Embed Size (px)
Citation preview

23 Production of Genetically Identical
Embryos by Electrofusion
L a w r e n c e Char les S m i t h
Centre de Recherche en Reproduction Animale, Faculté de Médecine Vétérinaire, Université de Montréal, Saint Hyacinthe, Québec J2S 7C6, Canada
I. Introduction
II. The Amphibian Cloning Model
A. Methodology
Β. Development Potential
III. The Mammalian Cloning Model
A. Methodology
B. Development Potential
IV. Nuclear-Cytoplasmic Interactions
A. Cell Cycle Stage Effects
B. Cytoplasmic Effects
V. Applications and Conclusions
References
I. Introduction
Recent improvements in micromanipulation techniques have opened new possibil-
ities for experimental research in mammalian embryology. The combined use of the
techniques of nuclear transplantation and cell electrofusion have enabled further
investigations into the genetic and epigenetic mechanisms by which early embryonic
development is controlled. These studies have clarified our understanding of deter-
mination and differentiation in mammalian embryonic cells during the preimplan-
tation stages and also of the abilities of oocyte cytoplasm to reprogram these de-
velopmental events. Live offspring have been derived from nuclear transplanted
Guide to Electroporation and Electrofusion
Copyright © 1992 by Academic Press, Inc. All rights of reproduction in any form reserved. 371

372 Lawrence Charles Smith
embryos in laboratory and domestic species with the purpose of producing genetically
identical animals ("clones").
This review aims to provide readers with some background knowledge on factors
influencing the development of nuclear transplanted embryos, hoping that this will
stimulate further research on nucleocytoplasmic interactions in early embryos. The
first section is devoted to the cloning model used for amphibia eggs, since this has
been studied in more detail and for a longer t ime than the mammalian model.
Although nuclear transplantation techniques in mammals have been around for at
least one decade, cloning procedures have only become viable in the last 5 years
when they were first successfully applied to farm animals (Willadsen, 1986). The
mammalian section begins by describing the steps leading to the current method-
ology and then outlines the effects the donor nucleus and the recipient cytoplasm
have on the ability of nuclear transplanted embryos to develop normally. The section
on the nuclear-cytoplasmic interactions discusses more fully the events taking place
at fusion, such as the effects of cell cycle stage, cytoskeleton interactions, and others.
The final section concludes by detailing some practical uses for nuclear transplan-
tation and the production of genetically identical mammals .
II. The Amphibian Cloning Model
A. Methodo logy
Current procedures for cloning mammalian embryos originated from nuclear trans-
plantation experiments in amphibians performed initially by Briggs and King (1952).
The transplantation operation in amphibia is carried out in two main steps. First,
the recipient frog eggs are activated by pricking with a glass needle; this causes
rotation and brings the animal pole to lie uppermost . The chromosomes are visualized
as a "black dot" ( i .e . , p igment granules clustered about the second metaphase meiotic
spindle) that can be extirpated surgically by lifting the spindle away from the surface
of the egg with a glass needle (Briggs and King , 1953). Effective enucleation can
also be attained by ablating the chromosomes with either ultraviolet (Gurdon, I960)
or microbeam ruby laser irradiation (Ellinger et al., 1975). The second step involves
the isolation of the donor cells and the nuclear transplantation itself. Donor cells
are isolated, either mechanically with a fine-diameter pipette or chemically with
enzymatic treatment. A given cell is then drawn into the t ip of a sharp micropipette,
the inner diameter of which is somewhat smaller than that of the cell. This causes
rupture of the plasma membrane, but the cytoplasm contents are not dispersed and
remain surrounding the nucleus. The pipet te is then inserted through the membrane
of the recipient enucleated egg and the contents of the broken cell are ejected,
liberating the nucleus into the egg cytoplasm.

Chapter 23 Production of Genetically Identical Embryos 373
B . D e v e l o p m e n t Po t en t i a l
Amphibian nuclear transplantation experiments using several embryonic cell types
have revealed that , as donor nuclei are tested from progressively older stages of
embryogenesis, there is a decrease in the number of individuals that develop normally
(reviews by King , 1966; DiBerardino and Hoffner, 1970; Gurdon, 1974; Briggs,
1977; McKinell , 1978). Al though already containing approximately ten thousand
cells with some degree of morphological differentiation, most , if not all, blastula
nuclei are thought to be able to support normal development when fused to enu-
cleated eggs. However, only a few early gastrula-stage nuclei have been shown to
be totipotent , and a very low percentage of nuclei taken at any later embryonic
stage are fully able to support development (Briggs and King, I960) . Studies carried
out using nuclei derived from regions of larvae already commited to a specific
developmental pathway have shown that only around 0 . 2 % develop to adults and
another 4 % arrest at the larva stage (pluripotent nuclei). Together, these results
suggest that the developmental capacity of nuclei becomes progressively restricted
during the process of cell type determination and differentiation.
Several investigators have tested the developmental capacity of amphibian nuclei
from adult cells. The most advanced stage of development reported from using
nuclei of adult cells has been a feeding larva obtained from spermatogonia cells of
Rana pipens (DiBerardino and Hoffner, 1971). Among adult somatic nuclei tested
in Xenopus, there are 33 cases of early larvae obtained in support of nuclear pluri-
potency (Laskey and Gurdon, 1970; Gurdon et al., 1975; McAvoy et al., 1975;
Wabl et al., 1975; Brun, 1978). One of these larvae was reported to be normal and
originated from a crest cell nucleus of the intestine, but this apparent normal larva
died during an early larval stage (McAvoy et al., 1975). The remaining nuclear
transplant larvae were morphologically abnormal and the percent success ranged
from 0.3 to 6 . 0 % . N o adult frog has yet developed from transplanted adult nucleus.
In order to determine the nature of the developmental restrictions displayed by
nuclei from advanced cell types, an extensive series of studies was performed on
abnormal embryos and larvae of Rana nuclear transplants (reviewed by DiBerardino,
1979)· Most abnormal nuclear transplants examined exhibited abnormalities in
chromosome number and/or structure that , in most cases, arose during the first cell
cycle of the egg (DiBerardino and Hoffner, 1970). These abnormalities are now
known to be the cause of developmental arrest in this species. The most severe
chromosomal alterations cause developmental arrest at the blastula stage, whereas
relatively minor karyotypic alterations permit development to early larval stages.
Evidence has been presented that most of these numerical and structural changes in
the chromosomes (1) involve chromosomal loss, (2) are a reflection of chromosomal
differentiation acquired progressively though embryogenesis, and (3) are not a result,
for the most part , of technical damage (DiBerardino, 1979, 1980; DiBerardino and

374 Lawrence Charles Smith
Hoffner, 1980). It appears that the cytoplasmic cell cycle of the amphibian egg,
which is much faster than the nuclear cycle of advanced cell types, induces premature
changes in transplanted nuclei resulting in incomplete D N A replication, chromo-
some breaks and arrangements, and hypoaneuploidy in the most severely affected
embryos.
III. The Mammalian Cloning Model
A. Methodo logy
As the volume of individual mammalian eggs is close to one thousand times smaller
than that of frog eggs, it is not surprising that more refined methods of microsurgery
were required before the development of techniques for nuclear transplantation could
be usefully applied to mammals . The successful microinjection of embryonic nuclei
into the cytoplasm of one-cell rabbit (Bromhall, 1975) and mouse embryos (Mod-
linski, 1978) suggested that nuclei from later embryonic stages were able to par-
ticipate with the egg's genome in supporting preimplantation development. The
first report of mice born from nuclear transplantation came from the work of Illmensee
and Hoppe (1981), showing that nuclei derived from the inner cell mass rather than
trophectoderm of mouse blastocysts were able to support development to the m o r u l a -
blastocyst stage in 3 4 % of the transferred embryos, and that 19% of these would
develop to term. These reports brought many scientists to speculate that this tech-
nique would provide a means of making an infinite number of genetically identical
copies from a single embryo. However, it was later reported that a t tempts of other
laboratories using this technique were unsuccessful (Marx, 1983; McGrath and
Solter, 1984a; McLaren, 1984). Suggestions were made that Illmensee and Hoppe
could not have properly enucleated the recipient zygotes, which would have allowed
for their development to full term (McGrath and Solter, 1984a).
In support of these suggestions were the results from a novel method developed
for nuclear transplantation by McGrath and Solter (1983a). Their technique avoids
the necessity of penetrating the plasma membrane of either the donor or the recipient
egg. This is possible by placing the eggs in cytoskeletal inhibitors for a short period
before microsurgery, followed by zona pellucida penetration with a sharp micro-
pipette and sucking out the two pronuclei surrounded by a piece of plasma mem-
brane—a karyoplast. The donor karyoplast, which can be obtained from any em-
bryonic or somatic cell, is injected under the zona pellucida of the enucleated
recipient egg (Fig. 1). The following step of this noninvasive method for nuclear
transplantation relies on an effective means for fusing the nuclear (karyoplast) and
cytoplasmic (cytoplast) portions enclosed in the zona pellucida. Contrary to the poor
levels of success obtained using Illmensee and Hoppe's invasive technique, McGrath
and Solter's method provided virtually 100% of success, and the yield of live mice
is very high when fertilized eggs are used as nuclear donors.

Chapter 23 Production of Genetically Identical Embryos 375
Figure 1 Technical procedure use for the enucleation of pronuclear zygotes followed by the collection and transplantation of karyoplasts. (a) Positioning of pronuclear zygote for enu-cleation; (b) removal of male and female pronuclei in a karyoplast; (c) nuclear donor four-cell stage embryo positioned for microsurgery; (d) removal of four-cell stage karyoplast; (e) positioning of donor karyoplast into zona pellucida; (f) recipient cytoplast and donor karyoplast ready for fusion.

376 Lawrence Charles Smith
Several methods are available for fusion, some of which are either unreliable and
toxic, such as polyethylene glycol (PEG) and lysolecithin or, as for inactivated Sendai
virus solutions, laborious to obtain and dangerous to raise due to its highly infectious
nature. The latter also has the inconvenience that it is impossible to ascertain exactly
the moment of fusion, since it occurs between 15 to 60 min after manipulation
according to the amount and activity of the solution injected into the perivitelline
space. However, possibly due to its high levels of success and to the ease with which
manipulations are performed, Sendai-mediated fusion has been the most widely used
approach in mouse nuclear transplantations. Willadsen (1986) reported the use of
electric pulses to fuse the nuclear donor cell to the cytoplasm of the enucleated egg.
Although Sendai virus-mediated fusion was successfully achieved in sheep, electro-
fusion techniques are more effective and less variable when species other than mice
are used.
Electrofusion has been successfully used to fuse blastomeres and for nuclear
transplantation experiments in mammalian embryos derived from species as diverse
as mice (Kubiak and Tarkowski, 1985; Tsunoda et al, 1987a; Kono and Tsunoda,
1988; Barra and Renard, 1988; Clement etal., 1988), rabbit (Ozil and Modlinski,
1986; Stice and Robl, 1988; Clement etal, 1988), sheep (Willadsen, 1986; Smith
and W i l m u t , 1989), cattle (Prather et al, 1987; Bondioli et al, 1990), and pigs
(Clement et al, 1988; Prather et al, 1989). Embryonic cells (karyoplasts and
cytoplasts) to be fused are placed in a chamber with electrofusion medium and
positioned between two electrodes connected to a pulse generator (Fig. 2). Successful
fusion can be attained using a large range of parameters for the direct current (DC)
fusing pulse. Normally, a field intensity of around 1.0 kV/cm with durations between
50 and 100 \ks is used. Two or three consecutive pulses separated by a fraction of
a second may, in some instances, improve the levels of success. Fusion seems to be
caused by the reversible electrical breakdown in the zone of membrane contact
between the cytoplasm and nuclear donor cells. This breakdown is followed im-
mediately by their repair into a single membrane, leading initially to the formation
of small pores between the two cells, which continue growing and joining together
until fusion is complete (Zimmerman and Vienken, 1982). Alternating current (AC)
pulses may be used to align the cells so as to position their membranes perpendicular
to the electrical field where conditions for fusion are most suitable. The preceding
AC pulse is particularly important when fusing enucleated oocytes to cells with
reduced diameters since the polarization caused by the AC field will aid in bringing
their membranes into contact for the DC fusing pulse (Smith and W i l m u t , 1989).
Willadsen (1986) reported the development to term of sheep embryos derived
from the transplantation of embryonic blastomere nuclei to the cytoplasm of sec-
ondary oocytes. The relevance of his findings was not exclusively related to the
ability of a single eight-cell blastomere to support development, since this had
already been indicated in earlier chimeric studies (Willadsen and Fehilly, 1983),
but also that , as for amphibian nuclear transfers, the resulting fused embryo would

Chapter 23 Production of Genetically Identical Embryos 377
Figure 2 Schematic diagram of a chamber used for the electrofusion of mammalian embryos. Two ΙΟΟ-μπι-diameter platinum electrodes are stuck to the bottom of a glass petri dish separated at 200 μπι from each other.
develop and differentiate as if commencing from the t ime of fertilization, indicating
an ability of the oocyte's cytoplasm for reprogramming the developmental "clock"
of the donor nucleus. Al though the technique used by Willadsen is similar to that
used with frogs, in that both techniques utilize the secondary oocytes as the recipient
cytoplasm, it also uses the noninvasive technique for transplanting the nucleus into
the cytoplasm.

378 Lawrence Charles Smith
B. D e v e l o p m e n t Po ten t i a l
As for amphibian, many factors are involved in the ability of mammalian nuclear
transplanted embryos to develop normally. Because these techniques are still fairly
recent in mammals , our understanding of each factor remains l imited. The following
sections will try to point out some possible reasons why many embryos fail to develop
after nuclear transplantation.
1. Nuclear Donor Cell
All cells in an embryo have a nucleus containing a comparable amount of D N A
{In), which is derived by continuous replication and segregation at mitosis from a
single fusion product of the D N A carried by male (n) and female (n) gametes at
fertilization. Assuming that this concept of nuclear equivalence is correct, and there
are many reasons to believe so, one can presume that because later embryonic stages
possess more cells with genetically identical nuclei, these are more suitable for
cloning. There are some factors that l imit the extent to which embryos should
develop when allocated for use as nuclear donors in the cloning procedure.
The first factor relates to the loss of developmental potential at the expense of
cellular determination or differentiation. As described earlier in amphibia, nuclei
derived from cells that have commit ted themselves to a particular pathway seem to
lose the ability to be readily reprogrammed and are unable to regain the totipotent
status of their ancestral fertilization genome. An early report in mice showed that ,
although 9 5 % of enucleated zygotes fused with zygote pronuclei will develop to
blastocyst, only 13% will develop to blastocyst when 2-cell nuclei are fused and
none when four-, eight-, 16-cell, and inner cell mass (ICM) nuclei are fused to
enucleated zygotes, indicating a rapid loss in potential already by the 2-cell stage
(McGrath and Solter, 1984a). These findings have been confirmed by other labo-
ratories (Robl et al., 1986; Surani et al., 1987; Tsunoda et al., 1987b; Smith et al.,
1988), and suggestions have been made that blastomere nuclei lose their potential
at the t ime of maternal-zygotic transition (MZT) due either to genome differentiation
or to toxicity of stage-specific factors (Solter et al., 1986; Solter, 1987).
More recent findings in several mammalian species have shown that nuclei derived
from stages beyond the M Z T are still able to support development (Table 1).
Comparisons between results from several mammals indicate that embryos derived
from species that experience M Z T within a single or two cells after oocyte activation
tend to perform poorly after nuclear transplantation. In mice, where M Z T occurs
at the early two-cell stage (Flach et al., 1982), it has been reported that a small
proportion of nuclei from the eight-cell (McGrath and Solter, 1986) and from the
inner cell mass (Tsunoda et al., 1988, 1990) will support development to the
blastocyst stage when fused to enucleated secondary oocytes. Again in mice, by
using karyoplasts derived from embryos at different stages within the two-cell cycle,

Chapter 23 Production of Genetically Identical Embryos 379
Table 1
Pre- and Postimplantation Development Potential of Nuclear Transplanted Embryos Derived from the Fusion of Embryonic Cells to the Cytoplasm of "Enucleated" Secondary
Oocytes in Several Mammalian Species*
Morula Embryonic cell Number or Number Established Full-term
Species type fused blastocyst transferred pregnancy development Reference
Mouse 2-Cell 86 35% n.a. n.a. n.a. (1) 8-Cell 36 None n.a. n.a. n.a. (1) ICM 80 3% n.a. n.a. n.a. (1) ICM 83 4% n.a. n.a. n.a. (2) PGC 135 10% 59 10% None (1)
Pig 2-Cell 11 9% 33 n.a. None (3) 4-Cell 83 8% 34 n.a. 3% (3) 8-Cell 57 19% 21 n.a. None (3)
Rabbit 8-Cell 70 68% 85 2% 1% (4) 16-Cell n.a. n.a. 110 n.a. 21% (5) 32-Cell 67 76% n.a. n.a. n.a. (5) 3 2-Cell 67 55% 67 n.a. n.a. (6) ICM 52 37% n.a. n.a. n.a. (6) TE 27 None n.a. n.a. n.a. (6)
Sheep 8-Cell 76% 42% 4 n.a. 75% (7) 16-Cell 29 48% 6 50% n.a. (7) 16-Cell 49 35% 14 21% 14% (8) ICM 16 56% 8 13% 13% (8)
Cattle <8-Cell 111 12% 12 n.a. None (9) < 16-Cell 50 16% 7 n.a. 29% (9) <32-Cell 24 8% n.a. n.a. n.a. (9) Day 5 morula 604 32% n.a. n.a. n.a. (10) Day 5.5 morula 139 40% n.a. n.a. n.a. (10) Day 6 morula 54 53% n.a. n.a. n.a. (10) Morula n.a. n.a. 463 23% 22% (10)
'References: (1) Tsunoda et al., 1990; (2) Tsunoda et al., 1988; (3) Prather et al., 1989; (4) Tsunoda et al., 1989; (5) Collas and Robl, 1990; (6) Collas and Robl, 1991; (7) Willadsen, 1986; (8) Smith and Wilmut, 1989; (9) Prather etal., 1987; (10) Bondioli et al., 1990; n.a. = not available; ICM = inner cell mass; PGC = primordial germ cell; TE = trophectoderm cells.
nuclei derived from embryos just before the M Z T are slightly less able to support
blastocyst development than those derived from embryos after the M Z T (Smith et
al., 1988). In pigs, where M Z T occurs at the four-cell stage (Norberg, 1973), no
significant difference could be detected between using nuclei from two- (9%), four-
(8%) or eight-cell (19%) blastomeres in nuclear transplantations to enucleated sec-
ondary oocytes (Prather et al., 1989).

380 Lawrence Charles Smith
Species that experience M Z T at or after the eight-cell stage (three or more cell
cycles after oocyte activation) tend to perform better in nuclear transplantation trials.
In sheep, where M Z T occurs at the eight-cell stage (Crosby et al., 1988), it has
been reported that eight-cell (42%) and 16-cell (48%) nuclei are able to support
development to blastocyst at similar proportions (Willadsen, 1986) and also that
nuclei from 16-cell (35%) and the ICM (56%) stages of early blastocysts are similarly
competent in supporting development both to blastocyst and to term (Smith and
Wi lmu t , 1989). In cattle, where M Z T occurs around the eight-cell stage (Camous
et al., 1986), reports have indicated similar development potentials are obtained
from nuclei derived from embryos collected at day 5 (32%), day 5.5 (40%), and
day 6 (53%) (Bondioli et al., 1990). Therefore, it seems that , at least up to the
embryonic stages tested in farm animals, there is no detectable loss of developmental
potential of embryonic nuclei dur ing the preimplantat ion stages utilized to this
data. However, reports using rabbit nuclear transplanted embryos have indicated a
slight decrease in potential at later embryonic stages. Al though M Z T occurs at the
eight-cell stage in rabbits (Manes, 1977), the potential of nuclei derived from eight-
cell (68%), 32-cell (55%), or from the inner cell mass cells (37%) seems to decrease
slightly during development (Tsunoda et al., 1989; Collas and Robl, 1990, 1991).
Another important finding made in this species is the striking fall in potential (0%)
when using commit ted blastocyst trophectoderm cells in nuclear transplantations
(Collas and Robl, 1991).
Technical aspects should also be considered when deciding on the best embryonic
stage from which to derive viable cells for nuclear transplantation. At compaction,
cells from the outer layer of the embryo develop t ight junctions making disaggre-
gation into single cells increasingly more difficult. Al though calcium-free medium
and/or trypsinization can be used to aid in separating cells, such treatments may be
detrimental to the viability of their nuclei even after short periods of exposure. The
compact outer layers of late morula and blastocysts can be destroyed through an
immunosurgical technique (Solter and Knowles, 1975), permit t ing an easier dis-
aggregation of their inner cells as reported for the use of cells from the inner cell
mass of blastocysts in sheep nuclear transplantations (Smith and W i l m u t , 1989).
Moreover, cell diameter decreases considerably during the cleavage stages, and their
reduced size causes problems during the fusion procedure.
2. Recipient Cytoplasm
Recipient cytoplasm for mammalian nuclear transplantation was derived initially
from enucleated zygotes at early stages after fertilization (McGrath and Solter,
1983a). As described in the previous section, nuclear transplantation experiments
in mice using cytoplasm from pronuclear zygotes provided very limited support
from nuclei beyond the two-cell stage in mice (McGrath and Solter, 1984a). Nuclear
transplantations to zygotic cytoplasm have also been at tempted in other mammals

Chapter 23 Production of Genetically Identical Embryos 381
with very poor results. In rats, al though 2 2 % of the embryos derived from pronuclear
transfers developed to term, no pregnancies could be attained when using nuclei
derived from two-, four-, or eight-cell blastomeres (Kono et al, 1988). In cattle,
pronuclear zygotes were centrifuged to reveal the pronuclei for removal and later
fused to karyoplasts from early stage blastomeres (Robl et al., 1987). Bovine results
were similarly discouraging, with only 17% of the pronuclear transplantations de-
veloping to the morula-blastocyst stage after 5 days culture in a sheep's oviduct and
not a single normal development after transplantations of two-, four-, and eight-
cell blastomere nuclei.
When using cytoplasm from two-cell enucleated blastomeres, Robl et al. (1986)
were able to obtain morulae and blastocysts after fusion with karyoplasts derived
from eight-cell mouse blastomeres. However, they also reported that these apparently
normal blastocysts were unable to support development beyond mid-gestation after
transfer to synchronized females. W i t h a slight modification of the technique for
using two-cell recipient cytoplasm, Tsunoda et al. (1987) were able to produce a
small number of offspring from nuclei derived from eight-cell embryos but concluded
that this technique would not prove useful in cloning mammalian embryos. Gen-
erally, recipient cytoplasm from secondary oocytes utilized in mouse nuclear trans-
plantations has provided very little improvement when compared to the postfertil-
ization cytoplasm sources (Fig. 3). However, some development to blastocyst was
achieved with eight-cell (5%) and ICM (3%) stages after fusion to enucleated oocytes
(McGrath and Solter, 1986; Tsunoda et al., 1990). Moreover, primordial germ cell
(PGC) nuclei have also been reported to support development to blastocyst (10%)
and to mid-gestation (10%) after fusion to secondary oocyte cytoplasm (Tsunoda et
al, 1990).
Secondary oocytes have been the only cytoplasm recipient stage to support full-
term development after nuclear transplantation in mammals . Liveborn have been
produced in several species including sheep (Willadsen, 1986; Smith and W i l m u t ,
1989), cattle (Prather etal, 1987; Marx, 1988; Bondioli etal, 1990), rabbit (Stice
and Robl, 1988), and pigs (Prather et al, 1989). The optimal stage for util izing
secondary oocytes for nuclear transplantations ( t iming after ovulation or maturation)
has not yet been determined, but there seems to be an increased potential after a
short period of aging (Ware et al, 1989). However, this beneficial effect is likely
to be related to improved levels of success in causing activation by artificial stimuli
after aging. The beneficial effect of aging the oocyte has also been observed in rabbit ,
where further improvement can be obtained by increasing the number of pulses at
or after electrofusion (Collas and Robl , 1990).
In general, secondary oocytes have been obtained directly from the oviduct either
during surgical intervention or at slaughter. W i t h the current efficiency of the
cloning procedure, the collection of in vivo matured oocytes comprises a substantial
proportion of the total cost for obtaining a nuclear transplanted offspring. Al though
Prather et al (1987) reported l imited development using in vitro matured oocytes,

382 Lawrence Charles Smith
Figure 3 Microsurgical methods used in the preparation of sheep secondary oocytes and embryos for nuclear transplantation by electrofusion. (a) Secondary oocyte positioned for enucleation with the first polar body (1PB) facing the beveled microsurgical pipette, (b) Removal of approximately one-quarter of the cytoplasm surrounding the 1PB in a membrane-bound cell fragment. A single cell derived from either (c) 16- or 32-cell embryos or (d) inner cell mass of early blastocysts has been positioned within the zona pellucida of an enucleated secondary oocyte for the electrofusion procedure.
recent improvements in the methods for in vitro maturat ion (Gordon and Lu, 1990)
are likely to make this the most cost-effective route for obtaining recipient oocytes
for nuclear transplantation. This slaughterhouse source of cytoplasm has been suc-
cessfully used for obtaining live offspring in cattle and sheep nuclear transplantation.
One limitation for the use of secondary oocytes as recipient cytoplasm concerns
the enucleation procedure. W i t h the exception of the rabbit (Stice and Robl, 1988),
metaphase chromosomes cannot be readily visualized in secondary oocytes from other
farm species due to the presence of large lipid vesicles in the cytoplasm. This leads
to the need for using the position of the first polar body as an indicator of the
position of the chromosomal plate followed by the aspiration of cytoplasm from the
adjacent area (Fig. 4). However, possibly due to displacement or degeneration of

Chapter 23 Production of Genetically Identical Embryos 383
1 0 0 T
1 -ce l l 2 -ce l l 4 -ce l l 8 -ce l l I.C.M.
Nuclear donor s t age
Figure 4 Comparison between the development potential of different recipient cytoplasm
stages with regard to their ability in supporting the development to the morula or blastocyst
stage after fusion to karyoplasts derived from preimplantation stage embryos. MZT = timing
of the maternal-zygotic transition. O, 1-cell cytoplasm; · , 2-cell cytoplasm; Δ, oocyte
cytoplasm.
the first polar body, an improper removal of chromosomes still occurs in a substantial
proportion of manipulated secondary oocytes. Unsuccessful enucleation has been
reported to occur in approximately one-third of the a t tempts , which, possibly due
to aneuploidy, lead to abnormal cleavage and developmental arrest during the early
preimplantation stage (Willadsen, 1986; Smith and W i l m u t , 1989). Al though the
success of enucleation can be ascertained using D N A vital stains, further research
is required to assess the effect this may have on development due to the possible
damaging of mitochondrial D N A and other cytoplasmic components.
IV. IMuclear-Cytoplasmic Interactions
When fusion is complete, donor nucleus and recipient cytoplasm will initiate in-
teractions, which, when compatible, will lead to the normal development of a nuclear
transplanted embryo. This section discusses the cellular and molecular interactions
that take place at and after fusion.

384 Lawrence Charles Smith
A. Cell Cycle Stage Effects
Experiments with mouse pronuclear zygotes have indicated that cell cycle stage
synchrony between nucleus and cytoplasm is beneficial for further development in
vitro (Smith et al., 1988). This observation has been further extended to transplan-
tations between two-cell embryos where asynchronous exchanges were highly del-
eterious to further development (unpublished observation). This effect may be ex-
plained either by the disruption of the cell cycle oscillatory mechanisms or by
incompatible nuclear-cytoplasmic interactions in controlling critical developmental
steps (Smith et al., 1990). Possibly resulting from incompatible nuclear-cytoplasmic
interactions, sheep embryos recovered from ligated oviducts 5 days after nuclear
transplantation either do not cleave at all (and may fragment after a period of aging)
or cleave a few times but cannot compact and blastulate as normal embryos (Smith
and W i l m u t , 1989). Because observations have been performed mostly after a long
period of in vivo culture, it is unclear exactly at what stage these embryos arrest
development. Better methods for culturing livestock embryos in vitro ( i .e . , coculture
with oviductal cells) coupled with detailed biochemical studies on their transcrip-
tional and translational activities will certainly enable a clearer understanding of
these nuclear-cytoplasmic incompatibilit ies.
B . Cytop lasmic Effects
Nucleo—cytoplasmic compatibili ty has been shown to improve when nuclear trans-
planted embryos are placed into a medium containing cytochalasin Β (CB) for a
short period immediately after fusion. In sheep, nuclei derived from both 16-cell
stages ( 3 5 % vs. 11%) and ICM cells (56% vs. 0%) of early blastocysts were sig-
nificantly better able to support development to morulae and blastocysts after nuclear
transplantation when placed in medium containing 7.5 p g CB for 1 hr after the
electric stimulus as compared with placing them into medium alone (Smith and
W i l m u t , 1989). A similar t reatment was also effective in improving development
to blastocyst (44% vs. 11%) in rabbit 32-cell nuclear transplantations (Collas and
Robl, 1990). Together, these results indicate that fused nuclei are affected by
cytoskeletal mechanisms operating during the activation of the recipient cytoplasm,
possibly leading to chromosomal anomalies by extrusion into pseudo first polar
bodies. However, an abstract report has shown that the effect of cytochalasin Β is
not apparent in cattle nuclear transplantation, which may indicate species variations
in nucleo-cytoplasmic interactions after fusion (Levanduski and Westhusin , 1990).
Czolowska et al. ( 1984) observed the swelling of thymocyte nuclei in activated
ovum cytoplasm to equal that of pronuclei when nuclear introduction coincided
with activation, but diminished with increasing t ime between activation and nuclear
introduction. Nuclear swelling has also been observed in mouse and rabbit nuclear

Chapter 23 Production of Genetically Identical Embryos 385
transplantations (Robl et al., 1986; Stice and Robl, 1988). Moreover, Szollosi etal.
(1988) showed that nuclear membrane breakdown and further reassembly of a new
nuclear envelop occurred only when fusions were performed immediately before or
up to 30 min after oocyte activation. Possibly, the nuclear membrane functions as
a barrier to a few specific factors in the oocyte cytoplasm required to reprogram the
genome's developmental pathway. Another explanation for the poor performance of
nuclear transplantations to enucleated zygotes as compared with secondary oocytes
is that developmentally important factors are sequestered within or around the
nuclear membrane and removed dur ing pronuclear enucleation.
Peri- and postimplantational studies on nuclear transplanted embryos are limited
in mammals (Table 1). It has been reported that al though apparently normal in
morphology, many of the morula—blastocyst stage embryos derived from nuclear
transplantation fail to produce pregnancy after transplantation to the uteri of syn-
chronized recipients (Prather et al., 1987; Smith and W i l m u t , 1989; Bondioli et
al., 1990). Al though many seem to die soon after implantat ion, causing an extended
estrous cycle, others may develop much further, leading to abortions at later stages
of gestation. It has also been noted that fetuses derived from nuclear transplanted
embryos are likely to be larger than usual and/or to extend slightly beyond the
normal gestation t ime (Smith and W i l m u t , 1989; Dr. K. Bondioli, personal com-
munication).
V. Applications and Conclusions
The technique of nuclear transplantation in mammals is the most rigorous test to
ascertain whether embryonic or somatic nuclei retain all of the genetic information
found in a zygote nucleus. However, other biological applications of this technique
have included (1) studies on the role of paternally and maternally derived genes in
development ("imprinting") (McGrath and Solter, 1984b; Surani et al., 1984), (2)
studies to test whether events that take place dur ing embryonic development are
inherited through the nucleus or cytoplasm (McGrath and Solter, 1983b; McGrath
and Solter, 1984c; Mann, 1986), and (3) the control of cleavage in early embryos
(Smith etal., 1990).
In practice, the technique of nuclear transplantation already provides a means
for cloning embryos in many mammalian species (Fig. 5). Therefore, it is likely
that this technique will soon become more widely available for producing genetically
identical animals either for research or for commercial application. In research, clones
could be used to eliminate the variation caused by the genotype allowing for better
assessment of nongenetic factors such as environment, disease, experimental treat-
ments , etc. For animal breeders, cloning will bring the advantage of speeding the
annual genetic gain of selection programs and enable a much shorter interval to
disseminate the gain to the commercial herds. Commercial organizations using these
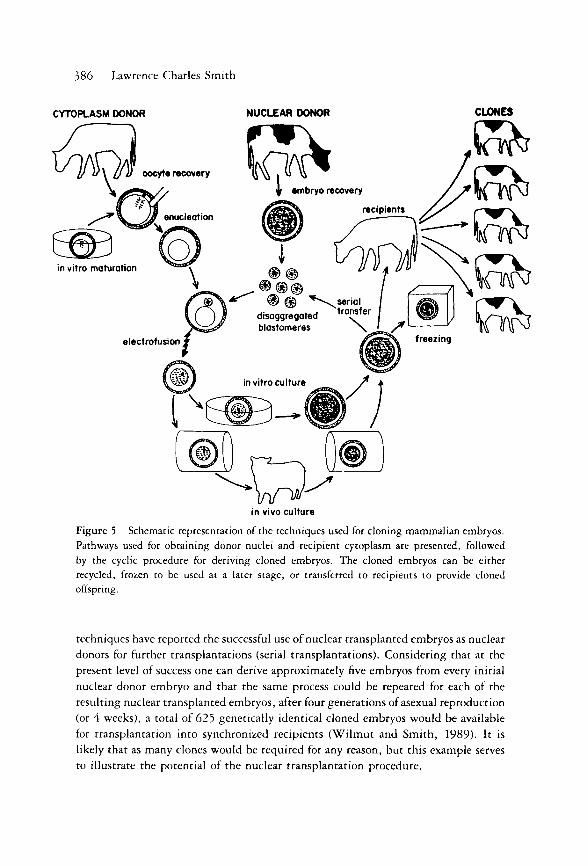
386 Lawrence Charles Smith
CYTOPLASM DONOR NUCLEAR DONOR CLONES
in vivo culture
Figure 5 Schematic representation of the techniques used for cloning mammalian embryos. Pathways used for obtaining donor nuclei and recipient cytoplasm are presented, followed by the cyclic procedure for deriving cloned embryos. The cloned embryos can be either recycled, frozen to be used at a later stage, or transferred to recipients to provide cloned offspring.
techniques have reported the successful use of nuclear transplanted embryos as nuclear
donors for further transplantations (serial transplantations). Considering that at the
present level of success one can derive approximately five embryos from every initial
nuclear donor embryo and that the same process could be repeated for each of the
resulting nuclear transplanted embryos, after four generations of asexual reproduction
(or 4 weeks), a total of 625 genetically identical cloned embryos would be available
for transplantation into synchronized recipients (Wi lmut and Smith, 1989). It is
likely that as many clones would be required for any reason, but this example serves
to illustrate the potential of the nuclear transplantation procedure.

Chapter 23 Production of Genetically Identical Embryos 387
Recent reports have indicated progress in deriving embryonic stem (ES) cells
from farm animals, which may become a valuable source of genetically identical
nuclei for cloning (Evans et al., 1990). The ease of selecting for integration and
expression of transfected ES cells in vitro will also allow for a more efficient route
for producing transgenic livestock than the current methodologies of pronuclear
injection and chimeras produced from transfected ES cells. At this point , it is also
important to note some of the possible disadvantages in the cloning procedure. One
aspect concerns the decrease in the genetic variability by inbreeding. Large foun-
dation populations should be obtained when using cloning in conjunction with
M O E T (multiple ovulation and embryo transfer) selection schemes. It is also im-
portant to verify the degree to which cytoplasmic inheritance can influence animal
production, since clones will be exposed not only to different uterine and neonatal
environments but also to a different ooplasm. The maternal inheritances of mi to-
chondrial genes are examples of differences that may arise between nuclear trans-
plantation-derived clones. Other potential factors of variation to be considered are
the random inactivation of X chromosomes in females and the possibility of differ-
ential expression or mutat ion after nuclear transplantation in cloned embryos. These
final comments help to strengthen the notion that many aspects of the interactions
between nucleus and cytoplasm after transplantation remain unknown and that it
is only by both fundamental and applied research that further improvements may
be achieved with the cloning of mammalian species.
Acknowledgments
The author thanks Dr. A. K. Goff for helpful comments and NSERC, CAAB, and FCAR for their financial support during the preparation of this manuscript.
References
Barra, J . , and Renard, J . - .P . (1988). Diploid mouse embryos constructed at the late 2-cell
stage from haploid parthenotes and androgenotes can develop to term. Development 102,
773-779. Bondioli, K. R., Westhusin, M. E., and Looney, C. R. (1990). Production of identical
bovine offspring by nuclear transfer. Theriogenology 33, 165-174. Briggs, R. (1977). Genetics of cell type determination. In "Cell Interactions in Differentiation"
(L. Saxen and L. Weiss, eds.), pp. 23—44. Academic Press, New York. Briggs, R., and King, T. J. (1952). Transplantation of living nuclei from blastula cells into
enucleated frog eggs. Proc. Natl Acad. Sei. USA 38, 455-467. Briggs, R., and King, T. J. (1953). Factors affecting the transplantability of nuclei of frog
embryonic cells. J. Exp. Zool. 122, 485-505. Briggs, R., and King, T. J. (I960). Nuclear transplantation studies on the early gastrula
{Rana pipiens). Dev. Biol. 2, 252-270.

388 Lawrence Charles Smith
Bromhall, J. D. (1975). Nuclear transplantation in the rabbit egg. Nature (Lond.) 258, 719— 722.
Brun, R. B. (1978). Developmental capacities of Xenopus eggs, provided with erythrocyte or erythroblast nuclei from adults. Dev. Biol. 65, 271—289.
Camous, S., Kopecny, V., and Flechon, J.-E. (1986). Autoradiographic detection of the earliest stage of (3H)-uridine incorporation. Biol. Cell 58, 195-200.
Clement, Α., Meyer, J . , and Brem, G. (1988). Electrofusion of early mammalian embryos. In "New Developments in Biosciences; Their Implications for Laboratory Animal Sciences" (A. C. Beynen and H. A. Solleveld, eds.), pp. 343-347. Martins Nijhoff, Dorrecht, Netherlands.
Collas, P., and Robl, J. M. (1990). Factors affecting the efficiency of nuclear transplantation in the rabbit embryo. Biol. Reprod. 43 (in press).
Collas, P., and Robl, J. M. (1991). Development of rabbit nuclear transplant embryos from morula and blastocyst stage donor nuclei. Theriogenology (in press).
Crosby, I. M., Gandolfi, F., and Moor, R. M. (1988). Control of protein synthesis during early cleavage of sheep embryos. J . Reprod. Fertil. 82, 769-775.
DiBerardino, M. A. (1979). Nuclear and chromosomal behavior in amphibian nuclear trans-plants. Inter. Rev. Cytol. Suppl. 9, 129-160.
DiBerardino, M. A. (1980). Genetic stability and modulation of metazoa nuclei transplanted
into eggs and oocytes. Differentiation 17, 17—30. DiBerardino, Μ. Α., and Hoffner, N. J. (1970). Origin of chromosomal abnormalities in
nuclear transplants—A réévaluation of nuclear differentiation and nuclear equivalence in amphibians. Dev. Biol. 23, 185-209.
DiBerardino, Μ. Α., and Hoffner, N. J. (1971). Development and chromosomal constitution of nuclear transplants derived from male germ cells. J . Exp.Zool. 176, 61-78.
DiBerardino, Μ. Α., and Hoffner, W. J. (1980). The current status of cloning and nuclear reprogramming in amphibian eggs. In "Differentiation and Neoplasia," Vol. 2 (R. G. McKinnel et al., eds.), pp. 53—64. Springer-Verlag, Berlin.
Ellinger, M. S. (1978). The cell cycle and transplantation of Blastula Nuclei in Bombina orientalis. Dev. Biol. 65, 81-89-
Evans, M. J. , Notarianni, Ε., Laurie, S., and Moor, R. M. (1990). Derivation and preliminary characterization of pluripotent cell lines from porcine and bovine blastocysts. Theriogenology 33, 125-128.
Flach, G , Johnson, M. H. , Braude, P. R., Taylor, R. A. S., and Bolton, V. N. (1982). The transition from maternal to embryonic control in the 2-cell mouse embryo. EMBO J. 1, 681-686.
Gordon, I., and Lu, Κ. Η. (1990). Production of embryos in vitro and its impact on animal production. Theriogenology 33, 77—88.
Gurdon, J. B. (I960). The effects of ultra-violet irradiation on uncleaved eggs of Xenopus laevis. Q.J. Microsc. Soc. 101, 299-311.
Gurdon, J. B. (1974). The genome in specialized cells as revealed by nuclear transplantation in amphibia. In "The Cell Nucleus," Vol. 1 (H. Bush, ed.), pp. 471-488. Academic Press, New York.
Gurdon, J. B., Laskey, R. Α., and Reeves, O. R. (1975). The developmental capacity of nuclei transplanted from keratinized skin cells of adult frogs. J. Embryol. Exp. Morphol. 34, 93-112.

Chapter 23 Production of Genetically Identical Embryos 389
Illmensee, K., and Hoppe, P. C. (1981). Nuclear transplantation in Mus musculus: Devel-opmental potential of nuclei from preimplantation embryos. Cell 23, 9-18.
King, T. J. (1966). Nuclear transplantation in amphibia. Methods Cell Biol. 2, 1. Kono, T., and Tsunoda, Y. (1988). Effects of induction current and other factors on large
scale electrofusion for pronuclear transplantation of mouse eggs. Gamete Res. 19, 349— 357.
Kono, T., Shioda, Y., and Tsunoda, Y. (1988). Nuclear transplantation of rat embryos. J . Exp. Zool. 248, 303-305.
Kubiak, J. Z. , and Tarkowski, A. K. (1985). Electrofusion of mouse blastomeres. Exp. Cell Res. 157, 561-566.
Laskey, R. Α., and Gurdon, J. B. (1970). Genetic content of adult somatic cells tested by nuclear transplantation from cultured cells. Nature (Lond.) 228, 332-358.
Levanduski, M. J . , and Westhusin, M. E. (1990). Effect of cytoskeletal inhibitors on fusion and development of bovine nuclear transfer embryos. Theriogenology 33, 273 (abstr.).
Manes, C. (1977). Nucleic acid synthesis in preimplantation rabbit embryo. III. A "dark period" immediately following fertilization and the early predominance of low molecular weight RNA synthesis. J . Exp. Zool. 201, 247-258.
Mann, J. L. (1986). DDK-egg foreign sperm incompatibility in mice is not between the pronuclei. J. Reprod. Fertil. 76, 779-781.
Marx, J. L. (1983). Swiss research questioned. Science 220, 1023. Marx, J. L. (1988). Cloning sheep and cattle embryos. Science 239, 463-464. McAvoy, J. W. , Dixon, R. E., and Marshall, J. A. (1975). Effects of differences in mitotic
activity, stage of cell cycle and degree of specialization of donor cells of nuclear trans-plantation in Xenopus laevis. Dep. Biol. 45, 330—352.
McGrath, J . , and Solter, D. (1983a). Nuclear transplantation in the mouse embryo by microsurgery and cell fusion. Science 220, 1300-1302.
McGrath, J . , and Solter, D. (1983b). Nuclear transplantation in mouse embryos. J . Exp. Zool. 288, 355-362.
McGrath, J . , and Solter, D. (1984a). Inability of mouse blastomere nuclei transferred to enucleated zygotes to support development in vitro. Science 226, 1317-1319.
McGrath, J . , and Solter, D. (1984b). Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell 37, 179—183.
McGrath, J. , and Solter, D. (1984c). Maternal Thp lethality in the mouse is a nuclear, not cytoplasmic defect. Nature (Lond.) 308, 550-551.
McGrath, J . , and Solter, D. (1986). Nucleocytoplasmic interactions in the mouse embryo. J. Embryol. Exp. Morphol. 97, suppl., 277-289.
McKinnel, R. G. (1978). "Cloning, Nuclear Transplantation in Amphibia." University of Minnesota, Minneapolis.
McLaren, A. (1984). Methods and success of nuclear transplantation in mammals. Nature (Lond.) 309, 671-672.
Modlinski, J. A. (1978). Transfer of embryonic nuclei to fertilized mouse eggs and devel-opment of tetraploid blastocysts. Nature (Lond.) 273, 466-467.
Norberg, H. S. (1973). Ultrastructural aspects of the preattached pig embryo: cleavage and early blastocyst stages. A. Anat. EnstGesch 143, 45-114.
Ozil, J . -P. , and Modlinski, J. A. (1986). Effect of electric field on fusion rate and survival of 2-cell rabbit embryos. J . Embryol. Exp. Morphol. 96, 211-228.

390 Lawrence Charles Smith
Prather, R. S., Barnes, F. L., Sims, M. M., Robl, J. M., Eyestone, M. H. , and First, N. L. (1987). Nuclear transplantation in the bovine embryo: Assessment of donor nuclei and recipient oocyte. Biol. Reprod. 37, 859-866.
Prather, R. S., Sims, M. L., and First, N. L. (1989). Nuclear transplantation in early pig embryos. Biol. Reprod. 41 , 414-418.
Robl, J. M., Gilligan, B., Critser, E. S., and First, N. L. (1986). Nuclear transplantation in mouse embryos: Assessment of recipient cell stage. Biol. Reprod. 34, 733-739.
Robl, J. M., Prather, R. S., Barnes, F., Eyestone, W. , Northey, D., Gilligan, B., and First, N. L. (1987). Nuclear transplantation in bovine embryos./ . Anim. Sei. 64, 6 4 2 -
647. Smith, L. C , and Wilmut, I. (1989). Influence of nuclear and cytoplasmic activity on the
development in vivo of sheep embryos after nuclear transplantation. Biol. Reprod. 40, 1027-1035.
Smith, L. C , Wilmut, I., and Hunter, R. H. F. (1988). Influence of cell cycle stage at nuclear transplantation on the development in vitro of mouse embryos. J. Reprod. Fertil. 84, 619-624.
Smith, L. C , Wilmut, I., and West, J. D. (1990). Cleavage control in single-cell recon-stituted mouse embryos. J. Reprod. Fertil. 88, 655-663.
Solter, D. (1987). inertia of the embryonic genome in mammals. Trends Genet. 3, 23—27. Solter, D., and Knowels, Β. B. (1975). Immunosurgery of mouse blastocysts. Proc. Natl.
Acad. Sa. USA 72, 5099-5102. Solter, D., Aronson, J . , Gilbert, S. F., and McGrath, J. (1986). Nuclear transfer in mouse
embryos: activation of the embryonic genome. In "Molecular Biology of Development," 50th Symp. Quant. Biol. Vol. L, pp. 45-50. Cold Spring Harbour, New York.
Stice, S. L., and Robl, J. M. (1988). Nuclear reprogramming in nuclear transplant rabbit embryos. Biol. Reprod. 39, 657-664.
Surani, Μ. A. H., Barton, S. C , and Norris, M. L. (1984). Development of reconstituted eggs suggested imprinting of the genome during gametogenesis. Nature (Lond.) 308, 548-550.
Surani, Μ. A. H. , Barton, S. C , and Norris, M. L. (1987). Experimental reconstruction of mouse eggs and embryos: An analysis of mammalian development. Biol. Reprod. 36, 1-16.
Szollosi, D., Czolowska, R., Szollosi, M. S., and Tarkowski, A. K. (1988). Remodeling of mouse thymocyte nuclei depends on the time of their transfer into activated homologous oocytes. J. Cell Set. 91 , 603-613.
Tsunoda, Y., Kato, Y., and Shioda, Y. (1987a). Electrofusion for the pronuclear transplan-tation of mouse eggs. Gamete Res. 17, 15—20.
Tsunoda, Y., Yasui, T., Shioda, Y., Nakamura, K., Uchida, T., and Sugie, T. (1987b). Full term development of mouse blastomere transplanted into enucleated two-cell embryos. J. Exp. Zool. 242, 147-151.
Tsunoda, Y., Shioda, Y., Onodera, M., Nakamura, Κ., and Uchida, T. (1988). Differential sensitivity of mouse pronuclei and zygote cytoplasm to Hoeschst staining and ultraviolet irradiation. J. Reprod. Fertil. 82, 173-178.
Tsunoda, Y., Maruyama, Y., and Kila, M. (1989). Nuclear transplantation of 8- to 16-cell embryos into enucleated oocytes in the rabbit. Jpn. J. Zootech. Sei. 60, 846-851.

Chapter 23 Production of Genetically Identical Embryos 391
Tsunoda, Y., Tokunaga, T., Imai, H. , and Uchida, T. (1990). Nuclear transplantation of male primordial germ cells in the mouse. Development 107, 407-411.
Wabl, M. R., Brun, R. B., and Pasquier, L. (1975). Lymphocytes of the toad Xenopus laevis have the gene set for promoting tadpole development. Science 190, 1310.
Ware, C. B., Barnes, F. L., Maija-Lauiro, M., and First, N. L. (1989). Age dependence of bovine oocyte activation. Gamete Res. 22, 265—275.
Willadsen, S. M. (1986). Nuclear transplantation in sheep embryos. Nature (Lond.) 320,
63-65. Willadsen, S. M., and Fehilly, C. B. (1983). The developmental potential and regulatory
capacity of blastomeres from 2-, 4- and 8-cell sheep embryos. In "Fertilization of the Human Egg in Vitro; Biological and Clinical Applications" (H. M. Beier and H. R. Lindner, ed.), pp. 27-52. Springer-Verlag, Berlin.
Wilmut, I., and Smith, L. C. (1989). Biotechnology and the bovine embryo: At present and in the future. In "Colloque Scientifique", 4th ed., pp. 19-31. Α. Ε. T. E., Lyons, France.
Zimmermann, U., and Vienken, J. (1982). Electric field induced cell-to-cell fusion. J. Membrane Biol. 67, 165-182.





























