Embed Size (px)
Citation preview

Comp. Biochem. Physiol. Vol. 85B, No. 1, pp. 23-30, 1986 0305-0491/86 $3.00 + 0.00 Printed in Great Britain Pergamon Journals Ltd
HETEROGENEOUS H UMORAL A N D HEMOCYTE- ASSOCIATED LECTINS WITH N-ACYLAMINOSUGAR
SPECIFICITIES FROM THE BLUE CRAB, CALLINECTES SAPIDUS RATHBUN*
FREDERICK J. CASSELS, JOHN J. MARCHALONIS and GERARDO R. VASTA~" Department of Biochemistry, Medical University of South Carolina, Charleston, SC 29425, USA
(Tel: 803-792-4321)
(Received 11 November 1985)
Abslraet--l. CaUinectes sapidus serum and hemocyte microsomal fraction agglutinated a panel of untreated and enzyme treated vertebrate erythroeytes and cultured lymphoid cell lines.
2. Crossed absorption experiments suggested the presence of multiple specific lectins in the serum. 3. The microsomal fraction showed a 35-fold increase in specific activity when compared to the
hemocyte lysate suggesting that hemocyte lectins are membrane-associated. 4. Agglutination by serum and hemocyte lectins was inhibited by low concentrations of N-acylamino
compounds including sialic, N-acetylmurarnic and N-acetylglutamic acids, GalNAc, GIcNAc, ManNAc, and glycoproteins and polysaccharides which contain these carbohydrates: bovine submaxillary mucin, human orosomucoid, porcine stomach mucin and colominic acid.
5. Hemagglutination by lectins of both serum and hemocyte microsomal fraction required divalent cations as suggested by the reduction in hemagglutination titer in the presence of the chelators EDTA, EGTA, CDTA and citrate.
INTRODUCTION
Lectins in the invertebrates have so far proven to be ubiquitous (Gold and Balding, 1975; Cohen and Vasta, 1982), with serum lectins first studied (Noguchi, 1903) then much later tissue lectins (Prokop et al., 1965) and hemocyte plasma mem- brane lectins (Tyson and Jenkin, 1974; Amirante and Mazzalai, 1978; Vasta et al., 1982, 1984). Hemocyte associated membrane lectins have been demonstrated in a variety of arthropod and molluscan species including the crayfish Parachaeraps bicarinatus (Tyson and Jenkin, 1974); the cockroach Leucophaea maderae (Amirante and Mazzalai, 1978); the oyster Crassostrea virginica (Vasta et al., 1982, 1984); the mussel Mytilus edulis (Renwrantz and Stahmer, 1983); the mantis shrimp Squilla mantis (Amirante and Basso, 1984) and the blue crab Callinectes sapidus (Vasta and Cassels, 1983). In addition, lobster (Homarus americanus) hemocyte lysates have demon- strated high lectin specific activities (Cornick and Stewart, 1973; Hall and Rowlands, 1974a). In lob- sters (Cornick and Stewart, 1973) and cockroaches (Amirante and Mazzalai, 1978) hemocytes are the proposed site of synthesis of serum lectins.
A number of studies have demonstrated the presence of humoral lectins in crustacean species
*This work was supported by National Institutes of Health Grant No. GM 30672 to JJM and GRV. Support for F.J.C. was provided from the College of Graduate Studies, Medical University of South Carolina Institu- tional Funds. This work is in partial fulfillment of the requirements for the degree of Doctor of Philosophy.
1"To whom correspondence should be addressed.
(Vasta and Marchalonis, 1983, in review list 34 species). However, the carbohydrate specificities of serum lectins from crustaceans have been elucidated only in a few species. The lobster Homarus ameri- canus exhibits one lectin specific for sialic acids and a second lectin specific for N-acetyl-D-galactosamine (Hall and Rowlands, 1974b); the freshwater prawn (Macrobrachium rosenbergii) has one lectin fraction specific for sialoconjugates and a second undeter- mined (Vasta et al., 1983); the coconut crab Birgus latro exhibits one lectin fraction specific for sialic acids and a second for undetermined carbohydrate moieties (Vasta and Cohen, 1984). Ravindranath et al. (1985) described a lectin from the crab Cancer antennarius with specificity toward O-acetylated sialic acids. Brown et al. (1968) found that Callinectes danae serum agglutinated human erythrocytes; Pauley (1974) reported on a serum lectin from Calli- nectes sapidus reactive to rabbit, chicken and human erythrocytes; and McCumber et al. (1979) demon- strated that a viral neutralization factor was present in Callinectes sapidus hemolymph and that it was distinct from the lectin which binds to mouse erythro- cytes. We reported earlier, in the blue crab Callinectes sapidus, heterogeneous populations of serum and hemocyte microsomal lectins that exhibit specifici- ties for N-acetylated acids and amines (Vasta and Cassels, 1983).
This study describes the preliminary characteriz- ation of Callinectes serum and hemocyte associated lectins through agglutination and crossed absorption experiments; a comparison of their carbohydrate specificities through inhibition experiments with mono-, oligo-, polysaceharides and glycoproteins; and describes evidence for their divalent cation requirements.
23

24 FREDERICK J. CASSELS et aL
MATERIALS AND METHODS
Source of materials
Live adult Callinectes sapidus were collected in Charleston Harbor, South Carolina by trawl and by trapping. The crabs were taken to the laboratory, where bleeding pro- ceeded immediately or after maintaining the crabs overnight in a 10°C tank with filtered, recirculated brackish harbor water (salinity 12 parts per thousand). Crabs stored over- night were bled after warming to room temperature.
Collection of hemocytes and serum
Crabs of 12.7 cm carapace width or larger were bled with 18 or 20 gauge needles from the arthrodial membrane of the cheliped merus. To obtain serum the blood was withdrawn into a 20 ml syringe, pooled on ice in lots of ten individuals or greater and allowed to clot at 4°C. For plasma and hemocyte collection, hemolymph was withdrawn into a syringe containing precooled (4°C) anticoagulation buffer (Sfderh~ll and Smith, 1983) (0.45M NaC1, 0.1 M glucose, 30 mM trisodium citrate, 26 mM citric acid and 10mM EDTA, pH 4.6) for a final ratio of approximately 1:1. Plasma from females and males was pooled separately on ice and then centrifuged at 800 g for 5 rain at 4°C. The superna- rant (plasma) was exhaustively dialyzed against crustacean Ringers solution (McCumber et al., 1979) (0.525 M NaC1; 13 mM KC1; 12 mM CaC12; 25 mM MgCI 2, pH 7.0 with NaHCO3). Sodium azide was added to samples of serum and plasma to a final concentration of 0.05% and stored at -25°C until use.
Cell fractionation
The cell pellet was washed once in the anticoagulation buffer, twice in crustacean Ringers solution, then Dounce homogenized with 25 strokes. From this point the procedure follows the differential centrifugation protocol of Vasta et al. (1982). Briefly, the lysate was centrifuged at 300g for 5 rain (all centrifugation was done at 4°C) and the nuclear pellet removed; the supernatant was centrifuged at 2000g for 10 min to yield the mitochondrial pellet; this supernatant was then centrifuged at 48,000g for 60 rain to yield the microsomal pellet. The microsomal pellet was resuspended, Dounce homogenized in hypotonie buffer (10 mM Tris-HC1 pH 7.4), recentrifuged and resuspended in 10 mM Tris- HCI, pH 7.4, 0.14M NaC1.
Reagents
N-acetylneuraminic acid (NeuAc) type VI from E. coli, N-glycolylneuraminic acid (NeuGc), methoxyneuraminic acid, N-acetylneuraminlactose grade I from bovine colostrum (sialyllactose), D-glucuronic acid sodium salt, D-galacturonic acid monohydrate, N-acetyl-D-glucosamine grade III (GlcNAc), N-acetyl-o-galaetosamine (GalNAe), N-acetyl-fl-D-mannosamine grade II (ManNAc), N-acetyl- muramic acid, N-acetylglutamic acid, muramic acid, D-galactosamine, D(+)-mannose, o-glucose, D-galactose, "-L-(--)-fucose, "-L-rhamnose, 2-keto-3-deoxyoctonate ammonium salt, arabinogalactan, colominic acid sodium salt from E. coli, bovine thyroglobulin type I, bovine submaxillary mucin (BSM), bovine glycoprotein fraction VI orosomucoid, human ,,t-acid giycoprotein purified from Cohen fraction VI orosomucoid, porcine stomach mucin (PSM), ovomucoid Type III-o purified, free of ovoinhibitor and pronase P (protease) type VI purified from Strepto- mvces griseus (3-4 units/mg) (Pr), EGTA (ethylene glycol- bis-(fl-amino ethyl ether)N,N,N',N'-tetraacetic acid), and CDTA (trans- 1,2-diaminocyclo-hexanetetraacetic acid), were purchased from Sigma Chemical Company, St. Louis, Missouri. Neuraminidasc (VCN) from Vibrio cholerae, 500 U/ml, and fetuin were purchased from (3IBCO, Grand Island, New York. N-acetylneuraminic acid ,a-methyl glyco- side was kindly supplied by Dr. M. Sharma, RosweU Park Memorial Institute, Buffalo, New York. EDTA (disodium
ethylenediamine tetraacetic acid), and sodium citrate were purchased from Fisher, Norcross, Georgia.
Desialylation of glycoproteins
Glycoproteins were dissolved in 0.I N H2SO4 0.85% NaC1 at concentrations of 10 mg/ml. Hydrolysis was carried out for 1 hr at 80°C. Hydrolyzed giycoproteins were exhaus- tively dialyzed against 0.85% NaCI (24 hr, with 2 changes of saline) and Tris°buffered saline (TBS; 0.10M NaC1, 0.05 M Tris-HCl, 0.02M CaC12, 0.05% NaN 3, pH 7.6) (24 hr, with 2 changes TBS), aliquoted and stored at - 25°C.
Erythrocytes and lymphoid cell lines
Horse erythrocytes (RBC) in Alsevers solution were pur- chased from Chisolm Labs, Seabrook, South Carolina. Human blood samples collected in ACD (citrate-dextrose) were obtained from the blood bank at the Medical Univer- sity of South Carolina. Clear nose skate (Raja eglanteri) blood was obtained by cardiac puncture and RBC were separated from the teased clot. The blood samples from all other species were obtained by venous or cardiac puncture and collected in Alsevers solution. RBC were washed once in 0.85% NaC1 and twice with TBS and were suspended at concentrations of 5 x l0 6 cells per ml TBS. Packed RBC were treated with Pr (3-4 U/ml) or VCN (500 U/ml) as described by Uhlenbruck et al. (1968) and were suspended in TBS as above. Lymphoid cells tested were established T cell lines (70-N2 and 22-CM-37) and B cell lines (1605L) from the cotton topped marmoset (Sanguinus oedipus) and from a human T cell line (HUT78). All were maintained on RPMI 1640 medium supplemented with 5% Nu-Serum, 2 mM L-glutamine, 50 U/ml penicillin and 50 #g/ml strepto- mycin. Washing, enzyme treatment and cell suspension preparations were done as described above.
Agglutination test
5#1 of 2-fold serial dilutions of Callinectes serum or hemocyte microsomal preparation in TBS were placed in Terasaki 96 well plates (Robbins Scientific, Mountain View, California) and equal volumes of the erythrocyte sus- pensions were added to each well. The plates were gently vortex-mixed, incubated at room temperature for 45 min and read under a microscope (I00 x ) with the degree of agglutination graded from 0 (negative) to +4. Titers were recorded as the inverse ratio of the highest dilution showing a + 1/2 degree of agglutination. The substitution of serum or microsomal preparation with TBS served as controls for all titrations.
Crossed absorption test
Serum diluted 1/8 in TBS was mixed with one third of its volume in washed, packed, Pr- and VCN-treated RBC and incubated for 45 min with gentle mixing every 15 min. After centrifugation at 800g for 3 min the supernatant was with- drawn and titrated as described above.
Hemagglutination-inhibition tests
Five microliters of serial two-fold dilutions of serum or microsomal preparation were added to Terasaki plates, followed by 5 #I of the selected inhibitor (mono-, oligo-, polysaccharide or glycoprotein) at the given concentration, gently vortex-mixed and incubated for 30 rain. Two micro- liters of selected RBC cell suspension (107 cells per ml) was added to each well, vortex-mixed and incubated for 45 rain. The percent reduced score at each inhibitor concentration was plotted and 50% inhibition value was recorded.
Divalent cation requirements
Studies on divalent cation requirements were performed in the same manner as the hemaggiutination-inhibition assays, with the use of the divalent cation chelators EDTA, EGTA, CDTA and sodium citrate at different concen- trations in place ofinhibitors, and with an incubation period
Heterogeneous humoral and hemocyte crab lectins 25
of 45 rain. The reduction in titer relative to the control (chelator substituted by TBS) was plotted with results expressed as the mean of three independent values plus or minus the SE.
R E S U L T S
The association of lectin activity with hemocytes is suggested by the results given in Table 1. All cell fractions agglutinated both horse and rabbit RBC, with the exception of the nuclear (300g) pellet. The microsomal (48,000 g) pellet showed an increase in specific activity over each subfraction and a 34.7-fold increase in specific activity over that of the lysate, indicating that Callinectes membranes are a rich lectin source. The final supernatant retained a considerable level of specific activity indicating the presence of lectin in the cytosol.
Hemagglutination by the serum and microsomal preparation (48,000 g pellet) was assayed by using a panel of vertebrate RBC both untreated and enzyme treated (Table 2). Callinectes serum exhibited slightly higher agglutination titers with human A RBC as compared to B and O. This level of discrimination
was not exhibited by the microsomal preparation. Pr treatment of the RBC led to increased serum titers with the majority of RBC, and all RBC except rabbit when tested with the microsomal preparation indi- cating that a number of additional receptors for lectin were uncovered on the RBC by the proteolytic activ- ity. Decreased serum titers after VCN treatment of erythrocytes were observed for human A, B, horse, dog, mouse and duck. Typical sialic acid specific agglutination patterns for serum were suggested with mouse and dog RBC, while only dog showed this pattern when tested with microsomal preparation. Although titers for untreated and enzyme treated human RBC were similar for both serum and micro- somal preparation, titers against horse, cat, rabbit and rat were markedly different suggesting that the distribution of those multiple specificities is different for serum and microsomal preparation. Three T cell lines, HUT78, 70-N2 and 22-CM-37 and a B cell line, 1605L were also tested for agglutination with Calli- nectes plasma (Table 3). These cultured cell lines were agglutinated at relatively low titers as compared to RBC. 22-CM-37 was not agglutinated whether untreated or enzyme treated. Pr-treated HUT78,
Table 1. Agglut inat ing activity o f cen fractions
Fraction Recovered AU 1
Horse Rabbit
from disrupted Callinectes sapidus hemocytes
Percentage Total Specilic Act iv i ty 2 Recover),
Horse Rabb i t Horse Rabbit
Lysate 1024.0 256.0 100.0 100.0 3.68 0.92
Nuclear pellet (300 g) 19.2 0.0 1.9 0.0 0.18 0.00
Mitochondrial pellet (2,000 g) 128.0 64.0 12.5 25.0 56.89 28.44
Microsomal pellet (~g,000 g) 96.0 24.0 9.4 9.4 128.00 32.00
Supernatant t40.8 70.4 13.8 27.5 6.40 3.20
tFigures are in agglutination units (AU), defined as the amount of lectin present in the highest dilution of serum capable of agglut inat ing RBC with a score o f + 2 in a scale from 0 to +4.
2Specific activity expressed as AU/prote in .
Table 2. Agglut inat ion of vertebrate erythrocytes by Callinectes sapidus serum and microsomal preparation ~
Cell Type Serum 2 Microsomal Preparation
Unt 3 Pr VCN Unt Pr VCN
Human A 512 512 32 64 256 16
Human B 64 64 32 32 64 32
Human O 32 64 64 32 1215 g
Horse 4096 20gg 1024 g 32 16
Goat 2 16 8 0 g 0
Dog 16 32 I 2 8 0
Cat 0 5124 g 0 4 16
Rabbit 512 2048 0096 32 32 32
Rat 256 4096 512 ~ 16 8
Mouse 64 32 0 I 8 8
Duck 128 64 16 0 $ 1
Skate 0 16 2 0 2 0
~Figures are highest di lut ion at which + 1/2 reaction observed on a scale from 0 to +4 .
2Serum and microsomal preparation was obtained from pools of 10 adult male specimens.
3Abbreviations: Unt , untreated; Pr, pronase-treated; VCN, neuraminid- ase-treated.
4Prozone noted.

26 FREDERICK J. CASSELS et al.
Table 3. Agglut inat ion o f lymphoid cells lines by Callinectes sapidus plasma I
Cell Line Enz),me Treatment Unt 2 Pr VCN
HUT7g 8 # 2
70-N2 16 0 16
22-CM-37 0 0 0
160.~L 16 0 16
~Figures are highest di lut ion at which + 1 / 2 reaction observed on a scale from 0 to + 4.
2Abbreviations: Unt , untreated; Pr, pronase- treated; VCN, neuraminidase-treated.
70-N2 and 1605L exhibited decreased agglutination titers suggesting that Pr-sensitive glycopeptides were involved in the binding of Callinectes serum lectins. VCN treatment decreased Callinectes serum titers for HUT78 indicating that cell surface sialic acids are important for agglutination. Cleavage Of terminal sialic acids by VCN did not affect titers for 70-N2 and 1605L. 22-CM-37 cells very likely exhibit some sialic acids on the cell surface, as do cat RBC (Table 2), but neither was agglutinated by Callinectes lectins. This is not surprising, however, and it has been observed for several lectins. Since lectins are very large molecules a particular conformation of the cell surface glyco- peptide might interfere with the binding and/or agglutination. Hemagglutinating activity of pooled serum and plasma from male specimens and pooled serum and plasma from female specimens was tested with selected untreated and enzyme-treated RBC (data not shown). No significant differences in agglutination titers were found between pooled male and female serum and between pooled male and female plasma. The agglutination titers of pooled male serum and plasma showed no significant differences from pooled female serum and plasma.
The crossed absorption experiments (Table 4) sug- gest that multiple lectin specificities exist in the serum of Callinectes. One distinct lectin specificity is sug- gested by the cross-reactions between Pr-treated duck and human O RBC, where absorption with the first RBC removed agglutinating activity for the second. This pattern of cross-reactions was repeated with the
Pr-treated rat and horse RBC and VCN-treated horse RBC, suggesting a second lectin specificity. A third specific lectin fraction appears to bind to Pr-treated rabbit RBC, as Pr-treated rabbit RBC did not absorb out activity for the other RBC tested. Pr-treated goat and VCN-treated duck RBC were not agglutinated by Callinectes serum and did not significantly modify titers when used as the test absorbants.
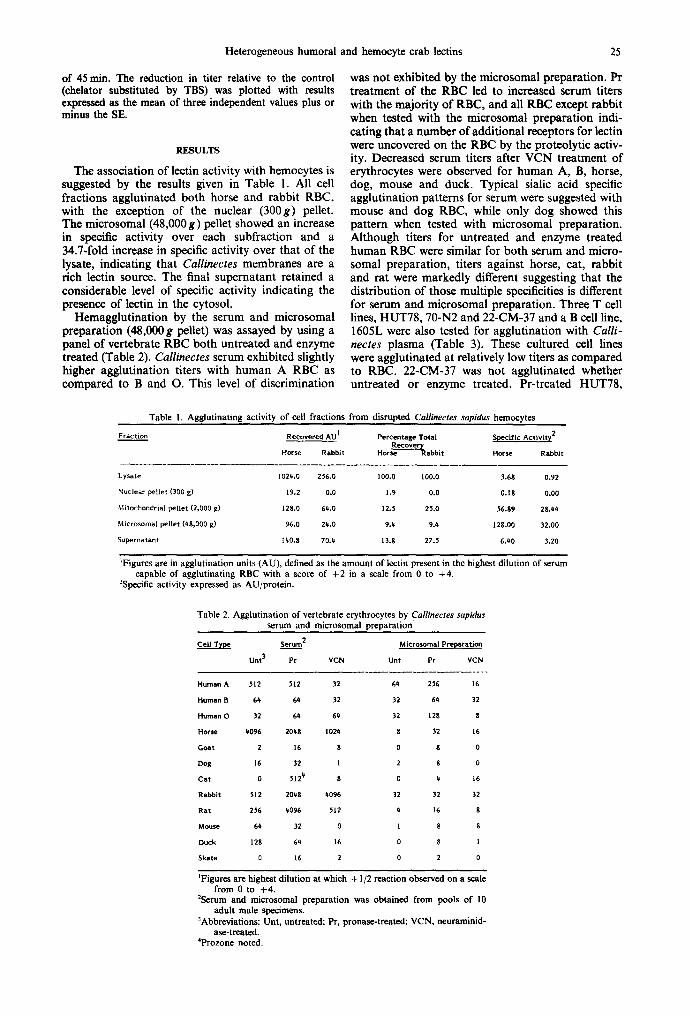
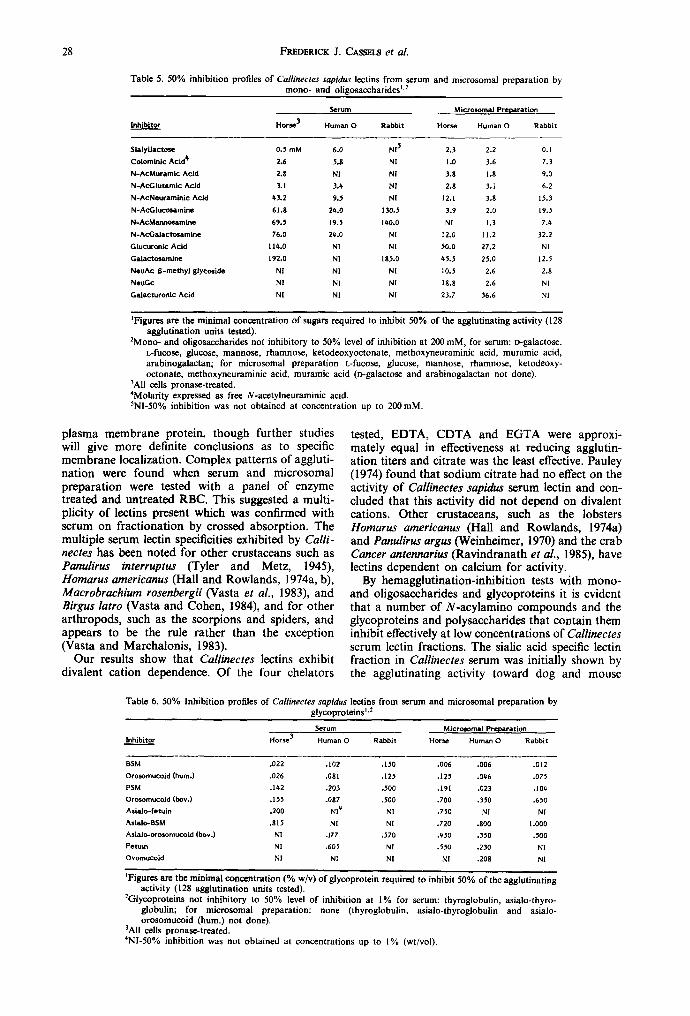
The existence of non-cross-reacting multiple spe- cific serum lectins was confirmed by comparison of qualitative and quantitative differences in inhibitory capabilities of mono- and oligosaccharides for differ- ent RBC [Figs l(a--c) and Table 5]. Hemagglutin- ation-inhibition results were plotted as percent inhibition versus inhibitor concentration (Fig. 1). The concentrations of inhibitor at the 50% level of inhibition are shown in Table 5. Those mono- and oligosaccharides that inhibited hemagglutination by both serum and microsomal preparation at the lowest concentrations are N-acetylated acids and amines. Although free N-acetylneuraminic acid behaved as a strong inhibitor, particularly for the microsomal preparation, sialyllactose (NeuAc-a-2-3(6)-Gal-fl-l- 4-Glc) and colominic acid (-NeuAc-~-2-8-NeuAc-) inhibited hemagglutination at lower concentrations, indicating that the anomeric linkage to the sub- terminal sugar in addition to the terminal sugar is an important factor for the lectin binding. Only high concentrations of GlcNAc, ManNAc and galactos- amine inhibited the hemagglutination of rabbit RBC by serum. The inhibition of hemagglutination between microsomal preparation and Pr-treated rab- bit RBC did not follow this pattern as a number of mono- and oligosaccharides inhibited at low concen- trations indicating different lectin specificities from serum. Although the best inhibitors for Callinectes lectins were those with bound sialic acids and free N-acetylneuraminic acid, the fact that these lectins were inhibited also by other N-acyl amino sugars present on the cell surface (i.e. GalNAc) either linked to glycoproteins or glycolipids would explain our observation that for most RBC tested VCN treat- ment did not prevent lectin binding. A complex array of carbohydrate binding molecules appears to be present in Callinectes serum and the possibility of existence of lectins binding to the carbohydrate
Table 4. Crossed absorption patterns o f Callinectes sapidus serum ~
A b ~ ' b e d with Tested with Goa~ Duck Duck Human 0 Rat Horse Horse Rabbit
Pr" VCN Pr Pr Pr Pr VCN Pr
None (control) 0 0 8 16 64 32 128 6#
Goat Pr 0 0 16 0 16 32 64 32
Duck VCN 0 0 16 ~ 32 32 128 32
Duck Pr 0 0 1 0 16 32 60 16
Human O Pr 0 0 0 0 16 32 64 60
Rat Pr 0 0 0 0 1 0 2 32
Horse Pr 0 1 8 2 0 0 0 32
Horse VCN 0 0 2 2 2 0 0 12~
Rabbit Pr 0 0 64 0 16 32 6t~ 0
~Figures are highest dilution at which + 1/2 reaction observed, on a scale from 0 to +4 . 2Pr, pronase-treated; VCN, neuraminidase-treated.
Heterogeneous humoral and hemoeyte crab lectins 27
1 0 0
SO
0
100 B
.o • ~ so t -
_=
0
1 0 0 C
5 0
0 , , 1.e 3.1 0.3 12.s 2s so Ioo 200
Inhibitor Concentrat ion {mMI
Fig. 1. Representative hemagglutination-inhibition profiles of Callinectes serum with several mono- and oligosacchar- ides (see 50°/, inhibition values for all sugars tested in Table 5). (A) Horse Pr-treated erythroeyte profile, (B) human O Pr- treated erythrocyte profile, (C) rabbit Pr-treated erythro- cyte profile. Inhibitors: sialyUactose, ~ ; N-acetylneuraminic acid, /X; glucuronic acid, &; N-acetyl glutamic acid, Irl; N-acetylgalactosamine, I-1; N-acetyl glucosamine, II; N- acetyl mannosamine, I]; D-galactosamine, []; L-fucose, O; D-glucose, tD; mannose, O; colominic acid, O. (See
Materials and Methods for experimental details.)
residues subterminal to sialic acids appears likely. In addition, RBC from horse, cat and rat exhibit VCN-resistant cell surface sialic acids.
Hemagglutination-inhibition results obtained with untreated and desialylated glycoproteins are shown in Table 6. Inhibition data for serum show that BSM inhibited hemagglutination of Pr-treated horse, human O and rabbit RBC at low concentrations but that on removal of sialic acid by acid hydrolysis the glycoprotein was no longer inhibitory to a 50% level (for Pr-treated human O and rabbit RBC) or decreased the level of inhibition 37-fold (Pr-treated horse RBC). The inhibition values for BSM inhibit- ing the microsomal preparation were similar to the values for BSM inhibiting serum. Agglutination of the three RBC types was inhibited with low concen- trations of BSM; when desialylated BSM was tested, inhibition levels decreased 83-113-fold. A similar pattern of removing or reducing inhibitory capabili- ties for Callinectes serum lectins of a glycoprotein by
desialylation was observed with orosomucoid (both human and bovine), while the desialylation of bovine orosomucoid did not reduce the level of inhibition with microsomal preparation. The results of hem- agglutination-inhibition experiments for serum and microsomal preparation when fetuin was desialylated did not follow a discernable pattern, reflecting the complexity of binding interactions in this mixture of lectins. Both thyroglobulin and asialothyroglobulin were not inhibitory to the 50% level at a concen- tration of 1% or less for serum.
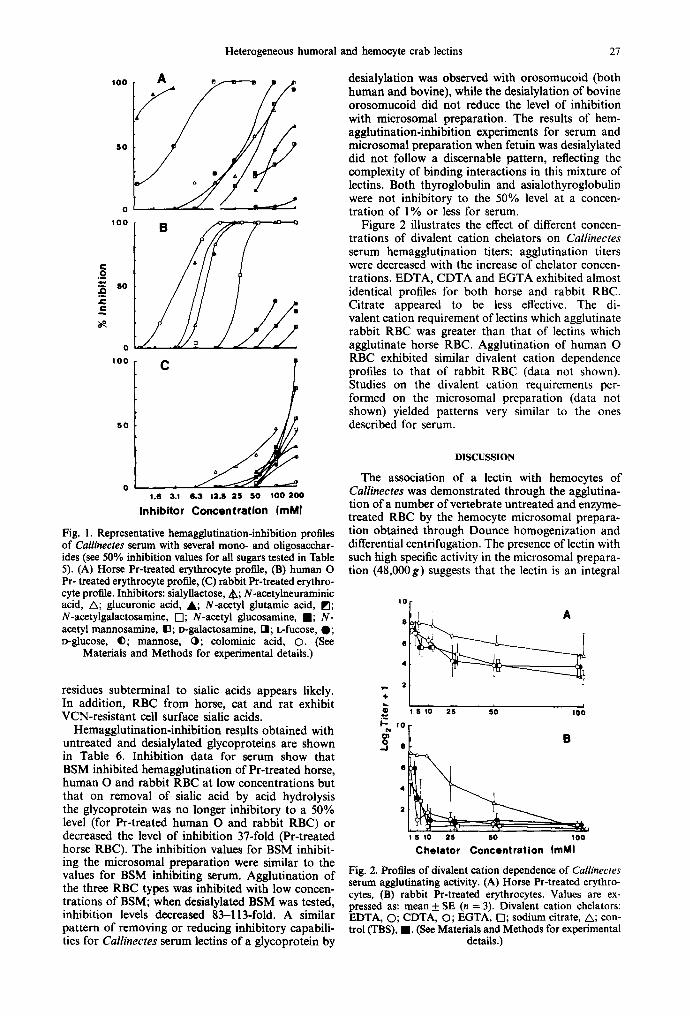
Figure 2 illustrates the effect of different concen- trations of divalent cation chelators on Callinectes serum hemagglutination titers: agglutination titers were decreased with the increase of chelator concen- trations. EDTA, CDTA and EGTA exhibited almost identical profiles for both horse and rabbit RBC. Citrate appeared to be less effective. The di- valent cation requirement of lectins which agglutinate rabbit RBC was greater than that of lectins which agglutinate horse RBC. Agglutination of human O RBC exhibited similar divalent cation dependence profiles to that of rabbit RBC (data not shown). Studies on the divalent cation requirements per- formed on the microsomal preparation (data not shown) yielded patterns very similar to the ones described for serum.
D I S C U S S I O N
The association of a lectin with hemocytes of Callinectes was demonstrated through the agglutina- tion of a number of vertebrate untreated and enzyme- treated RBC by the hemocyte microsomal prepara- tion obtained through Dounce homogenization and differential centrifugation. The presence of lectin with such high specific activity in the microsomal prepara- tion (48,000g) suggests that the lectin is an integral
I 0
8
e
4
2
I-- 10
1 A
, i 100
---?g, ~ I 5 10 25 50 100
Chelator Concentration ImMl
Fig. 2. Profiles of divalent cation dependence of Callinectes serum agglutinating activity. (A) Horse Pr-treated erythro- cytes, (B) rabbit Pr-treated erythrocytes. Values are ex- pressed as" mean +_ SE (n = 3). Divalent cation chelators: EDTA, O; CDTA, Q; EGTA, Q; sodium citrate, A; con- trol O'BS), I . (See Materials and Methods for experimental
details.)
28 FREDERICK J. CASSELS et al.
Table 5. 50% inhibition profiles of Callinectes sapidus lectins from serum and microsomal preparation by mono- and oligosaccharides ~.2
Serum .... Microsornal Preparation
Inhibitor Horse 3 Human O Rabbit Horse Human O Rabbit
Sialynactose 0.5 mM 6.0 NI 5 2.3 2.2 0.1
Colominic Acid e 2.6 5.8 NI 1.0 3.6 7.3
N-AcMuramic Acid 2.8 NI N1 3.8 l.g 9.0
N-AcGlutamic Acid 3.1 3.# NI 2.8 3. i 6.2
N-AcNeuraminic Acid It3.2 9.5 NI 12. I 3.8 15.3
N-AcGiucmamine 61.8 24.0 130.5 3.9 2.0 19.5
N-AcMannmamlne 69.5 19.5 I~t0.0 NI 1.3 7.4
N-Ac'Galacto~mmine 76.0 24.0 NI 12.0 l 1.2 32.2
Glucuronic Acid 1 lit.0 NI NI 50.0 27.2 NI
Galactosamine 192.0 NI 185.0 45.5 25.0 12.5
NeuAc 13-methyl glycoside NI NI NI 10.5 2.6 2.8
NeuGc NI NI NI 18.8 2.6 NI
Galacturonic Acid NI NI NI 23.7 36.6 NI
~Figures are the minimal concentration of sugars required to inhibit 50% of the agglutinating activity (128 agglutination units tested).
2Mono- and oligosaccharides not inhibitory to 50% level of inhibition at 200 mM, for serum: o-galactose, L-fucose, glucose, mannose, rhamnose, ketodeoxyoctonate, methoxyneuraminic acid, muramic acid, arabinogalactan; for microsomal preparation L-fucose, glucose, mannose, rhamnose, ketodeoxy- octonate, methoxyneuraminic acid, muramic acid (D-galactose and arabinogalactan not done).
3All cells pronase-treated. (Molarity expressed as free N-acetylneuraminic acid. 5NI-50% inhibition was not obtained at concentration up to 200 mM.
plasma membrane protein, though further studies will give more definite conclusions as to specific membrane localization. Complex patterns of aggluti- nation were found when serum and microsomal preparation were tested with a panel of enzyme treated and untreated RBC. This suggested a multi- plicity of lectins present which was confirmed with serum on fractionation by crossed absorption. The multiple serum lectin specificities exhibited by Calli- nectes has been noted for other crustaceans such as Panulirus interruptus (Tyler and Metz, 1945), Homarus americanus (Hall and Rowlands, 1974a, b), Macrobrachium rosenbergii (Vasta et al., 1983), and Birgus latro (Vasta and Cohen, 1984), and for other arthropods, such as the scorpions and spiders, and appears to be the rule rather than the exception (Vasta and Marchalonis, 1983).
Our results show that Callinectes lectins exhibit divalent cation dependence. Of the four chelators
tested, EDTA, CDTA and EGTA were approxi- mately equal in effectiveness at reducing agglutin- ation titers and citrate was the least effective. Pauley (1974) found that sodium citrate had no effect on the activity of Callinectes sapidus serum lectin and con- cluded that this activity did not depend on divalent cations. Other crustaceans, such as the lobsters Homarus americanus (Hall and Rowlands, 1974a) and Panulirus argus (Weinheimer, 1970) and the crab Cancer antennarius (Ravindranath et al., 1985), have lectins dependent on calcium for activity.
By hemagglutination-inhibition tests with mono- and oligosaccharides and glycoproteins it is evident that a number of N-acylamino compounds and the glycoproteins and polysaccharides that contain them inhibit effectively at low concentrations of Callinectes serum lectin fractions. The sialic acid specific lectin fraction in Callinectes serum was initially shown by the agglutinating activity toward dog and mouse
Table 6. 50% Inhibition profiles of Callinectes sapidus leetins from serum and microsomal preparation by glycoproteins ~' 2
Serum Microsornal Preparation
Inhibitor Horse 3 Human O Rabbit Horse Human O Rabbit
BSM .022 .102 .150 .006 .006 .012
Orosomucoid (hum.) .026 .081 .125 .125 .0/~6 .075
PSM .102 ,203 .500 .19[ .023 .104
Orosomucoid (boy.) .155 .087 .500 .700 .350 .650
Asialff-fetuin .200 NI ~ NI .750 NI NI
Astalo~BSM .SIS NI NI .720 .800 1.000
Asialo~or osomucoid (boy.) NI .177 .570 .450 °350 .500
Fetuin NI .605 NI .550 .230 NI
Ovomucoid NI NI NI Nl .208 NI
JFigures are the minimal concentration (% w/v) of glyeoprotein required to inhibit 50% of the agglutinating activity (128 agglutination units tested).
2Glycoproteins not inhibitory to 50% level of inhibition at 1% for serum: thyroglobulin, asialo-thyro- globulin; for microsomal preparation: none (thyroglobulin, asialo-thyroglobulin and asialo- orosomucoid (hum.) not done)
3All cells pronase-treated. 4NI-50% inhibition was not obtained at concentrations up to 1% (wt/vol).

Heterogeneous humoral
untreated and Pr-treated RBC and the lack of activity after VCN treatment of those RBC, and was con- firmed in the hemagglutination-inhibition tests. These sialic acid specificities are similar to the other charac- terized crustacean lectins including Homarus ameri- canus (Hall and Rowlands, 1974b), Macrobrachium rosenbergii (Vasta et al., 1983), Cancer antennarius (Ravindranath et aL, 1985), and Birgus latro (Vasta and Cohen, 1984). The specificity for sialic acids of at least one serum fraction extends to other arthro- pods including merostomes (horseshoe crabs), arach- nids (scorpions and spiders) and insects (Vasta and Marchalonis, 1983). Sialic acids do not appear to be present within the arthropods as self components (Warren, 1963), suggesting that the lectin fraction reactive to sialic acids may function in nonself recog- nition. This function might not only be accomplished through the binding of lectin to sialic acids or colo- minic acid but to other carbohydrates present on the bacterial cell wall or capsule. These carbohydrates include N-acetylmuramic acid, N-acetylglucosamine, glutamic acid, glucuronic acid, 2-keto-3-deoxy- octonate, D-galactose and mannose (Pistole, 1981; Vasta and Marchalonis, 1983). Inhibition profiles on Callinectes serum and microsomal preparation lectins indicate that Callinectes lectins will bind to N-acetyl muramic acid, N-acetylglucosamine and glucuronic acid in addition to NeuAc and colominic acid. This ability to bind to carbohydrate moieties present on microorganisms further suggests that the lectins function in non-self recognition.
Crustaceans are susceptible to a number of viral, bacterial and protozoan infections (Provenzano, 1983; Sindermann, 1977) with Callinectes the subject of numerous reports. Of these many pathogens, information on the surface carbohydrate present exists only for the lethal pathogenic bacterium Vibrio parahemolyticus. This bacterium is also responsible for about half of the cases of seafood-derived human bacterial food poisoning in Japan. At least 121 serotypes are recognised (Miwatani and Takeda, 1976) with the K antigens containing (of 46 serotypes tested) glucose, galactose, mannose, fucose, rhamnose, ribose, and glucosamine (23 of 46 strains) and sialic acid (4/46), Kudoh (1969). The O antigen of V. parahemolyticus appears to contain ~-N-acetyl-D- galactosaminyl, D-galactosyl, D-glucosyl, D-glucos- aminyl, N-acetyl-D-glucosaminyl and ~-N-acetyl-D- galactosaminyl residues in antigenic positions (Torii and Sakakibara, 1972). The Callinectes lectin frac- tions as described would potentially agglutinate a number of serotypes of V. parahemolyticus, but more definitive conclusions as to the role of Callinectes lectin fractions in self defense must await more information as to the serotypes pathogenic to Calli- nectes and to the ability of the lectin fractions to agglutinate V. parahemolyticus and other pathogens of Callinectes. Continuing work on the purification of Callinectes sapidus serum and hemocyte membrane lectins and the elucidation of their physicochemical properties and relationship to the phagocytic process is now in progress.
Acknowledgements--We would like to thank Mr. Charles Farmer and Mr. David Whittaker of the South Carolina Wildlife and Marine Resources Department for collecting
and hemocyte crab lectins 29
animals, Mrs. Teresa Vasta for the preparation of figures and Miss Lesley Seeker for typing the manuscript.
REFERENCES
Amirante G. A. and Basso V. (1984) Analytical study of lectins in Squilla mantis L. (Crustacea, Stomatopoda) using monoclonal antibodies. Devl. Comp. Immunol. 6, 721-726.
Amirante G. A. and Mazzalai F. G. (1978) Synthesis and localization of hemoaggiutinins in hemocytes of the cock- roach Leucophaea maderae L. Devl. Comp. lmmunol. 2, 723-740.
Brown R., Almodavar L. R., Bathia H. M. and Boyd W. C. (1968) Blood group specific agglutinins in invertebrates. J. Immunol. 100, 214-216.
Cohen E. and Vasta G. R. (1982) Immunological signifi- cance of ubiquitous lectins. In Developmental Immun- ology: Clinical Problems and Aging (Edited by Cooper E. L. and Brazier M. A. B.), pp. 99-108. Academic Press, New York.
Cornick J. W. and Stewart J. E. (1973) Partial characteriz- ation of a natural agglutinin in the hemolymph of the lobster, Homarus americanus. J. Invert. Pathol. 21, 255- 262.
Gold E. R. and Balding P. (1975) Receptor-Specific Proteins, Plant and Animal Lectins. Excerpta Medica, Amsterdam.
Hall J. L. and Rowlands D. T. (1974a) Heterogeneity of lobster agglutinins. I. Purification and physicochemical characterization. Biochemistry 13, 821-827.
Hall J. L. and Rowlands D. T. (1974b) Heterogeneity of lobster agglutinins. II. Specificity of agglutinin-erythro- cyte binding. Biochemistry 13, 828-832.
Kudoh Y. (1969) Studies on K antigen of Vibrio para- hemolyticus. 2. Immunochemical specificity of K antigens and their sugar constituents. Jap. J. Bacteriol. 24, 331-337 (Japanese with English summary).
McCumber L. J. and Clem L. W. (1977) Recognition of viruses and xenogeneic proteins by the blue crab, Calli- nectes sapidus. I. Clearance and organ concentration. Devl. Comp. Immunol. 1, 5-14.
McCumber L. J., Hoffmann and Clem L. W. (1979) Recog- nRion of viruses and xenogeneic proteins by the blue crab, Callinectes sapidus: a humoral receptor for T 2 bacterio- phage. J. lnvert. Path. 33, 1-9.
Miwatani T. and Takeda Y. (1976) Vibrio parahaemolyticus a Causative Bacterium of Food Poisoning. Saikon, Tokyo.
Noguchi H. (1903) A study of immunization-hemolysins, agglutinins, precipitins and coagulins in cold blooded animals. Zwt. F. Bact. Parasitenk. Inf. 33, 353-360.
Pauley G. B. (1974) Comparison of a natural agglutinin in the hemolymph of the blue crab, Callinectes sapidus, with agglutinins of other vertebrates. In Contemporary Topics in Immunology (Edited by Cooper E. L.), Vol. 4, pp. 241-260. Plenum Press, New York.
Pistole T. G. (1981) Interaction of bacteria and fungi with lectins and lectin-like substances. Ann. Rev. Microbiol. 35, 85-112.
Prokop O., Rackwitz A. and Schlesinger D. (1965) A "new" human blood group receptor Ahd tested with saline extracts from Helix hortensis (garden snail). J. Forensic Med. 12, 108-110.
Provenzano A. J. (1983) The Biology of Crustacea, Vol. 6, Pathobiology. Academic Press, New York.
Ravindranath M., Higa H., Cooper E. and Paulson J. (1985) Purification and characterization of an O-acetylsialic acid-specific lectin from a marine crab Cancer antenna- rius. J. biol. Chem. 260, 8850-8856.
Renwrantz L. and Stahmer A. (1983) Opsonizing properties of an isolated hemolymph agglutinin and demonstration of lectin-like recognition molecules at the surface of

30 FREDERICK J. CASSELS et al.
hemocytes from Mytilus edulis. J. Comp. Physiol. 149, 535-546.
Sindermann C. J. (1977) Disease Diagnosis and Control in North American Marine Aquaculture. Elsevier, New York.
S6derhiill K. and Smith V. J, (1983) Separation of the haemocyte populations of Carcinus rnaenas and other marine decapods and prophenoloxidase distribution. DevL Cornp. Irnrnunol. 7, 229-239.
Torii M. and Sakakibara K. (1972) Immunochemical studies on O-antigens of Vibrio parahernolyticus. III. Studies on inhibition of the precipitation reaction by known sugars. Biken J. 15, 17-22.
Tyler A. and Metz C. B. (1945) Natural heteroagglutinins in the serum of the spiny lobster, Panulirus interruptus. I. Taxonomical range of activity, electrophoretic and immunizing properties. J. exp. ZooL 100, 387-406.
Tyson C. J. and Jenkin C. R. (1974) Phagocytosis of bacteria in vitro by haemocytes from the crayfish (Para- chaeraps bicarinatus). Aust. J. Exp. Biol. Med. Sci. 52, 341-348.
Uhlenbruck G., Rothe A. and Pardoe G. I. (1968) Bemerkenswerte chemische, serologische un elektro- kinetische phanomene bei katzenerythrozyten: Zugleich ein beitrag zur bestimmung und bedeuntung von mucoid- und glykolipoidgebundener erythrozytenneuraminsafire. Z. Imrnun. Forsch. 1.36, 79-97.
Vasta G. R., Sullivan J. T., Cheng T. C., Marchalonis J. J. and Warr G. W. (1982) A cell membrane-associated lectin of the oyster hemocyte. J. Invert. Path. 40, 367-377.
Vasta G. R. and Cassels F. J. (1983) N-acylaminosugar- specific lectins in the serum and hemocyte microsomal fraction from the blue crab Callinectes sapidus. Am. ZooL 23, 901.
Vasta G. R. and Marchalonis J. J. (1983) Humoral recognition factors in the Arthropoda. The specificity of Chelicerata serum lectins. Am. ZooL 23, 157-171.
Vasta G. R., Warr G. W. and Marchalonis J. J. (1983) Serological characterization of humoral lectins from the freshwater prawn Macrobrachium rosenbergii. Devl. Comp. InnnunoL 7, 13-20.
Vasta G. R., Cheng T. C. and Marchalonis J. J. (1984) A lectin on the hemocyte membrane of the oyster (Crasso- strea virginica). Cell. InununoL 88, 475-488.
Vasta G. R. and Cohen E. (1984) Carbohydrate specificities of Birgus latro (coconut crab) serum lectins. Devl. Cornp. ImmunoL 8, 197-202.
Warren L. (1963) The distribution of sialic adds in nature. Cornp. Biochern. Physiol. 10, 153-171.
Weinheimer P. F. (1970) Characterization of erythrocyte factors of Panulirus argus. A contribution to immuno- phylogeny. Ph.D. Diss., University of Alabama, Birming- ham.





























