Embed Size (px)
Citation preview

II. TINJAUAN PUSTAKA
Potensi Serat Sawit untuk Pakan Ruminansia
Serat sawit yang diperoleh dari industri minyak sawit di Indonesia akan
terus meningkat sejalan dengan meningkatnya luas area penanaman kelapa sawit.
Di Indonesia saat ini penanaman kelapa sawit (Elais gueneensis JACK) sedang
dikembangkan dengan peningkatan luasan yang pesat dari 120.000 hektar tahun
1969 menjadi 7 juta Ha pada tahun 2010 dengan produksi minyak sawit (crude
palm oil) lebih dari 19 juta ton (Ditjen Perkebunan 2010), dengan meningkatnya
luas per-kebunan kelapa sawit tiap tahunnya 12,6 % (Liwang 2003), akan
meningkatkan limbah pengolahan kelapa sawit yang dihasilkan dan berpotensi
mengganggu lingkungan. Salah satu limbah pengolahan kelapa sawit adalah serat
sawit (palm press fibre). Setiap Ha luasan kebun kelapa sawit dihasilkan limbah
berupa serat sawit sebanyak 2.681 kg bahan kering per tahun (Diwyanto et al
2004), dengan luas perkebunan kelapa sawit yang ada di Indonesia yakni 7 juta
Ha (90 % nya berproduksi), jumlah serat sawit yang dihasilkan adalah sebesar
16,888 metrik ton BK/th. Hal ini merupakan potensi yang besar untuk dijadikan
pakan ternak, terutama ternak ruminansia. Terbatasnya penggunaan serat sawit
dalam ransum karena tingginya kandungan sellulosa (38.69 %) dan lignin
(20.99%) yang mengakibatkan rendahnya daya cerna serat kasar. Untuk mengatasi
ini diperlukan suatu teknologi, salah satu diantaranya memberikan perlakuan
secara kimia (NaOH) dan biologis yakni melakukan fermentasi menggunakan
kapang Aspergillus niger.
Menurut hasil penelitian Purwaningrum (2003) bahwa pemanfaatan SS
yang mendapatkan pengolahan dengan Trichoderma harzianum diperoleh SS dan
LSKS (limbah serat kelapa sawit) dengan rasio 1:2, dan digunakan sebagai
pengganti hijauan konvensional dengan taraf 50% dan lebih dari itu akan
menurunkan kecernaan dan keracunan amonia. Jamarun et al (2000)
mendapatkan hasil penelitian bahwa serat sawit yang telah direndam dengan
NaOH 2,5% dengan lama perendaman 24 jam telah mampu memberikan hasil
yang terba ik dalam menurunkan kandungan NDF, ADF, selulosa, lignin dan silika
serat sawit tanpa mempengaruhi nilai protein kasar. Mutu suatu bahan pakan

8
ditentukan oleh interaksi antara konsentrasi unsur gizi, tingkat kecernaan dan
tingkat konsumsi. Kandungan unsur gizi merupakan indikator awal yang
menunjukkan potensi suatu bahan pakan. Kandungan gizi beberapa produk
hasil samping perkebunan kelapa sawit disajikan pada Tabel 1.
Tabe l 1 Komposisi nutrien produk samping tanaman dan pengolahan buah kelapa sawit
Bahan/produk samping
BK (%)
Abu PK SK L BETN Ca P GE (kal/g) …% BK …
Daun tanpa lidi (5) Pelepah (4) Solid (4) Bungkil (2) Serat perasan (5) Tandan kosong (3)
46,18 26,07 24,08 91,83 93,11 92,10
13,40 5,10 14,40 4,14 5,90 7,89
14,12 3,07 14,58 16,33 6,20 3,70
21,52 50,94 35,88 36,68 48,10 47,93
4,37 1,07 14,78 6,49 3,22 4,70
46,59 39,82 16,36 28,19
-- --
0,84 0,96 1,08 0,56
-- --
0,17 0,08 0,25 0,84
-- --
4461 4841 4082 5178 4684
-- ( ) jumlah contoh Sumber : Mathius et al. (2004)
Tingkat kecernaan akan menentukan seberapa besar unsur gizi yang
terkandung dalam bahan pakan secara potensial dapat dimanfaatkan untuk
produksi ternak. Bahan pakan dengan kandungan serat tinggi seperti pelepah,
daun, serat perasan buah dan batang sawit merupakan sumber utama energi
untuk produksi. Bahan tersebut tetap akan menjadi sumber utama energi bagi
ternak, karena berperan sebagai pakan dasar (pokok) sehingga dikonsumsi dalam
jumlah yang relatif lebih besar.
Unsur kimiawi yang terkandung dalam serat atau dinding sel yang secara
efektif menentukan potensi energi dari suatu bahan pakan adalah konsentrasi dan
keterikatan selulosa, hemiselulosa, lignin, kutin dan silika (Tabel 2). Dari unsur
penyusun dinding sel atau serat tersebut pada dasarnya yang berpotensi sebagai
sumber energi bagi ternak adalah selulosa dan hemiselulosa melalui proses
fermentasi di dalam sistem pencernaan ternak.
Kandungan selulosa dan hemiselulosa dalam keseluruhan serat merupakan
yang terbesar (60-83%) atau setara dengan 44-69% dari bahan kering. Lignin,
selain tidak dapat dimanfaatkan oleh ternak, juga merupakan indeks negatif bagi
mutu bahan pakan, karena ikatannya dengan selulosa dan hemiselulosa
mempersulit pemanfaatan selulosa dan hemiselulosa sebagai sumber energi bagi

9
Tabe l 2 Kandungan senyawa kimia penyusun serat pada beberapa bahan pakan asal perkebunan kelapa sawit
Komponen serat Fraksi kelapa sawit Daun Pelepah Serat perasan buah Batang
Selulosa (%) Hemiselulosa (%) Lignin (%) Silika (%) Total
16,6 27,6 27,6
3,8 75,6
31,7 33,9 17,4
0,6 83,6
18,3 44,9 21,3
tt 84,5
34 35,8 12,6
1,4 83,8
Sumber : Bejo (1995)
ternak (Fengel & Wegener 1995).
Oleh karena itu ketersediaan selulosa sebagai sumber energi bervariasi dan
amat ditentukan oleh intensitas ikatannya dengan senyawa lignin. Silika yang
merupakan elemen struktural dan bersama lignin secara komplementer
memperkuat rigiditas serat/ dinding sel (Fengel & Wegener 1995) juga
menghambat pemanfaatan selulosa dan hemiselulosa sebagai sumber energi.
Kandungan lignin dan silika secara bersama yang relatif tinggi (18-40% dari total
dinding sel) merupakan indikator bahwa tingkat kecernaan bahan pakan
merupakan salah satu kendala penting dan membutuhkan teknik untuk
mengatasinya. Data ini memberi indikasi bahwa masalah utama pemanfaatan
hasil samping perkebunan kelapa sawit adalah bagaimana meningkatkan
kecernaan. Peningkatan kecernaan selanjutnya diharapkan dapat memberi
pengaruh positif bagi peningkatan konsumsi.
Peningkatan Kualitas Serat Sawit dengan NaOH
Umumnya kecernaan dari ransum pakan serat sekitar 40-45 %, karena itu
sangat diperlukan upaya perbaikan nutrisi untuk meningkatkan kecernaannya.
Usaha yang dapat dilakukan antara lain dengan perlakuan pretreatment yaitu
untuk mendapatkan hasil yang tinggi di mana penting untuk pengembangan
teknologi biokonversi dalam skala komersial (Mos ier et al., 2005), secara kimia
diantaranya perendaman dengan NaOH, fisik (uap panas, menggiling dan
memotong), dan biologi (suplemen nitrogen dan fermentasi media padat) untuk
memperkaya nilai nutrisi dari sekam dan sekam yang digiling (Vadiveloo et al.
2009). Tujuan dari pretreatment adalah untuk membuka struktur lignoselulosa
agar selulosa menjadi lebih mudah diakses oleh enzim yang memecah polymer

10
polisakarida menjadi monomer gula. Tujuan pretreatment secara skematis
ditunjukkan pada Gambar 1.
Amorphous
Crystalline Region
Gambar 1. Skema pretreatment biomassa lingo-selulosa (Mosier et al.,
2005). Selain cara hidrolisis kimiawi, menurut Hendriks & Zeeman (2009) bahan
bermutu rendah dapat ditingkatkan kegunaannya dengan cara fisika, secara fisika
usaha lain unt uk memperba iki kualitas bahan makanan berserat dapat dilakukan
dengan pemanasan dan steam bertekanan (autoclave). NaOH adalah alkali yang
paling efektif untuk menaikkan kecernaan zat makanan limbah pertanian/industri
karena mampu merenggangkan ikatan ligno-selulosa yang lebih besar sehingga
kecernaan lebih tinggi, hal ini sesuai menurut Moss et al. (1993) bahwa perlakuan
dengan NaOH adalah suatu metode yang efektif untuk meningkatkan kualitas
jerami padi yang rendah, walaupun penambahan NaOH membuat defisiensi
nitrogen lebih buruk pada jerami padi. Ringkasan berbagai teknik pretreatment
yang dikembangkan ditampilkan pada Tabel 3 di bawah ini.
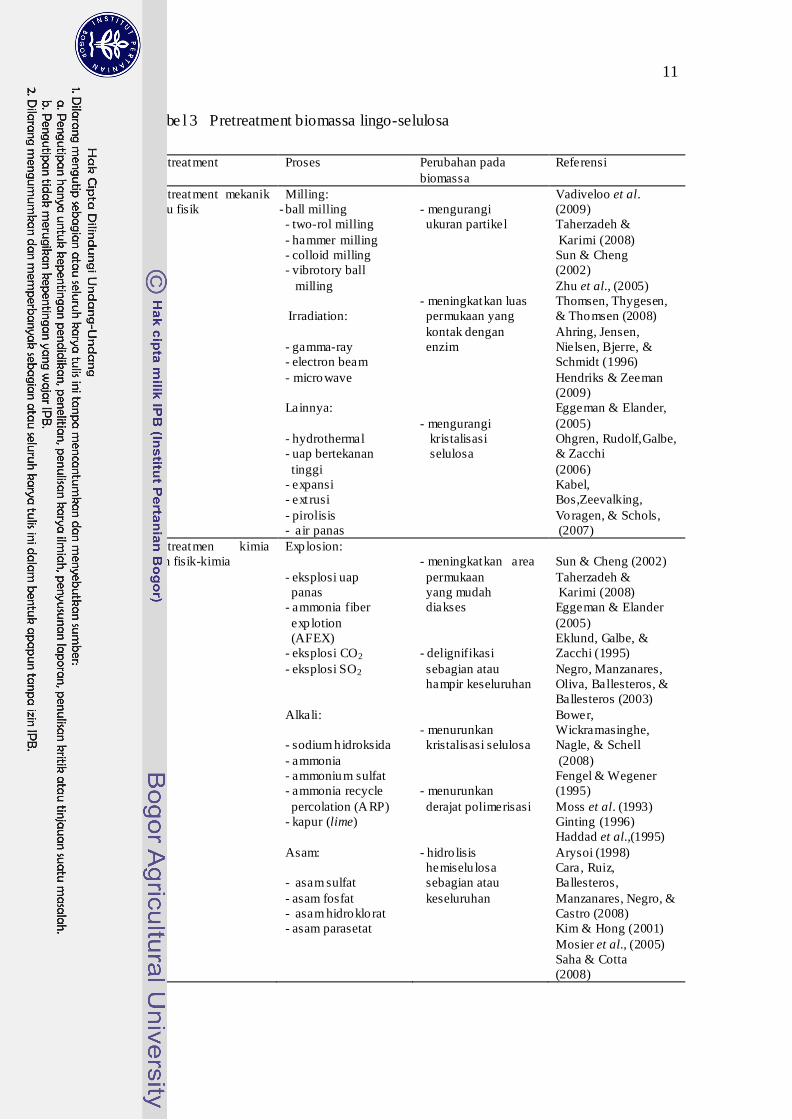
11
Tabe l 3 Pretreatment biomassa lingo-selulosa
Pretreatment Proses Perubahan pada biomassa
Referensi
Pretreatment mekanik atau fisik
Milling: - ball milling - two-rol milling - hammer milling - colloid milling - vibrotory ball milling Irradiation: - gamma-ray - electron beam - microwave Lainnya: - hydrothermal - uap bertekanan tinggi - expansi - ext rusi - pirolisis - air panas
- mengurangi ukuran partikel - meningkatkan luas permukaan yang kontak dengan enzim - mengurangi
kristalisasi selulosa
Vadiveloo et al. (2009) Taherzadeh & Karimi (2008) Sun & Cheng (2002) Zhu et al., (2005) Thomsen, Thygesen, & Thomsen (2008) Ahring, Jensen, Nielsen, Bjerre, & Schmidt (1996) Hendriks & Zeeman (2009) Eggeman & Elander, (2005) Ohgren, Rudolf,Galbe, & Zacchi (2006) Kabel, Bos,Zeevalking, Voragen, & Schols, (2007)
Pretreatmen kimia dan fisik-kimia
Exp losion: - eksplosi uap panas - ammonia fiber exp lotion (AFEX) - eksplosi CO- eksplosi SO
2
2
Alkali: - sodium h idroksida - ammonia - ammonium sulfat - ammonia recycle percolation (ARP) - kapur (lime) Asam: - asam sulfat - asam fosfat - asam hidroklorat - asam parasetat
- meningkatkan area permukaan yang mudah diakses - delignifikasi sebagian atau hampir keseluruhan
- menurunkan kristalisasi selulosa - menurunkan derajat polimerisasi - hidro lisis hemiselu losa sebagian atau keseluruhan
Sun & Cheng (2002) Taherzadeh & Karimi (2008) Eggeman & Elander (2005) Eklund, Galbe, & Zacchi (1995) Negro, Manzanares, Oliva, Ballesteros, & Ballesteros (2003) Bower, Wickramasinghe, Nagle, & Schell (2008) Fengel & Wegener (1995) Moss et al. (1993) Ginting (1996) Haddad et al.,(1995) Arysoi (1998) Cara, Ruiz, Ballesteros, Manzanares, Negro, & Castro (2008) Kim & Hong (2001) Mosier et al., (2005) Saha & Cotta (2008)

12
Pretreatment Proses Perubahan pada biomassa
Referensi
Shimizu, Sudo, Ono, Ishihara, Fujii, & Hishiyama, 1998)
Pretreatment Proses Perubahan pada biomassa
Referensi
Gas: - Clorin d ioksida - Nitrogen dioksida -Sulfur d ioksida Agen Oksidasi: -Hidrogen peroksida - oksidasi basah - Ozone Pelarut untuk ekstraksi lignin : - ekstrasi ethanol-air - ekstrasi benzene-air - ekstraksi etilen Gliko l - ekstraksi butanol- air - agen pemekar (swelling)
Sun & Chen Organosolv pretreatment by crude glycerol from oleochemicals industry for enzymatic hydrolysis of wheat straw (2008) Sun & Chen, Enhanced enzymatic hydrolysis of wheat straw by aqueous glycerol pretreatment (2008) Sun & Cheng (2005) Zhang, et al.,(2008) Kim & Lee (2002) Zhao, Zhang, & Liu (2008) Lloyd & Wayman (2005) Ahring, Jensen, Nielsen, Bjerre, & Schmidt (1996) Silverstein, Chen, Sharma-Shivappa, Boyette, & Osborne (2007)
Biologi - Fungi pelapuk putih - Aktinomicetes
- delignifikasi - penurunan derajat polerisasi selulosa - penurunan derajat kristalisasi selulosa
Taniguchi, Suzuki, Watanabe, Sakai, Hoshino, & Tanaka (2005) Shi, Ch inn, & Sharma-Shivappa, (2008) Keller, Hamilton, & Nguyen (2003) Kirk & Chang, Potential applicat ion of bio-ligninolyt ic System (1981)
Keterangan: modifikasi dari Taherzadeh and Karimi (2008)
Tersedianya jumlah energi dari bahan yang kaya serat kasar dapat ditingkatkan
dengan perlakuan alkali, menurut Haddad et al (1995) mengatakan bahwa
hidrolisis bahan berserat kasar dengan NaOH, NH4OH, urea dan Ca(OH)2

13
menurunkan kadar lignin dan peningkatan daya cerna secara proporsional dengan
turunnya kadar lignin. Perlakuan NaOH pada serat sawit ternyata dapat
meningkatkan bahan kering, bahan organik, abu, energi dan retensi N, namun
tidak terjadi peningkatan kecernaan serat kasar (Arysoi 1998), tetapi pada
penelitian Ginting (1996) perlakuan NaOH dengan konsentrasi 5 % memberikan
koefisien cerna bahan ke ring in-vitro serat sawit yang terbaik dibanding dengan
konsentrasi NaOH 2.5 dan 7.5 %.
Fermentasi
Banyak cara yang dicoba untuk meningkatkan biomassa bagi kepentingan
manusia atau ternak dan dengan cara tersebut semuanya berdasarkan kemampuan
mikroba terutama jamur dan bakteri dalam merubah biomassa menjadi glukosa,
etanol, protein sel tunggal dari makanan ternak. Fermentasi adalah proses
metabolisme dimana enzim yang dihasilkan mikroorganisme menstimulasi reaksi
oksidasi, reaksi hidrolisa dan reaksi kimia lainnya sehingga mengakibatkan
perubahan struktur kimia pada substrat organik dengan menghasilkan produk
tertentu (Dwidjoseputro 2003). Fermentasi merupakan salah satu proses
pengolahan dan pengawetan dengan bantuan mikroba. Fermentasi dapat
meningkatkan nilai gizi bahan makanan menjadi lebih tinggi dari bahan asalnya,
sebab mikroba katabolik akan memecah komponen kompleks menjadi (zat-zat)
yang lebih sederhana.
Proses fermentasi menurut medianya dibagi atas dua golongan yaitu
fermentasi medium padat dan fermentasi medium cair. Fermentasi medium padat
adalah fermentasi yang substratnya tidak larut dan tidak mengandung air bebas
tetapi cukup mengandung air untuk keperluan mikroba. Sebaliknya fermentasi
medium cair adalah fermentasi yang substratnya larut atau tersuspensi di dalam
fase cair (Rahman 1990).
Fermentasi medium padat secara alami umumnya berlangsung pada
medium dengan kadar air berkisar antara 60-80%, karena pada keadaan ini
medium mengandung air yang cukup untuk pertumbuhan mikroba (Bentley &
Bennett 2008, Krishna 2005). Pada hakekatnya kadar air substrat pada fermentasi

14
medium padat tergantung pada sifat alamiah substrat, jenis organisme dan tipe
produk akhir dikehendaki.
Fermentasi medium padat mempunyai beberapa keuntungan antara lain
memiliki kesederhanaan dalam persiapan mediumnya, persiapan inokulum lebih
sederhana, kontrol terhadap kontaminasi lebih mudah, kondisi mediumnya
mendekati keadaan tempat tumbuh kapang yang biasa dijumpai di alam dan
fermentasi memiliki kekurangan (Krishna 2005). Ada ms dan Moss (2008)
menyatakan bahwa kandungan asam amino, lemak, karbohidrat, vitamin, dan
mineral bahan akan mengalami perubahan akibat aktifitas dan
perkembangbiakkan mikroorganisme selama fermentasi. Selanjutnya Fardiaz
(1992) mengatakan bahwa pada proses fermentasi akan terjadi perubahan pH,
kelembaban, aroma, serta perubahan nilai gizi yang mencakup terjadinya
peningkatan protein, vitamin, dan beberapa zat gizi lainnya walaupun mungkin
terjadinya penurunan vitamin B1 dan mineral fosfor.
Terjadinya peningkatan kadar air selama fermentasi disebabkan aktifitas
enzim yang dihasilkan oleh mikroorganisme. Menurut Fardiaz (1992)
mikroorganisme menggunakan karbohidrat sebagai sumber energi yang setelah
terlebih dahulu dipecah menjadi glukosa. Pemecahan glukosa selanjutnya
dilakukan melalui jalur glikolisis sampai akhirnya dihasilkan energi. Pada proses
tersebut juga dihasilkan molekul air dan CO2
Perubahan kadar serat kasar setelah fermentasi terjadi pada dedak padi dan
bungkil inti sawit yang meningkat selama fermentasi berlangsung. Meningkatnya
kadar serat tersebut disebabkan oleh pertumbuhan miselia kapang yang
mengandung serat serta terjadinya kehilangan dari sejumlah padatan lainnya (Nur
2006).
. Sebagian air akan keluar dari
produk sehingga berat kering produk cenderung berkurang setelah fermentasi.
Faktor-faktor yang mempengaruhi fermentasi
Untuk meningkatkan nilai gizi serat sawit, dilakukan fermentasi dengan
menggunakan kapang Aspergillus niger. Dalam proses fermentasi akan terjadi
pemecahan oleh enzim-enzim tertentu terhadap zat-zat yang tidak dapat dicerna
oleh ternak seperti selulosa, hemiselulosa dan polimer-polimernya menjadi gula

15
sederhana dan alkhohol sehingga bahan yang telah difermentasi mempunyai daya
cerna yang lebih tinggi dari bahan asalnya (Bentley & Bennett 2008). Menurut
Fardiaz (1992) untuk mendapatkan pertumbuhan kapang yang baik, perlu
diperhatikan faktor-faktor yang mempengaruhinya seperti suhu, pH, ketersediaan
O2 dan H2
Ikram et al, (2005) menyatakan bahwa, enzim yang dapat menghirolisis ikatan
β(1-4) pada selulosa adalah selulase. Hidrolisis enzimatik yang sempurna
memerlukan aksi sinergis dari tiga tipe enzim ini, yaitu :
O. Media yang diinokulasi pHnya diusahakan sesuai dengan kebutuhan
kapang. Aspergillus niger merupakan kapang yang tumbuh cepat, banyak
digunakan secara komersil dalam produksi asam sitrat, asam glukonat, dan
beberapa enzim seperti amilase, amiloglukosidase dan selulase (Bentley &
Bennett 2008, Iyayi 2004). Kapang A. niger berperan dalam meningkatkan
kandungan protein kasar bahan sehingga meningkatkan daya cerna bahan kering
dan bahan organik yang difermentasi. Jenis fungi yang biasa digunakan dalam
produksi selulase adalah Aspergillus niger (Immanuel et al. 2006, Ikram et al. 2005,
Omojasola et al. 2008, Narasimha et al. 2006).
Endo-1,4-β-D-glucanase (endoselulase, carboxymethylcellulase atau CMCase),
yang mengurai polimer selulosa secara random pada ikatan internal α-1,4-
glikosida untuk menghasilkan oligodekstrin dengan panjang rantai yang
bervariasi.
Exo-1,4-β-D-glucanase (cellobiohydrolase), yang mengurai selulosa dari ujung
pereduksi dan non pereduksi untuk menghasilkan selobiosa dan/atau glukosa.
β–glucosidase (cellobiase), yang mengurai selobiosa untuk menghasilkan glukosa.
Mekanisme hidrolisis selulosa oleh enzim selulase dapat dilihat dalam Gambar 2.
Kompleks selulase digunakan secara komersial dalam pengolahan kopi.
Selulase digunakan secara luas dalam industri tekstil, deterjen, pulp dan kertas
bahkan kadang-kadang digunakan dalam industri farmasi. Dalam krisis energi
sekarang ini, selulase dapat digunakan dalam fermentasi biomassa menjadi
biofuel, walaupun proses ini sifatnya masih eksperimental. Di bidang kesehatan
selulase digunakan sebagai treatment untuk phytobezoars salah satu bentuk
selulosa bezoar di dalam perut manusia (en.wikipedia.org/wiki/cellulase). Seperti
yang dijelaskan di atas, selulosa dapat dihidrolisis menjadi glukosa dengan
menggunakan asam atau enzim. Hidrolisis menggunakan asam biasanya dilakukan

16
pada temperatur tinggi. Proses ini relatif mahal karena kebutuhan energi yang
cukup tinggi. Baru pada tahun 1980-an, mulai dikembangkan hidrolisis selulosa
dengan menggunakan enzim selulase (Gado et al. 2007). Selulosa diproduksi
oleh fungi, bakteri, tumbuhan, dan ruminansia. Produksi komersial selulase pada
umumnya menggunakan fungi atau bakteri yang telah diisolasi. Meskipun banyak
mikroorganisme yang dapat mendegradasi selulosa, hanya beberapa
mikroorganisme yang memproduksi selulase dalam jumlah yang signifikan yang
mampu menghidrolisa kristal selulosa secara invitro. Fungi adalah
mikroorganisme utama yang dapat memproduksi selulase, meskipun beberapa
bakteri dan actinomycetes telah dilaporkan juga menghasilkan aktivitas selulase.
Fungi berfilamen seperti Tricoderma dan Aspergillus adalah penghasil selulase dan
crude enzyme secara komersial fungi-fungi tersebut sangat efisien dalam
memproduksi selulase (Ikram et al. 2005).
Gambar 2 Mekanisme hidrolisis selulosa (en.wikipedia.org/wiki/cellulase).
Hidrolisis selulosa secara biologik dapat dilakukan baik menggunakan
enzim selulase (Vrije et al., 2002) maupun mikroorganisme selobios glukosa
penghasil selulase (Aderemi et al ., 2008). Hidrolisis selulosa dipengaruhi oleh
jenis sumber subsrat (seperti serbuk gergaji, jerami padi, sabut sawit) dan ukuran
partikel. selulotik, jumlah β-glukosidasenya lebih rendah dari yang dibutuhkan

17
untuk hidrolisis selulosa menjadi glukosa secara efisien, sehingga produk utama
hidrolisisnya bukan glukosa melainkan selobiosa (Juhasz et al., 2005; Martins et
al., 2008; Ahamed dan Vermette, 2008), yang merupakan inhibitor kuat terhadap
endo dan eksoglukanase. Mikroorganisme yang mempunyai kemampuan
memprod uksi β-glukosidase yang kuat yaitu Aspergillus niger (Juhasz et al.
2005).
Perubahan zat-zat makanan selama fermentasi
Makanan yang mengalami fermentasi biasanya mempunyai nilai gizi yang
lebih baik dari asalnya. Hal ini tidak hanya disebabkan oleh mikroorganisme yang
memecah komponen-komponen kompleks menjadi zat-zat yang lebih sederhana
sehingga mudah dicerna, tetapi mikroorganisme juga dapat mensintesa beberapa
vitamin seperti riboflavin, vitamin B12, provitamin A dan faktor pertumbuhan
lain-nya, juga dapat terjadi pemecahan gula oleh enzim tertentu misalnya
hemiselulosa, sellulosa dan polimer-polimernya menjadi gula sederhana atau
turunannya (Winarno 2008).
Bentley dan Bennett (2008) menjelaskan bahwa kapang yang mempunyai
pertumbuhan dan perkembangbiakan yang baik yang akan dapat merubah lebih
banyak komponen penyusun media menjadi suatu massa sel, sehingga akan
terbentuk protein yang berasal dari tubuh kapang itu sendiri dan dapat
meningkatkan protein kasar dari bahan. Fardiaz (1992) menambahkan selama
proses fermentasi mikroba akan mengeluarkan enzim dimana enzim tersebut
adalah protein dan mikroba itu sendiri juga merupakan sumber protein sel
tunggal.
Menurut hasil penelitian Jamarun et al. (2000) bahwa serat sawit fermentasi
dapat digunakan sampai level 45% dari total ransum (menggantikan 75%
kebutuhan hijauan). Pemberian 60% serat sawit yang difermentasi dengan
Aspergillus niger Cz 51 VI/I dalam ransum atau pengganti 100% hijauan dengan
serat sawit, menyebabkan penurunan berat badan ternak domba. Terbatasnya
penggunaan serat sawit dalam ransum, karena tingginya kandungan lignoselulosa
(selulosa 38.60% dan lignin 20.99%) yang mengakibatkan rendahnya daya cerna
serat kasar. Untuk mengatasi hal ini diperlukan suatu teknologi, salah satunya

18
dengan memberikan perlakuan secara biologis yakni melakukan fermentasi
menggunakan kapang Aspergillus niger Cz 51 VI/I pensintesa Cr-organik.
Fermentasi dengan Aspergillus niger
Aspergillus niger merupakan kapang yang dapat tumbuh cepat, banyak
digunakan secara komersial dalam produksi asam sitrat, asam glutamat serta
beberapa enzim, seperti amilase, pektinase, amiloglukosidase dan selulase.
Kapang ini dapat menghasilkan berberapa vitamin yang larut dalam air seperti B6
, B12 , dan niasin. Aspergillus niger dapat tumbuh pada kisaran pH antara 2,8 –
8,8 dengan pH optimum berkisar antara 3,0 – 6,0 dalam pertumbuhannya
Aspergillus niger membutuhkan mineral Mg, Fe, K, Zn, Mn, tiamin dan urea.
Enzim selulsase yang dihasilkan Aspergillus niger menunjukkan aktivitas
optimum pada kisaran pH 4,5 – 5,5. Aspergillus niger bersifat aerobik, sehingga
dalam pertumbuhannya membutuhkan oksigen dalam jumlah yang cukup. Suhu
pertumbuhan optimum Aspergillus niger adalah 35 – 37oC (Iyayi 2004),
sedangkan suhu untuk produksi enzim selulase adalah 25 – 28oC (Bentley &
Bennett 2008).
Aspergillus niger adalah kapang penghasil komplek enzim selulase yang
memiliki aktivitas tinggi dan berpotensi untuk dimanfaatkan dalam menkonversi
bahan lignoselulosik menjadi bioenergi. Dari hasil penelitian kompleks ensim
selulase yang dihasilkan dari Aspergillus niger terdiri da ri CMC-ase (1,4-ß-D-
glucan glucanohydro- lase), Avicelase (1,4-ß-cellobiosidase) dan ß-glukosidase (ß-
D-glucosidase gluco-hydrolase) dengan masing-masing memiliki kemampuan
yang berbeda dalam mendegradasi selulase. Dalam mendegradasi selulosa
kompleks enzim tersebut bertipe endo berbeda dengan enzim selulase yang
dihasilkan dari bakteri umumnya bertipe exo. Mod el aksi da ri enzim tersebut
akan berpengaruh pada kemampuan dalam mendegradasi bahan lignoselulos ik
menjadi komponen gula yang lebih sede rhana (Bentley & Bennett 2008 ).
Aspergillus niger merupakan salah satu spesies yang paling umum dan
mudah diidentifikasi dari genus Aspergillus, famili Moniliaceae, ordo Monoliales
dan kelas Fungi imperfecti. Aspergillus niger dapat tumbuh dengan cepat,
diantaranya digunakan secara komersial dalam produksi asam sitrat, asam
glukonat dan pembuatan berapa enzim seperti amilase, pektinase,

19
amiloglukosidase dan sellulase. Aspergillus niger dapat tumbuh pada suhu
35ºC - 37ºC (optimum), 6 - 8ºC (minimum), 45ºC-47ºC (maksimum) dan
memerlukan oksigen yang cukup (aerobik). Aspergillus niger memiliki bulu dasar
berwarna putih atau kuning dengan lapisan konidiospora tebal berwarna coklat
gelap sampai hitam. Kepala konidia berwarna hitam, bulat, cenderung memisah
menjadi bagian-bagian yang lebih longgar dengan bertambahnya umur.
Konidiospora memiliki dinding yang halus, hialin tetapi juga berwarna coklat.
Aspergillus niger memerlukan mineral (NH4)2SO4, KH2PO4, MgSO4,
urea, CaCl2.7H2O, FeSO4, MnSO4.H2O untuk menghasilkan enzim sellulase,
sedangkan untuk enzim amilase khususnya amiglukosa diperlukan (NH4)2SO4,
KH2PO4 .7H2O, Zn SO4, 7H2O. Bahan organik dengan kandungan nitrogen
tinggi dapat dikomposisi lebih cepat dari pada bahan organik yang rendah
kandungan nitrogennya pada tahap awal dekomposisi. Tahap selanjutnya bahan
organik yang rendah kandungan nitrogennya dapat dikomposisi lebih cepat
daripada bahan organik dengan kandungan nitrogen tinggi. Penurunan bahan
organik sebagai sumber karbon dan nitrogen disebabkan oleh Aspergillus niger
sebagai sumber energinya untuk bahan penunjang pertumbuhan atau growth
factor. Aspergillus niger dalam pertumbuhannya berhubungan langsung dengan
zat makanan yang terdapat dalam substrat, molekul sederhana yang terdapat
disekeliling hifa dapat langsung diserap sedangkan molekul yang lebih kompleks
harus dipecah dahulu sebelum diserap ke dalam sel, dengan menghasilkan
beberapa enzim ekstra seluler. Bahan organik dari substrat digunakan oleh
Aspergillus niger untuk aktivitas transport molekul, pemeliharaan struktur sel dan
mobilitas sel. Amonia dapat digunakan oleh kapang untuk pembentukan asam
amino. Sedangkan perubahan kandungan SK dipengaruhi oleh intensitas
pertumbuhan miselia kapang , kemampuan memecah SK untuk memenuhi
kebutuhan energi, dan kehilangan BK selama fermentasi. Penurunan SK diduga
karena Aspergillus niger pada inkubasi 4 hari mulai mensintesa enzim pengurai,
yaitu selulose yang akan merombak selulosa dalam produk. Aspergillus niger
merupakan kapang yang dapat tumbuh cepat an menghasilkan beberapa enzim
seperti amylase, pektinase, amiloglukosidase dan selulase (Bentley & Bennett
2008).

20
Fermentasi yang dilakukan dengan menggunakan Aspergillus niger dapat
meningkatkan kecernaan dan kandungan protein kasar serat sawit. Hasil
penelitian yang telah dilakukan menggunakan berbagai level inokulum
Aspergillus niger dan lama fermentasi serat sawit dengan NaOH, terhadap
kecernaan bahan kering (KCBK) dan kecernaan bahan organik (KCBO)
meningkat, dengan meningkatnya level inokulum dan lama fermentasi (Jamarun
et al 2000). Pemanfaatan hasil fermentasi bungkil inti sawit dengan Aspergillus
niger dalam ransum ayam broiler terhadap warna daging, memperlihatkan hasil
yang signifikan dibandingkan dengan tanpa perlakuan. Pemakaian 7.5% bungkil
inti sawit fermentasi memberikan warna daging merah ceri dibandingkan dengan
tanpa bungkil inti sawit fermentasi (Nur 2001). Hasil dari fermentasi ini beraroma
wangi yang disenangi ayam dan dapat disimpan dalam jangka waktu 1 (satu)
bulan dan lemak pada abdomen tidak begitu banyak dan rasa dagingnya manis.
Setelah lumpur sawit difermentasi selama 4 hari, kandungan PK nya naik menjadi
35,43 % dari 13,25% dan serat kasarnya menjadi 13,8% dari 16,3%. Kenaikan PK
LSF ini dikarenakan setelah fermentasi 4 hari terjadi kehilangan bahan kering
yang tinggi (28,77%), kapang ini juga mempunyai intensitas pertumbuhan yang
tinggi, kemudian diduga juga kapang ini telah mensintesis enzim ureasi untuk
mencegah urea menjadi amonia dan CO2 pada fermentasi 4 hari.
Peranan kromium dalam sistem transport dan metabolisme nutrien
Kromium (Cr) diketahui merupakan mineral esensial sejak tahun 1959.
Schwart dan Mertz adalah orang pertama yang menemukan bahwa yeast
mengandung suatu substansi yang mampu meningkatkan uptake glukosa dan
meningkatkan potensi aktifitas insulin. Substansi ini kemudian diketahui sebagai
faktor toleransi glukosa (Glucose Tolerance Factor, GTF). Struktur GTF
tersusun dari kompleks antara Cr3+
dengan 2 molekul asam nikotinat dan 3 asam
amino yang terkandung dalam glutation yaitu glutamat, glisin dan sistein (Linder
1992, Underwood & Suttle 2001) seperti disajikan pada Gambar 3. Di dalam
struktur GTF kromium adalah komponen aktifnya sehingga tanpa adanya Cr pada
pusat atau intinya, GTF tidak dapat bekerja mempengaruhi insulin (Burton 1995).

21
Gambar 3 Struktur faktor toleransi glukosa (Linder 1992)
Linder (1992) menyatakan kerja GTF pada mempengaruhi insulin (Burton 1995).
Linder (1992) menyatakan kerja GTF pada sistem transport glukosa dan asam
amino adalah meningkatkan pengikatan insulin dengan reseptor spesifik pada
organ target. Saat insulin mengikat reseptor spesifik-nya, uptake seluler glukosa
dan asam amino dipermudah dalam hal fungsi GTF adalah meningkatkan
efektifitas potensi insulin.
Fungsi GTF sebenarnya lebih berpusat pada sel target, kerja GTF dalam
transfer gula pada sel ragi tidak bergantung pada kehadiran insulin. Hasil-hasil
penelitian Cr menunjukkan bahwa selain esensial dalam metabolisme karbohidrat,
Cr juga dibutuhkan dalam metabolisme lemak dan protein, dalam hal ini difisiensi
Cr dapat menyebabkan hiperkolesterolemia dan arterosklerosis serta rendahnya
inkorporasi asam amino pada protein hati. Asam amino yang dipengaruhi oleh Cr
adalah metionin, glisin dan serin (Anderson 1994).
Fungsi utama Cr ialah untuk meningkatkan aktivitas insulin di dalam
metabolisme glukosa dan untuk mempertahankan transport glukosa dari darah ke
dalam sel-sel. Kromium membentuk suatu komplek dengan insulin dan reseptor
insulin memfasilitasi respon jaringan yang sensitive terhadap insulin (NRC 1997).
Kegunaan Cr sebagai suatu faktor nutrien ditetapkan untuk pertama kalinya ketika
diketahui bahwa brewer`s yeast secara positif dapat mempengaruhi metabolism
karbohidrat pada organisme tingkat tinggi dan meningkatkan aktifitas hormon
insulin (NRC 1997; Demirci & Pornetto 2000; Vincent 2000 ). Kromium trivalent
mempunyai kecenderungan yang sangat kuat untuk membentuk komplek

22
octahedral dengan ligand biologis pada membrane sel (Zetic et al. 2001).
Kromium merupakan suatu elemen yang dapat menstabilkan struktur tersier dari
protein (Demirci & Pornetto 2000). Hampir semua sumber Cr di alam terdapat
dalam bentuk trivalen (Cr III), tetapi produk dari pabrik (K2Cr2O7, K2Cr2O4 dan
Na2Cr2O4
Pada sel kelenjar ambing hewan ruminansia uptake glukosa tidak
ditentukan oleh insulin, namun insulin sangat dibutuhkan untuk pengambilan
asam amino khususnya asam aspartat, valin, isoleusin, leusin dan tirosin
(McGuire et al. 1995; Manalu 1999). Hasil penelitian Lyons (1995) mendapatkan
) terdapat dalam bentuk heksavalen (Cr VI). Bentuk Cr juga dapat
mempengaruhi ketersediaannya secara secara biologis (bioavailabilitas)
contohnya oksalat, meningkatkan absorbsi Cr pada tikus, sedangkan EDTA
(Ethylene Diamine Tetraacetic Acid) sitrat tidak meningkatkan absorbsi Cr.
Bentuk bentuk organik sintetik lainnya seperti kromium nikotinat dan kromium
pikolinat juga telah digunakan sebagai sumber kromium yang mudah tersedia.
Inkopo rasi kromium ke dalam jaringan sangat tergantung pada bentuk
kromiumnya dan inkoporasi kromium paling tinggi terjadi pada kromium
dinicotinic diglycerine glutamic acid, kromium pikolinat, kromium asetat,
kromium potassium sulfat dan komplek glycine kromium. Komplek kromium
yang terjadi secara alam, juga diketahui mempunyai bioavailabilitas yang relative
tinggi. Percobaan pada tikus menunjukkan bahwa 10-25% dari Cr diabsorbsi di
dalam ragi bir (NRC 1997). Kromium selain penting di dalam metabolisme
karbohidrat, juga dibutuhkan dalam metabolism lemak dan protein (Davis &
Vincent 1997), asam nukleat dan mencegah stress. Kromium juga berperan dalam
sistem kekebalan tubuh dan konversi tiroksin (T4) menjadi triiodotironin (T3),
yaitu hormon yang berperan dalam meningkatkan laju metabolisme karbohidrat,
lemak dan protein di dalam hati, ginjal, jantung dan otot serta meningkatkan
sintesis protein (Burton 1995; Stipanuk 2000). Suplementasi Cr ke dalam pakan
lebih menguntungkan apabila diberikan dalam bentuk Cr organik. Kromium
dalam bentuk trivalen yang tidak beracun sangat sulit diserap. Pada beberapa
kasus, Cr organik yang dikonsumsi manusia lewat makanan 98% tidak diserap
dan dikeluarkan lewat feses, sebaliknya ketersediaan Cr organik cukup tinggi
yaitu antara 25 sampai 30% (NRC 1997).

23
bahwa selain asam amino di atas, asam amino lain yang uptake selulernya ke
dalam sel kelenjar ambing meningkat oleh perlakuan infusi insulin adalah
metionin, lisin, asam glutamat, treonin, asparagin dan serin, fungsi GTF adalah
meningkatkan efektifitas potensi insulin (Gambar 4). Burton (1995)
menambahkan bahwa Cr berperan dalam sitem kekebalan tubuh dan konversi
tiroksin (T4) menjadi triiodotironin (T3) yaitu hormon yang berperan
meningkatkan laju metabolisme karbohidrat, lemak dan protein da lam hati, ginjal,
jantung dan otot serta meningkatkan sistesis protein.
Permukaan Membran sel Insulin
Insulin
GTF
Reseptor insulin Insulin pada permukaan membransel Gambar 4 Mekanisme kerja GTF dalam meningkatkan potensi aktifitas insulin
(Lyons 1995)
Spears (1999) yang menghimpun beberapa hasil penelitian tentang
peranan Cr dalam sistem kekebalan tubuh mengatakan bahwa Cr berpengaruh
baik pada pembentukan sistem kekebalan humoral (HI) maupun kekebalan yang
diperantarai oleh sel (CMI). Dalam HI suplementasi Cr meningkatkan produksi
antibodi atau imunoglobin (Igs), sedangkan dalam CMI suplementasi Cr
menyebabkan peningkatan respons blastogenik (lymphocit blasgonesis) terhadap
imunostimulan. Sohn et al. (2000) menyatakan bahwa peningkatan produksi
antibodi adalah sebagai akibat penurunan konsentrasi kortisol. Hormon ini
bekerja meningkatkan glukoneogenesis pada saat ternak dalam kondisi stres.
Proses glukoneogenesis akan menekan sintesis protein dalam hati sehingga

24
sintesis antibod i juga ditekan, dengan kata lain hormon kortisol bekerja
berlawanan dengan terbentuknya sistem kekebalan dalam tubuh ternak.
Kebutuhan kromium dan bentuk suplemen dalam pakan
Kebutuhan Cr pada ternak belum diketahui dengan pasti. Pada kondisi
stres kebutuhan Cr akan meningkat. Hal ini disebabkan oleh cadangan Cr dalam
tubuh berkurang akibat peningkatan mobilisasi cadangan glukosa jaringan perifer
untuk mencukupi kebutuhan glukosa pada otak, yang selanjutnya sebagian Cr itu
akan hilang melalui urine (Burton 1995). Selama kebuntingan dan laktasi
kebutuhan Cr juga meningkat, hal ini karena pada saat kebuntingan ternak
mengalami stres akibat perubahan fisiologis, fisik dan metabolik (Yang et al.
1996).
Cr-GTF dapat diserap 15-20 %, dayagunanya lebih tinggi. Berdasarkan
pada banyaknya kromium yang hilang : 0,5-1 µg/hari dan rata-rata penyerapannya
1 %, dengan demikian kebutuhan minimum 50 µg/hari, dengan rekomendasi
konsumsi 50-200 µg. Setelah penyerapan, kromium diangkut pada protein
pengangkut Fe (iron carrier protein) dari plasma darah : transferin. Tidak
diketahui apakah GTF yang diserap melalui intestin akan masuk ke dalam darah
tanpa perubahan bentuk atau juga terikat dengan transferin. Dari intestin, hampir
semua kromium masuk ke dalam hati dimana akan terinkoperasi ke dalam GTF.
Sejumlah GTF tertentu disekresi ke dalam plasma dimana akan tersedia dalam
menolong aktivitas insulin. Kalau kadar glukose darah meningkat, dan/atau
insulin disekresi, meningkatkan aliran GTF dan/atau kromium ke da lam plasma,
GTF akan meningkatkan pengaruh insulin yang disekresi tersebut dan kemudian
keluar melalui urin. Aktivitas GTF (dan/atau Cr) masih banyak yang belum
diketahui; mungkin terlibat pengaruhnya pada struktur insulin dan/atau pengikatan
resptor (Linder 1992).
Meskipun konsentrasi Cr dalam tubuh relatif kecil, toleransinya dalam pakan
cukup besar yaitu 3000 ppm dalam bentuk Cr2O3 dan 1000 ppm CrCl3 (NRC
1997). Efektivitas suplementasi Cr selain tergantung pada jenis ternak juga
tergantung pada kondisi fisiologis dan bentuk Cr yang digunakan. Kompleks
organik Cr terdapat dalam bentuk Cr-chelate, Cr proteinat ragi (high Cr-yeast) dan

25
Cr-pikolinat. Kromium pikolinat terbentuk dari Cr3+ yang mengikat 3 molekul
asam pikolinat. Asam pikolinat adalah metabolik sekunder yang dihasilkan pada
metabolisme triptopan sebelum membentuk niasin atau asam nikotinat (Combs
1992; Groff & Gropper 2000). Tahapan metabolisme triptopan menjadi niasin
disajikan pada Gambar 5.
Gambar 5 Metabolisme triptopan menjadi niasin (Combs 1992)
Pengaruh suplementasi kromium terhadap produksi ternak
Page et al (1993), yang meneliti tentang suplementasi Cr pada babi sedang
tumbuh, mendapatkan bahwa suplementasi Cr pikolinat sebanyak 200 ppb
meningkat-kan pertambahan bobot badan 0.87 kg/hari lebih tinggi dibanding
kontrol 0.81 kg/hari. Pertambahan bobot badan yang tinggi hasil penelitian di
atas, menggambarkan terjadinya peningkatan sintesis protein dan lemak pada
jaringan perifer akibat meningkatnya uptake asam amino dan glukosa oleh
efektifitas kerja insulin akibat adanya Cr. Namun demikian suplementasi Cr pada
kondisi laktasi akan berpengaruh menurunkan sensitifitas jaringan perifer

26
terhadap insulin sehingga asam amino dan glukosa dialirkan ke dalam sel kelenjar
ambing untuk p roduks i susu.
Fenomena di atas dibuktikan oleh hasil penelitian Yang et al. (1996) pada
sapi perah laktasi bahwa suplementasi chelate sebesar 5 mg Cr per hari
menghasilkan peningkatan produksi susu sebesar 13.25 % (27.5 vs 24.3 kg/hari).
Lebih lanjut dijelaskan bahwa suplementasi Cr juga meningkatkan konsumsi
bahan kering sebesar 15 % (13.76 vs 11.95 kg/hari), mengubah resistensi insulin
pada sel kelenjar ambing, menurunkan kejadian milk fever dan teat edema.
Penelitian senada yang dilakukan oleh Subiyatno et al. (1996) mendapatkan hasil
suplementasi Cr pada sapi perah laktasi sebesar 7.25 mg/hari mampu
meningkatkan produksi susu sebesar 24 % (22.9 vs 18.5 kg) pada 2 minggu
pertama laktasi. Selain itu juga meningkatkan konsentrasi hormon IGF-1 (39.05
vs 49.42 µg/ml) dan ratio insulin/glukosa (7.27 vs 5.76 U/mol). Pada domba
pengaruh penambahan kromium disajikan pada Tabel 4.
Tabe l 4 Pengaruh penambahan kromium dalam ransum domba
Referensi Kebutuhan konsentrasi per kg BK Berat badan pada awal dan selama penelitian
Peningkatan pertumbuhan rata-rata
Peningkatan efisiensi pakan
Britton et al. 1968 Samsell dan Spears 1989 Samsell dan Spears 1989 William et al. 1994 Kitchalong et al. 1995 DePew et al. 1996 Sano et al. 1996
Basal diet=tdk diketahui Cr diet=0,037 mg/d and basal (molasses ash or CrCl3Basal diet, low fiber=0,175 mg;
)
Basal diet, low fiber=0,295 mg; Cr diet,low fiber=0,185 mg Cr diet,high fiber=0,305 mg (CrCl3Basal diet, low fiber=0,175 mg;
)
Basal diet, low fiber=0,295 mg; Cr diet,low fiber=0,185 mg Cr diet,high fiber=0,305 mg (CrCl3Basal diet=tdk diketahui
)
Cr diet Basal diet= 1 mg Cr diet Basal diet=tdk diketahui Cr diet Basal diet=tdk diketahui Cr diet
Growing lambs 16 anak domba-45kg 28 hari percobaan 16 anak domba-50kg 28 hari percobaan 24 anak domba-29kg Heat stressed 24 anak domba-38kg 85 hari percobaan 24 anak domba-33kg 42 hari percobaan 6 domba jantan
ND ND ND ND ND Tidak ada pengaruh Yes-no Statistics provided
ND ND ND No effect ND ND ND
Sumber: NRC (1997) Keterangan: Cr = kromium ND= not determined CrCl3 = kromium klorida

27
Tang et al. (2008) meneliti pengaruh Cr terhadap pembentukan antibodi
dengan memberikan Cr-yeast sebanyak 200 µg/kg ransum pada babi yang disuntik
antigen virus demam. Empat belas hari setelah penyuntikan diperoleh hasil
bahwa jumlah titer antibodi lebih tinggi pada babi yang diberi Cr-yeast dibanding
kontrol yaitu 0.09 vs 0.17 U. Hasil senada diperoleh Burton et al. (1994) bahwa
suplementasi Cr meningkatkan titer antibodi sapi yang disuntuk dengan antigen
sel darah manusia (HRBC) dan vaksin virus rhinotracheitis.
Pada kondisi stres kebutuhan Cr akan meningkat. Hal ini disebabkan oleh
mobilisasi cadangan Cr dalam tubuh meningkat akibat peningkatan mobilisasi
cadangan glukosa jaringan perifer untuk mencukupi kebutuhan glukosa pada otak,
yang selanjutnya sebagian Cr itu akan hilang melalui urine . Ternak yang
mengalami stress, kebutuhannya akan kromium meningkat karena pada kondisi
stress terjadi peningkatan metabolism glukosa secara cepat yang ditandai dengan
meningkatnya sekresi hormone kortisol di dalam darah, sedangkan hormone
kor tisol memiliki aks i yang antagonis denga n insulin yaitu mencegah masuknya
glukosa ke dalam sel jaringan tubuh, akibatnya glukosa yang masuk ke dalam sel
menurun, sehingga menyebabkan kadar glukosa darah meningkat yang disebut
dengan hiperglisemia. Ternak yang mengalami stres, kebutuhannya akan Cr
meningkat karena pada kondisi stres terjadi peningkatan metabolisme glukosa
secara cepat yang ditandai dengan meningkatnya sekresi hormon kortisol di dalam
darah sedangkan hormon kortisol memiliki aksi yang antagonis dengan insulin
yaitu mencegah masuknya glukosa ke dalam sel jaringan tubuh, akibatnya glukosa
yang masuk ke dalam sel menurun, sehingga menyebabkan kadar glukosa darah
meningkat yang disebut dengan hiperglisemia. Peningkatan kadar glukosa darah
merangsang mobilisasi Cr dari penyimpanannya di dalam tubuh. Unsur Cr yang
telah dimobilisasi bersifat tidak balik (irreversible) dan keluar melalui urin
sehingga pada kondisi stres peluang terjadinya defisiensi Cr meningkat. Selain
hiperglisemia, stress akan mengganggu pertumbuhan. Oleh ka rena itu perlu untuk
menormalkan kadar glukosa darah agar tidak mengganggu pertumbuhan dan
performa ternak (Burton 1995).
Moonsie dan Mowat (1993) menyatakan bahwa, suplementasi Cr ragi
(0.2, 0.5 dan 1,0 ppm) pada anak sapi yang mengalami stres meningkatkan berat

28
badan dan konsumsi pakan masing-masing sebesar 29 dan 15% dibandingkan
dengan kontrol selama 30 hari pertama di feedlot. Suplementasi 0.4 ppm Cr yeast
pada sapi yang mengalami stres, setelah 28 hari hasilnya menunjukkan tidak
adanya perbedaan terhadap konsumsi bahan kering per minggu selama 4 minggu
dan pertambahan bobot badan sapi juga tidak berbeda, tetapi Cr nyata
menurunkan kadar kortisol serum (75.0 vs 55.6 nmol/L) dan meningkatkan IgM
dalam serum (kandungan Cr ransum basal (12.12 pp m). Tidak ada respon
terhadap vaksinasi PI-3 (para influenza-3) pada sapi yang stres akibat transportasi
yang disuplementasi 0.16 mg Cr chelate (Burton et al. 1995). Suplementasi 0.8
mg Cr pikolinat pada 24 ekor domba pada kondisi cekaman panas (heat stress)
dengan bobot badan 29 kg tidak mempengaruhi efisiensi penggunaan ransum,
kadar glukosa plasma, keseimbangan nitrogen, dan kemampuan untuk mencerna
serat (kandungan Cr ransum basal tidak diketahui) (Williams et al. 1994).
Suplementasi 0.25 mg Cr pikolinat pada 24 ekor domba dengan bobot badan 38
kg selama 85 hari tidak mempengaruhi total kolesterol, albumen, total protein, T3
dan T4, glukosa dan urea tetapi terjadi penurunan NEFA (non esterifed fatty acid)
(kandungan Cr ransum basal < 1 mg) (Kitchalong et al. 1995). Selanjutnya
menurut Uyanik (2001) suplementasi Cr tidak mempengaruhi pertambahan bobot
badan, kadar Cr darah, LDL dan kolesterol tetapi menurunkan kadar glukosa
darah pada domba yang disuplementasi dengan 0.2 dan 0.4 Cr organik. Hasil uji
in vitro ransum yang disuplementasi Cr anorganik maupun Cr organik (1, 2, 3 dan
4 ppm) meningkatkan produksi total VFA tetapi kadar NH3 menurun dan
suplementasi Cr organik lebih efisien dibandingkan dengan bentuk anorganik.
Level terbaik penggunaan Cr organik adalah 1.0 ppm (Jayanegara et al. 2006).
Suplementasi Cr organik asal Rhizopus orizae dalam ransum sebesar 1 dan 3
mg/kg memberikan hasil tertinggi pada kecernaan bahan kering dan bahan
organik (secara invitro) (Astuti 2006).
Suplementasi Cr ke dalam pakan akan lebih menguntungkan apabila
diberikan dalam bentuk kompleks organik. Hal ini karena dalam bentuk
anorganik, Cr dapat meracuni terutama yang berbentuk heksavalen (Cr6+),
walaupun tingkat absorbsinya di usus tinggi, sedangkan bentuk trivalen (Cr3+)
yang tidak beracun sangat sulit diserap. Dalam beberapa kasus Cr-anorganik yang

29
dikonsumsi manusia lewat makanan 98% tidak diserap dan dikeluarkan lewat
feses (Amatya et al. 2004). Sebaliknya ketersediaan Cr-organik cukup tinggi,
tercatat 25 sampai 30 persen (Mordenti et al. 1997). Hasil-hasil penelitian Cr
menunjukkan bahwa selain esensial dalam metabolisme karbohidrat, Cr juga
dibutuhkan dalam metabolisme lemak dan protein dalam hal ini difisiensi Cr
dapat menyebabkan hiperkolesterolemia dan arterosklerosis serta rendahnya
inkorporasi asam amino pada protein hati. Ditambahkan bahwa asam amino yang
dipengaruhi oleh Cr adalah metionin, glisin dan serin (Anderson 1994). Burton
(1995) menambahkan bahwa Cr berperan dalam sitem kekebalan tubuh dan
konversi tiroksin (T4) menjadi triiodotironin (T3) yaitu hormon yang berperan
dalam meningkatkan laju metabolisme karbohidrat, lemak dan protein dalam hati,
ginjal, jantung dan otot serta meningkatkan sintesis protein. Kompleks Cr organik
terdapat dalam bentuk Cr-chelate, Cr proteinat ragi (high Cr-yeast) dan Cr-
pikolinat. Kromium pikolinat terbentuk dari Cr3+
Pencernaan mikroba pada ruminansia
yang mengikat 3 molekul asam
pikolinat. Asam pikolinat adalah metabolit sekunder yang dihasilkan pada
metabolisme triptofan sebelum membentuk nisin atau asam nikotinat (Combs
1992, Groff & Gropper 2000). Menurut Sahin et al (2011) yang melakukan
percobaan pada tikus yang diberi diet tinggi lemak mengatakan bahwa kondisi di
mana metabolisme glukosa terganggu karena resistensi insulin berkaitan dengan
gangguan memori, tambahan kromium (Cr) dapat mengurangi resistensi insulin
pada diabetes tipe 2 dan akibatnya meningkatkan akuisisi memori, tergantung
pada sumber dan tingkat. Selanjutnya hasil penelitian Toghyani et al (2010)
menunjukkan bahwa diet suplementasi Cr-yeast meningkatkan kualitas daging
ayam, paha ayam ras pedaging dalam kondisi stres panas
Ruminansia merupakan ternak yang unik, karena mempunyai sistem
pencernaan yang mampu merubah secara efisien sumber-sumber karbohidrat,
maupun bahan makanan kasar yang tedapat cukup di alam, hal ini dapat
berlangsung karena didalam rumenya terdapat mikroba yang mampu mencerna
serat kasar (Tillman et al 1998).

30
Pencernaan ada lah suatu rangka ian proses perubahan secara fisik dan kimia
yang terjadi pada pakan di dalam alat pencernaan hewan. Alat pencernaan
ruminansia terdiri dari mulut, perut, usus halus, dan alat pencernaan bagian
belakang (Hind gut). Perut ternak ruminansia terdiri dari empat bagian yaitu
rumen (pe rut beludru), retikulum (perut laja/sarang lebah), omasum (perut buku),
dan abomasum (perut sejati/ perut kelenjer). Tiga pertama disebut perut bagian
depan (fore gut) dan abomasum disebut perut sejati . Rumen dan retikulum
tergabung menjadi satu bagian dan disebut retikolorumen, didalamnya terdapat
bakteri, protozoa, fungsi dan virus yang kesemuanya berperan pada metabolisme
dalam rumen (Van Soest 1987).
Proses degradasi dan fermentasi pakan karbohidrat di dalam rumen di bagi
menjadi 3 tahap yaitu 1) pemecahan partikel makanan menghasilkan polimer
karbohidrat; 2) polimer dihidrolisir menjadi sakharida sederhana; 3) sakharida
sederhana menghasilkan Volatile fatty Acid (VFA). Pakan yang kaya akan
karbohidrat dirombak menjadi gula sederhana (maltosa, selobiosa, silosa dan
pentosa). Selanjutnya pruduk tersebut di konversi oleh enzim yang diproduks i
oleh bakteri rumen menjadi glukosa atau glukosa l- fosfat dan melalui proses
glikolisis dibentuk menjadi asam piruvat dan energi berupa ATP. Asam piruvat
yang terbentuk difermentasi di rumen dan menghasilkan VFA, yang dapat
menggambarkan fermentasi suatu pakan. Peningkatan kosentrasi VFA dapat
mencerminkan peningkatan protein asal pakan dan karbohidrat yang mudah larut
(readily available carbohydrate/RAC), lebih kurang 75% dari total VFA yang
dihasilkan diserap ke dalam retikulo-rumen, kemudian dalam abomasum dan
omasum, sedang sisanya 5% diserap dalam usus. Secara umum kandungan VFA
individual cairan rumen sapi 50-65% untuk asetat, 18-24% propionat dan 13-21%
butirat, sedangkan untuk domba mengandung 53-66% asetat, 19-27% propionat
dan 12-17% butirat dari persen molar total VFA. Produksi VFA cairan rumen
mencerminkan tingkat fermentasi suatu bahan. Semakin rendah suatu bahan
difermentasi semakin besar pula produksi VFA yang dihasilkan. VFA yang
dibutuhkan untuk pertumbuhan dan aktifitas mikroba maksimum 80-160 mM
(Arora 1995).

31
Kosentrasi amonia dalam rumen bervariasi dari 0-130 mM cairan rumen.
Protein yang berasal dari ransum masuk ke dalam rumen akan mengalami proses
degradasi oleh mikroba rumen menjadi peptida dan asam-asam amino.
Selanjutnya asam amino mengalami deaminasi amonia, CO2 dan mikroba dan
untuk menilai keefisienan penggunaan N pada ruminansia. Konsentrasi amonia
dipengaruhi oleh beberapa faktor, antara lain: 1) kandungan protein dalam ransum
serta kelarutannya, 2) jumlah karbohidrat dalam ransum, dan 3) waktu setelah
makan (Sutardi 1990).
Peningkatan karbohidrat yang mudah difermentasi (RAC) akan menurunkan
produk amonia, karena terjadi peningkatan penggunaan amonia untuk
pertumbuhan mikroba. Kondisi yang ideal adalah sumber energi sama cepatnya
difermentasi dengan pembentukan amonia, sehingga pada saat terbentuk amonia
terdapat juga rantai karbon dari fermentasi karbohidrat yang akan digunakan
sebagai sumber kerangka karbon protein mikroba yang telah tersedia (McDonald
et al. 2002).
Kebutuhan nutrisi pada domba
Nutrisi diperlukan oleh ternak untuk kebutuhan hidup pokok membangun
jaringan ba ru dan jaringa n tubuh yang mengalami kerusakan serta produks i,
sehingga dalam pemberian pakan untukternak domba harus memperhatikan
kandungan nutrisi dalam ransumnya serta disesuaikan dengan kebutuhannya
(Tillman et al. 1991). Standar kebutuhan nutrisi yang memenuhi persyaratan
kebutuhan domba di Indonesia, dengan pertambahan bobot badan harian (PBBH)
50 – 100 g/ekor/hari adalah BK 3.1 – 3.4% BB, PK 73.7 – 135.8 g/ekor/hari dan
energi 6.23 - 11.63 MJ/ekor/hari (Haryanto dan Djajanegara 1993). Ternak
domba lokal jantan umur 10 – 12 bulan dengan bobot badan 24.88± 3.77 kg yang
diberi pakan konsentrat 2% bobot badan dengan kandungan PK ±15%dan TDN
±70% menghasilkan rataan PBBH 122 g/ekor/hari (Ernawati dan Sunarso 2001).
Standar kebutuhan nutrisi per ekor per hari untuk domba di Indonesia disajikan
pada Tabel 5.
Anggorodi (1994) mengatakan bahwa faktor- faktor yang dapat menentukan
kebutuhan nutrisi adalah laju pertumbuhan, ukuran, jenis kelamin, kondisi

32
fisiologis dan lingkungan. Individu ternak dalam suatu bangsa yang memiliki laju
pertumbuhan cepat akan memerlukan nutrisi yang lebih banyak dibandingkan
ternak dengan laju pertumbuhan lambat (Tulloh dan William dalam Soeparno
1994). Selanjutnya menurut Berg dan Butterfield dalam Soeparno (1994), ternak
dengan ukuran tubuh yang besar memerlukan nutrisi yang lebih banyak daripada
ternak dengan ukuran kecil.
Beberapa faktor yang dapat mempengaruhi tingkat konsumsi pakan seekor
domba antara lain meliputi jenis dan tipe ternak, umur dan bobot badan, tingkat
produksi, pakan yang diberikan erta lingkungan tempat domba tersebut dipelihara
(Anggorodi 1994). Devendra dan Burns (1994) menyatakan bahwa bangsa
unggul dengan tingkat
Tabel 5 Standar kebutuhan energi dan protein per ekor per hari untuk domba di Indonesia
BB (kg) PBB (g)
Kebutuhan Energi Kebutuhan Protein Konsumsi Bahan kering (% BB)
DE (MJ) ME (MJ) TP (g) DP (g) 10 12 14 16 18 20
0 50 100 0 50 100 0 50 100 0 50 100 0 50 100 0 50 100
4.84 6.23 8.28 4.90 6.91 8.95 5.57 7.58 9.62 6.23 8.24
10.29 6.91 8.95
10.96 7.57 9.62
11.63
3.43 5.11 6.78 4.02 5.65 7.32 4.56 6.23 7.91 5.11 6.78 8.45 5.65 7.32 8.99 6.32 7.87 9.54
44.7 73.7 102.7 51.3 80.3 109.3 57.9 86.9 116.0 64.5 93.6 122.6 71.2 100.2 129.2 77.8 106.8 135.8
16.5 35.2 54.0 24.9 43.6 62.3 33.2 52.0 70.7 41.6 60.3 79.1 40.0 68.7 87.4 58.4 77.1 95.8
3.4
3.3
3.2
3.2
3.1
3.1
Sumber: Haryanto dan Djajanegara (1993) Keterangan:BB = bobot badan PBB= pertambahan bobot badan DE = d igestible energy ME= metabolizable energy TP = total protein DP= d igestible protein produktivitas tinggi cendrung mengkonsumsi pakan yang lebih banyak dibanding
dengan bangsa lokal. Haryanto dan Djajanegara (1993) menyatakan bahwa domba
tipe pedaging mengkonsumsi lebi banyak daripada tipe wool, dan domba dengan
bobot badan (BB) yang besar cendrung mengkonsumsi pakan lebih banyak

33
dibanding BB yang kecil. Cekaman panas sebagai akibat suhu lingkungan yang
tinggi akan menurunkan konsumsi pakan oleh ruminansia (Moose et al dalan
Rianto 1997). Jumlah konsumsi pakan dipengaruhi oleh kebutuhan BK dan zat-
zat pakan lainnya sesuai dengan BB dan fungsi fisiologisnya (Herman 1977).
Komposisi kimia dan kualitas dag ing
Daging adalah seluruh bagian karkas, tidak hanya terdiri dari jaringan otot,
tulang dan lemak, tetapi termasuk juga organ-organ tubuh dan kelenjar yang dapat
atau lazim dimakan. Daging terdiri dari otot, lemak dan sejumlah jaringan ikat
seperti jaringan epithel, syaraf dan pembuluh darah. Komposisi kimia daging
bervariasi dan dipengaruhi oleh beberapa faktor antara lain ; bangsa, umur, pakan
dan perbedaan pertumbuhan, termasuk perbedaan waktu penggemukan (Soeparno
1998).
Daging didefinisikan sebagai semua jaringan hewan dan semua produk hasil
pengolahan jaringan-jaringa n tersebut yang sesuai unt uk dimakan serta tidak
menimbulkan gangguan kesehatan bagi yang memakannya, keadaan fisik daging
dapat dikelompokkan menjadi (1) daging segar yang dilayukan atau tanpa
pelayuan, (2) daging yang dilayukan kemudian didingin-kan (daging dingin), (3)
daging yang dilayukan, didinginkan, kemudian dibekukan (daging beku), (4)
daging masak, (5) daging asap, dan (6) daging olahan. Karkas tersusun atas
kurang lebih enam ratus jenis otot yang berbeda ukuran dan bentuknya, susunan
syaraf dan persediaan darahnya serta perlekatannya pada bagian tulang dan tujuan
serta jenis geraknya. Kesehatan daging merupakan bagian yang penting bagi
kesehatan makanan dan selalu menjadi pokok permasalahan yang mendapatkan
perhatian khusus dalam penyediaan daging bagi konsumen (Aberley et al. 2001).
Daging yang dapat dikonsumsi adalah daging yang berasal dari hewan
yang sehat. Saat penyembelihan dan pemasaran berada dalam pengawasan
petugas rumah potong hewan serta terbebas dari pencemaran mikroorganisme.
Secara fisik, kriteria daging yang baik adalah berwarna merah segar, berbau
aromatis, memiliki konsistensi
yang kenyal dan bila ditekan tidak terlalu banyak mengeluarkan cairan. Daging
sebagai sumber protein hewani memiliki nilai hayati (biological value) yang

34
tinggi, mengandung 19% protein, 5% lemak, 70% air, 3,5% zat-zat non protein
dan 2,5% mineral dan bahan-bahan lainnya (Forrest et al. 1992). Komposisi
daging menurut Lawrie (2003) terdiri atas 75% air, 18% protein, 3,5% lemak
dan 3,5% zat-zat non
protein yang dapat larut. Secara umum, komposisi kimia daging terdiri atas 70%
air, 20% protein, 9% lemak dan 1% abu. Jumlah ini akan be rubah bila
hewan
digemukkan yang akan menurunkan persentase air dan protein serta
meningkatkan persentase lemak (Romans et al. 1994). Daging merupakan sumber
utama untuk mendapatkan asam amino esensial. Asam amino esensial terpenting
di dalam otot segar adalah alanin, glisin, asam glutamat, dan histidin. Daging sapi
mengandung asam amino leusin, lisin, dan valin yang lebih tinggi daripada daging
babi atau domba. Pemanasan dapat mempengaruhi kandungan protein daging.
Daging sapi yang dipanaskan pada suhu 70oC akan mengalami pengurangan
jumlah lisin menjadi 90 persen, sedangkan pemanasan pada suhu 160o
Lawrie (2003) menyatakan bahwa susunan kimia daging secara umum
terdiri dari 75% air, 19% protein, 2,5% lemak, 1,2% karbohidrat, dan 2,3% zat
terlarut dan vitamin. Protein daging terdistribusi pada miofibril 11,5%,
sarcoplasma 5,5% dan jaringan ikat 2%. Kadar protein daging relatif konstan
akan tetapi pada kasus tertentu perbedaan kadar protein pada urat daging
disebabkan karena perbedaan struktur daging yang terdiri atas protein miobril dan
jaringan ikat (kolagen, elastin dan retikulin). Actin dan myosin menyusun 75-
80% protein, sedangkan yang lain pada protein pengatur kontraksi yakni;
tropomin, tropomiosin, M-protein, C-protein, alfaactinin, dan beta actinin.
Kolagen ada lah komponen utama jaringan ikat, dan jaringan ini terdapat hampir
disemua komponen tubuh, sehingga kolagen paling banyak terdapat dalam tubuh
ternak.
C akan
menurunkan jumlah lisin hingga 50 persen. Pengasapan dan penggaraman juga
sedikit mengurangi kadar asam amino (Lawrie 2003).
Kualitas daging ditentukan oleh beberapa kriteria antara lain warna,
keempukan, flavor dan bau, cita rasa dan jueceness, kandungan lemak, susut
masak, retensi cairan dan pH daging (Soeparno 1998). Kadar kolagen daging

35
dipengaruhi oleh beberapa faktor, yaitu : kandungan lemak, umur ternak, dan
aktifitas gerak dari urat daging. Kualitas daging ditentukan oleh beberapa kriteria
antara lain: warna, keempukan, flavor, dan aroma (bau, cita rasa, dan juceness),
kandungan lemak intramusculair (marbling), Susut masak (cooking lost ), retensi
cairan, dan pH daging (Soeparno 1998).
Komponen lemak yang paling menentukan adalah lemak intramuskuler
(marbling), lemak tersebut sangat menentukan keempukan rasa dan aroma.
Daging yang dinilai baik adalah daging yang tingkat perlemakannya tidak terlalu
banyak, tetapi cukup mempunyai perlemakan di dalam urat daging (McPhee
2008). Daging yang hampir tidak mengandung marbling tampak kering dan yang
mempunyai flavor yang kurang baik, namun sebaliknya apabila marbling terlalu
banyak akan mengurangi palatabilitas.
Satuan produk karkas dinyatakan dalam bobot dan persentase. Persentase
karkas dipengaruhi oleh bobot karkas, bobot potong, kondisi ternak, bangsa,
proporsi bagian non karkas, dan makanan. Persentase karkas sangat bervariasi
antar 50-60% dari bobot hidup, rata-rata persentase karkas domba lokal adalah
43.60 persen (Sunarlin & Usmiati 2006). Faktor yang perlu diperhitungkan
dalam memperkirakan jumlah daging dari karkas dalah; 1) ketebalan lemak sub
kutan, 2) luas mata rusuk longisimus dorsi area, 3) persen lemak viscena
(penyelubung gijal, pelvis, dan jantung), dan 4) berat karkas (Swatland 1994).
Lawrie (2003) menyatakan bahwa pembagian karkas menjadi potongan-
potongan karkas sangat bervariasi pada beberapa negara atau daerah, berbeda dari
satu tempat ke tempat lainnya, disesuaikan dengan spesies ternak dan selera
konsumen. Karkas domba terbagi atas dua bagian besar yaitu forsadle (51%) dan
hindsadle (49%), forsadle (bagian depan) terdiri shuoulder, rack, freshank, dan
breast, sedangkan hindsadle (bagian belakang) terdiri dari loin, leg, dan flank.
Selanjutnya dinyatakan bahwa potongan utama dari karkas domba adalah leg,
rack, dan breast, Potongan-potongan komersial karkas tercantum pada Gambar 6.

36
Topside and silverside Steakmeal Aitchbone Rumo S irloin Forerib Midrib Clod and sticking
Leg Top rump Flank Briska l Shin Gambar 6 Potongan-potongan karkas ko mersial (Lawrie 2003)
Soeparno (1998) menyatakan bahwa, kandungan lemak pada daging
menentukan kualitas daging karena lemak menentukan cita rasa dan aroma
daging. Keragaman yang nyata pada kompos isi lemak terdapat antara jenis ternak
memamah biak dan ternak tidak memamah biak adalah karena adanya
hidrogenasi oleh mikroorganisme rumen. Protein daging terdiri dari protein
sederhana dan protein terkonjugasi, berdasarkan asalnya protein dapat dibedakan
dalam 3 kelompok yaitu protein sarkoplasma, protein miofibril, dan protein
jaringan ikat. Protein sarkop lasma adalah protein larut air karena umumnya dapat
diekstrak oleh air dan larutan garam encer. Protein miofibril terdiri atas aktin dan
miosin, serta sejumlah kecil troponin dan aktinin. Protein jaringan ikat ini
memiliki sifat larut dalam larutan garam. Protein jaringan ikat merupakan fraksi
protein yang tidak larut, terdiri atas protein kolagen, elastin, dan retikulin.
Kualitas fisik dag ing
Warna daging
Faktor utama yang menentukan warna daging yaitu konsentrasi pigmen
daging myoglobin, tipe molekul dan status kimia myoglobin. Faktor penentu
warna daging tersebut dipengaruhi oleh pakan, spesies, bangsa, umur, jenis
kelamin, stress (tingkat aktivitas dan tipe otot), pH dan oksigen (Purbowati et al.
2005). Pada umumnya, makin bertambah umur ternak, konsentrasi myoglobin

37
makin meningkat, tetapi peningkatan ini tidak konstan. Warna daging dapat
diukur dengan notasi atau dimensi warna “tristimulus”, yaitu: 1. hue = warna
(misalnya merah, hijau, dan biru), 2. nilai = terang atau gelap, dan 3. kroma =
jumlah atau intensitas warna. Warna daging domba bervariasi antara merah
terang hingga merah gelap. Dalam daging segar, sebelum dimasak bentuk kimia
yang paling penting adalah oksimioglobin. Walau itu terjadi dipermukaan saja,
pigmen ini sangat penting karena menggambarkan warna merah cerah yang
dikehendaki oleh konsumen (Lawrie 2003).
Nilai pH dag ing
Perubahan nilai pH sangat penting untuk diperhatikan dalam perubahan
daging postmortem. Nilai pH dapat menunjukkan penyimpangan kualitas daging,
karena berkaitan dengan warna, keempukan, cita rasa, daya mengikat air, dan
masa simpan. Konsentrasi glikogen otot pada saat pemotongan merupakan salah
satu faktor terpenting yang mempengaruhi kualitas daging. Glikogen adalah
subs trat metabolik dalam glikolisis postmortem yang menghasilkan asam laktat,
yang akan mempengaruhi pH otot. Proses glikolisis dan penurunan pH
berlangsung hingga cadangan glikogen habis atau terhentinya proses metabolik
terkait terhentinya proses enzimatik akibat pH yang rendah (Lukman et al 2007).
Aberley et al. (2001) menyatakan bahwa nilai pH daging ditentukan oleh
kadar glikogen dan asam laktat daging hewan setelah dipotong. Perubahan pH ini
tergantung dari jumlah glikogen sebelum dilakukan pemotongan. Bila jumlah
glikogen dalam ternak normal akan mendapatkan daging yang berkualitas baik,
tetapi bila glikogen dalam ternak tidak cukup atau terlalu banyak akan
menghasilkan daging yang kurang berkualitas, bahkan mendapatkan daging yang
berkua litas jelek. Laju penurunan pH daging secara umum dapat dibagi menjadi
tiga yaitu:
1. Nilai pH menurun secara bertahap dari 7.0 sampai berkisar 5.6 – 5.7 dalam
waktu 6 – 8 jam setelah pemotongan dan mencapai pH akhir sekitar 5.3 – 5.7.
Pola penurunan pH ini ada lah normal.

38
2. Nilai pH menurun sedikit sekali pada jam-jam pertama setelah pemotongan
dan tetap sampai mencapai pH akhir sekitar 6.5 – 6.8 . Sifat daging yang
dihasilkan adalah gelap, keras dan kering atau dark firm dry (DFD).
3. Nilai pH turun relative cepat sampai berkisar 5.4 – 5.5 pada jam-jam pertama
setelah pemotongan dan mencapai pH akhir sekitar 5.4 – 5.6 . Sifat daging
yang dihasilkan ialah pucat, lembek dan berair atau disebut pale soft
exudative (PSE).
Keempukan daging
Keempukan merupakan penentu kualitas daging domba. Komponen
utama yang menentukan keempukan ada lah jaringa n ikat dan lemak yang
berhubungan dengan otot (Aberle et al. 2001). Bertambahnya umur ternak akan
mengurangi tingkat keempukan dari daging karena ikatan silang intra dan
intermolekuler antara polipeptida kolagen meningkat. Pertumbuhan yang cepat
dapat mengurangi ikatan silang sehingga meningkatkan keempukan, perbedaan
bangsa juga dapat menimbulkan perbedaan keempukan daging, daging dari tipe
kecil lebih empuk dari pada daging dari tipe besar (Lawrie 2003). Menurut
Epley (2008) bahwa keempukan daging akan menurun seiring dengan
meningkatnya umur hewan. Jaringan ikat pada otot hewan muda banyak
mengandung retikuli dan memiliki ikatan silang yang lebih rendah jika
dibandingkan dengan hewan tua. Pemasakan daging dalam oven 135oC sampai
suhu dalam 50oC atau 60oC tidak mempengaruhi nilai daya putus Warner Bratzler
(Lawrie 2003). Perbedaan suhu dalam daging saat pemasakan (60oC, 70oC,
80oC) akan mempengaruhi keempukan dari daging, semakin tinggi suhu akhir
pemasakan akan menghasilkan daging yang lebih empuk. Suhu akhir (60oC,
70oC, 80oC) secara akurat dapat digunakan sebagai alat untuk klasifikasi
keempukan daging, tetapi pada suhu yang rendah (60oC) perbedaan suhu dalam
daging tidak dapat dijadikan patokan yang akurat untuk klasifikasi keempukan
daging karena dipengaruhi oleh waktu pemasakan, jumlah perubahan jaringan
dan rendahnya nilai klasifikasi keempukan (Wheeler et al. 1999). Combes et al.
(2002) menyatakan bahwa nilai keempukan daging dengan Warner Bratzler
mencapai minimum pada suhu dalam 60-65oC dan meningkat kembali mencapai
maksimum pada suhu dalam daging 80-90oC. Keempukan daging berkisar antara

39
3.83-5.49 dan secara statistik hampir sama, hal ini disebabkan komposisi PK dan
lemak yang hampir sama.
Faktor yang mempengaruhi keempukan daging dapat digolongkan menjadi
dua, yakni faktor antemortem dan faktor postmortem.
Kriteria keempukan menurut Suryati dan Arief (2005) berdasarkan panelis
yang terlatih menyebutkan bahwa daging sangat empuk memiliki daya putus WB
(Warner Bratzler) < 4.15 kg/cm
Faktor antemortem tersebut
meliput i genetik termasuk bangsa, spesies dan fisiologi, umur, manajemen, jenis
kelamin, dan stress. Faktor postmortem diantaranya adalah metode chilling,
refrigerasi, pelayuan, dan metode pengolahan. Jadi keempukan bisa bervariasi
antara spesies, bangsa, ternak dalam spesies yang sama, potongan karkas, dan
diantara otot, serta pada otot yang sama. Keempukan daging ditentukan oleh 3
komposisi daging yaitu : 1. Struktur miofibril dan status kontraksinya, 2.
Kandungan jaringan ikat dan tingkat ikatan silang dan 3. Daya ikat air oleh
protein daging dan marbling (Abe rle et al 2001). Perbedaan bangsa juga dapat
menimbulkan perbedaan keempukan daging, daging dari tipe kecil lebih empuk
daripada daging dari tipe besar (Lawrie 2003). Menurut Epley (2008)
keempukan daging akan menurun seiring dengan meningkatnya umur hewan.
Jaringan ikat pada otot hewan muda banyak mengandung retikuli dan memiliki
ikatan silang yang lebih rendah jika dibandingkan dengan hewan tua.
2, daging empuk 4.15 - < 5.86 kg/cm2, daging
agak empuk 5.86 - < 7.56 kg/cm2, daging agak alot 7.56 - < 9.27 kg/cm2, daging
alot 9.27 - < 10.97 kg/cm2, daging sangat alot = 10.97 kg/cm2
Keempukkan daging ditentukan oleh tiga komponen daging yaitu; 1)
struktur miofibril dan status kontraksi, 2) kandungan jaringan ikat dan ikatan
silang, dan 3) daya ikat air oleh protein daging dan marbling (Soeparno 1998).
Tingkat keempukkan daging dapat dihubungkan dengan tiga katagori protein
otot yaitu; 1) protein jaringan ikat (ko lagen, elastin, dan mukopolisakarida),
2) miofibril (miosin, actin, dan tropomiosin), dan 3) sarkoplasma (protein
sarkoplasmatik, dan sarkoplas-matik retikulum). Kontribusi masing-masing
kategori protein tersebut tergantung pada tingkat kontraksi miofibril, tipe otot dan
lama serta suhu pemasakan (Lawrie 2003).
.

40
Daya mengikat air (DMA)
Daya mengikat air (DMA) oleh protein daging atau water holding capacity
merupakan suatu nilai yang menunjukkan kemampuan untuk mengikat air atau
cairan baik yang berasal dari dalam maupun dari luar yang ditambahkan. Daya
mengikat air merupakan salah satu faktor yang menentukan kualitas dan daya
terima daging oleh konsumen. Pengukuran banyak air yang hilang atau drip
merupakan hal yang penting dalam penentuan rantai harga, karena mempengaruhi
bobot daging. Tingkat daya mengikat air ini ditentukan oleh spesies, genetik, laju
glikolisis, pH akhir, proses pemotongan dan waktu (Honikel 1998). Fungsi atau
gerakan otot yang berbeda mengakibatkan perbedaan jumlah glikogen yang
menentukan besarnya pembentukan asam laktat dan akhirnya menghasilkan DMA
yang berbeda. Lawrie (2003) menambahkan bahwa daya mengikat air daging
sangat dipengaruhi oleh pH, semakin tinggi pH akhir semakin sedikit penurunan
DMA. Daya mengikat air sangat penting dalam proses pengolahan daging
sebaga i protein yang mampu menahan lebih banyak air menjadi lebih mudah
larut. Daya mengikat air menurun dari pH tinggi (sekitar 7-10) sampai pada pH
titik isoelektrik protein-potein daging antara 5.0 – 5.1.
Daya ikat air (DMA) daging adalah kemampuan daging untuk mengikat
airnya atau air yang ditambahkan selama ada pengaruh kekuatan dari luar,
misalnya pemotongan daging, pemanasan, penggilingan, dan tekanan. Daya ikat
air dipengaruhi oleh perbedaan macam otot, species, umur dan fungsi otot. Fungsi
atau gerakan otot yang berbeda mengakibatkan perbedaan jumlah glikogen yang
menentukan besarnya pembentukan asam laktat dan akhirnya menghasilkan DIA
yang berbeda. Daya ikat air menurun dari pH tinggi (sekitar 7-10) sampai pada pH
titik isoelektrik protein-potein daging antara 5,0–5,1
Salah satu faktor yang
mempengaruhi DMA daging adalah umur ternak. Semakin tua umur ternak,
kapasitas memegang air daging lebih sedikit.
(Purbowati et al. 2005).
Lawrie (2003) mengatakan bahwa salah satu faktor yang mempengaruhi DMA
daging adalah umur ternak. Semakin tua umur ternak, kapasitas memegang air
daging lebih sedikit.

41
Susut masak
Susut masak dipengaruhi oleh temperatur dan lama pemasakan. Semakin
tinggi temperatur pemasakan maka semakin besar kadar cairan daging yang hilang
sampai mencapai tingkat yang konstan. Beberapa faktor yang memepengaruhi
susut masak adalah pH, panjang sarkomer serabut otot, panjang potongan serabut
otot, status kontraksi myofibril, ukuran dan berat sampel daging dan penampang
lintang daging (Soeparno 1998). Daging dengan susut masak yang lebih rendah
mempunyai kualitas yang lebih baik daripada daging dengan susut masak yang
lebih besar, karena kehilangan nutrisi selama pemasakan akan lebih sedikit.
Besarnya susut masak dapat dipergunakan untuk mengestimasi jumlah jus dalam
daging. Kesan jus daging atau juiciness mempunyai hubungan yang erat dengan
susut masak. Kadar jus daging yang rendah dapat disebabkan oleh susut masak
yang tinggi. Kadar jus daging yang rendah dapat disebabkan oleh susut masak
yang tinggi. Daging dengan susut masak yang lebih rendah mempunyai kualitas
yang relatif lebih baik daripada daging dengan susut masak yang lebih besar,
karena kehilangan nutrisi selama pemasakan akan lebih sedikit. Perebusan daging
pada suhu 60 – 90oC menyebabkan rusaknya jaringan epimisium, perimisium dan
endomisium sehingga myofibril menyus ut yang menstimulasi keluarnya cairan
daging. Susut masak merupakan fungsi dari temperatur dan lama pemasakan.
Beberapa faktor yang mempengaruhi susut masak adalah pH, panjang sarkomer
serabut otot, panjang potongan serabut otot, status kontraksi myofibril, ukuran dan
berat sampel daging dan penampang lintang daging. Daging dengan susut masak
yang lebih rendah mempunyai kualitas yang lebih baik daripada daging dengan
susut masak yang lebih besar, karena kehilangan nutrisi selama pemasakan
akan lebih sedikit. Besarnya susut masak dapat dipergunakan untuk
mengestimasi jumlah jus dalam daging, jus daging atau juiciness mempunyai
hubungan yang erat dengan susut masak, kadar jus daging yang rendah dapat
disebabkan oleh susut masak yang tinggi, umumnya susut masak bervariasi
dengan kisaran antara 15-40% (Soeparno 1998). Selanjutnya Lawrie (2003)
menjelaskan bahwa bobot potong dapat mempengaruhi susut masak apabila
terdapat perbedaan deposisi lemak intramuskuler (marbling). Daging dengan
susut masak yang lebih rendah mempunyai kualitas yang relatif lebih baik dari

42
pada daging dengan susut masak yang lebih besar, karena kehilangan nutrisi
selama pemasakan akan lebih sedikit.





























