins that we identified usingAbs specific to Ikaros exon 3 (Ik2 does not contain exon 3). We didnot examine Ikaros expression in murine erythroid lineage cells.Our human studies show abundant Ik1 expression in nucleatederythroid lineage cells; however, Ikx is only faintly detectable.Although there may be variation in Ikx expression between spe-cies, it is also possible that Ikx is selectively incorporated into thePYR complex.
In the first study to describe alternate splicing of murine Ikarostranscripts, Hahm et al. (22) describe a murine Ikaros sequencethat is homologous to the 60-base insert identified in human stud-ies. Using murine cell lines, their study identified transcripts forIkaros splice forms that represent Ik5� and Ik6� (designatedIkaros isoform IV and isoform II, respectively) (22). Our studiesshow that splice variants with the insert (primarily Ik1� and Ikx�)are expressed at the protein level in normal murine and humanhematopoietic cells of multiple lineages. The pattern of expressionfor Ikaros proteins with the insert mirrors that seen for parent iso-forms that do not include the insert.
The selective expression of Ikx proteins in early progenitors andin the myeloid lineage suggests that particular Ikaros isoforms may
have specific functions in hematopoiesis. Due to the relativelylarge numbers of cells required for immunoblot analyses, therehave been few reports of Ikaros isoform expression, at the proteinlevel, in primary murine or human hematopoietic cells of any lin-eage. The poor correlation between Ikaros RT-PCR products andIkaros protein expression underscores the importance of examin-ing Ikaros expression at the protein level. Given the tumor sup-pressor activity ascribed to Ikaros (30, 49–53), aberrant Ikarosexpression in leukemia and other cell lines seems likely. There-fore, an understanding of the role of Ikaros in normal hematopoi-esis may depend on knowledge of the specific Ikaros proteins ex-pressed in primary hematopoietic cells.
The studies described in this report suggest that the expression ofIk1 and Ikx could play a role in myeloid vs lymphoid and erythroidlineage commitment. Current studies in our laboratory are aimed atexamining the effects of overexpression of Ik1 and Ikx on lineagecommitment and hematopoietic differentiation in human HSCs.
AcknowledgmentsWe thank the staff of Kaiser Permanente Hospital Sunset (Los Angeles,CA) for collecting CB samples.
References1. Lo, K., N. R. Landau, and S. T. Smale. 1991. LyF-1, a transcriptional regulator
that interacts with a novel class of promoters for lymphocyte-specific genes. Mol.Cell. Biol. 11:5229.
2. Georgopoulos, K., D. D. Moore, and B. Derfler. 1992. Ikaros, an early lymphoid-specific transcription factor and a putative mediator for T cell commitment. Sci-ence 258:808.
3. Klug, C. A., S. J. Morrison, M. Masek, K. Hahm, S. T. Smale, andI. L. Weissman. 1998. Hematopoietic stem cells and lymphoid progenitors ex-press different Ikaros isoforms, and Ikaros is localized to heterochromatin inimmature lymphocytes. Proc. Natl. Acad. Sci. USA 95:657.
4. Nichogiannopoulou, A., M. Trevisan, S. Neben, C. Friedrich, andK. Georgopoulos. 1999. Defects in hematopoietic stem cell activity in Ikarosmutant mice. J. Exp. Med. 190:1201.
5. O’Neill, D. W., S. S. Schoetz, R. A. Lopez, M. Castle, L. Rabinowitz, E. Shor,D. Krawchuk, M. G. Goll, M. Renz, H. P. Seelig, et al. 2000. An Ikaros-con-taining chromatin-remodeling complex in adult-type erythroid cells. Mol. Cell.Biol. 20:7572.
6. Lopez, R. A., S. Schoetz, K. DeAngelis, D. O’Neill, and A. Bank. 2002. Multiplehematopoietic defects and delayed globin switching in Ikaros null mice. Proc.Natl. Acad. Sci. USA 99:602.
7. Molnar, A., and K. Georgopoulos. 1994. The Ikaros gene encodes a family offunctionally diverse zinc finger DNA-binding proteins. Mol. Cell. Biol. 14:8292.
8. Morgan, B., L. Sun, N. Avitahl, K. Andrikopoulos, T. Ikeda, E. Gonzales, P. Wu,S. Neben, and K. Georgopoulos. 1997. Aiolos, a lymphoid restricted transcriptionfactor that interacts with Ikaros to regulate lymphocyte differentiation. EMBO J.16:2004.
9. Kim, J., S. Sif, B. Jones, A. Jackson, J. Koipally, E. Heller, S. Winandy, A. Viel,A. Sawyer, T. Ikeda, et al. 1999. Ikaros DNA-binding proteins direct formationof chromatin remodeling complexes in lymphocytes. Immunity 10:345.
10. Ito, T., S. Nomura, M. Okada, Y. Katsumata, F. Kikkawa, T. Rogi, M. Tsujimoto,and S. Mizutani. 2002. Ap-2 and Ikaros regulate transcription of human placentalleucine aminopeptidase/oxytocinase gene. Biochim. Biophys. Acta 290:1048.
11. Sun, P., and H. H. Loh. 2002. Transcriptional regulation of mouse �-opioid re-ceptor gene: role of Ikaros in the stimulated transcription of mouse �-opioidreceptor gene in activated T cells. J. Biol. Chem. 277:12854.
12. Koipally, J., E. J. Heller, J. R. Seavitt, and K. Georgopoulos. 2002. Unconven-tional potentiation of gene expression by Ikaros. J. Biol. Chem. 277:13007.
13. Ernst, P., K. Hahm, and S. T. Smale. 1993. Both LyF-1 and an Ets protein interactwith a critical promoter element in the murine terminal transferase gene. Mol.Cell. Biol. 13:2982.
14. Brown, K. E., S. S. Guest, S. T. Smale, K. Hahm, M. Merkenschlager, andA. G. Fisher. 1997. Association of transcriptionally silent genes with Ikaros com-plexes at centromeric heterochromatin. Cell 91:845.
15. Koipally, J., J. Kim, B. Jones, A. Jackson, N. Avitahl, S. Winandy, M. Trevisan,A. Nichogiannopoulou, C. Kelley, and K. Georgopoulos. 1999. Ikaros chromatinremodeling complexes in the control of differentiation of the hemo-lymphoidsystem. Cold Spring Harbor Symp. Quant. Biol. 64:79.
16. Brown, K. E., J. Baxter, D. Graf, M. Merkenschlager, and A. G. Fisher. 1999.Dynamic repositioning of genes in the nucleus of lymphocytes preparing for celldivision. Mol. Cell 3:207.
17. Koipally, J., and K. Georgopoulos. 2000. Ikaros interactions with CtBP reveal arepression mechanism that is independent of histone deacetylase activity. J. Biol.Chem. 275:19594.
18. Cobb, B. S., S. Morales-Alcelay, G. Kleiger, K. E. Brown, A. G. Fisher, andS. T. Smale. 2000. Targeting of Ikaros to pericentromeric heterochromatin bydirect DNA binding. Genes Dev. 14:2146.
3097The Journal of Immunology
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

19. Sabbattini, P., M. Lundgren, A. Georgiou, C. Chow, G. Warnes, and N. Dillon.2001. Binding of Ikaros to the �5 promoter silences transcription through a mech-anism that does not require heterochromatin formation. EMBO J. 20:2812.
20. Trinh, L. A., R. Ferrini, B. S. Cobb, A. S. Weinmann, K. Hahm, P. Ernst,I. P. Garraway, M. Merkenschlager, and S. T. Smale. 2001. Down-regulation ofTDT transcription in CD4�CD8� thymocytes by Ikaros proteins in direct com-petition with an Ets activator. Genes Dev. 15:1817.
21. Dorsam, G., and E. J. Goetzl. 2002. Vasoactive intestinal peptide receptor-1(VPAC-1) is a novel gene target of the hemolymphopoietic transcription factorIkaros. J. Biol. Chem. 277:13488.
22. Hahm, K., P. Ernst, K. Lo, G. S. Kim, C. Turck, and S. T. Smale. 1994. Thelymphoid transcription factor LyF-1 is encoded by specific, alternatively splicedmRNAs derived from the Ikaros gene. Mol. Cell. Biol. 14:7111.
23. Molnar, A., P. Wu, D. A. Largespada, A. Vortkamp, S. Scherer, N. G. Copeland,N. A. Jenkins, G. Bruns, and K. Georgopoulos. 1996. The Ikaros gene encodesa family of lymphocyte-restricted zinc finger DNA binding proteins, highly con-served in human and mouse. J. Immunol. 156:585.
24. Sun, L., N. Heerema, L. Crotty, X. Wu, C. Navara, A. Vassilev, M. Sensel,G. H. Reaman, and F. M. Uckun. 1999. Expression of dominant-negative andmutant isoforms of the antileukemic transcription factor Ikaros in infant acutelymphoblastic leukemia. Proc. Natl. Acad. Sci. USA 96:680.
25. Sun, L., M. L. Crotty, M. Sensel, H. Sather, C. Navara, J. Nachman,P. G. Steinherz, P. S. Gaynon, N. Seibel, C. Mao, et al. 1999. Expression ofdominant-negative Ikaros isoforms in T-cell acute lymphoblastic leukemia. Clin.Cancer Res. 5:2112.
26. Sun, L., P. A. Goodman, C. M. Wood, M. L. Crotty, M. Sensel, H. Sather,C. Navara, J. Nachman, P. G. Steinherz, P. S. Gaynon, et al. 1999. Expression ofaberrantly spliced oncogenic Ikaros isoforms in childhood acute lymphoblasticleukemia. J. Clin. Oncol. 17:3753.
27. Nakayama, H., F. Ishimaru, Y. Katayama, K. Nakase, N. Sezaki, K. Takenaka,K. Shinagawa, K. Ikeda, K. Niiya, and M. Harada. 2000. Ikaros expression inhuman hematopoietic lineages. Exp. Hematol. 28:1232.
28. Olivero, S., C. Maroc, E. Beillard, J. Gabert, W. Nietfeld, C. Chabannon, andC. Tonnelle. 2000. Detection of different Ikaros isoforms in human leukaemiasusing real-time quantitative polymerase chain reaction. Br. J. Haematol. 110:826.
29. Payne, K. J., J. H. Nicolas, J. Y. Zhu, L. W. Barsky, and G. M. Crooks. 2001.Cutting edge: predominant expression of a novel Ikaros isoform in normal humanhematopoiesis. J. Immunol. 167:1867.
30. Wang, J. H., A. Nichogiannopoulou, L. Wu, L. Sun, A. H. Sharpe, M. Bigby, andK. Georgopoulos. 1996. Selective defects in the development of the fetal andadult lymphoid system in mice with an Ikaros null mutation. Immunity 5:537.
31. Georgopoulos, K., M. Bigby, J. H. Wang, A. Molnar, P. Wu, S. Winandy, andA. Sharpe. 1994. The Ikaros gene is required for the development of all lymphoidlineages. Cell 79:143.
32. Winandy, S., L. Wu, J. H. Wang, and K. Georgopoulos. 1999. Pre-T cell receptor(TCR) and TCR-controlled checkpoints in T cell differentiation are set by Ikaros.J. Exp. Med. 190:1039.
33. Avitahl, N., S. Winandy, C. Friedrich, B. Jones, Y. Ge, and K. Georgopoulos.1999. Ikaros sets thresholds for T cell activation and regulates chromosome prop-agation. Immunity 10:333.
34. Winandy, S., P. Wu, and K. Georgopoulos. 1995. A dominant mutation in theIkaros gene leads to rapid development of leukemia and lymphoma. Cell 83:289.
35. Boggs, S. S., M. Trevisan, K. Patrene, and K. Georgopoulos. 1998. Lack ofnatural killer cell precursors in fetal liver of Ikaros knockout mutant mice. Nat.Immun. 16:137.
36. Wu, L., A. Nichogiannopoulou, K. Shortman, and K. Georgopoulos. 1997. Cell-autonomous defects in dendritic cell populations of Ikaros mutant mice point toa developmental relationship with the lymphoid lineage. Immunity 7:483.
37. Dumortier, A., P. Kirstetter, P. Kastner, and S. Chan. 2002. Ikaros regulatesneutrophil differentiation. Blood. 5:1336.
38. Hao, Q. L., A. J. Shah, F. T. Thiemann, E. M. Smogorzewska, and G. M. Crooks.1995. A functional comparison of CD34�CD38� cells in cord blood and bonemarrow. Blood 86:3745.
39. Hao, Q. L., E. M. Smogorzewska, L. W. Barsky, and G. M. Crooks. 1998. In vitroidentification of single CD34�CD38� cells with both lymphoid and myeloidpotential. Blood 91:4145.
40. Hao, Q. L., J. Zhu, M. A. Price, K. J. Payne, L. W. Barsky, and G. M. Crooks.2001. Identification of a novel, human multilymphoid progenitor in cord blood.Blood 97:3683.
41. Kansas, G. S., M. J. Muirhead, and M. O. Dailey. 1990. Expression of the CD11/CD18, leukocyte adhesion molecule 1, and CD44 adhesion molecules duringnormal myeloid and erythroid differentiation in humans. Blood 76:2483.
42. Gooi, H. C., S. J. Thorpe, E. F. Hounsell, H. Rumpold, D. Kraft, O. Forster, andT. Feizi. 1983. Marker of peripheral blood granulocytes and monocytes of manrecognized by two monoclonal antibodies VEP8 and VEP9 involves the trisac-charide 3-fucosyl-N-acetyllactosamine. Eur. J. Immunol. 13:306.
43. Davidson, S. E., J. L. McKenzie, M. E. Beard, and D. N. Hart. 1988. The tissuedistribution of the 3 �-fucosyl-N-acetyl lactosamine determinant recognized bythe CD15 monoclonal antibodies CMRF-7 and 27. Pathology 20:24.
44. Wright, S. D., P. S. Tobias, R. J. Ulevitch, and R. A. Ramos. 1989. Lipopoly-saccharide (LPS) binding protein opsonizes LPS-bearing particles for recognitionby a novel receptor on macrophages. J. Exp. Med. 170:1231.
45. Skubitz, K., K. Micklem, and E. van der Schoot. 1995. CD66 and CD67 clusterworkshop report. In Leukocyte Typing V, Vol. 1. B. L. Schlossmann, W. Gilks,J. Harlan, T. Kishimoto, C. Morimoto, J. Ritz, T. A. Springer, T. F. Tedder,R. F. Todd, eds. Oxford Univ. Press, Oxford, p. 889.
46. Eades-Perner, A. M., J. Thompson, H. van der Putten, and W. Zimmermann.1998. Mice transgenic for the human CGM6 gene express its product, the gran-ulocyte marker CD66b, exclusively in granulocytes. Blood 91:663.
47. Lian, Z., L. Wang, S. Yamaga, W. Bonds, Y. Beazer-Barclay, Y. Kluger,M. Gerstein, P. E. Newburger, N. Berliner, and S. M. Weissman. 2001. Genomicand proteomic analysis of the myeloid differentiation program. Blood 98:513.
48. Chen, G., T. G. Gharib, C. C. Huang, J. M. Taylor, D. E. Misek, S. L. Kardia,T. J. Giordano, M. D. Iannettoni, M. B. Orringer, S. M. Hanash, and D. G. Beer.2002. Discordant protein and mRNA expression in lung adenocarcinomas. Mol.Cell. Proteomics 1:304.
49. Nakayama, H., F. Ishimaru, N. Avitahl, N. Sezaki, N. Fujii, K. Nakase,Y. Ninomiya, A. Harashima, J. Minowada, J. Tsuchiyama, et al. 1999. Decreasesin Ikaros activity correlate with blast crisis in patients with chronic myelogenousleukemia. Cancer Res. 59:3931.
50. Nakase, K., F. Ishimaru, N. Avitahl, H. Dansako, K. Matsuo, K. Fujii, N. Sezaki,H. Nakayama, T. Yano, S. Fukuda, et al. 2000. Dominant negative isoform of theIkaros gene in patients with adult B-cell acute lymphoblastic leukemia. CancerRes. 60:4062.
51. Georgopoulos, K., S. Winandy, and N. Avitahl. 1997. The role of the Ikaros genein lymphocyte development and homeostasis. Annu. Rev. Immunol. 15:155.
52. Okano, H., Y. Saito, T. Miyazawa, T. Shinbo, D. Chou, S. Kosugi, Y. Takahashi,S. Odani, O. Niwa, and R. Kominami. 1999. Homozygous deletions and pointmutations of the Ikaros gene in gamma-ray-induced mouse thymic lymphomas.Oncogene 18:6677.
53. Karlsson, A., P. Soderkvist, and S. M. Zhuang. 2002. Point mutations and dele-tions in the znfn1a1/ikaros gene in chemically induced murine lymphomas. Can-cer Res. 62:2650.
3098 Ikx IS SELECTIVELY EXPRESSED IN MYELOID DIFFERENTIATION
by guest on August 26, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





























