Embed Size (px)
Citation preview

Maydica 45 (2000): 95-103
GRAIN FILLING IN MAIZE: THE EFFECT OF NITROGEN NUTRITIONON THE ACTIVITlES OF NITROGEN ASSIMIlATING ENZY'MES
IN THE PEDICEL-PIACENTO-CHAIAZA REGION
ANDRADC.L. I
2000
A.Á.C.Purcino", M.R. e Silva, S.R.M. Andrade, c.L. Belele, S.N. Parentoni, M.X. dos Santos
Ernbrapa Maize and Sorghum, Caixa Postal 151, 35.701-970 Sete Lagoas, MG, Brazil
Received February 11, 2000
ABSTRACf - In a developing maize kernel, the pedicel-placento-chalaza (PPCh) is a heterogeneous tissue be-rween the cob vascular system and the endosperm. Its ma-jor role is to modify the cob vascular system amino acidpool composition to better suit the requirements of the de-veloping endosperm. Nitrogen metabolism in the PPCh tis-sue may, therefore, influence individual kernel develop-ment and consequently play an important role in deter-mining grain yield capacity in maize. The objectives of thetwo experiments reported in this work were: (1) to deter-mine the effect of nitrogen supply on the activities of thenitrogen assimilating enzymes glutamine synthetase (GS),NADH-dependent glutamate synthase (NADH-GOGAT),glutamate dehydrogenase (GDH) and of phospho-enolpyruvate carboxylase (PEPC) in the pedicel-placento-chalaza (PPCh) tissue of maize genotypes contrasting intheir responsiveness to nitrogen (N) fertilization, and (2)to verify if the activities of these enzymes in the PPCh tis-sue are correlated to individual kernel weight. Hybridswere grown in the field under low (16 kg!ha) and high(116 kg/ha) supply and hand pollinated ar silking. En-zyme activities were measured 30 days after pollination(DAP) and individual kernel weight was determined afterblack layer formation. Nitrogen supply did not influenceindividual kernel weight, soluble protein, GS and PEPCactivities. However inconsistent results were observed forNADH-GOGAT and GDH over experimental years. En-zyme activities varied among genotypes, but correlationdeterminations indicated that individual kernel weight wasnot associated with soluble protein content and GS activi-ty, with discrepant results observed for NADH-GOGATand GDH over years. However, a positive and significantcorrelation was observed between kernel weight andPEPC activity in both experiments. This finding merits fur-ther investigations over a larger number of genotypes anddifferent environmental conditions. It suggests that geno-types with increased PEPC activity in the PPCh tissue haveincreased kernel weight and that this enzyme plays an im-portant role in determining grain yield in maize.
• For correspondence Cfax+55 31 779 1088; e-mail: [email protected]).
KEY WORDS: Grain filling; Pedicel-placento-chalaza; GS;GOGAT; GDH; PEPe.
INTRODUCTION
In a developing maize kernel, the pedicel-pla-cento-chalaza (PPCh) is a heterogeneous maternaltissue between the incoming cob vascular systemand the endosperm and embryo. Before entering in-to the basal part of the endosperm, nutrients andsugars are unloaded into the pedicel, enter theapoplast of the placento-chalaza region and subse-quently are taken up by the endosperm via severallayers of transfer cells. Whether this transfer systemis an active process remains uncertain (PORTERet al.,1987 and references therein, LYZNIK et ai., 1989) butone striking feature of the PPCh is that the concen-tration of sugars and amino acids in the vegetativecob vascular termini is qualitatively and quantita-tively different from the endosperm. These differ-ences may indicate that the PPCh is not just a barri-er between the vascular system of the cob and theendosperm, but it may in fact play an importantmetabolic role in the kernel nitrogen metabolismo
Glutarnine and, to a lesser extent, aspartic acid arethe major components of the cob vascular sap buttheir \evels tend to decline with a simultaneous in-crease in the contents of valine, methionine,isoleucine, leucine, tyrosyne and phenylalanine askernel maturation occurs (ARRUDA and SILVA, 1979,1983). On the other hand, the PPCh contains moreglutamine, glutamate and proline than the vascularsap and more asparagine, aspartic acid, serine and ly-sine than de endosperm. Moreover, although gluta-mine content in the vascular sap is only half of that inthe PPCh, it tends do decline with kernel develop-ment, whereas glutamine content in the PPCh re-mains fairly constant (LYZNIK et al., 1985), Zeins, themajor storage proteins in maize, function as N sinks

96 A.Á.C. PURCINO, M.R. E SILVA.S.R.M. ANDR.A.DE. c.t, BELELE, S.N. PARENTONI, M.X. DOS SANTOS
TABU: 1 - Characteristics of the maize genotypes used in the field experiment (adapted from PURClNO et al., 1994, SAN1DS et ai. and PARE.\7DS/
et al., unpuhlisbed results),
Genotype Characteristics
Three way cross, Swan x Tuxpeno, short. early, yields better than HT(1167 x 1143) x 16.3.1Three way cross, Swan x Tuxpeno, short, earlySingle cross hybrid, Tuxpeno, short, early, high yíeld under low N, responsive to N fertilizationSingle cross hybrid, Tuxpeno, short, early, high yield under low N, less responsive to N fertilization than HS 22 x 20Single cross hybrid, Tuxpeno dent, short, early. Under low N yields less than HS 22 x 20, HS 22 x 724 and BR 201-M but responsive to N fertilizationSingle cross hybrid, Caribbean flint, short, early, yields better than BR 201-F under low N but less responsive to Nfertilization
HT (1167 x 1143) x 6.1.3.1HT 0167 x 1143) x 16.3.1HS 22 x 20HS 22 x 724BR 201-F
13R201-M
for nitrogenous compounds in the endosperm (TSAIetal., 1986) and its four classes of proteins (27, 22, 14and 10 kD proteins) tend to be rich in proline, gluta-mine and leucine, but low in aspartic acid, Iysine andtriptophane (LOPES,1993). It seems, therefore, that thePPCh region has a "stabílizing effect" on the aminoacid flux into the endosperm and its major function isto modify the vascular system amino acid pool com-position to better fit the endosperm requirements forthe synthesis of storage proteins (LYZNIKet al., 1982;RAcZYNSKA-Bo]ANOWSKAet al., 1986).
As should be expected from these intense metabol-ic activities, several key enzymes responsible for aminoacíd metabolism as well as for catabolic and anabolicactivities have been reported to occur in the PPCh, en-dosperm and ernbryo of the developing kernel (SODEKand DA SILVA,1977; TSAl, 1979; MISRAand OAKS,1981;LYZNIKet al., 1985). For the inbred line w64A, the spe-cific activities of glutarnine synthetase (GS), glutamatesynthase (GOGAT) and glutamate dehydrogenase(GDH) are significantly higher in the PPCh than in theendosperm at 15 days after pollination (DAP) (RACZYNS-KA-BO]ANOWSKAet al., 1986), but more important is thatthese activities vaty significantly among inbred lines(Tsxr 1979, RACZYNSKA-BojANOWSKAet al., 1986). Theseenzymes may, therefore, play an importam role in de-termining the yield capacity of maize.
Considering that information about the effect ofN nutrition on the activities of the PPCh N-assimilat-ing enzymes of maize genotypes contrasting in theiryield potential and responsiveness to N supply -islacking, this study was initiated with the following
.objectives: (1) to determine the effect of nitrogensupply on the activities of GS, GOGAT, GDH andphosphoenolpyruvate carboxylase (PEPC) in thePPCh tissue of maize genotypes with contrasting re-sponsiveness to N fettilization, and (2) to verify ifthe activities of these enzymes in the PPCh tissue iscorrelated to individual kernel weight.
MATERIALS AND METHODS
Field ExperimentSix maize genotypes (Table 1) were evaluated in combina-
tion with two nitrogen levels [16 and 116 kg/ha N as (NH4)zS04l.The three way crosses HT (1167 x 1143) x 6.1.3.1 and HT (1167x 1143) x 16.3 1 and the single cresses HS 22x20 and HS 22x724were compared in 1994/95 whereas in 1996/97 the three waycrosses were substituted for the singles crosses BR 201-F and BR201-M. The experiments were carried out as a randomized com-plete block design with genotypes as a split plot on nitrogen lev-els. Subplots consisted of four rows five rneters long and 0.90 mapart with five plants/m. Each treatment combination was repli-cated four times. The experimental site soil was a kaolinitic iso-hypertherrníc Typic Haplustox originally under savanah-typevegetation, known in Brazil as Cerrado, 760 m above sea level,19° 27' S and 44° 14' W. Before plantíng, the area was limed anelfertilizeel with macro- anel micronutrients according to soil fertili-ty tests. Cultural practices wcre those normally recommeneled forrnaíze, and supplemental irrigation was used when needed. Up-on silking, 20 main ears in the two central rows of each treat-ment were sib pollinated and baggeel with Kraft paper. Thirtydays after pollínatíon (DAP) - when enzyme activities in thePPCh tend to reach its maximum (RACZYNSKA-BoJANOWSKAet al.,1986; MISRAand OAKS, 1981) - bagged cars from 10 compctítiveplants of each treatment cornbination were harvested and imme-diately taken to the laboratory. The pedicel-placento-chalaza tis-sue was carefully excised frorn the basal portion of the ear,wrapped in aluminum foil and stored at -80°C until analysis. Re-maining unbagged ears from competitive plants were used forthe determination of indivielual kernel weight (weight of 1000kernels/l000 expressed in mg) corrected to 13% moísture. Analy-sis of variances and correlations were cornputed with the MSTACsoftware package and means were compared using the Duncan'srnultíple range test at P<O.OS.
Enzyme ExtractionAll reagents for extraction and determination of enzyme ac-
tivities were obtained from Sigma Chemical Co. Glutamine syn-thetase (GS, EC 6.3.1.2), NADH-dependent glutarnate synthase(NADH-GOGAT, EC 1.4.7.14) and glutamate dehydrogenase(GDH, EC 1.4,1.2) were extracted with 50 mM Tris-HCI pH 7.5containing 10 mM 2-mercaptoethanol (f:\ME) and 1 111M ethyl-enediaminetetraacetic acid (EDTA). One gram of tissue wasground with sand in 5 mL of the extraction buffer, centrifugedat 18.000 g for 20 minutes and 1 mL of the clear supernatantwas desalted in 5 mL Sephadex G-25 columns pre-equilibrated

ENZYMEACTIVITIESIN THE PPCH TISSUE 97
with the extraction buffer. Ali procedures were carried out be-tween O-4°C.
Phosphoenolpyruvate carboxylase (PEPC, EC 4.1.1.31) wasextracted with 100 mM phosphate buffer pH 7.5 containing 10mM MgClz.6Hp, 1 mM EDTA, 2 mM dithiothreitol (DTT) , 0.8mM phenylmethylsulfonyl fluoriele (PMSF) and 10% polyvi-nylpolypyrrolidone (PVVP) with the same protocol used for theextraction of the other enzymes.
Determination of enzyme activityThe híosynthetic activity of GS was measureel in a 1 mL reac-
tion mixture containing 100 mM Tris-HCI pH 7.5, 10 mM 2-(,;ME,20 mM MgS04.7H20, 15 mM hydroxylamine hydrocloride(NHzOH.HCl), 10 mM glutamate and 300 pL of the crude extract.After incubation at 30°C for 30 minutes the reaction was stoppedwith the addition of 1 mL of the FeCl3 reagem (300 mM FeCI3,
670 mM HCI, 200 mM TCA). The forrnation of glutamyl hidroxa-mate (GHA) was determined at 540 nm and enzyme activity wascalculated from a standard curve prepared with pure GHA.NADH-GOGAT actívíty was determined in a 3 mL reaction mix-ture containing 50 mM Tris-HCI pH 7.5,5 mM 2-oxoglutarate (pHadjusteel to 7), 5 mM glutamine, 0.2 mM NADH and 100 ul. of thecrude enzyme extract. This mixture was incubatcd at 30°C andthe rate of change in absorbance at 340 nm was determined be-tween zero anel 15 minutes. GDH activity in the aminating direc-tion was determined by monitoring the decrease in absorbanceat 340 nm at 30°C, in a reaction mixture comaining 100 mM Tris-HCI pH 7.5, 100 mM (NH4)2S04, 20 mM 2-oxoglutarate (pH ad-justed to 7), 10 mM CaClj, 4 mM NADH and 100 !JLof cruele en-zyme extract diluteel 3X. PEPC activity was measured spec-trophotometrically in a coupled reaction with malate dehydroge-nase (MDH, EC 1.1.1.37). The 1 mL reaction mixture contained50 mM phosphate buffer pH 7.5, 10 mM 2-mercaptoethanol, 12lU MDH, 5 mM glucose-6-phosphate, 0.2 mM NADH, 5 mM PEPand 100 ul, of the crude enzyme extract eliluteel 5X. The reactionwas started with PEP and the oxidation of NADH was monitoredat 340 nm. Corrections for substrate-índependcnt activities weremaele for all NADH-dependent activities.
Determination of soluble proteínTotal pedicel-placento-chalaza tissue soluble protein concen-
tration was determined according to the method of BRADFORD(1976) using bovine serum albumin as the standarel.
RESULTS AND DISCUSSION
Previous field experiments have indicated thatthe single cross hybrids HS 22 X 20 and HS 22 X
724 produce more grain yield than the single cross-es BR 201-F and BR 201-M. In terms of responsive-ness to N fertilization these singles crosses wereranked HS 22 X 20 > HS 22 x 724 > BR 201-F > BR201-M (PURCINO et al., 1994; PARENTONI et al.; MAR-RIELet al. unpublished results). Hybrids HS 22 x 20,HS 22 x 724 and BR 201-F share a common inbredline, but BR 201- M is totally unrelated to all ofthem. The three way cross hybrid HT (1167 x 1143)x 6.1.3.1 produces more grain yield than HT (1167x 1143) x 16.3.1 but information about their re-sponsiveness to N is not available (Santos, unpub-lished results).
Individual Kernel WeightValues obtained for individual kernel weight
measurements obtained in these experiments arepresented in Table 2 and are comparable to figuresreported for the inbred line w64A (CULLY et al.,1984) and several other commercial hybrids (TOL-LENAARet al., 1992). In 1994/95 kernel weight wasnot influenced by N fertilization but it varied amonggenotypes, and in 1996/97 it was influenced by asignificam genotype x N leve! interaction. In thefirst experiment, kernels of HS 22 x 724 were signif-icantly Iighter than kernels of HS 22 x 20 but theirvalues were not different in 1996/97. These two hy-brids produced heavier kernels in the 1996/97 ex-periment and apparently this "year effecr" influ-enced kernel weight more strongly than both N lev-el and genotype characteristics. In 1996/97, geno-types HS 22 x 20 and HS 22 x 724 produced lighter
TAI3LE2 - Effect of nitrogen supply on leernel weight (mgleernelr! ) of six tropical maize genotypes.
1994/95CUltivar
16 kg!ha N 116 kg/ha N Average
HT(1167X1143)x6.1.3.1 261 270 266AHT(1167X1143)XI6.3.1 260 269 264AHS 22x20 244 249 247AHS 22x724 189 195 192BBR 201-FBR 201-MAverage 239a 246aCV 11.3%
1996/97
Average16 kg!ha N 116 kg!ha N
328Aa337Aa248Ba332Ab
305Bb305Bb245Ca350Aa
5.9%
Means followed by the same letters are not significantly different by the Duncan's test at the 5% level. Capital letters for comparísons with-ín columns, small letters within rows.
98 A.Á.C.PURCIl\'O,:V1.R.ESILVA.S.R.M.ANDRADE,c.i. BELELE,S.N.PARENTONI,M.X.DOSS_\t\TOS
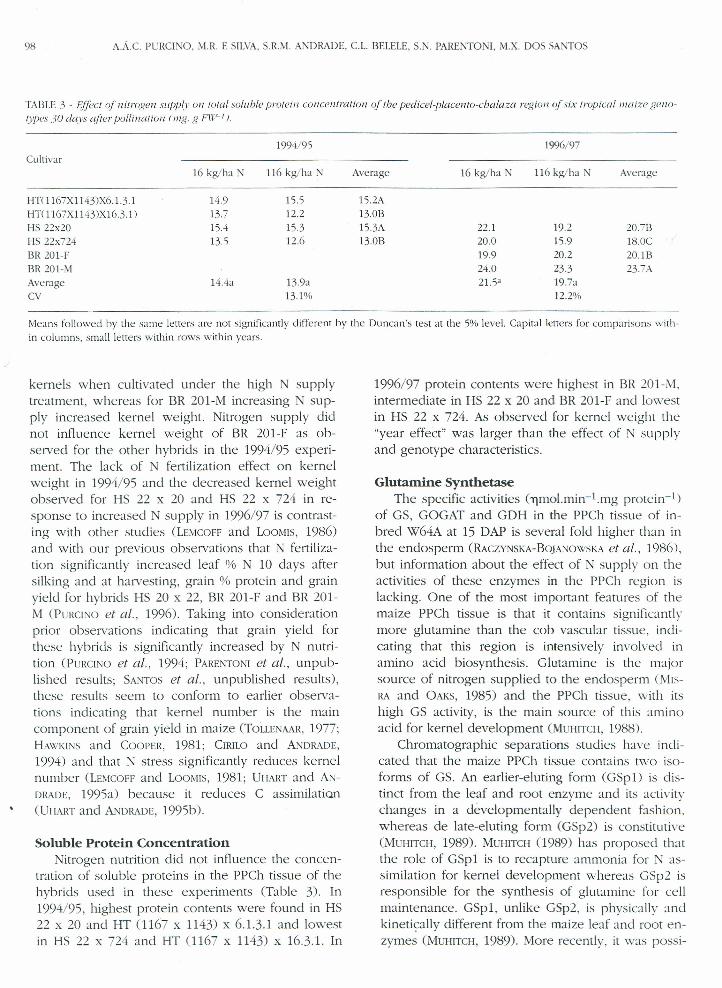
TABU:3 - EJ/ect ofnitrogen suppl» 0/1 total soluble proteiu concentration oftbe pedicet-ptacento-cbataza region ofsix tropical maire gel/o-types 30 days alia pollination t mg, g FI'(f-1).
1994/95Cultivar
16 kg/ha N 116kglha N Average
HTOI67XI143)X6.1.3.1 14.9 15.5 15.2AHT(1167X1143)XI6.3.1) 13.7 12.2 13.013I-IS22x20 15.4 15.3 15.3A]-]S22x724 13.5 12.6 13.013I3R201-FBR201-MAverage 14.4a 13.9aCV 13.1%
1996/97
16 kglha N 116kglha N Average
22.120.019924.021.5"
19.215.920.223.319.7a12.2%
20.71318.0C20.lB23.7A
Me.ms followed by the same letters are not significantlydífferent by the Duncan's test at the 5%leveI.Capital letters for cornparisons with-in columns, srnall letters within rows within years.
kernels when cultivated under the high N supplytreatment, whereas for BR 201-M increasing N sup-ply increased kernel weight. Nitrogen supply didnot influence kernel weight of BR 201-F as ob-servee.! for the other hybrie.!s in the 1994/95 experi-ment. The lack of N fertilization effect on kernelweight in 1994/95 and the decreased kernel weightobserved for HS 22 x 20 and HS 22 x 724 in re-sponse to íncreased N supply in 1996/97 is contrast-ing with other studies (LEMCOFFand LOOMIS,1986)ane.! with our previous observations that N fertiliza-tion significantly increased leaf % N 10 days aftersilking and at harvesting, grain % protein and grainyield for hybrids HS 20 x 22, BR 201-F and BR 201-M (PURCINOet ai., 1996). Taking into considerationprior observations indicating that grain yield forthese hybrids is significantly increased by N nutri-tion (PURCINOet ai., 1994; PARENTONIet al., unpub-lished results, SANTOSet al., unpublished results),these results seem to conform to earlier observa-tions indicating that kernel number is the maincomponent of grain yield in maize (TOLLENAAR,1977;HAWKINSand COOPER, 1981; CIRlLO and ANDRADE,1994) and that N stress significantly reduces kernelnumber (LEMCOFFand LOOMIS,1981; UHARTand AN-DRADE, 1995a) because it reduces C assimilaticn(UHARTand ANDRADE,1995b).
Soluble Protein ConcentrationNitrogen nutrition did not influence the concen-
tration of soluble proteins in the PPCh tissue of thehybrids used in these experiments (Table 3). In1994/95, highest protein contents were found in HS22 x 20 and HT (1167 x 1143) x 6.1.3.1 and lowestin HS 22 x 724 and HT (1167 x 1143) x 16.3.1. In
1996/97 protein contents were highest in BR 201-M.intermediate in HS 22 x 20 and BR 201-F anel lowestin HS 22 x 724. As observee.! for kernel weight the"year effect" was larger than the effect of N supplyand genotype characterístícs.
Glutamine SynthetaseThe specific activities (nmol.mirr+.mg proteírr+)
of GS, GOGAT and GDH in the PPCh tissue of in-bred w64A at 15 DAP is several fole.! higher than inthe enelosperm (RACZYNSKA-Bo.IANOWSKAet al., 1986),but information about the effect of N supply on theactivities of these enzymes in the PPCh region islacking. One of the most important features of themaíze PPCh tissue is that it contains significantlymore glutamine than the cob vascular tissue, indi-cating that this region is intensively involvee.! inamino acid biosynthesis. Glutamine is the majorsource of nitrogen supplied to the endosperm (MIS-RA and OAKS, 1985) ane.! the PPCh tissue, with itshigh GS activity, is the main source of this aminoacid for kernel development (MUHITCH,1988).
Chromatographic separations studies have indi-cated that the maize PPCh tissue contains two iso-forms of GS. An earlíer-eluting form (Gôp l ) is dis-tinct from the leaf and root enzyme and its activirychanges in a developmentally dependent fashion.whereas de late-eluting form CGSp2) is constitutive(MUHITCH,1989). MUHITCH(1989) has proposed thatthe role of GSp1 is to recapture ammonia for N as-similation for kernel development whereas GSp2 isresponsible for the synthesis of glutamine for cellmaintenance. GSp1, unlike GSp2, is physically andkínetically different from the maize leaf and root en-zymes (MUHITCH,1989). More recently, it was possi-

ENZYMEACTIVITIESINTI-lEPPCH TISSUE 99
TABLE4 - Effect of nitrogen supply on CSactivity of the pedicel-placento-cbalaza region of six tropical maize genotypes 30 days after polli-natlon (rimoles.Giin.g FW1mirrl).
1994/95Cultivar
16 kg/ha N 116 kglha N Average
HTCl167X1143)x6.1.3.1 490 639 564CHT0167Xl143)X16.3.1 606 602 604BHS 22x20 683 714 698AHS 22x724 630 677 653ABBR 201-FBR 201-MAverage 602b 658aCV 13.0%
1996/97
Average16 kg/ha N 116 kg/ha N
332385453715471a
435482480722530a24.5%
384B434B466B719A
Means followed by the same letters are not significantly different by the Duncan's test at the 5% leveI. Capital letters for comparisons with-in columns, small letters within rows within years.
ble to demonstrate that out of the 5 GS1 genesknown to occur in maize, only GSl-2 and GSl-3 areexpressed in the pedicel, but only GSl-2 is uniqueto the kernel tissue and is developmentally regulat-ed (RASTOGIet ai., 1998).
Nitrogen nutrition increased GS activity in bothyears, but results were signíficantly different onlyfor 1994/95, probably because of a larger coeffi-cient of variation associated with the 1996/97 mea-surements (Table 4). Hybrids HS 22 x 20 and HS 22x 724 had comparable GS activities in both experi-ments, but activities were considerably higher in1994/95 than in 1996/97, again indicating an im-portant "year effect". The three way cross hybridHT 0167 x 1143) x 16.3.1 had higher GS activitythan HT 0167 x 1143) x 6.1.3.1 and actívíty washigher in BR 201-M than in BR 201-F. Unlike theseresults obtained in the PPCh tissue, the total GS ac-tivities in the leaves of HS 20 x 22, BR 201-F andBR 201-M are neither influenced by N nutrition norsignificantly different among themselves (PURCINOet ai., 1996). MISRAand OAKS(981) found no sig-nificant differences for GS activity in the en-dosperm of three maize genotypes and suggestedthat the differences observed by TSAI(979) weredue to the use of undesalted crude extracts by thelatter investigator. Similarly, SINGLETARYet ai. (990)observed that in desalted endosperm extract of in-bred w64A cultured in uitro, GS activity was notinfluenced by N supply. Our present results withcrude desalted enzyme extract clearly indicate thatGS activity may vary significantly in the PPCh tis-sue of maíze genotypes and, moreover, that thisvariation is strongly influenced by an uncontrolled"year effect".
NADH-dependent GOGATIn the endosperm, the role of GOGAT is to metab-
olize glutamine into glutamate which is a substrate forseveral transaminases responsible for the biosynthesisof other seed amino acids. In the normal single crosshybrid HS 1406, GOGAT activity peaked at about 31days after pollination in a trend similar to N accumu-lation in this tissue (SODEKand DASILVA,1977), but ina opaque-2 mutant, GOGAT activity increased sharplyafter termination of zein synthesis, possibly to preventaccumulation of glutamine, a major component ofzeins (TsAI,1979). The PPCh is a fully mature tissuewhen the endosperm begins to accumulate dry matterafter pollination and has higher GOGAT activity thanthe endosperm (RACZVNSKA-BoJANOWSKAet ai., 1986).However, GOGAT activity in the PPCh does notchange as grain filling progresses, whereas activity inthe endosperm increases by ca 6 fold between 15and 30 DAP (LYZNIKet ai., 1985).
Nitrogen nutrition did not influence NADH-de-pendent GOGAT activity in the PPCh tissue of thegenotypes planted in 1994/95, but activities weresignificantly lower in plants fertilized with the high-er N levei in 1996/97 (Table 5). In 1994/95, HT(1167 x 1143) x 6.1.3.1 had higher activity than HT(1167 x 1143) x 16.3.1 and HS 22 x 20 had higheractivity than HS 22 x 724. Conversely, highestNADH-GOGAT activity was observed in HS 22 x724 in 1996/97, again demonstrating that uncon-trolled environmental conditions between growingseasons have a large effect on PPCh enzyme activi-ties. On the other hand, we have observed that Nsupply favored the activity of ferredoxine-depen-dent GOGAT in the leaves of HS 20 x 22 and BR201-M but not of BR 201-F (PURCINOet ai., 1996).
100 A.Á.e PURCINO, M.R. E SILVA. S.R.M. ANDRADE, c.t. BELELE, S.N. PARENTONI, M.X. DOS SANTOS
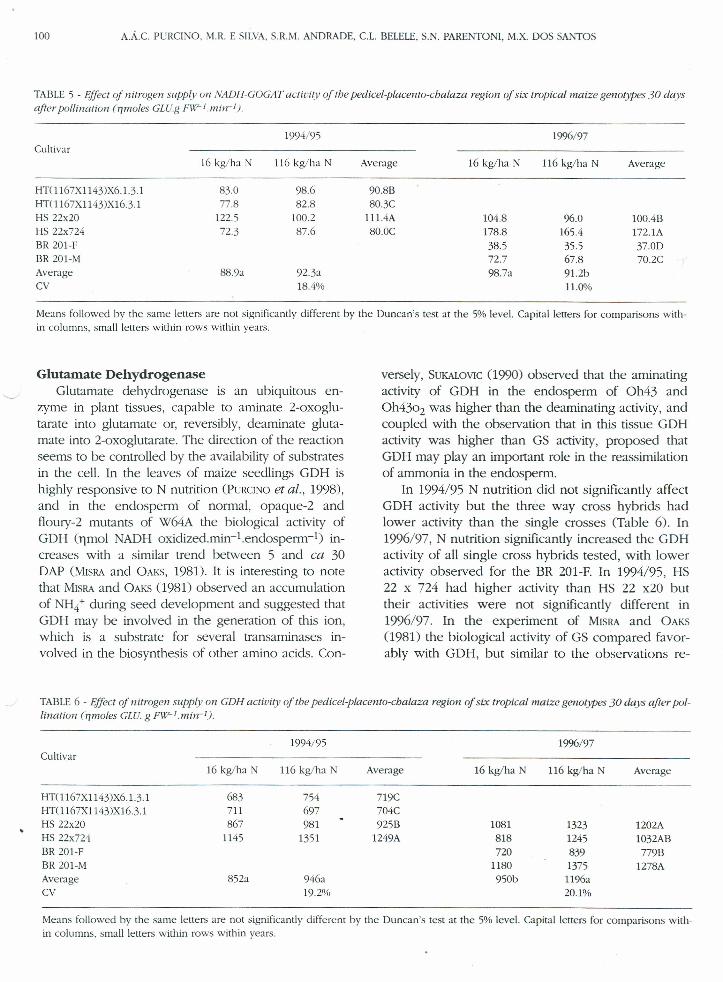
TABLE 5 - EjJecIof nilrogen supply on NADH-COCAT actiuitv of lhe pedicel.placento-cbalaza region of six tropical maize genotypes 30 daysafter poilination (nmoles CLU.g FIV-lmir,I).
1994/95Cultivar
16 kg!ha N 116 kg!ha N Average
HT(l167XI143)x6.1.3.1 83.0 98.6 90.8BHT( 1167XI143)XI6.3.1 77.8 82.8 80.3CHS 22x20 122.5 100.2 I1l.4AHS 22x724 72.3 87.6 80.0CBR 201-FBR 201-MAverage 88.9a 92.3aCV 18.4%
16 kg!ha N Average
104.8178.838.572.7
98.7a
10/)6/97
116 kg!ha N
%.0165.435.567.891.2b11.0%
100.4B172.1A37.0D70.2C
Means followed by the same letters are not significantly different by the Duncan's test at the 5% leveI. Capital letters for comparisons with-in columns, small letters within rows within years.
Glutamate DehydrogenaseGlutamate dehydrogenase is an ubiquitous en-
zyme in plant tissues, capable to aminate 2-oxoglll-tarate into glutamate or, reversibly, deaminate gluta-mate into 2-oxogllltarate. The direction of the reactionseems to be controlled by the availability of substratesin the cell. In the leaves of maize seedlings GDH ishighly responsive to N nutrition (PURCINOet al., 1998),and in the endosperm of normal, opaque-Z andfloury-Z mutants of w64A the biological activity ofGDH (nrnol NADH oxidized.mirr+endosperrrr+) in-creases with a similar trend between 5 and ca 30DAP (MISRAand OAKS, 1981). It is interesting to notethat MISRAand OAKS(981) observed an accumulationof NH4 + during seed development and suggested thatGDH may be involved in the generation of this ion,which is a substrate for severa I transaminases in-volved in the biosynthesis of other amino acids. Con-
versely, SUKALOVlC(990) observed that the aminatingactivity of GDH in the endosperm of Oh43 andOh4302 was higher than the deaminating activity, andcoupled with the observation that in this tissue GDHactivity was higher than GS activity, proposed thatGDH may play an important role in the reassimilationof ammonia in the endosperm.
In 1994/95 N nutrition did not significantly affectGDH activity but the three way cross hybrids hadlower activity than the single crosses (Table 6). In1996/97, N nutrition significantly increased the GDHactivity of all single cross hybrids tested, with loweractivity observed for the BR 201-F. In 1994/95, HS22 x 724 had higher activity than HS 22 x20 buttheir activities were not significantly different in1996/97. In the experiment of MISRA and OAKS(981) the biological activity of GS compared favor-ably with GDH, but similar to the observations re-
TABLE 6 - EjJect of nitrogen supply on CDH aaioity of lhe pedicel-placento-cbalaza region of six tropical maize genotypes 30 days after pol-lination (nmoles CLU. g FWlmil',l).
1994/95Cultivar
16 kg!ha N 116 kg/ha N Average
HT(l167XI143)X6.1.3.1 683 754 719CHT0167X1143)XI6.3.1 711 697 704CHS 22x20 867 981 925BHS 22x724 1145 1351 1249ABR 201-FBR 201-MAverage 852a 946aCV 19.2%
16 kg/ha N Average
1081818720
1180950b
19%/97
116 kg/ha N
1323124583913751196a20.1%
1202A1032AB779B
1278A
Means followed by the same letters are not significantly different by the Duncan's test at the 5% leveI. Capital letters for comparisons with-in columns, small letters within rows within years.
ENZYMEACTIVITIESIN THEPPCHTISSUE 101
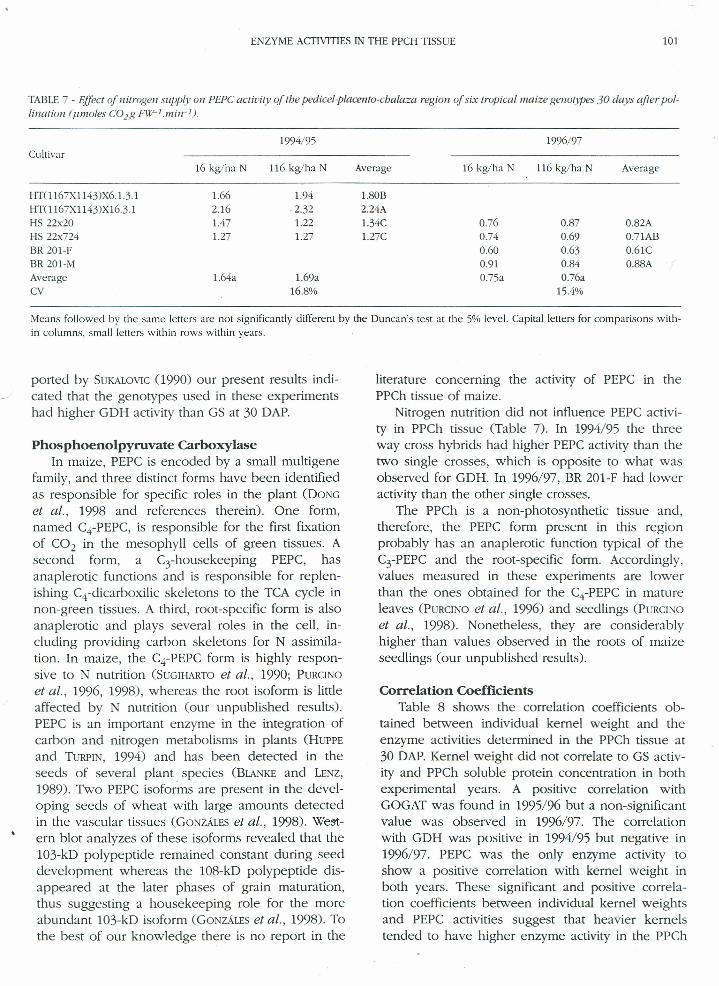
TABLE7 - Effect of nitrogen supply 0/1 PEPC actioity of the pedicel-placento-cbalaza region of six tropical maize genotypes 30 days after pol-lination (umoles CO2B Fll7-lmin-l).
1994/95Cultivar
16 kg!ha N 116kg!ha N Average
HT(1167XI143)x6.1.3.1 1.66 1.94 1.80BHTOI67XI143)XI6.3.1 2.16 ·2.32 2.24AHS22x20 1.47 1.22 1.34CHS22x724 1.27 1.27 1.27CBR201-FBR201-MAverage 1.64a 1.69aCV 16.8%
1996/97
16 kg!ha N 116kg!ha N Average
0.760.740.600.910.75a
0.870.690.630.840.76a
15.4%
0.82A0.71AB0.61C0.88A
Means followed by the same letters are not significantlydifferentby the Duncan's test at the 5%leveI.Capitalletters for comparisons with-in columns, small letters within rows within years.
ported by SUKALOVlC(990) our present results indi-cated that the genotypes used in these experimentshad higher GDH activity than GS at 30 DAP.
Phosphoenolpyruvate CarboxylaseIn maize, PEPC is encoded by a small multigene
family, and three distinct forms have been identifiedas responsible for specific roles in the plant (DONGet al., 1998 and references therein). One form,named C4-PEPC, is responsible for the first fixationof CO2 in the mesophyll cells of green tissues. Asecond form, a C3-housekeeping PEPC, hasanaplerotic functions and is responsible for replen-ishing C4-dicarboxilic skeletons to the TCA cyde innon-green tissues. A third, root-specific form is alsoanaplerotic and plays severa I roles in the cell, in-duding providing carbon skeletons for N assimila-tion. In maize, the C4-PEPC form is highly respon-sive to N nutrition (SUGIHARTOet al., 1990; PURCINOet al., 1996, 1998), whereas the root isoform is littleaffected by N nutrition (our unpublished results).PEPC is an important enzyme in the integration ofcarbon and nitrogen metabolisms in plants (HUPPEand TURPIN, 1994) and has been detected in theseeds of severa I plant species (BLANKEand LENZ,1989). Two PEPC isofor·ms are present in the devel-oping seeds of wheat with large amounts detectedin the vascular tissues (GONZÃLESet al., 1998). West-ern blot analyzes of these isoforms revealed that the103-kD polypeptide remained constant during seeddevelopment whereas the 108-kD polypeptide dis-appeared at the later phases of grain maturation,thus suggesting a housekeeping role for the moreabundant 103-kD isoform (GONZÃLESet al., 1998). Tothe best of our knowledge there is no report in the
literature concerning the activity of PEPC in thePPCh tissue of maize.
Nitrogen nutrition did not influence PEPC activi-ty in PPCh tissue (Table 7). In 1994/95 the threeway cross hybrids had higher PEPC activity than thetwo single crosses, which is opposite to what wasobserved for GDH. In 1996/97, BR 201-F had loweractivity than the other single crosses.
The PPCh is a non-photosynthetic tissue and,therefore, the PEPC form present in this regionprobably has an anaplerotic function typical of theC3-PEPC and the root-specific formo Accordingly,values measured in these experiments are lowerthan the ones obtained for the CrPEPC in matureleaves (PURCINOet al., 1996) and seedlings (PURCINOet al., 1998). Nonetheless, they are considerablyhigher than values observed in the roots of maizeseedlings (our unpublished results).
Correlation CoefficientsTable 8 shows the correlation coefficients ob-
tained between individual kernel weight and theenzyme activities determined in the PPCh tissue at30 DAP. Kernel weight díd not correlate to GS activ-ity and PPCh soluble protein concentration in bothexperimental years. A positive correlation withGOGAT was found in 1995/96 but a non-significantvalue was observed in 1996/97. The correlationwith GDH was positive in 1994/95 but negative in1996/97. PEPC was the only enzyme activity toshow a positive correlation with kernel weight inboth years. These significant and positive correla-tion coefficients between individual kernel weightsand PEPC activities suggest that heavier kernelstended to have higher enzyme activity in the PPCh

102 A.Á.C.PCRCITO.M.R.ESILVA,S.R.M.ANDRADE,c.i. BELELE.S.N.PARENTONI,M.X.DOSSANTOS
TAl.lLE8 - Correlation coefficients betu.een rnature leernel iceigbt (rng.leernet !) and enzyme actiuity in lhe pedtcel-placento-cbalaza tissue0/ maizc bybrids at 30 days afterpollination. t n = 32 obsercations/year).
Year GS PEPCGOGAT ProteinGDH
Kernel Weight 1994/951996/97
0.22ns-D.14ns
0.47"0.06ns
0.46~0.15"
0.19n50.07ns
0.37'-D.64··
• and •• denotes sígníficance at the 0.05and 0.01 levei respectively,ns, non-significant.
than líghter kernels. Although these results wereobtained with a limited number of genotypes in twogrowing seasons and considering that we have ob-served that both kernel weight and enzyme activi-ties in the PPCh tissue were influenced by uncon-trolled environmental conditions, these significantand positive correlation coefficients certainly meritfurther investigation for confirmation. Taken togeth-er with the fact that PEPC activity in the second leafabove the main ear corre lates positively to grainyield in maize (SASAKAWAand SUGIYAMA,1989; PURCI-NO et al., 1996) these results gives rise to the hy-pothesis that PEPC plays a key role in determininggrain yield in tropical maize.
CONCLUSIONS
Application of nitrogen fertílizer did not sígnífí-cantly influence individual kernel weight, solubleprotein, GS and PEPC activities in the PPCh tissue ofthe maize genotypes used in this study in 1994/95and 1996/97. Nitrogen supply did not influenceNADH-GOGAT activity in 1994/95 but lower activi-ties were observed in the genotypes cultivated un-der high N in 1996/97. Higher GDH activities wereobserved in the PPCh tissue of genotypes cultivatedunder high N supply in 1996/97, but this favorableN effect was not significant in 1994/95. Taken to-gether these results seem to indica te that for thegenotypes used in this study, increased N supply isnot a key factor for enhancing individual kernelweight, soluble protein content and the activities ofthe N assimilating enzymes measured in this study.Therefore, as increased N supply is a major factorfor increasing grain yield of these genotypes, itseems that the beneficial N effect on yield occurs 9nparameters other than the ones studied in these ex-periments, e.g kernel number (UHARTand ANDRADE,1995b) and enzyme activity in source tissues (PURCI-NOet ai., 1996). On the other hand, genotypic díffer-ences were observed for ali enzyme activities mea-sured in this srudy, but correlation determinations
indicated that individual kernel weight was not asso-ciated with soluble protein contem and GS activityin the PPCh. Inconsistent correlations observed forkernel weight x NADH-GOGAT and kernel weight xGDH activities suggest that these relationships wereinfluenced by uncontrolled environmental condi-tions which varied between experimental years. Ir isnoteworthy, however, the fact that the correlationkernel weight x PEPC activity was positive and sig-nificant both years. Collectively, these observationssuggest that the genotypic variation for PEPC activityobserved in the PPCh tissue of the hybrids used inthis study is positively associated with individualkernel weight, i.e. genotypes with higher PEPC ac-tivity had higher individual kernel weight. Therefore,these results suggest that PEPC activity in the PPChtissue may be an useful breeding criterion for in-creasing individual kernel weight, an important com-ponent of grain yield in maize.
ACKNOWLEGMENTS- The authors are thankful 10 José MoreiraCampos and his crew for carrying our the field experiments. Ed-na Pallo Pacheco de Pinho for skillfulllaboratorial assistance. Dr.Antônio Carlosde Oliveira for the statistical analysís of the dataand Dr. MauricioAmônia Lopes for teaehing us how to obtainthe PPChtissue.This project was partiallyfunded by CNPq,grant522072-94-1to AACP.
REFERENCES
ARRUDAP.,W.J.DASILVA,1979 Aminoacid composition of vascu-lar sap of maize ear peduncle. Phytochemistry18: 409--tl0.
ARRlJDAP.,W.J.DASILVA,1983 Lysineketoglutarate reductase ac-tivity in maize. Its possible role in Iysinemetabolism of e1e-velopingenelosperm.Phytochernistry22: 2687-2689.
BLANKEM.M.,f. LENZ,1989 Fruitphorosynthesís. Piam Cell Envi-ron 12: 31-46.
BRADFORDM.,1976 A rapielanelsensitive methoelfor the quanti-tation of microgramquantities of protein utilizing the princi-pie of e1yebiding. Ana\. Bíochern.72: 248-254.
CIRILOA.G., f.H. ANDRADE,1994 Sowingdate anelmaize proeluc-tiviry:lI. Kernel nurnber deterrninatíon.Crop Sei. 34: 1044-1046.

ENZYME ACTIVITIES IN THE PPCH TISSUE
CULLYD.E., B.G. GENGENBACH,J.A. SMITH,I. RUllENSTEIN,J.A. CON-NELLY,W.D. PARK,1984 Endasperm protein synthesis and L-[3';Slmethionine incorporation in maize kernel eultured in uit-ro. Plant Physiol. 74: 389-394.
DONG L-Y., T. MASUDA, T. KAwAMURA,S. HATA, K. Izur, 1998Cloning, expression, and eharacterization of a root-forrn ofphosphoenolpyruvate carboxylase from Zea mays. cornpar-isons with the c4-form enzyme. Plant Cell Physiol. 39: 865-873.
GONzAu:s M-C., L. OSUNA,C. ECHEVARRÍA,J. VIDAL,F.J. CEjUDO, 1998Expression and loealization of phosphoenolpyruvate carboxy-lase in developing and germinating wheat grains. Plant Physi-01. 116: 1249-1258.
HAWKINSR.C., P.J.M. COOPER, 1981 Growth, development andgrain yield of maize. Exp. Agrie. 17: 203-207.
HUI'I'E H.C., D.H. TURPIN, 1994 Integratíon of earbon and nitro-gen metabolism in plant and alga I eells. Annu. Rev. PlantPhysiol. Plant MoI. Biol. 45: 577-607.
LEMCOFFJ.H., R.S. LOOMIS,1986 Nitrogen influences on yield de-termination in maize. Crop Sei. 26: 1017-1022.
LOPES M.A., 1993 Genetic and bioehemical eharacterization ofthe maize mutants floury-Z and modified opaque-ê. Ph.DThesis, The University of Arizona.
LYZNIK1.., A. RAFALSKI,K. RAcZYNSKA-BojANOWSKA,1985 Aminoacid metabolism in the pedícel-placento-chalaza region ofthe developing maize kernel. Phytochemistry 24: 425-430.
LYZNIKL.A., J. KANABuS,C.Y. TSAI, 1989 L-Glutamine and Dvglu-cose uptake by developíng endorsperms of maize. Phyto-chemistry 28: 3271-3275.
LYZNIKL.A., W. ZDROjEWSKI,M. NEUMANN,J. MACEWICZ,K. RAczYNS-KA-BOjANOWSKA,1982 A possible role of pedicel-placento-chalazal tissues in the arnino acids supply to the developingmaize endosperm. Maydica 27: 1991-198.
MISRAS., A. OAKS, 1981 Enzymes of nitrogen assimilatian duringseed development in normal and high-lysine mutants inmaize Zea mays, (W64A). Cano J. Bor. 59: 2725-2743.
MISRAS., A. OAKS, 1985 Glutamine metabolism in corn kernelscultured in vitro. Plant Physiol. 77: 520-523.
MUHITCH M., 1988 Glutamine synthetase activity of the en-dosperm, embryo and pedicel-placento-chalazal regions of de-veloping maize (Zea mays) kernels. Physiol Plant. 74: 176-180.
MUHITCHM., 1989 Purification and characterization of two formsof glutamine synthetase from the pedicel region of maize(Zea mays L.) kernels. Plant Physiol. 91: 868-875.
PORTERG.A., D.P. KNIEVEL,J. SHAJ'iNON,1987 Assimílate unload-ing from maize (Zea mays L.) pedicel tissue. lI. Effects ofchemícal agents on sugar, arnino acid, and 14C-assimilate un-loading. Piam Physiol. 85: 558-565.
PURCINOA.A.C., R. MAGNAVACA,A.T. MACHADO,I.E. MARRIEL,1994Atividade da redutase do nitrato em genótipos antigos -emodernos de milho, cultivados sob dois níveis de nitrogênio.R.Bras. Fisiol. Veg 6: 41-46.
103
PURelNOA.A.C., E. PAIVA,M.R. E SII.VA,S.R.M. DE ANDRADE,1996 In-fluenee of Azospirillum inoeulation and nitrogen supply angrain yield , and carbon- and nitrogen-assimilating enzymes inmaize. J. Plant Nutr. 19: 1045-1060.
PURCINOA.A.C., C. ARELLANO,G.S. ATHWAL,S.c. HUBER, 1998 Ni-trate effeet on carbon and nitrogen assimilating enzymes ofmaize hybrids representing seven eras of breeding. Maydiea43: 83-94.
RACZYNSKA-BojANOWSKAK., A. RAFALSKI,L. LYZNIK,1986 Nitrogentransfer into the maize kernel. pp. 159-174. In. Shannon, j.C;D.P. Knievel, C.D Boyer (eds) Regulation of earbon and ni-trogen reduetion and utilization in maize, ASPP, UniversityPark, USA.
RASTOGIR., P.S. CHOUREY,M.J. MUHITCH,1998 The maize gluta-mine synthetase GSI_2 gene is preferentially expressed in thekernel pedicels and is developmentally-regulated. Plant CellPhysiol. 39: 443-446.
RHODESD., G.A. RENDON,G.R. STEWART,1975 The control of glu-tamine symhetase levels in Lemma minor. Planta. 125: 201-211.
SINGLETARYG.W., D.C DOEHLERT,C.M. WILSON. M.]. MUHITCH,F.E.BELOW,1990 Response of enzymes and storage proteins ofmaize endosperm to nitrogen supply. Piam Physiol. 94: 858-864.
SODEK,L., W.J. DASILVA,1977 Glutamate synthase: a possible rolein nitrogen metabolism of the developing maize endosperm.Plant Physiol. 60: 602-605.
SUGIHARTO,8., K. MIYATA,H. NAKAMOTO,H. SASAKAWA,T. SUGIYAMA,1990 Regulation of expression of carbon-assimilating en-zymes by nitrogen in maize leaf. Piam Physiol. 29: 963-969.
SUKALOVICV.H-T., 1990 Properties of glutamate dehydrogenasefrom developing maize endosperm. Physiol. Plant. 80: 238-242.
TOLLENAARM., 1977 Sink-source relationships during reproduc-tive development in maize. A review. Maydica 22: 49-75.
TOLLENAARM., L.M. DWYER, D.W. STEWART,1992 Ear and kernelformation in maize hybrids representing three decades ofgrain yield improvement in Ontario. Crop Sei. 32: 432-438.
TSAI CY, 1979 Early termination of zein aecumulation in theopaque-Z mutant. Maydica 24: 129-140.
TSAI c.v., D.M. HUBER,H.L. WARREN,c.i, TSAI, 1986 Sink regula-tion of souree aetivity by nitrogen utilization. pp. 247-258. In.Shannon, J.c., D.P. Knievel, C.D Boyer (eds) Regulation ofearbon and nitrogen reduction and utilízation in maize,ASPP, University Park, USA.
UHARTS.A., F.H. ANDRADE,1995a Nitrogen deficiency in maize:I. Effects on crop growth, development, dry matter partition-ing and kernel ser. Crop Sei. 35: 1376-1383.
UHARTS.A., F.H. ANDRADE,1995b Nitrogen defieieney in maize:I!. Carbon-nitrogen interaction effects on kernel number andgrain yield. Crop Sei. 35: 1384-1389.





























