Embed Size (px)
Citation preview

ELSEVIER Journal of Experimental Marine Biology and Ecology,
201 (1996) 233-252
JOURNAL OF EXPERIMENTAL MARINE BIOLOGY AND ECOLOGY
Geratology and decomposition of Spartina alternifh-a Loisel in a New Brunswick saltmarsh
Joko Samiaji”, Felix B%rlocherb’*
‘Laboratorium Ekologi, Fak. Perikanan, Univ. Riau Jl., Simpang Panam KM 12,5, Pekanbaru, Riau, Indonesia
‘Department of Biology, Mount Allison University, Sackville, N.B., Canada
Received 4 September 1995; revised 27 November 1995; accepted 19 December 1995
Abstract
In a saltmarsh in the Bay of Fundy, New Brunswick, Spartina alterniJora had a 5 month growing period. Shoot extension was highest between July and August, and the largest number of leaves were added between June and July. Some leaves began to senesce in the first month of the
growing season, but over 50% remained alive until September. Approximately 20% of the leaves became detached from the plant between May and December. Between September and December (snowfall), such leaves lost over 60% of their mass. Compared to predried leaves in litterbags,
naturally decaying leaves accumulated more fungal species (16 vs. 9) and ergosterol (up to 125
Pg g--l vs. I50 pug g-‘), and fewer bacteria. Losses of mass, phenolics, and sugars in the first 14 days were higher in litterbag leaves.
Keywords: Decomposition; Ergosterol; Geratology; Litterbag; Saltmarsh; Standing dead
1. Introduction
In saltmarshes on the Atlantic coast of North America, the smooth cordgrass Spartina alternz)‘lora Loisel dominates the area between mean sea level and mean high water (Mann, 1982). S. aZterni$ora extends from the Gulf of St. Lawrence (latitude 46”) to Texas (latitude 27”). Average net primary productivity varies from 1000
gdrymassm-‘year-’ . m the south to a little more than half of that in the north (Mann,
1982). At optimal conditions, it can be as high as 4000 (e.g., tall Spartina growing along creeks on Sapelo Island, Odum and Fanning, 1973). This places saltmarshes among the most productive ecosystems of the world. Multiple stable isotope analyses demonstrated
*Corresponding author.
0022-0981/96/$15.00 0 1996 Elsevier Science B.V. All rights reserved
SSDl 0022-0981(95)00211-l

234 J. Sumi@, F. Biirlochrr I .I. Exp. Mar. Bid. Ed 201 (1996) 2.7.7-2-52
that much of the animal production of salt marshes is ultimately derived from cordgrass
(Peterson and Howarth, 1987; Bertness, 1992). Generally more than 90% of the Spurtina biomass is not eaten while alive (e.g., Teal,
1962). It dies and is colonized by microorganisms before it becomes available to
invertebrate feeders. Research into the decomposition of S. alterniJoru is therefore vital
for understanding the contribution of saltmarshes to the food web of coastal areas and
has attracted the attention of over 100 scientists in the last 30 years (Newell, 1993). It seems obvious that “Researchers interested in accurately describing natural microbial
participation in the decay of portions of vascular plants must try to avoid altering
genuine conditions of decay via their methods” (Newell, 1993). Unfortunately, in most studies of Spartina decomposition, questionable techniques have been used. The most serious consequence has been a drastic underestimation of fungal participation.
Traditionally, decomposition studies have relied on the litterbag technique. This involves confining dried and often preleached plant material within netting of variable
mesh size. Periodically, subsamples are recovered from the field, and mass loss and various chemical changes are monitored. Analogues of the litter bag technique were first
used by soil ecologists and date back at least 60 years (e.g., Falconer et al., 1933; for review, see Wieder and Lang, 1982). Drying and preleaching the plant material is aimed
at reducing the variability due to fluctuations in the water content or to different degrees
of preleaching by dew or rainfall. Recently, the use of dried, rather than fresh leaves, to study decomposition in streams
has been criticized by Gessner and Schwoerbel (1989). They found that leaching (rapid, abiotic loss of soluble substances) was much less pronounced in naturally shed alder and willow leaves that were not dried before being placed in water. It seems likely that the distorting effects of drying will be most pronounced when decay occurs in permanently
submerged habitats. In saltmarshes, tidal cycles ensure alternating dry/wet periods. This may retard drying of decaying Spurtinu leaves, and therefore leaching. There is no study addressing this aspect. Valiela et al. (1985) attributed an early phase (first month of
decay) of rapid weight loss to leaching. They based their conclusion on the use of predried material exposed in litter bags.
Of greater significance is the fact that S. ulternijoru, in common with most grasses,
does not abscise leaves or stems (Newell, 1993). After senescence and death of a leaf, its decay begins while still attached to the plant, in the standing-dead condition. Neverthe- less, the earliest studies of cordgrass decomposition used cut and dried leaves permanently submerged in boxes, or ground up leaves incubated in seawater with marsh sediment (Burkholder and Bornside, 1957). These conditions effectively inactivated
many fungi that may have been present, and, not surprisingly, bacteria were considered to be the main agents of decay of saltmarsh detritus. As Newell (1993) pointed out, the word ‘fungus’ was not mentioned once in Burkholder and Bomside’s 18-page paper. These early conclusions inspired a famous model of saltmarsh processes by Teal (1962), and have continued to influence the approach of saltmarsh ecologists.
An important modification was introduced by Gessner and Goos (1973). They compared the number of fungal colonies recovered from cut and dried S. altemiJEoru leaves subjected to immersion only at high tide and from leaves submerged in a creek. Material that was periodically exposed to air allowed recovery of more than twice as

.I. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ed. 201 (1996) 233-2_f2 235
many colonies. Subsequently, detailed studies revealed that over 100 higher filamentous
fungi are commonly found on decaying leaves of Spartina spp. along the east coast of North America (Boland and Grund, 1979; Gessner, 1977; Gessner and Kohlmeyer,
1976).
The first non-mycologists to attribute a major role to fungi in the decomposition of Spartina spp. leaves were Odum et al. (1979b). Their primary goal was to characterize sources of organic nitrogen on decomposing detritus. They found that chitin, presumably
originating from fungal cell walls, contributed 2550% to the non-protein nitrogen content of detritus exposed in aerobic habitats (including saltmarsh creeks and sediment). However, the involvement of fungi and bacteria was not investigated
systematically until 10 years later. Newell and Fallon (1989) and Newell et al. (1989)
compared the decomposition of dried S. alterni$ora leaves in litterbags with that of standing leaves marked with electric cable ties. Their main conclusion was that
ascomycetous fungi dominate the microbial biomass that accumulates on naturally decaying leaves (8 weeks after senescence, the ratio of bacterial to fungal biomass can
be as low as 1:600). Fungi were able to capture 90% of the nitrogen present in decaying
S. alterniflora leaves within 8-10 weeks. Estimates of fungal standing crop depend somewhat on the technique used (for
review, see Newell, 1992). With direct microscopic counts, fungal percentage of total
leaf mass was estimated to range between 4 and 33% (Newell and Hicks, 1982). With chitin as indicator molecule, these values increase to 18-44%. Neither microscopic
counts nor chitin measurements allow differentiation between living and dead mycelia. In contrast, ergosterol (a sterol typical of higher fungi) is believed to disappear rapidly
after death of the fungal cell. It has therefore become a popular indicator of living fungal
biomass. Using conservative ratios, the contribution of living fungal hyphae to total organic mass of naturally decaying S. alterni’ora leaves has been estimated to reach a
maximum of roughly 10% (Newell, 1992). The corresponding values for leaves decaying in litterbags are 5-lo-times lower (Newell and Fallon, 1989).
Thus, several studies from Georgia saltmarshes have provided convincing evidence that the use of dried material in litter bags will seriously underestimate fungal production and biomass during the decomposition of Spartina spp. leaves. It is conceivable that at
higher latitudes the natural action of ice and snow during late fall and winter simulate to some extent decay of detached leaves on the sediment or in tidal creeks. Floating sheets
of ice driven back and forth by the tides may mow down cordgrass shoots, which may subsequently be deposited on the marsh or decay while suspended in creeks or estuaries. In fact, drifting mats of dead plant material (‘wracks’) were identified as a major source of physical disturbance in spring and early summer in a saltmarsh on Rhode Island
(Bertness and Ellison, 1987). Nevertheless, one study from a saltmarsh in Maine (latitude 45”) demonstrated that approximately 66% of the yearly leaf production died
between June and October, and decay was initiated well before ice and snow were
present, and while leaves were still attached to the plant (Hardisky and Reimold, 1977). The objectives of the present study were to determine to what extent some of the
results from southern latitudes apply to a saltmarsh at higher latitudes (New Brunswick). In all cases, decomposition of attached dead leaves (natural conditions, individual plants tagged with plastic ties) were compared with dried leaves in litterbags (traditional

236 J. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ecol. 201 (1996) 233-2-52
technique). One of the majors goals was to follow fungal biomass (estimated by
ergosterol content) and bacterial numbers during the early stages of decay (from end of growing season in September, to first snow fall in December). In addition, loss of leaf area and mass were measured. Due to the build-up of snow and ice, regular sampling in
New Brunswick marshes becomes impossible after December. To estimate the removal of organic matter from the marsh before and after ice-cover, leaf mass per shoot and number of shoots per unit area of marsh were measured in fall and the following spring.
We also monitored changes in the content of sugars and phenolics of decaying leaves. These compounds are generally water soluble; their dynamics may therefore reflect how
pretreatment and exposure technique influenced leaching in S. altemijora leaves. We
expected increased leaching in predried litterbag leaves. Finally, the changes in a saltmarsh in terms of height increments, addition of new
leaves, transition from live to dead components (geratology), and loss of leaves during
the growing season were monitored. These data allow some estimate of how much of the annual leaf production decays in a standing-dead state, and therefore, how serious potential errors due to the use of the litterbag technique are.
2. Materials and methods
2.1. Study sites
The study was carried out in the Cumberland Basin, Bay of Fundy (New Brunswick, Canada). The salinity of offshore water varies between 30-33%0 and the temperature between 1 and 14°C on a yearly basis (Thomas, 1983). Tidal range can reach 16 m in
the upper bay (Mann, 1982). Decomposition and geratology studies were conducted in a saltmarsh near Fort
Beausejour (45”53’ N, 64”20’ W). The study site is relatively flat (rise of 1 m every 10 m), measures approximately 10 X 200 m, and lies within the low marsh zone (subject to
daily inundation). It is covered by Spartina altemijlora. Above this zone, dense patches of S. patens occur. No permanent tidal creeks are present, but a small pool lies at the
upper range of S. alterniforu. On each sampling occasion, the temperature of this pool and, when possible, the temperature of the incoming tidal water were measured. The two temperatures never deviated by more than 1 .S’C. Tidal range is approximately 11 m and
salinity > 22%0. Between December and April, thick, shorefast ice develops between neap and springwater levels. Other parts of the marsh are periodically affected by moving or stranded drift ice (Gordon et al., 1985). The loss of plants due to these disturbances between fall 1992 and spring 1993 was estimated at the Beausejour site as well as at a smaller marsh near the Tantramar Bridge, Sackville. This second marsh borders Tantramar River, and salinity of the water covering it is 2-4%0 lower than in the Fort Beausejour marsh.
2.2. Litterbug studies
Two experiments were conducted, one in the fall of 1992, and one in the fall of 1993. Five days before the start of the experiment, yellow-green leaf blades (excluding

J. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ecol. 201 (1996) 233-252 231
sheaths) of S. alternij?ora were collected, rinsed with sea water in the field to remove
attached clay, and rinsed more thoroughly in the laboratory with distilled water.
Air-dried leaves (5 days, 25°C) were placed in 1.5 mm mesh nylon litterbags (7 X 18
cm). This mesh size proved to be too large to retain all the leaves; in the second field
experiment, leaves were therefore tied together with nylon line before being placed in the bags.
The litterbags were exposed on 16 September, 1992, and on 20 September, 1993, at six low-marsh sites (5 bags per site, sites at 10 m intervals). The bags were attached with a 50 cm long piece of fishing line to a tent peg pushed into the sediment, and thus were
able to freely move with the tidal currents. Five randomly chosen replicates were collected after 14, 35, 63 and 91 days. In the laboratory, samples were washed carefully
with distilled water to remove sediment. Subsamples from each bag were used to estimate mass loss, fungal biomass by ergosterol measurement, fungal colonization,
bacterial numbers and changes in phenolics and sugar concentrations of the leaves.
2.3. Tagged leaves
On the same dates that litterbags were placed on the saltmarsh, 300 individual plants
of comparable size were tagged with nylon electrical cable ties immediately below the
lowest leaf that had just turned yellow-green. It was not possible to tag individual leaves, as suggested by Newell and Fallon (1989), since they did not retain the tags. The
tagging was spread over the same area where the bags were placed. On each sampling date, 50 randomly chosen leaves were collected for the various analyses.
2.4. Loss of leaf area and leaf mass
The loss of leaf area and mass was estimated using the techniques described by
Newell and Fallon (1989). On each sampling date, ten leaves (excluding leaf sheaths) were collected. The leaves were rinsed in seawater and distilled water, and cut into three
3 cm long pieces: base, tip and center. Leaf sections were photocopied and their areas
measured to the nearest mm2. They were then freeze-dried and weighed. Organic mass was determined by exposure in a muffle furnace (450°C 24 h). To determine average mass losses of entire leaves, intact leaves were collected, freeze-dried and weighed (ten
replicate leaves per sample). Their organic content was determined.
2.5. Ergosterol content of leaves
Entire leaves (standing dead or from litterbag) recovered from the field were repeatedly rinsed with distilled water to remove sediment and then freeze-dried.
Ergosterol extraction and analysis were adapted from Miller et al. (1983) and Newell et al. (1988). Preweighed amounts of leaf material were refluxed in high-performance liquid chromatography (HPLC) grade methanol (80°C 2 h), followed by refluxing of the supematant (30 min) after addition of 5 ml of a 4% solution of KOH in 95% ethanol. Sterols were partitioned into pentane, and after evaporation of the pentane, redissolved in methanol. Ergosterol content of the extract was estimated with a Gilson HPLC system

23X .I. Samiaji, F. Biirlocher I J. Exp. Mm. Biol. Ed. 201 (1996) 233-252
with a UV detector at 282 nm. Ergosterol (Fluka) dissolved in methanol served as
standard.
2.6. Microscopic examination of leaves
In the 1992 decomposition study, tagged and litterbag leaves were scanned for
reproductive fungal structures. Leaves were washed with distilled water, cut in 3 cm sections, and bleached by exposure to the following sequence of solutions (5 ml per
section, with three distilled water rinses after each treatment; Newell and Hicks, 1982): 5% NaOH in distilled water, 80°C 5 min; 30% H,O,, room temperature, 1.5 h; 0.1%
Trypan blue in lactophenol, room temperature, 2 h. The leaf sections were then stored (0-4°C) in 0.2 pm filtered seawater with 2% formaldehyde. For each sampling date, a
total of 5 randomly selected leaf sections were scanned. Fungal identification and
classification were based on Kohlmeyer and Kohlmeyer (1979, 1991).
2.7. Bacterial counts
Bacterial numbers on decaying leaves were estimated with an epifluorescence method
adapted from Newell and Hicks (1982). Leaves were rinsed in distilled water, cut into 3
cm sections, and stored in vials at 0-4°C with 0.2 km filtered seawater and 2% formaldehyde. Leaf sections were measured to the nearest mm’, homogenized with mortar and pestle, sonicated and suspended in 100 ml of autoclaved and filtered
seawater. The suspension was passed through a Nytal nylon filter (125 ,um). A subsample was passed through a black Nuclepore filter (0.2 pm), which was then stained with a 0.01% solution of acridine orange. Bacterial numbers were counted with
an epifluorescence microscope (10 replicate filters, 10 fields per filter).
2.8. Chemical analyses
Freeze-dried leaves were ground in a Wiley mill (250 pm mesh size). Five replicates of 100 mg leaf powder were extracted twice with 5 ml of 50% aqueous methanol (95°C
25 min). The extracts were combined, and the final volume readjusted to 10 ml. Phenolics were estimated with Folin-Ciocalteu’s reagent, using ferulic acid (Sigma F 3500) as standard (Rosset et al., 1982). Reducing sugars in the extract were measured
with the technique by Somogyi (1952) and Nelson (1944). Glucose served as the standard.
2.9. Saltmarsh geratology
In early spring, after ice and snow disappear, only dead material is present on the marsh surface. New shoots appear in late April. On 21 May 1993, 100 live S. altemiJlora shoots were selected at random and tagged with plastic cable ties (Reimold and Hardisky, 1977). The height of each shoot was measured to the nearest cm (from ground to tallest vegetative component, includes stem and leaves). At the same time, live (green) and dead (yellow/brown; only leaves from the current season) leaves were

.I. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ed. 201 (1996) 233-252 239
counted. These three variables were monitored at 4-week intervals for each of the tagged
plants during the growing season.
2.10. Total mass loss between September and spring
On 15 September 1992 (beginning of decomposition experiment), and on 5 May 1993
(shortly after all the ice had melted and the sites became accessible) the number of individual S. altemzjlora shoots per unit area was counted (15 replicates, 30 X 30 cm squares). At the same time, 60 plants were harvested. Their height was determined. After
rinsing the plants in distilled water, all leaves were detached and freeze-dried, and their
ash-free dry mass was determined.
2.1 I. Statistical analysis
Analyses were done with the SYSTAT 5.1 program (Wilkinson, 1989). Values for
percentage mass remaining, and phenolics and sugar concentrations were fitted to an
exponential decay model (linear regression after In transformation, days as time units; means+SE). The slopes of decay curves were compared with analysis of covariance
(ANCOVA), following the procedures by Snedecor and Cochran (1967). Remaining
mass, phenolics and sugar concentrations of standing dead and litterbag leaves after 14
days exposure (when leaching losses would be most pronounced) were compared with t-tests.
3. Results
In both years, temperature of a tidal pool at the study site declined linearly from 1.5 to 0°C between the first sample in September and the last in December.
Near the end of the 1992 growing season (15 September), shoot density was significantly higher in the Fort Beausejour Marsh (630+70 plants m-2) than at the
Tantramar Bridge marsh (338290 plants mp2; t-test, P < 0.01). By the following May,
the density had dropped to 355+90 (Fort Beausejour) and 186250 (Tantramar Bridge), a loss of almost 50%. On the other hand, plants were significantly taller in the Tantramar
Bridge marsh (43.428.9 cm) than in the Beausejour marsh (26.228.3; P -=c 0.01). By the following spring, the average heights of the remaining shoots were 31.0?7.9 (Tan-
tramar) and 12.3k2.8 cm (Beausejour). Average number of leaves per shoot in September 1992 was higher in the Tantramar
marsh (5.520.3) than in the Beausejour marsh (4.5kO.3; P < 0.01). By spring, these numbers had dropped to 2.42 1.6 and 1.420.9, respectively. Similarly, average leaf biomass (AFDM in g) per shoot was higher in the Tantramar marsh (0.30?0.07) than in
the Beausejour marsh (0.2720.06; PcO.01). By 5 May, these values had declined to 0.067+0.037 and 0.052+0.020, respectively. Thus, the average loss of leaf mass in plants that were not removed between May and September was 80.7% (Beausejour) and
77.6% (Tantramar). Overall, 89.7% (Beausejour) and 87.6% (Tantramar) of the leaf mass present in
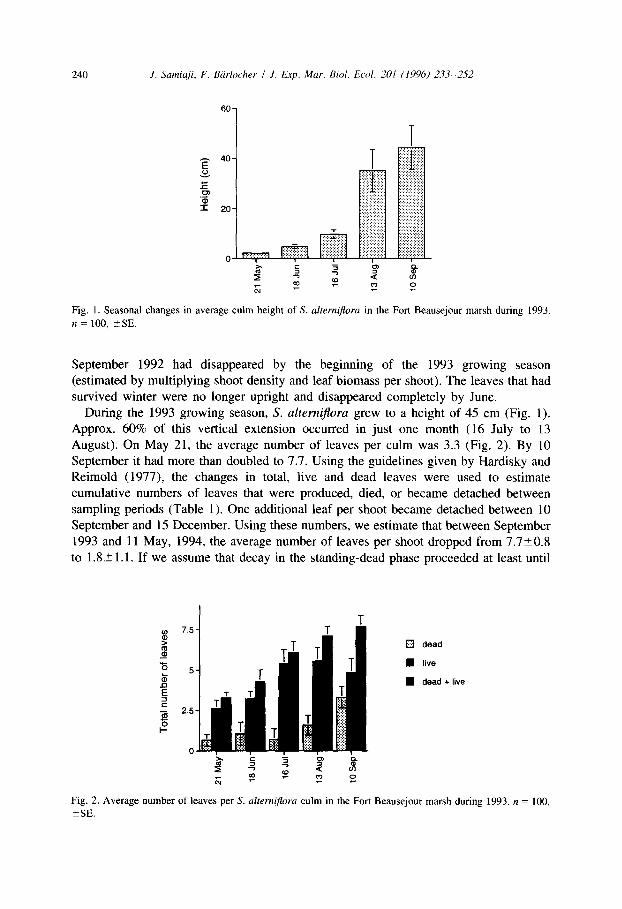
240 J. Samiaji, F. Biirlocher I J. Exp. Mar. Bid. Ecol. 201 (1996) Zi’.3-2.52
Fig. 1. Seasonal changes in average culm height of S. alferniflora in the Fort Beausejour marsh during 1993.
n = 100, +SE.
September 1992 had disappeared by the beginning of the 1993 growing season
(estimated by multiplying shoot density and leaf biomass per shoot). The leaves that had
survived winter were no longer upright and disappeared completely by June. During the 1993 growing season, S. altemi@-a grew to a height of 45 cm (Fig. 1).
Approx. 60% of this vertical extension occurred in just one month (16 July to 13 August). On May 21, the average number of leaves per culm was 3.3 (Fig. 2). By 10
September it had more than doubled to 7.7. Using the guidelines given by Hardisky and Reimold (1977), the changes in total, live and dead leaves were used to estimate cumulative numbers of leaves that were produced, died, or became detached between
sampling periods (Table 1). One additional leaf per shoot became detached between 10 September and 15 December. Using these numbers, we estimate that between September
1993 and 11 May, 1994, the average number of leaves per shoot dropped from 7.750.8
to 1.8 + 1.1. If we assume that decay in the standing-dead phase proceeded at least until
I 7.5
zl fl dead a, B n live
t 5
2 n dead + live
z 3 2.5
ij I-
Fig. 2. Average number of leaves per S. alter@ora culm in the Fort Beausejour marsh during 1993. n = 100,
&SE.

.I. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ecol. 2001 (1996) 233-252 241
Table 1 Cumulative numbers of leaves per culm of S. altemijloru that were produced, died or became detached during
the 1993 growing season (n = 100, 2%)
to 21 May May-June June-July July-August August-September
New 3.2 4.4 6.2 7.2 7.8
Dead 0.7 1.1 1.1 2.2 4.0 Detached _ 0.4 0.4 0.4
December, we can estimate the average time a leaf was decaying while still attached. It
is approximated by the area under a straight line connecting dead leaves in Fig. 2 (product of number of dead leaves and time) divided by total number of leaves
produced. This assumes that the decay of any detached leaf occurred entirely after detachment. Despite this conservative assumption, the numbers suggest that, on the average, the initial period of decomposition in an attached phase lasted 2 months.
In 1992, but not in 1993, litterbag leaves decayed more rapidly than standing-dead
leaves (Fig. 3; comparison of regression slopes by ANCOVA after In transformation of data; P = 0.025 in 1992, 1993: 0.15 in 1993). Decay coefficients k (days as time units) were - 0.011 for naturally decaying leaves in both years, and - 0.019 and - 0.013 for
litterbag leaves in 1992 and 1993. After 14 days, remaining AFDM was significantly lower in litterbags than in standing dead leaves (z-test, P < 0.002).
1992
EI Standing dead
25 ';; 2
z 0
2
F 1993 .- .s
E 100
B 75
25
14 35 $3 91
Time (days)
Fig. 3. Changes in ash-free dry mass of entire S. alternifbra leaves in the Fort Beausejour marsh. Day 0 = 16
September 1992 and 20 September 1993. Standing dead: plants were tagged with electrical cable ties;
Litterbag: leaves were cut, dried and exposed in nylon litterbags. n = 10, tSE.

242 J. Samiqji, F. Biirlocher I J. Exp. Mar. Bid. Ed. 201 (1996) 233-2.52
The results of decay experiments with leaf sections (tip, middle and base) are summarized in Table 2. Daily decay coeffcients k, estimated after In transformation of the data, were analyzed with 3-factorial ANOVA, with the data from the 2-year serving as replicates. All three factors were significant: exposure technique (standing dead,
litterbag; P = 0.026), parameter measured (leaf area or mass; P < O.OOl), and leaf section (tip, middle, base; P < 0.001). No interaction between the three factors was
significant (P 2 0.28). The ergosterol content of standing dead leaves increased steeply between 14 and 35
days of exposure; in litterbag leaves, the increase was delayed and less steep (Fig. 4).
Both time and exposure technique had a highly significant effect on ergosterol content (P 5 0.001). Ergosterol values declined after 91 days; this trend apparently continued through the winter. On April 14, 1993, the average ergosterol content of leaves still
attached to the plant was 25.920.8 pg g-’ (&SE, n = 10).
In 1992, reproductive structures of 16 species of marine fungi (11 Ascomycetes, 1 Basidiomycete, 4 Deuteromycetes) were identified on naturally decaying S. alterni$oru
leaves (Table 3). 10 species were found on leaves in litterbags. Throughout the study,
Table 2
Summary of daily loss rates of area or ash-free dry mass (AFDM) of S. alternifora leaf sections (Base, Center,
Tip)
Year Leaf section Measured Exposure k R
1992 Base Area SD -0.002 0.974
LB -0.002 0.997
AFDM SD -0.005 0.978
LB -0.006 0.965
Center Area SD -0.002 0.993
LB -0.002 0.988
AFDM SD -0.006 0.996
LB -0.006 0.995
Tip Area SD -0.005 0.987
LB -0.006 0.934
AFDM SD -0.008 0.999
LB -0.010 0.999
1993 Base Area SD -0.002 0.994
LB -0.002 0.977
AFDM SD -0.006 0.994
LB -0.006 0.969
Center Area SD -0.002 0.944
LB -0.003 0.929
AFDM SD - 0.004 0.994
LB -0.005 0.987
Tip Area SD -0.005 0.970
LB -0.005 0.985
AFDM SD -0.007 0.984
LB -0.009 0.987
Standing dead (SD): plants tagged with electrical cable ties; Litterbag (LB): leaves were cut, dried and
exposed in nylon litterbags. k = decay coefficient of In transformed data, time in days. R = correlation coefficient.

.I. Samiaji, F. B&ocher I .I. Exp. Mar. Biol. Ecol. 201 (1996) 233-252 243
Standing dead
Litterbag
s 0 14 35 63 91
& iii $ 15 150 1 1993
0 14 35 63 91
Time (days)
Fig. 4. Ergosterol content (pg g-’ AFDM) of decaying 5’. alternijlora leaves. Same symbols as in Fig. 3.
n = StSE.
Table 3
Fungal species reproducing on S. alferniflora leaves decaying in the Fort Beausejour marsh between
September and December 1992
Class Species Standing
dead
Litterbag
Ascomycetes Buergenenda spartinae Kohlm. and Gessner
Halosarpheia spartinae (Jones) Shearer and Crane
Halosphaeria hamata (HGhnk) Kohlm.
Leptosphaeria neomaritima Gessner and Kohlm.
L. oraemaris Linder
Lulworthia sp.
Passeriniella obiones (Crouan and Crouan)
Hyde and Mouzouras
Basidiomycetes
Deuteromycetes
Phaeosphaeria spartinae (Ellis and Everhart)
Shoemaker and Babcock
Ph. spartinicola Leuchtman
Pleospora pelagica Johnson
Pl. spartinae (Webster and Lucas) Apinis and Chesters
Nia vibrissa Moore and Meyers
Dendryphiella arenaria Nicot
Drechslera halodes (Drechsler) Subramanian and Jain
Phoma sp.
Stagonospora sp.
+ + + + + + + _ + _ + + _
4
+ + + + + + + + + + + + + + ._

244 .I. Samiaji, F. Btirlocher I J. Exp. Mar. Biol. Ecol. 201 (19%) 233-25.2
q Standing dead
n Litterbag
14 35 63 91
3 .s $ 5 c%
2
0 0 14 35 63 91
Time (days)
Fig. 5. Bacterial counts (per g AFDM) on decaying S. altemijlora leaves. Same symbols as in Fig. 3.
n = 1O”SE.
standing dead leaves carried more fungal species than leaves in litter bags. The largest number of species were found on day 63 (14 species on 5 sections of standing dead, 8
on litterbag leaves). In the final sample (day 91), the numbers declined to 6 and 4,
respectively. Buergenerula spartinae, Halosarphia spartinae, Halosphaeria hamata,
Phaeosphaeria spartinicola and Phoma sp. were present on 2 80% of all samples. Bacterial counts increased linearly throughout the study period (Fig. 5; for standing
dead leaves, regression slope b = 0.018 X lo9 in both years; for litterbag leaves, b = 0.026 X lo9 in 1992, b = 0.023 X lo9 in 1993). In 1993, slopes between standing dead and litterbag leaves were significantly different (ANCOVA; P = 0.03; in 1992,
P = 0.17).
Both phenolics and sugar contents decreased during decay (Fig. 6 and Fig. 7). In the
first field sample of 1992 (after 14 days of exposure), no difference was found in the phenolics content of standing dead and litterbag leaves (t-test, P = 0.36). In the other three cases, values were higher in standing dead leaves (phenolics 1993, P = 0.04;
sugars 1992, P = 0.002; sugars 1993, P = 0.03). When all samples are considered, sugars disappeared more rapidly in 1992 from litterbag than from standing dead leaves (ANCOVA after In transformation; sugars 1992, P = 0.036); no significant differences were found in the other cases (sugars 1993, P = 0.16; phenolics 1992, P = 0.24; phenolics 1993, P = 0.31).
4. Discussion
The shoot elongation and leaf production data show that S. alternijlora in the Fort

J. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ecol. 201 (1996) 233-252 24s
q Standing dead
0 14 35 63 91
Time (days)
Fig. 6. Phenolics content (ferulic acid equivalents) of decaying S. alremiflora leaves. Same symbols as in Fig.
3. n = S+SE.
0.6
0.4
0.2
q Standing dead
n Litterbag
4 0 14 35 63 91
E
LI 1993
0 1’4 35 s3 91
Time (days)
Fig. 7. Sugar content (glucose equivalents) of decaying S. alteniflora leaves. Same symbols as in Fig. 3.
n = 5fSE.

246 J. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ecol. 201 (1996) 233-252
Beausejour marsh had essentially a 5 month growing period (May-September). This agrees with a previous study on eight other saltmarshes in the same basin (Gordon et al.,
1985). Maximum average shoot height was 45 cm (Fig. l), which again falls within the range found in other marshes of the Cumberland basin (38-48 cm, Gordon et al., 1985) but is lower than values reported from Rhode Island (average stem height 39-l 14 cm, Bertness, 1985), Georgia and Louisiana (up to 200 cm; Odum and Fanning, 1973;
Latham et al., 1991). Primary production per unit area of saltmarshes in the Bay of Fundy is therefore rather low; nevertheless, it has been estimated that it contributes
approximately half of the carbon fixed photosynthetically in the Cumberland Basin, because the high turbidity of the water limits phytoplankton productivity (Gordon et al.,
1985). Shoot extension was highest between July and August, while most of the leaves were
added between June and July. By comparison, Gordon et al. (1985) found a peak of net
aerial primary production in early July, while Hardisky and Reimold (1977) observed a distinct decline of leaf production after June in a Maine saltmarsh.
In the Fort Beausejour marsh, average height was considerably lower at the end of the
1992 growing season (26.2 vs. 45 cm in 1993) as was the average number of leaves per culm (4.5 vs. 7.5). This indicates that year-to-year variation of standing crop and, by implication, of production is quite high.
Senescence and death of newly produced leaves were initiated very early in the
season: on May 21, approx. 25% of all leaves were dead (Fig. 2). This percentage increased to 44% by September 10 (a decline in July was presumably due to the loss of
entire leaves). From the beginning of the growing season until December, approx. 1.4
leaves per shoot, or 17.9% of the 1993 production, became detached. Estimates from three other saltmarshes in the Cumberland Basin range from 6.8 to 16.7% (Gordon et al.,
1985). Plants that were not sheared off by ice and snow, retained between 30 and 44% of the leaves through the winter. By comparison, only 3% of tagged leaves were lost
during 137 days in a Georgia saltmarsh (Newell and Fallon, 1989). Leaves that remained attached (corresponding to > 80% of the yearly production) lost
over 60% of their ash-free dry mass between September and December (Fig. 3). The
inevitable conclusion is that standing dead leaves have to be used if one wishes to accurately describe the predominant early phase of decomposition in saltmarshes in the Bay of Fundy. Litterbags containing fungal-colonized leaves, or particles prepared from such leaves, will be suitable for investigating the second phase of decomposition.
The mass loss of 50% in 63 days in the present study compares to roughly 25-40% in the 50-60 days in a large number of other studies (Montagna and Ruber, 1980; Valiela et
al., 1985; Pozo and Colino, 1992; Reice and Stiven, 1983). Presumably, the extreme tidal conditions in the Bay play a role, and may accelerate mechanical fragmentation after microbial weakening of the leaf’s structures (average tidal currents are 0.7 m s- ’ ; Gordon et al., 1985). During winter (i.e., when the leaves were in close contact with the sediment, which may be adequately represented by litterbags), another 17% were lost. Gordon et al. (1985) who determined loss of the entire biomass (i.e., leaves, leaf sheaths and stems), estimated that roughly 45% of the current year’s production was lost before winter, another 22.5% during winter, and the rest in the following spring and summer.

J. Samiaji, F. Biirlocher I .I. Exp. Mar. Biol. Ecol. 201 (1996) 233-252 247
Leaf mass was consistently lost more quickly than leaf area (Table 2). This suggests
that large leaf-shredding invertebrates did not make a major contribution to decomposi-
tion. These animals quickly skeletonize leaves (leading to a rapid loss of leaf area);
feeding by the saltmarsh periwinkle Lirtorariu irroruru results in typically frayed leaves (Newell, 1993). This was not observed here, nor were any macroinvertebrates found feeding on dead leaves.
Both mass and area declined most rapidly in leaf tips (Table 2), the oldest parts of the leaves, which reflects the progression of senescence: the yellowing of the leaf and the
appearance of visible fungal structures generally begin at the tip. Again, this suggests that microbial activity was instrumental in controlling overall loss, either directly or by
increasing the likelihood of mechanical fragmentation.
Both area and mass were generally lost more rapidly in detached leaves than in naturally decaying leaves (Fig. 3, Table 2), though when taken over the course of the
study, the differences were small. However, as pointed out by Newell and Fallon ( 1989) this does not validate the litterbag technique, since similar losses might be brought about
by different patterns of decomposer activities. The present study suggests that this is indeed the case. The concentration of ergosterol (an indicator molecule for living fungal
cells; Newell, 1992) was 2-4 times higher in naturally decaying leaves than in litterbag
leaves (Fig. 4) while bacterial numbers were consistently higher on litterbag leaves (Fig. 5). Average ergosterol concentrations reached a maximum of 125 pg g- ’ AFDM of decomposing attached leaves. This compares to values of 462 (Newell and Fallon,
1989) or 359 ,ug gg’ (Newell et al., 1989) of tagged leaves in a Georgia saltmarsh. Cut
leaves in litterbags in our study accumulated a maximum of 59 pug g-’ in the 1993 experiment. A similar value (5 1) was found by Lee et al. (1980) in litterbagged leaves of a Massachusetts marsh, while in Newell and Fallon (1989) the corresponding values
were 33.6 (winter sample) and 90.6 ,ug g-’ (May sample). Thus, at least qualitatively, cutting leaves and placing them in litterbags had the same effect in this study as in Georgia saltmarshes: it resulted in a shift from fungal to bacterial biomass. There are
some distinct differences, however. To begin with, the differences, both for fungi and bacteria, were smaller in this study (ergosterol ratio of bagged to tagged leaves was
0.5-0.25, compared to 0.25-o. 11; ratio of bacterial numbers 1.2 compared to 4-67).
Possibly, the effects were less drastic because of details of the technique and
peculiarities of Bay of Fundy marshes. Leaves were air-dried, which presumably
allowed survival of fungal inocula. The litterbags were attached to 50 cm long strings, allowing them to move with the current. They rarely became covered with sediment; thus anaerobic conditions were generally avoided (anaerobiosis is believed to be the main factor regulating fungal colonization of sediment-covered Spartinu sp. leaves; Wagner, 1965, 1969). Finally, it is unlikely that the supply of water between standing dead and bagged leaves would differ as drastically as in a Georgia saltmarsh. Air
temperature was low through most of the experimental period; during unusually warm
days, fog formation is very common. A second striking difference is the much larger average ergosterol content of
standing-dead leaves decaying in Georgia marshes (roughly 3-4 times as high as in this study). Visual comparison of decaying Spurtinu spp. leaves suggests that the difference is real (Newell, 1993, observed lower densities of ascomata on standing-dead leaves in

248 .I. Sumiaji, F. Bdocher I J. Exp. Mar. Biol. Ecol. 201 (1996) 233-252
Maine and Ireland; Barlocher, pers. comm., standing-dead leaves with very high
densities of ascomata are more common in Georgia marshes than in New Brunswick marshes). Nevertheless, it is unlikely that the values reported here represent true
maxima. Highest averages were reached after 9 weeks of decomposition, with the two
closest samples taken 4 weeks before and after. It is possible that higher values would have been found within this S-week period. In addition, the values represent averages of
entire leaves; there were clearly visible differences among individual leaves, and
between leaf sections. The highest ergosterol value of randomly pooled intact leaves was
200 pug gg’; in 1994 samples of leaf tips, values above 300 were not uncommon, and a maximum of 452 was observed.
It is difficult to convert ergosterol values into fungal biomass, since many factors influence this ratio (Newell, 1992; Newell et al., 1987). Among these are extraction
techniques (e.g., Padgett and Posey, 1993; Newell et al., 1988; Newell, 1992); growth
conditions (Newell et al., 1987; Gessner and Chauvet, 1993); physiological state and age
of mycelium (Gessner and Chauvet, 1993; Newell, 1992). Using conservative ratios for marine fungi by Newell et al. (1987), the ergosterol content in the present study would
correspond to an average fungal mass of 2.5-3.4% of decaying leaf mass. One potentially important factor is the relatively low temperature at which decay occurred. In a mesophilic Mm-or sp., ergosterol content increased by 70% when incubation
temperature was raised from 0 to 20°C (Dexter and Cooke, 1984). The effect of
temperature on ergosterol content of marine fungi is currently under investigation. By the following spring, ergosterol content of leaves had declined to about 25% of the
fall maximum. This indicates either that fungi do not survive well or become dormant when buried under ice or snow, or that leaves (leaf sections) with high fungal content
were preferentially removed by tidal currents.
More species of marine fungi colonized and reproduced on standing dead leaves than bagged leaves. Gessner (1977) found a similar pattern in a Rhode Island salt marsh. He
identified 14 marine fungi on naturally decaying leaves through direct microscopic examination, compared to nine species from litterbag leaves. In a study in the United
Kingdom, 17 marine fungi were found on S. townsendii leaves (Apinis and Chesters, 1964).
The number of fungal species in this study peaked in November, coinciding with maximum ergosterol content. Kohlmeyer and Kohlmeyer (1979) concluded that fungal
colonization of Spartina sp. leaves is concentrated between September and January, i.e., during seed formation, senescence and death of the host.
Five marine fungi were found throughout the study. In a survey of a Rhode Island
estuary (Gessner, 1977), four species, namely B. spartinae, H. hamatu, P. spartinicolu
and Stugonosporu sp., were present throughout the growing season. Dead culms were mostly inhabited by L. obiones, L. pelagica, Lulworthia sp., P. pelagica and Pleospora
vagans. With the exception of P. vagans, all species were also found in this study.
One objective of the study was to estimate leaching, i.e., rapid, abiotic removal of soluble compounds, and how it is influenced by cutting and predrying leaves. Mass loss of litterbag leaves was highest during the first 14 days (Fig. 3), and exceeded mass loss of naturally decaying leaves by 28 (1992) and 36% (1993). Similarly, losses of sugars and phenolics after 14 days were higher in predried leaves (Fig. 6 and Fig. 7; not

.I. Samiaji, F. Biirlocher 1 J. Exp. Mar. Biol. Ecol. 201 (1996) 233-252 249
statistically significant with phenolics in 1992). This suggests that the early rapid mass
loss, which is generally attributed to leaching (Valiela et al., 1985), is indeed to some
extent an artifact of unnatural pretreatment of the substrate. Subsequent changes in
phenolic and sugar concentrations reflect the balance between abiotic and biological processes: microorganisms use carbohydrates and phenolics as energy sources, and may release them during their extracellular attack of polysaccharides and lignins (Bergbauer and Newell, 1992; Newell, 1993). There is no obvious way to predict which of these processes will dominate.
In many detritus-dominated ecosystems, microbial colonization is closely followed by invertebrate attack (B;irlocher, 1992; Newell, 1993). For example, Littoruria irroruta
(Say) selectively removes living fungal mass from standing-dead S. alterni$Ioru leaves
(Newell and Bailocher, 1993). We saw no evidence that this plays any role in the Fort
Beausejour marsh. The only snail found in substantial numbers was Littorinu suxutilis
Olivi, and its abundance on the marsh declined drastically between September and
October (unpublished obs.). It was generally found on the sediment, and we did not observe any obvious feeding marks on Spurtinu detritus. Because of high tidal currents
(average of 0.7 m ss’; Gordon et al., 1985), it also seems unlikely that much feeding
occurred when the marsh was submerged. The possibility remains that more cryptic shredders are involved (Newell, 1993). These might include harpacticoid copepods,
collembolans, or amphipods. We never saw any of these invertebrates in collected bags or leaves, though a tidal pool on the marsh does contain Gummarus mucronafus Say
(Barlocher and Howatt, 1986).
This raises the question of how relevant Spurtinu is to the food web in the Cumberland Basin. Available evidence suggests that it contributes roughly 50% of total
primary production (Mann, 1982; Gordon et al., 1985). In contrast to Georgia marshes, however, much of this production is not consumed in situ. Instead, it is exported and
widely distributed about the estuary. Once dislodged, vegetation appears to leave the
marsh. Odum et al. (1979a) concluded that estuarine geomorphology plays a major role in controlling the flux of particulate matter between wetlands and water. They predict the
greatest net export to occur from exposed marshes in basins that gradually deepen and widen toward their mouths. Many marshes in the Cumberland Basin fit this description.
The prediction by Odum et al. (1979a) is supported by direct observations showing that saltmarsh detritus accounts for the largest portion of non-living suspended organic
matter in the Cumberland Basin (Schwinghamer et al., 1983). Fine Spurtinu detritus is believed to make an important contribution to the diet of Corophium volututor (Pallas) (Stuart et al., 1985, but see Murdoch et al., 1986). This tube-dwelling amphipod is the dominant macro-invertebrate in intertidal mudflats (up to 60,000 mm’), and forms a
major part in the diet of ground-feeding fish and migrating shorebirds (Hicklin and Smith, 1979). Its major growth occurs between June and September (Murdoch et al., 1986), and any Spurtina particles it ingests may have been colonized by fungi in the
preceding autumn and subsequently further processed in the water column. In conclusion, the fate of dead Spurtinu leaves in the Bay of Fundy differs
significantly from that in Georgia. In both cases, decay is initiated by fungi in the standing-dead phase. In the Bay of Fundy, these losses are probably accelerated by physical fragmentation due to strong tidal currents; in Georgia, shredding invertebrates

250 .I. Samiaji, F. Bdocher I .I. Exp. Mar. Biol. Ecol. 201 (1996) 233-252
speed up the process. The resulting particles or intact detached leaves are widely dispersed in the Bay of Fundy; in Georgia, they are further broken down within the marsh (Newell, 1993). Roughly l/3 of the annual leaf production is pressed onto the sediment by ice and snow in the Bay of Fundy; no comparable process exists in the
south. Despite these differences, Newell’s criticism (Newell, 1993) of the litterbag technique to study the early stages of decomposition of S. alrevrziJIora is as relevant in
New Brunswick as it is in Georgia.
Acknowledgments
This research has been financed by the Marine Sciences Education Project of the Department of Education and Culture, Republic of Indonesia, and by an Operating Grant to FB from the Natural Science and Engineering Research Council of Canada.
References
Apinis, A.E. and C.G.C. Chesters, 1964. Ascomycetes of some salt marshes and sand dunes. Trans. Br. Mycol. sot., Vol. 47, pp. 419-43s.
Badocher, F. (editor), 1992. The ecology ofaquatic hyphomycetes. Ecological Studies 94, Springer, Berlin, 225
PP. Bariocher, F. and S.L. Howatt, 1986. Digestion of carbohydrates and protein by Gammarus mucronatus Say
(Amphipoda). /. Exp. Mar. Biol. Ecol., Vol. 104, pp. 229-237. Bergbauer, M. and S.Y. Newell, 1992. Contribution to lignocellulose degradation and DOC formation from a
saltmarsh macrophyte by the ascomycete Phaeosphaeria spartinicola. FEMS Microbid. Ecol., Vol. 86, pp.
3341-348.
Bertness, M.D., 1985. Fiddler crab regulation of Spartina alternijlora production on a New England salt
marsh. Ecology, Vol. 66, pp. 1042-1055.
Bertness, M.D., 1992. The ecology of a New England salt marsh. Am. Sci., Vol. 80, pp. 260-269.
Bertness, M.D. and A.M. Ellison, 1987. Determinants of pattern in a New England salt marsh plant
community. Ecol. Monogr., Vol. 57, pp. 129-147.
Boland, G.J. and D.W. Grund, 1979. Fungi from the salt marshes of Minas Basin, Nova Scotia. Proc. N.S. Inst. Sci., Vol. 29, pp. 393-404.
Burkholder, P.R. and G.H. Bomside, 1957. Decomposition of marsh grass by aerobic marine bacteria. Bull. Tow. Bot. Club,Vol. 84, pp. 366-383.
Dexter, Y. and R.C. Cooke, 1984. Fatty acids, sterols and caretonoids of the psychrophile Mucor strictus and
some Mucor species. Trans. Br. Mycol. Sot., Vol. 83, pp. 455-461.
Falconer, G.J., J.W. Wright and H.W. Beall, 1933. The decomposition of certain types of fresh litter under field
conditions. Am. J. Bot., Vol. 20, pp. 196-203.
Gessner, M.O. and E. Chauvet, 1993. Ergosterol-to-biomass conversion factors for aquatic hyphomycetes.
Appl. Environ. Microbial., Vol. 59, pp. 502-507.
Gessner, M. and J. Schwoerbel, 1989. Leaching kinetics of fresh litter with implications for the current concept
of leaf processing in streams. Arch. Hydrobiol., Vol. 115, pp. 81-90.
Gessner, R.V 1977. Seasonal occurrence and distribution of fungi associated with Spartina alternif7ora from a
Rhode Island estuary. Mycologia, Vo1.69, pp. 477-491.
Gessner, R.V. and R.D. Goes, 1973. Fungi from decomposing Spartina alternijlora. Can. J. Bot., Vol. 51, pp.
5 l-55.
Gessner, R.V and J. Kohlmeyer, 1976. Geographical distribution and taxonomy of fungi from salt marsh Spartina. Can. J. Bot., Vol. 54, pp. 2023-2037.

.I. Samiaji, F. Biirlocher I J. Exp. Mar. Biol. Ecol. 201 (1996) 233-252 251
Gordon, D.C. Jr., D.J. Cranford and C. Desplanque, 1985. Observations on the ecological importance of salt
marshes in the Cumberland Basin, a macrotidal estuary in the Bay of Fundy. Es&w. Coast. She/f Sci., Vol.
20, pp. 205-227.
Hardisky, M. and R. Reimold, 1977. Salt-marsh plant geratology. Science, Vol. 198, pp. 612-614.
Hicklin, P.W. and PC. Smith, 1979. The diets of five species of migrant shorebirds in the Bay of Fundy. Proc.
N.S. Inst. Sci., Vol. 29, pp. 483-488.
Kohlmeyer, J. and B.V. Kohlmeyer, 1991. Illustrated Key to the filamentous higher marine fungi. Bat. Mar.,
Vol. 34, pp.l-61.
Kohlmeyer, J. and E. Kohlmeyer, 1979. Marine Mycology. Academic Press, New York, NY, 690 pp.
Latham, P.J., LG. Pearlstine and W.M. Kitchens, 1991. Spatial distributions of softstem bulrush, Scirpus
validus, across a salinity gradient. Estuaries, Vol. 14, pp. 192-198.
Lee, C., R.W. Howarth and B.L. Howes, 1980. Sterols in decomposing Spnrrina alternij?ora and the use of
ergosterol in estimating the contributions of fungi to detrital nitrogen. Limnol. Oceanogr., Vol. 25. pp.
290-303.
Mann, K.H., 1982. Ecology of coastal waters. Studies in Ecology, Vol. 8. University of California Press,
Berkeley, CA, 322 pp.
Miller, J.D., J.C. Young and J.L. Trenholm, 1983. Fusarium toxins in field corn. I. Time course of fungal
growth and production of deoxynivalenol and other mycotoxins. Can. J. Bot., Vol. 61, pp. 3080-3087.
Montagna, P.A. and E. Ruber, 1980. Decomposition of Sparfina alfernij?ora in different seasons and habitats of
a northern Massachusetts salt marsh and a comparison with other atlantic regions. Estuaries, Vol. 3, pp. 61-64.
Murdoch, M.H., F. BIrlocher and M.L. Laltoo, 1986. Population dynamics and nutrition of Corophium
volururor (Pallas) in the Cumberland Basin (Bay of Fundy). J. Exp. Mar. Biol. Ecol.,Vol. 103, pp. 235-249.
Nelson, N., 1944. A photometric adaptation of the Somigyi method for the determination of glucose. J. Biol.
Chem., Vol. 153, pp. 375-380.
Newell, S.Y., 1992. Estimating fungal biomass and productivity in decomposing litter. In, The ,fungal
community, edited by G.C. Carroll and D.T. Wicklow, Marcel Dekker, New York, NY, pp. 521-561.
Newell, S.Y., 1993. Decomposition of shoots of a salt-marsh grass. Methodology and dynamics of microbial
assemblages. Adv. Microb. Ecol., Vol. 13, pp. 301-326.
Newell, S.Y. and F. Bklocher, 1993. Removal of fungal and total organic matter from decaying cordgrass
leaves by shredder snails. J. Exp. Mar. Biol. Ecol., Vol. 171, pp. 39-49.
Newell, S.Y., T.L. Arsuffi and R.D. Fallon, 1988. Fundamental procedures for determining ergosterol content
of decaying plant material by liquid chromatography. Appl. Environ. Microbial., Vol. 54, pp. 1876-1879.
Newell, S.Y., J.D. Miller and R.D. Fallon, 1987. Ergosterol content of salt-marsh fungi: effect of growth
conditions and mycelial age. Mycologia, Vol. 79, pp. 688-695.
Newell, S.Y., J.D. Miller and R.D. Fallon, 1989. Decomposition and microbial dynamics for standing, naturally
positioned leaves of the salt-marsh grass Spartina altemijora. Mar. Biol., Vol. 101, pp. 471-481.
Newell, S.Y. and R.D. Fallon, 1989. Litterbags, leaf tags and decay of non-abscised intertidal leaves. Can. J.
Bot., Vol. 67, pp. 2324-2327.
Newell, S.Y. and R.E. Hicks, 1982. Direct count estimates of fungi and bacterial biovolume in dead leaves of
smooth cordgrass (Spartina alrernifora Loisel). Esfuaries, Vol. 5, pp. 246-260.
Odum, E.P. and M.E. Fanning, 1973. Comparisons of the productivity of Spartina alterni~orcz and S.
cynosuroides in Georgia coastal marshes. Bull. Georgia Acad. Sci., Vol. 31, pp. I- 12.
Odum, W.E.. J.S. Fisher and J.C. Pickral, 1979a. Factors controlling the flux of particualte organic carbon from
estuarine wetlands. In, Ecological processes in coastal and marine ecosystems, edited by R.J. Livingston,
Plenum Press, New York, NY, pp. 69-80.
Odum, W.E., P.W. Kirk and J.C. Zieman, 1979b. Non-protein compounds associated with particles of vascular
plant detritus. Oikos, Vol. 32, pp. 363-367.
Padgett. D.E. and M.H. Posey, 1993. An evaluation of the efficiencies of several ergosterol extraction
techniques. M~ol. Rex., Vol. 97, pp. 1476-1480. Peterson, B.J. and R.W. Howarth, 1987. Sulfur, carbon and nitrogen isotopes used to trace organic matter flow
in the salt marsh estuaries of Sapelo Island, Georgia. Limnol. Oceanogr., Vol. 32, pp. 1195-1213.
Pozo, J. and R. Colino, 1992. Decomposition processes of Spartina mar&ma in a salt marsh of the Basque
Country. Hydrobiologia, Vol. 231, pp. 165-175.

252 .I. Sumiaji, F. Biirlocher I J. Exp. MIX. Biol. Ecol. 201 (1996) 233-252
Reice, S.R. and A.E. Stiven, 1983. Environmental patchiness, litter decomposition and associated fauna1
patterns in a Spartina alterniflora marsh. Estuar. Coast. Shelf Sci., Vol. 16, pp. 559-571.
Rosset, .I., F. Batlochet and J.J. Oertli, 1982. Decomposition of conifer needles and leaf litter in two Black
Forest and two Swiss Juta streams. Inf. Rev. Ges. Hydrobiol., Vol. 67, pp. 695-711.
Schwinghamer, P., F.C. Tan and D.C. Gordon, Jr., 1983. Stable carbon isotope studies on the Peck’s Cove
mudflat ecosystem in the Cumbetland Basin, Bay of Fundy. Can. J. Fish. Aquat. Sci.,Vol. 40, pp. 262-272.
Snedecot, G.W. and W.G. Cochtan, 1967. Stufistical methods. Iowa State University, Ames, IA, 593 pp.
Somogyi, M., 1952. Notes on sugar determination. J. Biol. Chem.,Vol. 19.5, pp. 19-23.
Stuart,V., E.J.H. Head and K.H. Mann, 1985. Seasonal changes in the digestive enzyme levels of the amphipod
Corophium volutator (Pallas) in relation to diet. .I. Exp. Mar. Biol. Ecol., Vol. 88, pp. 243-256.
Teal, J.M., 1962. Energy flow in the salt marsh ecosystem of Georgia. In, Readings in marine ecology, edited
by J.W. Nybakken, Harper and Row, New York, NY, pp. 345-355
Thomas, M.L.H., 1983. Marine and coasral systems of the Quoddy region, New Brunswick. Canadian Special
Publication of Fisheries and Aquatic Sciences 64. Department of Fisheries and Oceans, Ottawa, Canada.
Valiela, I., J.M. Teal, S.D. Allen, R. Van Etten, D. Goehringet and S. Volkman, 1985. Decomposition in salt
marsh ecosystems: the phases and major factors affecting disappearance of above-ground organic matter. J.
Exp. Mar. Biol. Ecol., Vol. 89, pp. 29-54.
Wagner, D.T., 1965. Developmental biology of Leptosphaeria discors (Saccahtdo and Ellis) Saccardo and
Ellis. Nova Hedwigia, Vol. 9, pp. 45-61.
Wagner, D.T., 1969. Ecological studies of Leptosphaeria discors, a graminicolous fungus of salt marshes.
Nova Hedwigia,Vol. 23, pp. 915-922.
Wiedet, R.K. and G.E. Lang, 1982. A critique of the analytical methods used in examining decomposition data
obtained from litter bags. Ecology, Vol. 63, pp. 163661642.
Wilkinson, L., 1989. SYSTAT: the system for statistics. SYSTATS Inc., Evanston, IL, 724 pp.